
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceAcidogenic fermentation of dairy by-products for the utilization of volatile fatty acids IN PHBV production by Haloferax mediterranei†
Leire
Urbina
 *a,
Eric
Rovira-Cal
bc,
Ibai
Nafarrate
a,
Ana
Urkiaga
a,
Josu
Berganza
a,
Enrique
Aymerich
bc and
Maria José
Suárez
*a
*a,
Eric
Rovira-Cal
bc,
Ibai
Nafarrate
a,
Ana
Urkiaga
a,
Josu
Berganza
a,
Enrique
Aymerich
bc and
Maria José
Suárez
*a
aGaiker Technology Centre, Basque Research and Technology Alliance (BRTA), Parque Tecnológico de Bizkaia, Edificio 202, 48170 Zamudio, Spain. E-mail: urbina@gaiker.es; nafarrate@gaiker.es; urkiaga@gaiker; berganza@gaiker.es; suarez@gaiker.es
bCEIT-Basque Research and Technology Alliance (BRTA), Paseo Manuel Lardizabal 15, 20018 Donostia – San Sebastián, Spain. E-mail: eroviracal@ceit.es; eaymerich@ceit.es
cUniversidad de Navarra, Tecnun, Paseo Manuel Lardizabal 13, 20018 Donostia - San Sebastián, Spain
First published on 5th December 2024
Abstract
In this study, the acidogenic fermentation (AF) of cheese whey (CW) has been conducted to obtain streams with different volatile fatty acid (VFA) profiles for polyhydroxyalkanoate (PHA) biosynthesis. AF was carried out in single-phase reactors for parameter optimization under mesophilic and thermophilic conditions, and streams with 2718.2 mg L−1 of VFA (predominantly acetic acid, 1288 mg L−1, and propionic acid, 1119 mg L−1) and 2270.2 mg L−1 of VFA (predominantly butyric acid, 1339 mg L−1), were obtained, respectively. The thermophilic conditions were scaled up in a two-phase leachate bed reactor to obtain a final stream with a VFA concentration of 7879 mg L−1 composed mainly of butyric acid. Then, the technical feasibility of using the obtained streams as substrates for the production of poly(3-hydroxybutyrate-co-valerate) (PHBV) by Haloferax mediterranei was demonstrated. The PHBV production was found to be related to the butyric acid content, as the PHBV production was 1.4 vs. 0.3 g L−1 when using the streams rich in butyric acid vs. propionic acid, but the monomeric composition was dependent on the presence of propionic acid. A maximum PHVB production of 2.43 g L−1 and PHA accumulation of 44.3% were reached after 120 h of cultivation in bioreactor conditions. The characteristics of the produced PHBV bioplastic were compared with those of a commercial-grade PHA and found to be similar.
Sustainability spotlightBioprocesses, which involve the use of biological systems to create valuable products, are at the forefront of sustainable industrial practices. A wide range of industries are adopting bioprocesses to create products in a more environmentally friendly way. This is case for the materials industry and the production of bioplastics. Among the various methods to produce bioplastics, microbial fermentation stands out for its sustainability, efficiency, and lower environmental impact. The interest of this work lies in the combination of two bioprocesses to obtain a value-added bioplastic starting from dairy by-products. Acidogenic fermentation of cheese whey is performed to obtain volatile-fatty-acid-rich streams, followed by fermentation with Haloferax mediterranei, an extremophile bacteria that produces bioplastic under non-sterile conditions. |
1. Introduction
At present, biotechnology and bioprocesses for the cleaner production of numerous products for many industrial sectors, such as the health, food, energy, agriculture, and other sectors, are the subject of intensive research and development. The development and improvement of these processes are crucial to provide technological alternatives for a wide range of materials that are currently produced by chemical processes or using petroleum-derived products.1 In recent years, there has been an increasing trend towards more efficient utilization of agro-industrial by-products, residues and wastewaters for the production of various fermentation-related products. This is the case for cheese whey (CW), a by-product derived from the dairy industry.2 Whey is obtained by the precipitation and removal of milk casein during the cheese-making process, resulting in the production of approximately 10 L of CW per kilogram of cheese.3 Its composition can vary depending on factors such as the origin (cheese variety and type of milk) and production and process conditions used during cheese production. In general terms, it is mainly composed of lactose (39–60 g L−1), fats (0.99–10.58 g L−1), proteins (27–60 g L−1), and mineral salts (4.6–8 g L−1),4,5 making this by-product interesting for valorization in food-related applications. Strategies for cheese whey valorization also include its use as a low-cost substrate in fermentations for microbial growth (yeasts and bacteria) and the synthesis of value-added products such as biosurfactants,6 food product additives,3 lactic acid, succinate, riboflavin (vitamin B2)7 and microbial plastics.4,8,9 This last application in particular has been gaining particular attention in recent years due to environmental concerns about the use of petroleum-derived durable plastics and the need to search for sustainable materials that can compete with commonly produced plastics.4The microbial plastics known as polyhydroxyalkanoates (PHAs) are biologically produced polyesters that present similar characteristics to common plastics but are also biocompatible and biodegradable, making them promising for a wide range of applications. They are one of the few plastics that are biodegradable in soil and seawater; thus, they offer benefits in applications including marine and agricultural ropes, gears and films or food packaging.10,11 At present, the homopolymer poly(3-hydroxybutyrate) (PHB) and its copolymer with valerate, poly((3-hydroxybutyrate)-co-(3-hydroxyvalerate)) (PHBV), are practically the only commercially available PHAs.12 The latter presents improved mechanical properties and lower melting and glass transition points, which in turn offer a wider processing window.13 Despite the potential of these bioplastics in several applications, expanding the commercial dissemination of bacterium-derived polymer technology presents some challenges, including the selection of industrially robust organisms, the production of tailored copolymers and the use of suitable carbon sources that do not compete with food production as affordable substrates.14 In this sense, Haloferax mediterranei (H. mediterranei) is an extremophile archaeon that represents a model organism for PHBV production. Minimal sterility is required for its growth due to the high salt concentration (from 100 to 350 g L−1 total salts). These conditions hinder the growth of non-halophilic microorganisms. Furthermore, it can accumulate up to 70% polymer and is versatile in terms of the utilisation of renewable substrates for growth, i.e., rice and corn starch, ethanol stillage, molasses wastewater, food waste and cheese whey.8,15–17
It is difficult to find microorganisms that efficiently grow and produce PHAs using whey because it is mainly composed of lactose, which is not a preferred sugar for these kinds of bacterial strains.4 For this reason, whey is usually pre-treated to remove proteins and other solids from the lactose and then hydrolyzed into glucose and galactose, requiring multi-step processes. For example, in 2020 Raho et al.8 produced PHBV using H. mediterranei by first implementing a multi-step fractionation to recover lactose from cheese whey and then an enzymatic treatment to release glucose and galactose, resulting in successful growth and PHA production. Recently, acidogenic fermentation (AF), which is the initial phase of anaerobic digestion (AD), has proved to be an efficient process for the valorization of organic matter such as food waste.18,19 AD consists of four steps, i.e., hydrolysis, acidogenesis, acetogenesis, and methanogenesis, and the products obtained (hydrogen, ethanol, volatile fatty acids (VFAs) and biomethane) depend on the step at which the process is interrupted. AF can be carried out using pure-culture fermentations in which a specific acid can be produced, or with mixed cultures derived from AD, which produce a heterogenic spectrum of VFAs. Mixed-culture fermentation has advantages over pure-culture fermentation, as a wider range of feedstocks can be used, including agricultural waste, food waste and wastewater sludges.20 When methanogenesis is inhibited, VFAs or short-chain fatty acids (this term normally refers to C2–C6 fatty acids) such as acetic, propionic, butyric, and lactic acid, which are considered to be key platform chemicals in multiple industries, can be produced.21 These VFAs can be used as feedstocks for polyhydroxyalkanoate (PHA) production.22 Moreover, it has been reported that PHBVs with tailored compositions and microstructures (mol% of HV, hydroxyvalerate) can be produced using H. mediterranei depending on the VFAs present in the feed.23,24
In this work, cheese whey has been used to obtain a VFA feedstock through AF for subsequent PHA production by H. mediterranei. For the first time, the combination of two bioprocesses to valorize this residue, i.e., AF followed by bacterial fermentation, has proven a successful alternative to other multi-step valorization routes. Batch experiments were carried out to study the organic matter that could be converted into VFAs under mesophilic and thermophilic conditions. Then, a continuous chemostat experiment was conducted to study the influence of the hydraulic retention time (HRT) and pH on the VFA yield and VFA composition. Three streams with different VFA compositions were obtained and used in fermentation experiments with H. mediterranei. The PHA production and composition were analyzed, and the most promising feedstock was utilized in a 3 L reactor to extract the bioplastic. The physico-chemical, thermal and optical properties of the bioplastic were analyzed and compared to those of a commercial-grade PHBV to assess the suitability of the obtained PHA for different applications.
2. Materials and methods
2.1. Chemicals
Cheese whey (CW) powder was purchased from Norken S.L. and dissolved in distilled water at 84 g L−1 to match the composition of industrial CW. Its composition is detailed in Table 1. No pre-treatments were applied to the CW, and its preparation was carried out before the assays were started to avoid degradation.| Cheese whey | Inoculum | |
|---|---|---|
| TS (%) | 7.46 | 2.13 |
| VS (%) | 6.81 | 1.31 |
| tCOD (mg L−1) | 80![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 634 634 |
18481 |
| sCOD (mg L−1) | 78835 |
1162 |
| TKN (mg L−1) | 1344 | — |
| Protein (mg L−1) | 8575 | — |
| N–NH4 (mg L−1) | 129 | 1596 |
| pH | 6.03 | 7.67 |
The chemicals for the bacterial growth medium were as follows: NaCl, MgCl2·6H2O, CaCl2·6H2O and NaHCO3, (Sigma Aldrich, Darmstadt, Germany); MgSO4·7H2O, NaBr and KH2PO4 (PanReac AppliChem, Castelar de Vallés, Barcelona, Spain); KCl (Scharlab, Sentmenat, Barcelona, Spain); FeCl3 (Thermo Scientific, Alcobendas, Madrid, Spain); mineral trace solution NaMoO4·H2O and NiCl2·6H2O (Sigma Aldrich); MnCl2·4H2O, CuSO4, CoCl2·6H2O and ZnSO4·7H2O (PanReac AppliChem) and H3BO3 (VWR chemicals, Geldenaaksebaan, Leuven, Belgium). Sodium hydroxide (NaOH) and Ringers solution were obtained from Thermo Scientific. Butyric and valeric acids (≥99%) for the GC-MS calibration curve were purchased from Sigma-Aldrich.
Commercial-grade PHBV with a density of 1.25 g cm−3 and a melting temperature range of 175–180 °C was purchased from TiaNan (ENMAT 1000P) for comparison purposes.
2.2. Acidogenic fermentation of cheese whey
The pilot plant was equipped with a PT100 temperature probe and an electrical heater to automatically control the temperature of the reactor at a setpoint value. A pH probe (InPro3100iUD/120, Mettler Toledo) was installed and displayed using a Mettler Toledo M300. Initially, the pH was set at 5.5 to avoid methanogenic activity, and it was controlled manually every day using 8 N NaOH. The complete mixing was controlled with a frequency variator (Yaskawa VS-606V7) at 25 rpm. The reactor had a sealed gas outlet to allow the produced gas to escape while ensuring anaerobic conditions in the reactor. VFA, sugars, total chemical oxygen demand (tCOD) and pH were measured daily.
For semi-continuous operation, the pilot reactor was manually operated at a hydraulic retention time (HRT) of 4 days and pH 5.5 to maximise VFA concentrations. An ultrafiltration pilot plant was used for VFA filtration. 300 kDa ceramic membranes (INSIDE CéRAM™ 300 kDa – TAMI INDUSTRIES) were used to remove the solids from the fermentation broth after the batch. The membrane surface was 0.02 m2. A recirculation centrifuge pump (Grunfos CM3-6A) provided a recirculation flow of 300 L h−1, which produced an operation transmembrane pressure of 0.9 bar. To maintain room temperature, a serpentine refrigerator (Selecta Frigedor) was installed in the recirculation tank.
Analyses of the total fraction were performed directly over raw samples. The soluble fraction was obtained by centrifugation at 12000 rpm for 5 min, and the supernatant was filtered (Millipore 0.7 μm). Total solids (TS), VS, tCOD, sCOD, ammonia and pH were measured according to Standard Methods, 21st Edition.25 VFA was measured for the soluble fraction.26
2.3. Inocula and media preparation for PHA production
In this work the H. mediterranei DSM 1411 bacterial strain was acquired from The Leibniz Institute DSMZ German collection of Microorganisms and Cell Cultures GmbH, Germany. Lyophilised cells were recovered following the supplier's instructions. Master and working stocks were created from the recovered lyophilised cells using CRIOTECA® vials from Microkit Laboratories (Valdemorillo, Madrid, Spain) for cryogenic preservation. Vials were stored at −80 °C prior to reactivation and use in experiments.For the inoculum preparation, cells from cryo-storage were cultured in HS agar plates at 37 °C for 72 h. A single colony of H. mediterranei from a Petri dish was used to inoculate 100 mL of HS broth,27 which was incubated at 37 °C for 72 h with 200 rpm shaking. After incubation, the suspension was centrifuged at 10000g, and the obtained pellet was used as the inoculum to analyse the ability of H. mediterranei to produce PHBV from VFA obtained via whey acidogenic fermentation (WAF) at concentrations of 100% and 50% (diluted with distilled water). All culture media prepared via WAF were supplemented with 156 g per L NaCl, 13 g per L MgCl2·6H2O, 20 g per L MgSO4·7H2O, 1 g per L CaCl2·6H2O, 4 g per L KCl, 0.2 g per L NaHCO3, 0.5 g per L NaBr, 0.5 g per L KH2PO4, 0.005 g per L FeCl3, and 10 mL per L SL-6 micronutrients solution17 which contained 0.03 g per L NaMoO4·H2O, 0.02 g per L NiCl2·6H2O, 0.025 g per L MnCl2·4H2O, 0.006 g per L CuSO4, 0.2 g per L CoCl2·6H2O, 0.1 g per L ZnSO4·7H2O and 0.3 g per L H3BO3. The pH was adjusted to 7.2 using NaOH. Experiments were carried out under non-sterile conditions.
To maximize cell growth, an initial adaptation stage was included. The adaptation stage was included after the HS broth growth and prior to PHBV production by inoculating the pellet from the HS broth culture in the corresponding WAF medium and incubating the flasks at 37 °C for 72 h with 200 rpm shaking.28 This medium was used to inoculate the growth media. A reinoculation strategy was carried out, which consisted of the centrifugation of 80 mL of exhausted WAF medium (37 °C for 72 h and 200 rpm) and inoculation of the pellet into a fresh identical medium at 37 °C and 200 rpm for 144 h. All experiments were carried out in duplicate or triplicate in 250 mL Erlenmeyer flasks with 100 mL working volumes under non-sterile conditions; the data reported are the mean ± standard deviation.
2.4. PHA production in fed-batch fermentation
For the bioreactor fermentation, 400 mL of HS broth in a 1 L flask was inoculated with a single colony from the HS agar plate. After incubation at 37 °C for 144 h with agitation at 200 rpm, the cells were harvested by centrifugation at 10000g, and the pellet was inoculated in a 3 L jacketed bioreactor (Applikon® biotechnologies) containing 1 L of supplemented WAF medium. To enhance PHBV production, an initial 96 h adaptation phase was conducted at 37 °C. During this phase, the pH of the bioreactor was maintained at 7.0 by adding either 1 M HCl or 1 M NaOH; dissolved oxygen was maintained at 80% by adjusting the stirrer speed to 650 rpm and providing an extra air supply of 0.5–1 L min−1 as needed. When the adaptation stage was finished, the cells were collected by centrifugation and used to inoculate 800 mL of the same fresh supplemented WAF medium in the same bioreactor. The fermentation was carried out under the same conditions described before and was monitored over 144 h. Samples were taken at 96, 120 and 144 h to analyse cell growth and PHBV production.
2.5. Analytical methods
Bacterial evolution over time was monitored via turbidimetry by measuring the optical density (OD625) at 625 nm using a Shimadzu UV-2550 (Shimadzu, USA) spectrophotometer.The total biomass was estimated gravimetrically as dry cell weight (DCW). 10 mL of fermented broth was obtained from the shake flasks or bioreactor and centrifuged at 10000g for 10 min. The cell pellet was resuspended in 500 μL of Ringers solution and dried at 50 °C using labelled, dried and pre-weighted supports to estimate the final DCW.
PHA quantification and composition determination were performed using methanolysis and gas chromatography (GC-MS). Methanolysis was carried out as described by Urbina et al.29 10 mg of biomass was dissolved in 2 mL of CH2Cl2, and then 2 mL of a CH3OH:H2SO4 (85:15 v/v) mixture was added to the vial. The mixture was incubated at 100 °C for 140 min and then cooled to room temperature. About 1 mL of distilled water was added, and vigorous vortexing was carried out. The resulting solution was allowed to stand for 10 min. The bottom organic layer was separated and filtered into GC vials, and the remaining H2O content was removed. A gas chromatography system (model 6890N, Agilent Technologies) with an HP – 5MS column (30 m × 0.25 mm × 0.25 μm) and helium as the carrier gas was used. The injection volume was 1 μL in splitless mode. The temperature was initially 40 °C and was increased gradually to 200 °C. A calibration curve was constructed using methyl (R)-3-hydroxybutyrate and methyl (R)-3-hydroxyvalerate (from the methanolysis of butyric and valeric acids, as explained above) with benzoic acid as an internal standard.
The PHA proportion of the total biomass was evaluated at the end of the fermentation process as a concentration (mass of PHA per litre of fermentation broth) and as a percentage of total biomass (PHA (%)) (eqn (1)). The percentage of hydroxyvalerate (HV) units in the PHBV polymer was determined based on the mass ratio of the HV units to the sum of HV and HB (hydroxybutyrate) units as measured via GC-MS (eqn (2)).
 | (1) |
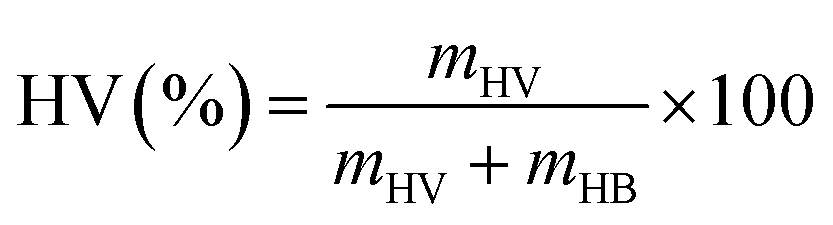 | (2) |
2.6. Polymer extraction and characterization
000g and 4 °C for 10 min, washed with ultrapure water, and pre-treated with acetone in order to remove lipids30 until a white pellet was obtained. For the PHA purification, the previously obtained pellet was washed in a Soxhlet apparatus using chloroform (1:100 g g−1) and subsequently precipitated with 10 volumes (1:10 v/v) of cold methanol (4 °C) under continuous stirring. The PHA was recovered by decantation or centrifugation at 10000g for 4 min and dried at 30 °C under vacuum.
:H2SO4 (85:15 v/v) mixture) and treated as above. About 2 μL of the methanolyzed sample was automatically injected into the column. The oven temperature was programmed to hold at 40 °C for 2 min and then increase to 240 °C at 5 °C min−1 and hold at this temperature for 15 min. The opacity of PHBV cast films was determined according to the method reported by Zhao, Wang & Liu in 2022.31 The film absorbance was measured at 600 nm using a UV-vis spectrophotometer (UV-1800 Shimadzu). The films were directly placed in a spectrophotometer test cell, and an empty test cell was used as the reference. The opacity of the films was calculated using eqn (3): | (3) |
3. Results and discussion
3.1. Acidogenic fermentation of cheese whey
In the present work, mixed-culture fermentation was carried out in order to examine different VFA profiles. In AF, the temperature is an operational parameter that impacts microbial kinetics, and digesters are usually operated in mesophilic (25–45 °C) or thermophilic (50–60 °C) temperature ranges. It has been reported in the literature that thermophilic conditions lead to higher hydrolytic and acidogenic activity, resulting in higher VFA production rates while reducing fermentation time.32,33To compare VFA yields and identify the optimum operating parameters in the batch mode, two pilot-scale batch fermentations were carried out in the mesophilic and thermophilic temperature ranges in single-phase reactors. An inoculum-to-substrate ratio of 0.8 gram VSIn per gram VSCW was used to compare the two conditions. This ratio was chosen based on previous studies26 and ensured complete substrate degradation, as it was added in low concentration. This also prevents possible substrate inhibition and facilitates an initial comparison between different operating conditions. A pH of 5.5 was selected based on bibliographic research, which indicated that an acidic pH of 5.5 is optimum for maximising the yield of VFA production from CW.34,35 The evolution of the VFA profile over time during the mesophilic and thermophilic fermentations is shown in Fig. 1.
 | ||
| Fig. 1 Daily evolution of the VFA fraction during mesophilic (a) and thermophilic (b) cheese whey fermentations. | ||
In the mesophilic fermentation, a heterogeneous profile was found in which propionic and acetic acids clearly predominated. Longer-chain fatty acids, i.e., valeric and caproic acids (C5–C6), appeared only after day 4 of fermentation. As can be observed, the fraction of acetic and propionic (C2–C3) acids decreased slightly during the last days of the fermentation, in agreement with the production of C5–C6 acids. This is due to the chain elongation process, in which longer volatile fatty acids are produced from acetic and propionic acids.36,37 This is corroborated by the fact that the sugars were consumed within the first 3 days and no other substrate that could explain the production of these acids was available.
Under thermophilic conditions, a shift in the product spectrum was observed. Chain elongation was not observed, and butyric acid was the main constituent at the end of the fermentation. In the final days of the assay, a slight increase in the fraction of acetic acid at the expense of that of butyric acid occurred, probably due to acetogenic bacteria present in the inoculum.38 As in the mesophilic fermentation, the sugars were completely consumed during the first 3 days. The production yield and concentration of both fermentations are listed in Table 2. As can be observed from the results, thermophilic conditions led to a slightly higher VFA yield. In 2019, Arras et al. also found a similar VFA distribution under the same conditions and observed that higher yields could be achieved at thermophilic temperatures.33 Another important variable is the substrate composition. As mentioned previously, CW is a carbohydrate-rich RS. Vázquez-Fernández et al. considered the effect of substrate composition in 2022, and the results shown in this work are representative of the fermentation of a carbohydrate-rich substrate.39
| Operation mode | Temperature | VFA yield (% gVFA ginitial sugars−1) | [VFA] (mg L−1) | Designation |
|---|---|---|---|---|
| Batch | Mesophilic | 47.2 | 2718.2 | WAF1 |
| Batch | Thermophilic | 49 | 2270.2 | WAF2 |
| Semi-continuous | Thermophilic | 16.2 | 7879 | WAF3 |
To increase the VFA concentration in the fermentation bulk, a semi-continuous operation was carried out under thermophilic conditions. Several studies involving batch mode operation at lab-scale have been reported, but the final objective is to scale-up these systems to full-scale continuous fermentations.40 The semi-continuous operation was carried out with higher substrate loads (i.e., 20 g VS per L per days) than the batch fermentations. The reactor was operated at pH 5.5 for an HRT of 4 days with the aim of recovering a VFA-rich effluent (according to a deeper analysis of pH and HRT carried out in our research group). Once a stationary phase was reached, the daily discharge was collected, and an ultrafiltration step was performed. When enough effluent for the PHA fermentation studies had been produced, the collection was stopped.
The yield of the semi-continuous fermentation (Table 2) was much lower than that achieved in the batch assays, but the final VFA concentration was almost three times greater. This was due to the initial sugar concentration, as the batch assays were carried out under controlled fermentation conditions for comparison purposes, while in the semi-continuous fermentation, the objective was to reach higher concentrations. Other studies in the literature have reported higher production yields,41 but most of them involve the use of a diluted substrate, which results in lower product concentrations. In the present work, the substrate was not diluted for two main reasons: first, high VFA concentrations are more favourable for the subsequent processes. Second, diluting the substrate would not be viable in a full-scale application due to water requirements, making the process less environmentally friendly.
After the AF of the cheese whey under the above-mentioned conditions, three different streams were obtained. The composition of each is listed in Table 3.
| Designation of stream | WAF1 | WAF2 | WAF3 |
|---|---|---|---|
| AF conditions | Mesophilic | Thermophilic | Thermophilic |
| pH | 5.5 | 5.5 | 5.5 |
| Operation mode | Batch | Batch | Semi-continuous |
|
|||
| Quantity (mg L− 1 ) | |||
| Acetic (Hac) | 1288 | 499 | 1869 |
| Propionic (Hpro) | 1119 | 318 | <1 |
| Isobutyric (IsoHbu) | 7 | 23 | <1 |
| Butyric (Hbu) | 177 | 1339 | 6010 |
| Isovaleric (IsoHva) | 38 | 91 | <1 |
| Valeric (Hva) | 104 | <1 | <1 |
| Caproic (Hcap) | 13 | <1 | <1 |
| Total VFA | 2746 | 2270 | 7879 |
As can be observed, the WAF1 stream was composed mainly of acetic acid (46.9%) and propionic acid (40.8%) along with small amounts of butyric and valeric acids, which led to the lowest production of PHBV. WAF1 and WAF2 had similar total VFA contents, but WAF2 was composed of 59% butyric acid, 21.9% acetic acid and 15% propionic acid. In contrast, WAF3 was richer in butyric acid.
3.2. PHA production from WAF streams
Although the use of cheese whey for the production of PHBV by this bacterial strain has been previously tested, the previous strategies focused on hydrolysis of the lactose.8,27 In those studies, the lactose was hydrolyzed to glucose and galactose, which were used as the carbon sources for the bacteria. To find alternatives, this work focuses on the utilization of the VFAs in AF rather than sugars. The ability to control the VFA composition can be an advantage, as it has been shown that the HV% is proportional to the C5% in the feed.24To analyse the ability of H. mediterranei to produce PHBV from these substrates and to check for inhibitory effects of VFA concentration, the three streams were used in shake flask experiments, either at the initial concentration or diluted to 50% of the initial concentration.42 In all cases, the streams were supplemented with salts and mineral trace solution. Fermentations were carried out at 37 °C for 144 h. The biomass over time (cell growth plus PHA production) was measured using turbidimetry, and the dry cell weight (DCW) and the amount of polymer synthesized were both determined at the end of incubation. Fig. 2 shows that H. mediterranei was able to grow and produce PHAs in all the tested streams. Table 3 lists the PHA accumulation (%) and DCW for all experiments. No inhibitory effects were observed, as greater bacterial growth and PHA production were observed when the higher concentration of VFAs (100%) was used in all cases. Among the three WAF streams tested, WAF1 was the least effective, while WAF2 and WAF3 gave similar results in terms of PHBV production (0.64 and 0.61 g L−1, respectively), DCW (4.9 and 5.4 g L−1, respectively) and PHA accumulation (13 and 10% respectively), even though the total VFA content of WAF3 was 3.4 times higher.
 | ||
| Fig. 2 PHA production and OD for the WAF streams at different concentrations. | ||
To maximize DCW and PHA production, a re-inoculation step was performed (denoted by R in Fig. 2). This is a similar strategy to that followed in other works with other bacterial strains to maximize PHA production: the addition of extra feed at different cultivation times, i.e., a fed-batch fermentation strategy, leads to remarkably higher polymer yields.29,43 When the reinoculation step was conducted, WAF3 gave significantly improved results, reaching 1.4 g L−1 of PHBV and 17.5 g L−1 of DCW. In H. mediterranei, the polymer accumulation is associated with cell growth, so no nutrient limitation is needed for PHA production. The results for the intracellular accumulation of polymer (PHA%) are listed in Table 4. For the WAF1 stream, which was mainly composed of acetic and propionic acids, the accumulation was almost negligible, with a maximum accumulation of 5%. For WAF2 and WAF3, which were rich in butyric acid, higher accumulation values were obtained. These results suggest that the production yield is related to the butyric acid content. The data were subjected to multifactorial analysis of variance (ANOVA) in which the factors were the WAF stream, reinoculation and % concentration used; the variable was the accumulation of PHA. The WAF stream was found to be significant (p < 0.05).
| PHA%|DCW (g L−1) | ||||||
|---|---|---|---|---|---|---|
| WAF1 | WAF2 | WAF3 | ||||
| 100% | 50% | 100% | 50% | 100% | 50% | |
| 1 | 5.3|3.8 ± 0.19 | 3.8|2.8 ± 0.05 | 13|4.9 ± 0.07 | 11|2.8 ± 0.04 | 10|5.4 ± 0.1 | 14|4.3 ± 0.02 |
| R | 5|5.9 ± 0.08 | 4|3.6 ± 0.05 | 11|6.3 ± 0.09 | 11.3|3.6 ± 0.03 | 11.1|17.5 ± 0.17 | 14.4|7.7 ± 0.2 |
As it has been previously reported in other works, the PHV precursors propionate (C3:0) and valerate (C5:0) are not the most suitable acids for cell growth of H. mediterranei, and acetic acid is consumed to a very low extent. In 2018, Ferre-Guell & Winterburn24 obtained an accumulation of 5.7% using 0.1 M of propanoic acid as the only carbon source. They performed a screening experiment with saturated fatty acids with chain lengths varying from C2:0 to C11:0, and cell growth was only observed for C2:0 to C5:0 at all carbon concentrations and C6:0 at low concentration. The experiments revealed that higher-purity PHB and PHV were obtained when H. mediterranei cultures were grown in C4:0 and C5:0 fatty acids, respectively, and C2:0 and C3:0 fatty acids led to lower PHBV (g L−1) concentrations. In addition, Wang & Zhang in 202117 studied the effect of a VFA mixture containing acetic acid, propionic acid and butyric acid on the microbial growth and PHBV production of this bacterial strain. By the end of cultivation, the butyric acid was completely consumed, around 81% of the propionic acid was consumed, and only 41% of acetic acid was consumed, although all species started with the same mol C% in the mixture. This indicates that acetic and propionic acids are not preferred acids for H. mediterranei, which agrees with the results of the present work.
In terms of polymer composition, the HV% value follows the trend WAF1 > WAF2 > WAF3, as shown in Fig. 3, regardless of the WAF concentration tested. As has been reported in literature, the main precursors of HV units for H. mediterranei are glucose and propionic and valeric acids.17 WAF3 contains only traces of the latter (<1 mg L−1), which led to only 2–1.4% HV units. In contrast, using WAF1 (1120 mg L−1 of propionic acid), a polymer containing 15–18% HV units was obtained. This is interesting, as the % HV affects the properties of the final bioplastic. Thus, by changing the HV precursor concentration, the polymer composition could be tuned for specific applications. The commercial-grade PHBV available at present usually contains 2–4% HV units. This fact provides an opportunity to compare the PHBV produced in this work to commercial-grade PHBV.
 | ||
| Fig. 3 PHA production and obtained composition after reinoculation for different WAF streams. | ||
3.3. PHA production in fed-batch fermentation
After the batch cultures in shake flask experiments were carried out, the WAF3 medium was scaled up in a bioreactor system under continuous control of the pH, temperature and O2 supply. WAF3 was selected because this medium led to the highest PHBV production due to its high butyric acid content.In the bioreactor experiment, samples were taken at 90, 120 and 144 h and the OD and PHA production were determined. As can be observed in Fig. 4, a maximum PHA production of 2.43 g L−1 and PHA accumulation of 44.3% were reached at 120 h of cultivation. However, the OD of the bacterial culture continued to grow at 144 h. The simultaneous increase of the OD and decrease of the PHBV concentration at 144 h could indicate that the accumulated PHA was being consumed, suggesting that the carbon source was exhausted at 120 h mark.
 | ||
| Fig. 4 Evolution of H. mediterranei in WAF3 medium. (●OD (600 nm), ▶PHA%, □ PHA production). | ||
These values are significantly higher than those obtained in the flask trials, indicating that the bioreactor conditions had been successfully scaled up. This is a common trend observed in other studies and is attributed to better control of the cultivation parameters, such as pH and dissolved oxygen, in the bioreactor.7,30 In 2020, Raho et al.8 obtained a PHA recovery of 1.18 g L−1 after growing this bacterial strain in a bioreactor system and observed remarkably higher values of polymer synthesis and DCW compared to those obtained in flask experiments. The results obtained in the present study suggest that the application of the acidogenic fermentation of an agroindustrial by-product to obtain a marketable bioplastic is promising, although further investigation could be performed for higher PHBV production and optimization.
3.4. PHBV characterization
The biosynthesised PHBV from WAF3, designated as PHBV (CW), was extracted from the biomass and characterized using different techniques to evaluate its properties. The films were optically translucent with a whitish colour, as shown in Fig. 5a. The thermal and optical properties were compared those of a commercial-grade PHA (1000P). | ||
| Fig. 5 (a) Casted PHBV (CW) film, (b) FTIR spectrum of PHBV (CW), (c) DSC thermograms and (d) TGA curves. | ||
The FTIR spectrum (Fig. 5b) exhibits the typical bands of the polyester structure.44 The bands at 2977, 2971 and 2934 cm−1 are associated with aliphatic C–H. The strong band at 1740–1720 cm−1 is a characteristic peak of PHAs and corresponds to the stretching vibration of the carbonyl of the ester group (C![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) O). The intensity and position of the carbonyl group are related to changes in the crystallinity of the PHA. The band shifts from a broad shoulder centered at 1740 cm−1 (related to the free CO) to a stronger and sharper peak at 1720 cm−1 (related to the hydrogen-bonded CO), which are assigned to the amorphous and crystalline phases, respectively.45 The bands at 1453 and 1378 cm−1 are assigned to CH2 and CH3 bending vibrations, respectively. The stretching vibration of the C–O–C bonds appears in the 1330–1000 cm−1 region.
O). The intensity and position of the carbonyl group are related to changes in the crystallinity of the PHA. The band shifts from a broad shoulder centered at 1740 cm−1 (related to the free CO) to a stronger and sharper peak at 1720 cm−1 (related to the hydrogen-bonded CO), which are assigned to the amorphous and crystalline phases, respectively.45 The bands at 1453 and 1378 cm−1 are assigned to CH2 and CH3 bending vibrations, respectively. The stretching vibration of the C–O–C bonds appears in the 1330–1000 cm−1 region.
Moreover, the transition temperatures and thermal stability were studied and compared to those of a commercial-grade PHBV (Y1000P). In the DSC thermogram (Fig. 5c) two endothermic melting peaks (Tm) were observed in PHBV (CW) at 148 and 163 °C, which correlated with the DSC thermograms of PHBV reported in the literature.8,43 This bimodal endotherm is associated with the formation of heterogenous crystal morphologies during crystallization phenomena.46 In the case of the commercial PHBV (Y1000P), a unique melting peak at 177 °C was observed. The TG analysis in Fig. 5d revealed one-step thermal degradation between 250 and 330 °C with residual mass loss until 450 °C in the case of PHBV (CW). PHBV 1000P exhibited slightly higher thermal stability, with the mass loss starting at 266 and continuing until 310 °C.
Finally, the opacity of the films was measured, and the values were found to be 5.3 and 9.7 mm−1, for PHBV (CW) and PHBV (Y1000P), respectively. The opacity values of short-chain chain-length PHAs are usually higher than the ones those for medium medium-chain chain-length PHAs due to the crystalline nature of PHBV.47
4. Conclusions
In the present work, streams with variable VFA concentrations were obtained through the acidogenic fermentation of cheese whey to produce bioplastic with a halophile bacterium. Single-phase bioreactor experiments showed similar yields and conversion of volatile fatty acids under mesophilic (47.2% and 2718.2 mg L−1) and thermophilic conditions (49%, 2270.2 mg L−1). However, the acid profiles of the streams were found to be different; the mesophilic-fermentation-derived stream was rich in acetic acid (1288 mg L−1) and propionic acid (1119 mg L−1), while the thermophilic-derived-stream was predominantly rich in butyric acid (1339 mg L−1). The thermophilic conditions were chosen for use in a two-phase leachate bed reactor for continuous production, with butyric acid being the main constituent. These streams were tested for the production of PHBV by Haloferax mediterranei; the stream composition was confirmed to be directly related to the production and monomer composition of the biopolymer. No inhibitory effects were observed, and the results indicated that cell growth and polymer production are related to the butyric acid content, with 1.4 g L−1 of PHBV being obtained in flask experiments using the thermophilic-AF-derived streams in contrast to 0.3 g L−1 of PHBV using the mesophilic-derived-stream. A maximum PHVB production of 2.43 g L−1 and PHA accumulation of 44.3% were obtained in controlled bioreactor experiments. In addition, the extracted PHBV showed similar properties to a commercial-grade product, which currently has potential applications, mainly in the packaging sector. Although this study requires further optimization in terms of the fermentation conditions, it represents a first step toward advancing bioprocessing technologies to obtain value-added bioplastics. This work gives an insight into a biorefinery process in which an agri-food by-product has been valorised via acidogenic fermentation to obtain carbon substrates that are currently mainly produced via chemical synthesis using petroleum-based feedstocks.Data availability
The data supporting this article have been included as part of the ESI.†Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors are thankful for the financial support of the Basque Government (Proyectos de Investigación Fundamental Colaborativa) in the frame of CIRCULAR BIOBASED project and the CDTI under the Ministry of Science, Innovation and Universities for the financial support in the frame of MARFIL network.References
- A. Barragán-Ocaña, P. Silva-Borjas, S. Olmos-Peña and M. Polanco-Olguín, Processes, 2020, 8, 1–11 CrossRef.
- T. S. Anand, H. Vahab, D. Chandran, A. Shanavas, M. Kumar, F. Nainu, M. Bagath, P. Mohankumar, R. K. Mohapatra, S. Chakraborty, M. Alagawany and K. Dhama, Indian Vet. J., 2022, 99, 07–19 Search PubMed.
- G. Iosca, M. Turetta, L. De Vero, C. H. Bang-Berthelsen, M. Gullo and A. Pulvirenti, Lwt, 2023, 176, 114524 CrossRef CAS.
- T. M. M. M. Amaro, D. Rosa, G. Comi and L. Iacumin, Front. Microbiol., 2019, 10, 1–12 CrossRef PubMed.
- A. Figueroa Pires, N. Garcia Marnotes, O. Díaz Rubio, A. Cobos Garcia and C. Dias Pereira, Foods, 2021, 10, 1067 CrossRef PubMed.
- V. Kachrimanidou, D. Alimpoumpa, A. Papadaki, I. Lappa, K. Alexopoulos and N. Kopsahelis, Biomass Convers. Biorefin., 2022, 4621–4635 CrossRef CAS.
- J. Ruchala, Y. A. Andreieva, A. O. Tsyrulnyk, S. M. Sobchuk, A. Najdecka, L. Wen, Y. Kang, O. V. Dmytruk, K. V. Dmytruk, D. V. Fedorovych and A. A. Sibirny, Microb. Cell Fact., 2022, 21, 1–9 CrossRef PubMed.
- S. Raho, V. E. Carofiglio, M. Montemurro, V. Miceli, D. Centrone, P. Stufano, M. Schioppa, E. Pontonio and C. G. Rizzello, Foods, 2020, 9(10), 1459 CrossRef CAS PubMed.
- S. RamKumar Pandian, V. Deepak, K. Kalishwaralal, N. Rameshkumar, M. Jeyaraj and S. Gurunathan, Bioresour. Technol., 2010, 101, 705–711 CrossRef CAS PubMed.
- M. Winnacker, Eur. J. Lipid Sci. Technol., 2019, 121, 1–9 CrossRef.
- S. Kliem, M. Kreutzbruck and C. Bonten, Materials, 2020, 13, 4586 CrossRef CAS PubMed.
- M. Koller and A. Mukherjee, Bioengineering, 2022, 9, 74 CrossRef CAS PubMed.
- M. I. Ibrahim, D. Alsafadi, K. A. Alamry and M. A. Hussein, J. Polym. Environ., 2021, 29, 1010–1030 CrossRef CAS.
- V. C. Kalia, S. K. Singh Patel, R. Shanmugam and J. K. Lee, Bioresour. Technol., 2021, 326, 124737 CrossRef CAS PubMed.
- A. Bhattacharyya, J. Saha, S. Haldar, A. Bhowmic, U. K. Mukhopadhyay and J. Mukherjee, Extremophiles, 2014, 18, 463–470 CrossRef CAS PubMed.
- B. Colombo, T. P. Sciarria, M. Reis, B. Scaglia and F. Adani, Bioresour. Technol., 2016, 218, 692–699 CrossRef PubMed.
- K. Wang and R. Zhang, J. Microbiol. Biotechnol., 2021, 31, 338–347 CrossRef CAS PubMed.
- J. Sun, L. Zhang and K. C. Loh, Bioresour. Bioprocess., 2021, 8, 68 CrossRef PubMed.
- J. Yin, X. Yu, Y. Zhang, D. Shen, M. Wang, Y. Long and T. Chen, Bioresour. Technol., 2016, 216, 996–1003 CrossRef CAS PubMed.
- E. C. Guarda, A. C. Oliveira, S. Antunes, F. Freitas, P. M. L. Castro, A. F. Duque and M. A. M. Reis, Appl. Sci., 2021, 11, 3222 CrossRef CAS.
- M. Ramos-Suarez, Y. Zhang and V. Outram, Rev. Environ. Sci. Biotechnol., 2021, 20, 439–478 CrossRef CAS.
- H. Al, B. Shatha, A. Kindi, V. Kumar and G. Nallusamy, J. Polym. Environ., 2021, 29, 250–259 CrossRef.
- A. Ferre-guell and J. Winterburn, Extremophiles, 2017, 21, 1037–1047 CrossRef CAS PubMed.
- A. Ferre-guell and J. Winterburn, Biomacromolecules, 2018, 19(3), 996–1005 CrossRef CAS PubMed.
- E. E. W. Rice, R. B. Baird and A. D. Eaton, Standard Methods for the Examination of Water and Wastewater, APHA (American Public Health Association)/AWWA (American Water Works Association), 23rd edn, 2017, p. 1496 Search PubMed.
- J. Garcia-Aguirre, E. Aymerich, J. González-Mtnez de Goñi and M. Esteban-Gutiérrez, Bioresour. Technol., 2017, 244, 1081–1088 CrossRef CAS PubMed.
- J. Pais, L. S. Serafim, F. Freitas and M. A. M. Reis, New Biotechnol., 2016, 33, 224–230 CrossRef CAS PubMed.
- F. Guzman Lagunes and J. B. Winterburn, Bioresour. Technol., 2016, 221, 336–343 CrossRef CAS PubMed.
- L. Urbina, P. Wongsirichot, M. Á. Corcuera, N. Gabilondo, A. Eceiza, J. Winterburn and A. Retegi, Eur. Polym. J., 2018, 108, 1–9 CrossRef CAS.
- M. Abbasi, D. Pokhrel, E. R. Coats, N. M. Guho and A. G. McDonald, Polymers, 2022, 14, 4140 CrossRef CAS PubMed.
- J. Zhao, Y. Wang and C. Liu, Food Anal. Methods, 2022, 15, 2840–2846 CrossRef.
- B. Steiniger, S. Hupfauf, H. Insam and C. Schaum, Fermentation, 2023, 9, 798 CrossRef CAS.
- W. Arras, A. Hussain, R. Hausler and S. R. Guiot, Waste Manag., 2019, 87, 279–287 CrossRef CAS PubMed.
- E. Jankowska, J. Chwialkowska, M. Stodolny and P. Oleskowicz-Popiel, Chem. Eng. J., 2017, 326, 901–910 CrossRef CAS.
- S. Greses, E. Tomás-Pejó and C. González-Fernández, Biomass Convers. Biorefin., 2022, 12, 4519–4529 CrossRef CAS.
- C. A. Contreras-Dávila, V. J. Carrión, V. R. Vonk, C. N. J. Buisman and D. P. B. T. B. Strik, Water Res., 2020, 169, 115215 CrossRef PubMed.
- C. Zhang, Z. Ling, L. Yang, Y. Liu, T. Cao, Y. Sun, W. Liu, S. Huo, Z. H. Zhang, H. Su, J. Baeyens and X. Qi, J. Clean. Prod., 2023, 383, 135394 CrossRef CAS.
- C. Huang, W. Wang, X. Sun, J. Shen and L. Wang, J. Environ. Manage., 2020, 255, 109842 CrossRef CAS PubMed.
- A. Vázquez-Fernández, M. E. Suárez-Ojeda and J. Carrera, J. Environ. Chem. Eng., 2022, 10, 107917 CrossRef.
- L. Zhang, K. C. Loh, Y. Dai and Y. W. Tong, Waste Manag., 2020, 109, 75–84 CrossRef CAS PubMed.
- M. Atasoy and Z. Cetecioglu, J. Environ. Manage., 2022, 319, 115700 CrossRef CAS PubMed.
- G. Agustín Martinez, L. Bertin, A. Scoma, S. Rebecchi, G. Braunegg and F. Fava, Biochem. Eng. J., 2015, 97, 92–100 CrossRef.
- A. Ferre-Guell and J. Winterburn, Biochem. Eng. J., 2019, 141, 35–42 CrossRef CAS.
- K. Chotchindakun, W. Pathom-aree, K. Dumri, J. Ruangsuriya, C. Pumas and J. Pekkoh, Plants, 2021, 10, 1–24 CrossRef PubMed.
- M. Kansiz, A. Domínguez-Vidal, D. McNaughton and B. Lendl, Anal. Bioanal. Chem., 2007, 388, 1207–1213 CrossRef CAS PubMed.
- S. Singh, A. K. Mohanty, T. Sugie, Y. Takai and H. Hamada, Composites, Part A, 2008, 39, 875–886 CrossRef.
- L. Urbina, A. Eceiza, N. Gabilondo, M. Á. Corcuera and A. Retegi, Food Packag. Shelf Life, 2019, 21, 100356 CrossRef.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4fb00279b |
| This journal is © The Royal Society of Chemistry 2025 |