
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Raman spectroscopy as a comprehensive tool for profiling endospore-forming bacteria†
Markus
Salbreiter
 ab,
Annette
Wagenhaus
ab,
Petra
Rösch
*ab and
Jürgen
Popp
abcd
ab,
Annette
Wagenhaus
ab,
Petra
Rösch
*ab and
Jürgen
Popp
abcd
aInstitute of Physical Chemistry and Abbe Center of Photonics, Friedrich Schiller University, Helmholtzweg 4, Jena, Germany. E-mail: Petra.roesch@uni-jena.de; Tel: +49 3641 948381
bInfectoGnostics Research Campus Jena, Center of Applied Research, Philosophenweg 7, 07743 Jena, Germany
cLeibniz-Institute of Photonic Technology, Member of the Leibniz Research Alliance – Leibniz Health Technologies, Albert-Einstein-Str. 9, 07745 Jena, Germany
dCluster of Excellence Balance of the Microverse, Friedrich Schiller University Jena, 07743 Jena, Germany
First published on 13th March 2025
Abstract
Accurate and reliable bacterial identification at the genus and species levels is essential for effective clinical diagnostics. Pathogens such as Clostridium perfringens, Bacillus cereus, Clostridioides difficile, and Paraclostridium sordellii pose significant challenges due to their unique cultivation requirements and developmental traits. Building on our previous work demonstrating the differentiation of vegetative Clostridium cells from non-Clostridium genera, we now aim to extend this approach to distinguish endospores of the same species. Raman spectroscopy was utilized to develop a comprehensive library of endospore spectra, encompassing both pathogenic and non-pathogenic species. This extensive dataset forms the foundation for advanced analytical capabilities. Chemometric analysis of single-endospore Raman spectra revealed significant discriminatory power across multiple hierarchical levels, facilitating the distinction between vegetative cells and endospores. Furthermore, this method enabled precise genus- and species-level classification of endospores, underscoring its potential for high-resolution bacterial endospore identification. These results highlight the versatility and efficacy of Raman spectroscopy in addressing the challenges associated with the identification of bacterial endospores in diverse clinical and environmental contexts. These findings present the first comprehensive library of endospore Raman spectra, demonstrating that Raman spectroscopy combined with chemometric analysis is a robust and reliable method for differentiating endospores of Clostridium species from those of Bacillus, Clostridioides, and Paraclostridium. This approach holds significant promise as a precise diagnostic tool for bacterial endospore identification in clinical settings.
Introduction
Under unfavourable conditions such as high temperatures, drought, exposure to chemicals, radiation, or scarcity of nutrients, vegetative cells of the bacilli and clostridia possess the ability to undergo a transformative process known as sporulation.1 This unique mechanism results in the production of the endospore, a dormant and extremely resistant phase that allows the bacteria to live for thousands of years.2 Endospores develop high resilience by embedding core macromolecules, enzymes, and metabolites in a dehydrated calcium dipicolinate (Ca-DPA) gel.3 The Ca-DPA gel is enclosed by a semi-crystalline inner spore membrane, which gives the metabolically inactive spore the ability to withstand external factors including heat, UV and solvents. However, endospores maintain a sensory mechanism, which is triggered by favourable environmental conditions and converts the spore back into a vegetative cell.1,4,5Spores from Clostridium perfringens, Clostridioides difficile (formerly known as Clostridium difficile6), Paraclostridium sordellii (formerly known as Clostridium sordellii7), and Bacillus cereus can be isolated from diverse environments, including animal gastrointestinal tracts and carcasses, wastewater, lawns, hospital rooms, and soil.8 Infections by these pathogens typically are initiated upon ingestion of spores, although C. perfringens can also enter the body via contaminating wounds. Upon sensing small-molecule germinants, spores from these pathogens germinate and outgrow into toxin-secreting vegetative cells.8C. perfringens is the second most prevalent bacterial cause of foodborne disease in the US and the most common Clostridium species linked to gas gangrene.9–11 Other pathogenic Clostridium species include the tetanus causing agent C. tetani and C. botulinum, which causes botulism.1 Furthermore, medical interest has increased in the designated “superbug” C. difficile in the past two decades, which is primarily linked to antibiotic-associated diarrhea.12,13P. sordellii has been associated with rare postabortion infections, pneumonia, endocarditis, arthritis, peritonitis, and myonecrosis.14,15 As a result, innovative techniques must be explored and developed for a rapid and reliable detection of endospores of pathogenic bacteria such as Clostridium.
One such novel approach is Raman spectroscopy (RS). This method converts the target's vibrational information into spectral information, allowing for identification and characterisation of the microorganism. RS is a cost-effective, quick, and easy-to-use identification approach that is non-destructive, culture- and label-free.16 RS records a sample's whole biochemical makeup (“spectroscopic molecular fingerprint”), allowing for chemometric evaluation. This allows for the characterisation, differentiation, and identification of bacteria at the species and subspecies levels.17,18 In a clinical environment, this would be extremely useful for identifying the causal pathogen and making a quick diagnosis.18
However, due to their widespread existence in nature, spore-forming bacteria have received increased attention in terms of sporulation,19,20 germination,21–23 the structure and content of endospores,19,24,25 and the evaluation of various inactivation strategies.26–29 Setlow et al. found that the water content of the core, the presence of small acid-soluble proteins (SASPs) associated to the DNA, spore coat proteins, core mineral ions, and dipicolinic acid (DPA) can all impact endospore resilience.30 Raman spectroscopy has already been used to detect specific cell components, with various research groups focusing on the Raman bands of DPA's calcium complex. Surface enhanced Raman spectroscopy (SERS) has been shown to be a reliable method to detect varying concentrations of Ca-DPA in single spores31,32 as well as pure Ca-DPA.33 Raman spectroscopy has also been applied to detect DPA in bacterial samples34 and to determine the Ca-DPA concentration in single spore samples.35
Öberg et al. showed that spore metabolic activity and germination of spores of B. cereus can be tracked using heavy water and micro-Raman spectroscopy.23 Calcium dipicolinate, due to its high concentration in bacterial spores, has been effectively detected in single spores using Raman spectroscopy,36–38 SERS,31,39–42 UV resonance Raman,35,43 coherent anti-Stokes Raman spectroscopy44,45 and confocal Raman spectroscopy coupled with laser tweezers (LTRS).46–48 LTRS allows for the quantitative assessment of Ca-DPA levels in individual spores,47 the detection of single spores in aqueous solution without the need for reagents,46 as well as the monitoring of the kinetics of germination processes.48 SERS was used to not only monitor the germination kinetics of Bacillus subtilis endospores31 but was also used to monitor the DPA release.39 Raman imaging has also been employed to monitor, detect and identify spore-forming bacteria and spores.49,50
Our previous study demonstrated the ability to accurately differentiate vegetative cells of Clostridium from non-Clostridium genera, including Bacillus, Clostridioides, and Paraclostridium, across multiple hierarchical levels using Raman spectroscopy combined with chemometric evaluation. Additionally, we demonstrated that cultivation conditions—whether aerobic or anaerobic—significantly influence both the Raman spectra and the subsequent chemometric evaluation. These findings underscore the importance of accounting for cultivation methods when analysing bacterial samples, as they can impact the accuracy and reliability of spectral differentiation and classification.51
Building on this foundation, the current study extends the scope by focusing on the same bacterial species while exploiting a key physiological feature—their ability to produce endospores. Endospores, known for their resilience and unique structural properties, present additional challenges and opportunities for precise classification. Previous studies have addressed the question of whether bacterial spores can be detected or monitored. They have identified specific biomarker bands in the Raman spectrum and the germination kinetics from spore to vegetative cell. However, to the best of our knowledge, no one has researched anaerobic spore-forming Clostridia, nor has a comprehensive endospore database been established based on Bacillus and Clostridium Raman spectra. To address this, Raman spectroscopy, with its sensitivity to molecular composition, is employed to analyse and distinguish the endospores of Bacillus and Clostridium species, as well as representative strains of Paraclostridium and Clostridioides. We then used Raman spectroscopy to compile an extensive database of Raman spectra from both pathogenic and non-pathogenic bacterial endospores. Once we had gathered our data, we proceeded to assemble it, carry out chemometric analysis, and create a computed model. Finally, we challenged our model using an independent validation process. This approach not only deepens our understanding of bacterial endospore differentiation but also highlights the potential of Raman spectroscopy in expanding the toolkit for advanced microbial diagnostics.
Materials and methods
Strains and cultivation conditions
A detailed description of each species/strain as well as cultivation condition used within this study is provided in Table S1.† Most of the pathogenic and apathogenic bacteria were either obtained from the German Collection of Microorganisms and Cell Culture GmbH (DSMZ), Braunschweig, Germany, the University Hospital Jena (UKJ), Germany, and the American Type Culture Collection (ATCC), Manassas, Virginia, USA.All Bacillus strains were grown either at 30 °C or at 37 °C (B. cereus DSM345 and DSM351) on modified nutrient agar (NA) for 8 to 10 days to promote sporulation. Briefly, the modified nutrient agar was prepared with peptone from meat 5.0 g l−1, yeast extract 2.0 g l−1, meat extract 1.0 g l−1, agar 15 g l−1, and MgSO4·7H2O 500 mg L−1 at pH 7.0.52
The Clostridium, Paraclostridium and Clostridioides strains were grown at 37 °C under anaerobic conditions within an anaerobic incubation system by Anaerocult and using Anaerocult A pads for the generation of an oxygen-deprived environment in an anaerobic jar (VWR, Darmstadt, Germany) for 10 to 14 days to promote endospore production. The BHI agar was prepared with HM infusion powder 12.5 g L−1, brain heart infusion powder 5 g L−1, proteose peptone 10 g L−1, dextrose (glucose) 2 g L−1, sodium chloride 5 g L−1, disodium phosphate 2.5 g L−1 and agar 15 g L−1 at pH 7.4 ± 0.2. Alternatively, 52 g L−1 of completed BHI agar can be dissolved and distributed in Petri dishes after autoclaving at 121 °C for 15 min. Most of the anaerobic strains had to be pre-cultured on Columbia Sheep Blood Agar (VWR, Darmstadt, Germany) before being able to efficiently grow and produce the desired number of endospores on BHI agar.
Furthermore, certain clostridial species such as C. butyricum and C. tertium were grown on modified nutrient agar (NA) containing 100 mg l−1 CaCl2·2H2O rather than MgSO4·7H2O.52 Moreover, C. perfringens was grown on tryptose sulfite cycloserine (TSC) agar containing enzymatic digest of casein 15 g l−1, soy peptone 5 g l−1, yeast extract 5 g l−1, sodium meta-bisulfite 1 g l−1, ferric ammonium citrate 1 g l−1, cycloserine 0.4 g l−1, egg yolk emulsion 50 ml, and 18 g l−1 agar (VWR, Darmstadt, Germany). Alternatively, pre-made Petri dishes are commercially available (VWR, Darmstadt, Germany). All the chemicals used for the media production were bought through VWR (Darmstadt, Germany) and Merck (Darmstadt, Germany).
Sample preparation
The cultured strains were scraped from the plate and put into 1 ml sterile dH2O-filled Eppendorf tubes (VWR, Darmstadt, Germany). On a case-by-case basis, some strains were suspended for one to three days in 1 ml sterile dH2O for vegetative cells to lyse and/or release endospores from their mother cells. The cell suspension was then rinsed and centrifuged three times with 1 ml sterile dH2O at 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 rcf for 5 minutes at 4 °C. Next, the pellet was resuspended in 200 μl dH2O. After taking 10 μl of the solution with an Eppendorf pipette, small droplets were deposited on nickel foil. If the sample was too turbid or the cell count was too high to assess single cells or spores, serial dilutions were used. After the suspension droplets were dried at room temperature on the nickel foil, they were measured by Raman spectroscopy.
000 rcf for 5 minutes at 4 °C. Next, the pellet was resuspended in 200 μl dH2O. After taking 10 μl of the solution with an Eppendorf pipette, small droplets were deposited on nickel foil. If the sample was too turbid or the cell count was too high to assess single cells or spores, serial dilutions were used. After the suspension droplets were dried at room temperature on the nickel foil, they were measured by Raman spectroscopy.
Raman measurements
The BioParticleExplorer (BPE4, rap.ID, Berlin, Germany) with a frequency-doubled (532 nm), solid-state diode pumped Nd:YAG laser (LCM-S-11-NNP25, Laser-export Co. Ltd, Moscow, Russia) and an Olympus MPL-FLN-BD 100× objective (Olympus Corporation, Tokyo, Japan) was used to perform Raman spectroscopic measurements of single endospores. The 100× objective focuses Raman excitation light onto the sample with a spot size of less than 1 μm and a laser power of 10 mW. The backscattered Raman light is diffracted using a single-stage monochromator with a 920 lines per mm grating (HE532, Horiba Jobin Yvon, Bensheim, Germany) before being detected using a thermoelectrically cooled CCD (DV 401 BV; Andor Technology, Belfast, UK). As a result, a spectral resolution of about 8 cm−1 is provided.Before measuring the endospore samples, 4-acetamidophenol (4-AAP) was measured for the calibration of the wavenumber axis at 50% laser intensity for 5 s. For each sample 50–60 spectra were obtained in three biological replicates. To make sure that the signal-to-noise ratios are comparable between the bacteria, most of the endospores were measured at 10% laser intensity with an exposure time of 5 s to ensure that the spores do not burst. Moreover, the B. cereus spores were more robust and were measured for 5 s with a 25% laser intensity. Additionally, some of the samples had to be measured slightly out of focus to not rupture the spores. Notably, some of the endospores appeared “coffee bean”-shaped.
Data pre-processing
The data analysis of the raw Raman spectra is a critical step since it removes unwanted artefacts and variation within the different spectra, while also enhancing certain key differences between the various bacterial spore spectra. The data pre-processing was performed using the Ramanmetrix software for Raman data analysis containing a vast array of different steps.53 The first step is the removal of artefacts such as cosmic interferences in form of spikes by cutting off below 350 and above 3150 cm−1 as well as the wavenumber axis calibration with the measured spectra of the 4-AAP to correct the peak position and make measurements of different days comparable.54,55 A sensitive nonlinear iterative peak (SNIP) clipping algorithm was used to make a baseline correction and to remove the fluorescent background.56 Finally, the silent region from 1800 to 2600 cm−1 was cropped and vector normalization was applied before the pre-processed mean spectra were calculated.Evaluation of the endospores
To compare the mean spectra of the bacterial endospores, a combination of principal component analysis (PCA) and linear discriminant analysis (LDA) was chosen. PCA-LDA models have already been proven to be highly effective models suited for classifying and identifying different bacterial species based on their Raman spectra. Hereby, the number of principal components was optimized to a certain amount and a random N-fold cross validation was performed. For species-level differentiation, principal component analysis (PCA) was combined with a support vector machine (SVM). SVM is a supervised learning algorithm designed for classification, which determines the optimal margin or hyperplane in an n-dimensional space (where n represents the number of features) to effectively separate data points into distinct categories. To enhance classification performance, the number of features was adjusted according to the specific classification task. The same model was then applied for the validation of the independent test data and identification of independent bacterial strains which were previously not included in the main data set.Results and discussion
In this study, the bacteria were chosen based upon their ability to produce endospores, phylogenetic relatedness and clinical relevance. Raman spectra of six Bacillus and eight Clostridium species as well as ten C. difficile and one P. sordellii strains were acquired. As already mentioned in the materials and methods section, each species was cultivated either aerobically (Bacillus) or anaerobically (Clostridium, Paraclostridium and Clostridioides), whereas individual species were cultivated using different media compositions. Additionally, Raman spectra of pure substances relevant to the Raman spectra of the endospores were measured as reference spectra. These reference spectra can be found in the ESI Fig. S1.†Raman band assignment
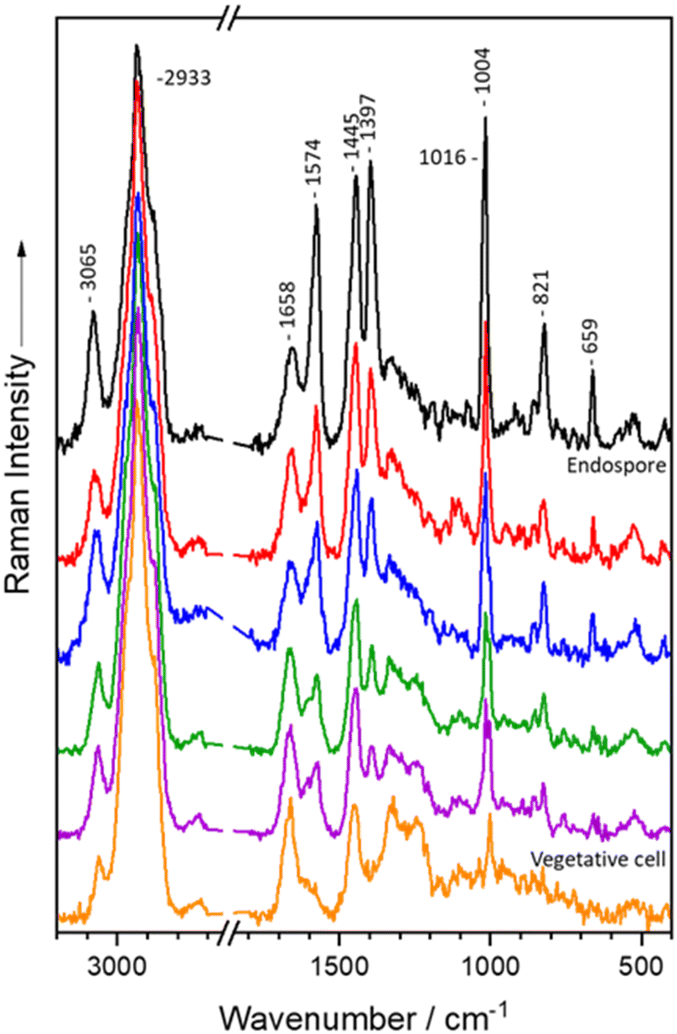
The mean Raman spectra from single vegetative cells and endospores of Bacillus and Clostridium are presented in Fig. 1. The Raman spectra of vegetative cells exhibit the characteristic spectral pattern found in bacterial spectra, exhibiting the whole spectrum of biomolecules.16,17,57Bacillus and Clostridium spectra are comparable in terms of the typical bacterial spectral signals: the C–H stretching vibrations are located at 2933 cm−1,58 whereas the CH2/CH3 deformation vibrations are found at 1451 cm−1,59–61 which are primarily found in lipids and proteins. Furthermore, the Raman signals at 1664 cm−1 (ref. 62 and 63) and 1250 cm−1 (ref. 64) correspond to amide I and amide III vibrations of proteins. Additionally, the ring breathing vibration of phenylalanine is located at 1004 cm−1,61 while the out of plane ring deformation vibration and the second ring breathing mode of tyrosine give rise to bands at 854 cm−1 (ref. 64 and 65) and 824 cm−1,27 respectively. Moreover, nucleic acids can be seen at 1583 cm−1.62,65 Finally, Bacillus vegetative cells have unique Raman signals at 1583, 1397, 1313, 1127, and 749 cm−1 (ref. 66–68) that correspond to cytochrome vibrations but are absent in Clostridium spectra. | ||
| Fig. 1 Mean spectra of endospores of Bacillus (A), Clostridium (B), Paraclostridium and Clostridioides (C) species: (Bt) B. thuringiensis, (Bs) B. spizizenii, (Ls) L. sphaericus, (Bm) B. mycoides, (Bl) B. licheniformis, (Bc) B. cereus, (Ct) C. tertium, (Csp) C. sporogenes, (Cse) C. septicum, (Cp) C. perfringens, (Cf) C. felsineum, (Cc) C. cadaveris, (Cbu) C. butyricum, (Cbe) C. beijerinckii, (Cd) C. difficile and (Ps) P. sordellii. | ||
In contrast, the endospore spectra revealed unique Raman signals at 1574, 1445, 1397, 1016, 821 and 659 cm−1 (ref. 69 and 70) which correspond to the calcium dipicolinate content in the spore core. At 1574 cm−1 the skeletal vibration of the pyridine ring is observed, whereas 821 cm−1 corresponds to the C–COO– stretching vibration.69,70 The pyridine ring breathing vibrations can be detected at 1445 cm−1 and 1016 cm−1, while 1397 cm−1 represents the OCO symmetric stretching vibration.69,70 Additionally, the aromatic C–H stretching vibration at 3080 cm−1 can be attributed to Ca-DPA.69,70 Ca-DPA also superimposes signals from different biomolecules such as the ring breathing mode of tyrosine at 821 cm−1,64,65 the CH2/CH3 deformation mode of proteins and lipids at 1450 cm−1,59–61 as well as the ring vibrations of adenine and guanine at 1574 cm−1.62,65
Additionally, the difference spectra in Fig. 2 were derived by subtracting the pre-processed vegetative cell mean spectra from the endospore mean spectra. This was performed once for Bacillus and once for Clostridium. Both spectra show the unique Ca-DPA Raman signals at 1574, 1445, 1397, 1016, 821, and 659 cm−1 on the positive side,69,70 emphasising the fact that Ca-DPA can only be found in endospores and not in vegetative cells. In addition to the Ca-DPA bands, Bacillus vegetative cells exhibited distinct cytochrome marker bands at 1313, 1127, and 749 cm−1.66–68 However, the cytochrome signals at 1583 and 1397 cm−1 (ref. 66–68) can not be seen in the difference spectrum for the vegetative cells because they are superimposed by the Ca-DPA signals that arise.69,70
 | ||
| Fig. 2 Mean Raman spectra of endospores and vegetative cells of Bacillus (A) and Clostridium (B) as well as difference spectra calculated with the mean spectra (endospore – vegetative cell). | ||
Endospore development
Endospore formation, or sporulation, is a tightly regulated developmental process employed by Bacillus and Clostridium species to endure adverse environmental conditions. This process allows these bacteria to transit from a vegetative state to a metabolically dormant and structurally robust endospore, capable of withstanding extreme stressors such as heat, desiccation, UV radiation, and chemical exposure.The specific Raman bands associated with calcium dipicolinate (Ca-DPA) arise from the skeletal vibrations of the pyridine ring (659, 1016, 1445, and 3065 cm−1) and the vibrational modes of the carboxylate group (821, 1397, and 1574 cm−1). These characteristic Raman signals serve not only as specific biomarkers for endospores but also as tools to monitor the process of endospore formation, from the initial stages within the mother cell to the release of the mature endospore. It is evident that the mean Raman spectra of different bacterial species exhibit significant similarities, with no distinct marker bands readily identifiable as species-specific (Fig. 1). This highlights the conserved nature of Ca-DPA-related spectral features among endospore-forming species.
Fig. 3 presents individual Raman spectra obtained from different bacterial cells within the same sample, capturing distinct stages of endospore development. These spectra highlight the dynamic changes in both band positions and relative intensities as the bacterial cells transit from a vegetative state (mother cell) to a maturing endospore. Notably, the spectra reveal that the spectral features associated with the vegetative cell differ significantly from those observed in the developing endospore, reflecting the biochemical and structural changes occurring during sporulation. As the endospore matures, there is a marked increase in the incorporation of Ca-DPA, which leads to an enhanced concentration of this compound within the developing endospore. This, in turn, results in a significant intensification of the Raman signal, further emphasizing the key role of Ca-DPA in the maturation process. The progressive evolution of the Raman spectra thus provides a valuable tool for tracking the temporal dynamics of endospore formation, offering insights into the molecular mechanisms underlying bacterial survival in response to environmental stresses.
 | ||
| Fig. 3 Raman spectra of single endospore development with increasing Ca-DPA concentration in C. difficile. | ||
It is essential to clarify that the Raman spectra presented in Fig. 3 correspond to individually acquired spectra from distinct vegetative cells and endospores, rather than capturing the continuous transition of a single vegetative cell into an endospore. Furthermore, to the best of our knowledge, previous studies have only conducted germination experiments that track the transition from an endospore to a vegetative cell.21–23
To determine whether the addition of calcium, manganese, or magnesium during the sporulation process influences the Raman spectra of endospores to a significant extent, Bacillus cereus and Clostridium butyricum were cultivated on nutrient agar plates enriched with calcium, manganese, and magnesium salts (Fig. 4). This approach allowed for the evaluation of how an addition of these metal ions might affect the biochemical composition and structural properties of the resulting endospores. By comparing the spectra of endospores formed under these enriched conditions to those formed under standard conditions, the aim was to assess any notable alterations in the spectral features that could be attributed to the presence of these additional ions.
 | ||
| Fig. 4 Bacillus cereus (A) and Clostridium butyricum (B) were cultured on nutrient agar (NA) supplemented with either CaCl2, MnSO4, or MgSO4. | ||
A study by Stöckel et al.24 demonstrated the specific effects of supplementary manganese and calcium ions on the Raman spectra of Bacillus endospores. In all strains examined, endospores cultivated on manganese-rich agar exhibited a reduction in the intensity of the characteristic Ca-DPA-associated Raman signals at 1016, 1400, and 1572 cm−1. Additionally, band broadening was observed, attributed to the emergence of new bands at 1030, 1390, and 1592 cm−1. This wavenumber repositioning indicates a partial substitution of intracellular Ca-DPA with Mn-DPA, which occurs when spore-forming bacteria are exposed to manganese concentrations significantly exceeding the levels required for vegetative growth.24
However, the expected wavenumber variations in the endospore spectra of bacteria grown on calcium- or manganese-supplemented nutrient agar, as reported by Stöckel et al., were not observed in this study. As illustrated in Fig. 4, neither Bacillus cereus nor Clostridium butyricum grown on agar enriched with calcium, manganese, or magnesium exhibited any significant wavenumber shifts. According to the literature, the stability and formation of metal-ion-DPA complexes are influenced by several factors, including charge density, orbital overlap, ionic radii, solution pH, and the competition and availability of metal ions in the medium.71 Theoretically, DPA is expected to preferentially bind to Mn2+ and Mg2+ over Ca2+ due to their smaller ionic radii and higher charge densities, which result in the formation of more stable complexes compared to Ca-DPA.72,73 However, when Ca2+ is present in significantly higher concentrations than other metal ions, DPA may bind preferentially to the more abundant calcium despite the lower stability of the resulting complex.71–74
The accumulation of metal ions in endospores is primarily influenced by their concentration in the growth medium, though the specific amounts of individual metals can vary significantly. These metals are also partially exchangeable.75 Among divalent metals, calcium is most closely associated with endospore heat resistance, and its substitution with other metals has been shown to reduce heat resistance.74,76 The high accumulation of Ca2+ in spores may be attributed to its high hydrolytic stability, its relative abundance in standard bacteriological media, and its nontoxic nature.75 In contrast, Ca2+ and Mg2+ form chelates with lower stability, making them more prone to dissociation or reversible binding to DPA. Mn2+, while forming more stable complexes, is less easily dissociated, which can lead to biological toxicity.71,77
Classification and validation
A more comprehensive evaluation of the Raman spectra was performed using chemometric techniques. Principal component analysis (PCA) combined with linear discriminant analysis (LDA) was applied to distinguish between vegetative cells and endospores, as well as among different genera. PCA is a dimensionality-reduction technique that condenses large data sets into a smaller set of principal components while retaining most of the original information. LDA was then employed for classification and validation, enabling differentiation between the two morphological types and among the genera.For species-level differentiation, PCA was coupled with a support vector machine (SVM). SVM is a supervised learning method designed for classification, which identifies the optimal margin or hyperplane in an n-dimensional space (where n is the number of features) to effectively separate data points into distinct categories. The number of features was adjusted based on the classification task to optimize performance. The validation of the training set was performed by using an independent batch of the selected species.
Table 1 summarizes the classification and the validation results for each level (morphology, genus, and species). Initially, a PCA-LDA was employed to discriminate vegetative cells from endospores with an accuracy of 92.9% and sensitivity and specificity of 94.0% and 91.8%, respectively. In this analysis, more vegetative cells were misclassified as endospores (398 out of 4875) than endospores were misclassified as vegetative cells (283 out of 4730). A plausible explanation for this discrepancy could be the varying developmental stages of endospore maturation. The model may struggle to classify partially matured endospores, potentially identifying them as either vegetative cells or endospores with uncertainty.
| Classification | Validation | ||||||
|---|---|---|---|---|---|---|---|
| Level | Name | Acc./% | Sens./% | Spec./% | Acc./% | Sens./% | Spec./% |
| Morphology | Endospore (ES) | 92.9 | 94.0 | 91.8 | 97.7 | 98.4 | 97.1 |
| Vegetative cells (VC) | 91.8 | 94.0 | 97.1 | 98.4 | |||
| Genus | Bacillus (B) | 97.0 | 97.1 | 99.1 | 93.7 | 97.8 | 100.0 |
| Clostridioides (CS) | 97.9 | 99.6 | 79.9 | 99.4 | |||
| Clostridium (CM) | 96.0 | 98.6 | 99.1 | 93.0 | |||
| Paraclostridium (P) | 96.8 | 98.7 | 96.7 | 98.4 | |||
| Species (Bacillus) | B. cereus (Bc) | 97.7 | 97.0 | 98.5 | 87.1 | 48.0 | 98.5 |
| B. licheniformis (Bl) | 98.0 | 100.0 | 100.0 | 100.0 | |||
| B. mycoides (Bm) | 97.1 | 99.7 | 82.4 | 82.8 | |||
| L. sphaericus (Ls) | 99.2 | 99.3 | 100.0 | 100.0 | |||
| B. spizizenii (Bs) | 97.7 | 100.0 | 100.0 | 97.3 | |||
| B. thuringiensis (Bt) | 98.0 | 99.6 | 92.0 | 100.0 | |||
| Species (Clostridium) | C. beijerinckii (Cbe) | 93.2 | 88.4 | 100.0 | 81.8 | 93.1 | 99.8 |
| C. butyricum (Cbu) | 93.8 | 98.4 | 100.0 | 93.3 | |||
| C. cadaveris (Cc) | 98.6 | 97.5 | 100.0 | 100.0 | |||
| C. felsineum (Cf) | 90.5 | 98.8 | 88.3 | 93.8 | |||
| C. perfringens (Cp) | 94.8 | 99.8 | 98.3 | 93.5 | |||
| C. septicum (Cse) | 87.2 | 98.6 | 58.3 | 97.7 | |||
| C. sporogenes (Csp) | 97.5 | 99.8 | 100.0 | 100.0 | |||
| C. tertium (Ct) | 92.3 | 99.3 | 16.7 | 100.0 | |||
A genus-level classification was also conducted to evaluate whether endospores from different genera could be reliably distinguished. The dataset included spectra from eight Clostridium species, one Paraclostridium sordellii, ten Clostridioides difficile, and six Bacillus species. Of the 4730 endospore spectra analysed, 142 were misclassified. Clostridium exhibited the highest misclassification rate with 67 spectra, followed by Bacillus and Clostridioides with 33 and 34 misclassifications, respectively. Paraclostridium, represented by only a single species, had the lowest misclassification rate, with 8 out of 250 Raman spectra being incorrectly classified. Thus, the genus-level LDA achieved an overall classification accuracy of 97.0% for the 4730 Raman spectra. Additionally, the model demonstrated sensitivities and specificities ranging from 96.0% to 99.6%, respectively, indicating a robust performance in distinguishing among the genera. The LDA plot (Fig. S2†) further visualizes the discriminatory power of the model, where each dot represents an individual Raman spectrum of one single bacterial cell corresponding to a specific genus.
At the species level, 97.7% of the 1153 Bacillus spectra were correctly classified, while the 1688 Clostridium spectra were classified with an accuracy of 93.2% (Table 1). Minor misclassifications occurred within the B. cereus group (B. cereus, B. licheniformis, and B. mycoides), which are phylogenetically closer to one another than to other Bacillus species in the dataset. Additionally, B. spizizenii and B. thuringiensis occasionally showed misclassifications with B. cereus. Notably, L. sphaericus achieved the highest classification accuracy, with only one misclassified spectrum. The Raman spectra for this species revealed signs of metabolic activity, such as the presence of cytochrome, suggesting that the endospore might still reside within the mother cell or was not yet fully matured. Despite these challenges, the model exhibited high sensitivities and specificities at the species level, ranging from 97.0% to 100.0%.
For Clostridia, 1573 out of 1688 Raman spectra were correctly classified, resulting in an overall accuracy of 93.2%. Minor misclassifications occurred primarily among C. beijerinckii, C. butyricum, C. felsineum, C. sporogenes, and C. tertium, which were occasionally misclassified as C. cadaveris. In contrast, C. cadaveris showed only a single misclassification with C. felsineum, C. septicum, and C. tertium. These errors can be attributed to the close phylogenetic relationships among these species. A more significant misclassification involved C. septicum, where 16 out of the 23 misclassified spectra (23/180) were incorrectly assigned to C. felsineum.
It is important to note that the position of the endospore within the mother cell can vary significantly between species and even among different strains within the same species. The endospore can either be found in a central, eccentric, subterminal, or terminal position with either swelling the mother cell or not.78 Additionally, structural variations such as the presence of appendages, an exosporium, or differences in the composition of the endospore cell walls can have a profound impact.79 These structural differences may influence the Raman spectra by altering the biochemical signals detected, ultimately affecting classification accuracy. Such variability highlights the need for careful consideration of these factors when interpreting Raman spectral data and developing classification models for bacterial endospores. Despite these misclassifications, the results underscore the overall robustness of the method in identifying and classifying Clostridia.
The classification of Paraclostridium sordellii and Clostridioides difficile on a species-level was not performed, because only one representative of each species was used in this dataset. Additionally, a strain-level classification of the ten C. difficile strains was not performed. The detailed classification results can be seen in Table 1 and Tables S3–6.†
Each finalized PCA-LDA and PCA-SVM model underwent a validation process using independently cultivated bacterial species previously included in the training set. The validation dataset comprised the same eight Clostridium species, six Bacillus species, one Paraclostridium sordellii species, and ten strains of Clostridioides difficile. Independent batches, defined as biological replicates separately cultivated and excluded from the training phase, were used to evaluate the sensitivity of the models. The constructed models were applied to predict the classifications of these independent validation sets, with the validation process performed at each classification level. For example, the morphology of the validation set was compared to the morphology classification model, and the same approach was applied for genus- and species-level validation (Table 1). However, species-level validation was not conducted for P. sordellii, nor was strain-level validation performed for the ten C. difficile strains. This procedure ensured the robustness of the models across different hierarchical levels of classification.
Of the 1159 endospore spectra, 1140 were correctly classified, while 1214 of 1250 spectra were accurately assigned to vegetative cells, resulting in an overall accuracy of 97.7% (Table S7†). At the genus level, validation achieved an accuracy of 93.7%, with Clostridioides exhibiting 59 misclassifications as Clostridium. Misclassifications among the other three genera were minimal. Sensitivity at the genus level ranged from 96.7% to 99.1%, except for Clostridioides, which achieved only 79.9% due to the higher rate of misclassification. Specificities were generally high, ranging from 93.0% to 100.0% (Table S8†). At the species level, validation accuracies were lower, with Bacillus achieving 87.1% and Clostridium achieving 81.8% (Tables S9–10†). These results highlight challenges in species-level differentiation, likely due to overlapping spectral features among closely related species (Table 1).
Compared to other genera, Bacillus achieved a species-level accuracy of 87.1% during validation, which is notably lower than the classification model's accuracy of 97.7% (Table 1). The primary source of misclassification was B. cereus, where 52 out of 100 spectra were incorrectly classified as B. mycoides. This misclassification aligns with findings from our previous study on vegetative cells of Bacillus species,51 which reported frequent misclassification of B. cereus as B. mycoides due to their close phylogenetic relationship within the B. cereus group. In contrast, B. mycoides exhibited only one misclassification as B. cereus, but eight misclassifications as B. spizizenii. These results underscore the challenge of distinguishing species within closely related groups, particularly the B. cereus group.
Clostridium achieved a validation accuracy of 81.8%, nearly identical to the validation performance of Bacillus, yet a decreased from its classification model (Table 1). The primary misclassifications involved C. tertium being incorrectly classified as C. butyricum and C. septicum as C. felsineum. Notably, C. tertium and C. butyricum are relatively close in the Clostridium sensu stricto phylogenetic tree, which likely explains their misclassification. Consequently, the sensitivity for C. tertium dropped significantly from 92.3% to 16.7%, while for C. septicum, it decreased from 87.2% to 58.3%. In contrast, C. butyricum, C. cadaveris, and C. sporogenes achieved 100.0% sensitivities, demonstrating robust validation performances for these species. Meanwhile, C. difficile and P. sordellii were validated only at the genus level since each was represented by a single species in the dataset. Tables S7–10† showed more detailed validation results for each hierarchical level of validation.
This study aimed to establish an extensive Raman spectroscopy dataset for endospores of Clostridium and Bacillus species, as well as C. difficile and P. sordellii. Anaerobic bacteria are often overlooked due to the challenges associated with their cultivation, despite their clinical and environmental significance. Notably, Raman spectroscopy proved highly effective in distinguishing these four genera across multiple hierarchical levels, achieving remarkably high classification and validation accuracies.
The method demonstrated the ability to easily differentiate between endospores and vegetative cells and yielded comparable species-level classification results for Bacillus and Clostridium. This highlights that despite their close phylogenetic relationships, the interspecies and intraspecies differences are pronounced enough to be resolved using Raman spectroscopy combined with appropriate chemometric methods. These findings underscore the critical role of selecting suitable chemometric assessments and model types in advancing the study of these often-underappreciated bacteria.
Conclusion
This study aimed to construct a comprehensive endospore Raman spectra dataset for Clostridium and Bacillus species, alongside C. difficile and P. sordellii, which are frequently underrepresented in research due to challenges associated with their phylogenetic relationships, cultivation requirements, and pathogenicity. The dataset was meticulously developed to address these challenges, incorporating samples grown under diverse conditions as well as applying rigorous spectral preprocessing methods and model constructions. Notably, Raman spectroscopy demonstrated high efficacy in distinguishing Clostridia from the recently reclassified former Clostridium species, C. difficile and P. sordellii, despite their close evolutionary relationships. This distinction is critical for both clinical diagnostics and environmental microbiology, where accurate identification of these species can significantly impact treatment strategies and ecological studies.Moreover, the findings underscore the importance of employing suitable chemometric approaches, such as principal component analysis (PCA) and machine learning classifiers, to extract meaningful patterns from the spectral data. The study also highlights how the choice of model types—linear versus nonlinear—can influence classification accuracy, particularly for challenging datasets with overlapping spectral features. This research emphasizes the potential of Raman spectroscopy as a powerful tool for robust differentiation and classification in microbial studies, offering insights into microbial taxonomy, pathogenicity, and environmental distribution. These results pave the way for broader applications of Raman-based techniques in microbiology, fostering advancements in both fundamental and applied research.
Author contributions
Markus Salbreiter: conceptualization, data curation, formal analysis, investigation, methodology, validation, visualization, writing – original draft, writing – reviewing and editing. Annette Wagenhaus: data curation, investigation. Petra Rösch: conceptualization, funding acquisition, project administration, supervision, writing – original draft, writing – reviewing and editing. Jürgen Popp: project administration, supervision, writing – original draft, writing – reviewing and editing.Data availability
Data, metadata, and a data description file are available on Zenodo at https://doi.org/10.5281/zenodo.14764655.Conflicts of interest
The authors do not have any conflicts of interests to declare.Acknowledgements
Financial support of the Federal Ministry of Education and Research, Germany (Bundesministerium für Bildung und Forschung (BMBF), Deutschland) in the project FastAlert (13GW0460B).References
- M. T. Madigan, K. S. Bender, D. H. Buckley, W. M. Sattley and D. A. Stahl, Brock Biology of Microorganisms, Pearson Education Limited, 2022, 16th edn, 2021 Search PubMed.
- L. McKane and J. Kandel, in Microbiology: Essentials and Application, 1996, pp. 66–95 Search PubMed.
- K. Johnstone, J. Appl. Bacteriol., 1994, 76, 17S–24S CrossRef PubMed.
- A. Keynan and H. Halvorson, in Spores III, American Society for Microbiology, Ann Arbor, Mich, 1965, pp. 174–179 Search PubMed.
- Spore Germination and Outgrowth, ed. M. Paidhungat and P. Setlow, ASM Press, 2001 Search PubMed.
- P. A. Lawson, D. M. Citron, K. L. Tyrrell and S. M. Finegold, Anaerobe, 2016, 40, 95–99 CrossRef PubMed.
- T. Sasi Jyothsna, L. Tushar, C. Sasikala and C. V. Ramana, Int. J. Syst. Evol. Microbiol., 2016, 66, 1268–1274 CrossRef CAS.
- M. C. Swick, T. M. Koehler and A. Driks, in Virulence mechanisms of bacterial pathogens, ed. I. T. Kudva, N. A. Cornick, P. J. Plummer, Q. Zhang, T. L. Nicholson, J. P. Bannantine and B. H. Bellaire, American Society for Microbiology, 2016, pp. 567–591, DOI:10.1128/9781555819286.ch20.
- J. E. Grass, L. H. Gould and B. E. Mahon, Foodborne Pathog. Dis., 2013, 10, 131–136 CrossRef PubMed.
- R. Kiu, J. Brown, H. Bedwell, C. Leclaire, S. Caim, D. Pickard, G. Dougan, R. A. Dixon and L. J. Hall, Anim. Microbiome, 2019, 1, 1–14 CrossRef.
- R. Kiu and L. J. Hall, Emerging Microbes Infect., 2018, 7, 1–15 CrossRef CAS PubMed.
- F. Barbut, J. Hosp. Infect., 2015, 89, 287–295 CrossRef CAS PubMed.
- T. Chopra, Clostridium Difficile Infection in Long-Term Care Facilities: A Clinician's Guide, Springer, 2019 Search PubMed.
- D. M. Aronoff, Anaerobe, 2013, 24, 98–101 CrossRef CAS PubMed.
- M. Aldape, A. Bryant and D. Stevens, Clin. Infect. Dis., 2006, 43, 1436–1446 CrossRef CAS PubMed.
- B. Lorenz, C. Wichmann, S. Stöckel, P. Rösch and J. Popp, Trends Microbiol., 2017, 25, 413–424 CrossRef CAS PubMed.
- A. Pistiki, M. Salbreiter, S. Sultan, P. Rösch and J. Popp, Transl. Biophotonics, 2022, 4, e202200011 CrossRef CAS.
- M. Salbreiter, A. Pistiki, D. Cialla-May, P. Rösch and J. Popp, in Raman Spectroscopy in Human Health and Biomedicine, ed. H. Sato, J. Popp, B. R. Wood and Y. Ozaki, 2023, pp. 337–410, DOI:10.1142/9789811264610_0010.
- J. De Gelder, P. Scheldeman, K. Leus, M. Heyndrickx, P. Vandenabeele, L. Moens and P. De Vos, Anal. Bioanal. Chem., 2007, 389, 2143–2151 CrossRef CAS PubMed.
- B. L. Liu and Y. M. Tzeng, Biotechnol. Bioeng., 2000, 68, 11–17 CrossRef CAS PubMed.
- M. Plomp, T. J. Leighton, K. E. Wheeler, H. D. Hill and A. J. Malkin, Proc. Natl. Acad. Sci. U. S. A., 2007, 104, 9644–9649 CrossRef CAS PubMed.
- D. Chen, S.-s. Huang and Y.-q. Li, Anal. Chem., 2006, 78, 6936–6941 CrossRef CAS PubMed.
- R. Öberg, T. Dahlberg, D. Malyshev and M. Andersson, Analyst, 2023, 148, 2141–2148 RSC.
- S. Stöckel, S. Meisel, R. Boehme, M. Elschner, P. Rösch and J. Popp, J. Raman Spectrosc., 2009, 40, 1469–1477 CrossRef.
- S. Stöckel, S. Meisel, M. Elschner, P. Rösch and J. Popp, Angew. Chem., Int. Ed., 2012, 51, 5339–5342 CrossRef PubMed.
- S. Stöckel, S. Meisel, M. Elschner, P. Rösch and J. Popp, Anal. Chem., 2012, 84, 9873–9880 CrossRef PubMed.
- S. Stöckel, W. Schumacher, S. Meisel, M. Elschner, P. Rösch and J. Popp, Appl. Environ. Microbiol., 2010, 76, 2895–2907 CrossRef PubMed.
- D. Malyshev, T. Dahlberg, K. Wiklund, P. O. Andersson, S. Henriksson and M. Andersson, Anal. Chem., 2021, 93, 3146–3153 CrossRef CAS PubMed.
- S. Wang, C. J. Doona, P. Setlow and Y.-q. Li, Appl. Environ. Microbiol., 2016, 82, 5775–5784 CrossRef CAS PubMed.
- P. Setlow, J. Appl. Microbiol., 2006, 101, 514–525 CrossRef CAS.
- J. K. Daniels, T. P. Caldwell, K. A. Christensen and G. Chumanov, Anal. Chem., 2006, 78, 1724–1729 CrossRef CAS PubMed.
- J. Chan, A. Esposito, C. Talley, C. Hollars, S. Lane and T. Huser, Anal. Chem., 2004, 76, 599–603 CrossRef CAS.
- S. E. Bell, J. N. Mackle and N. M. Sirimuthu, Analyst, 2005, 130, 545–549 RSC.
- S. Farquharson, L. Grigely, V. Khitrov, W. Smith, J. F. Sperry and G. Fenerty, J. Raman Spectrosc., 2004, 35, 82–86 CrossRef CAS.
- W. Nelson, R. Dasari, M. Feld and J. Sperry, Appl. Spectrosc., 2004, 58, 1408–1412 CrossRef CAS PubMed.
- A. P. Esposito, C. E. Talley, T. Huser, C. W. Hollars, C. M. Schaldach and S. M. Lane, Appl. Spectrosc., 2003, 57, 868–871 CrossRef CAS PubMed.
- P. Rösch, M. Harz, M. Schmitt, K.-D. Peschke, O. Ronneberger, H. Burkhardt, H.-W. Motzkus, M. Lankers, S. Hofer and H. Thiele, Appl. Environ. Microbiol., 2005, 71, 1626–1637 CrossRef PubMed.
- P. Rösch, M. Harz, K. D. Peschke, O. Ronneberger, H. Burkhardt and J. Popp, Biopolymers, 2006, 82, 312–316 CrossRef PubMed.
- D. D. Evanoff, J. Heckel, T. P. Caldwell, K. A. Christensen and G. Chumanov, J. Am. Chem. Soc., 2006, 128, 12618–12619 CrossRef CAS PubMed.
- J. Guicheteau, L. Argue, D. Emge, A. Hyre, M. Jacobson and S. Christesen, Appl. Spectrosc., 2008, 62, 267–272 CrossRef CAS PubMed.
- X. Zhang, M. A. Young, O. Lyandres and R. P. Van Duyne, J. Am. Chem. Soc., 2005, 127, 4484–4489 CrossRef CAS PubMed.
- C. Camerlingo, G. Di Meo, M. Lepore, M. Lisitskiy, A. Poli, M. Portaccio, I. Romano and P. Di Donato, Sensors, 2020, 20, 4150 CrossRef CAS PubMed.
- E. Ghiamati, R. Manoharan, W. Nelson and J. Sperry, Appl. Spectrosc., 1992, 46, 357–364 CrossRef CAS.
- N. Coluccelli, G. Galzerano, P. Laporta, K. Curtis, C. L. Lonsdale, D. Padgen, C. R. Howle and G. Cerullo, Sci. Rep., 2023, 13, 2634 CrossRef CAS PubMed.
- N. Coluccelli, G. Cichelli, P. Laporta and G. Cerullo, Opt. Express, 2023, 31, 23245–23259 CrossRef CAS.
- C. Xie, J. Mace, M. A. Dinno, Y. Q. Li, W. Tang, R. J. Newton and P. J. Gemperline, Anal. Chem., 2005, 77, 4390–4397 CrossRef CAS.
- S.-s. Huang, D. Chen, P. L. Pelczar, V. R. Vepachedu, P. Setlow and Y.-q. Li, J. Bacteriol., 2007, 189, 4681–4687 CrossRef CAS PubMed.
- L. Kong, P. Zhang, G. Wang, J. Yu, P. Setlow and Y.-q. Li, Nat. Protoc., 2011, 6, 625–639 CrossRef CAS PubMed.
- K. S. Kalasinsky, T. Hadfield, A. A. Shea, V. F. Kalasinsky, M. P. Nelson, J. Neiss, A. J. Drauch, G. S. Vanni and P. J. Treado, Anal. Chem., 2007, 79, 2658–2673 CrossRef CAS PubMed.
- A. Tripathi, R. E. Jabbour, P. J. Treado, J. H. Neiss, M. P. Nelson, J. L. Jensen and A. P. Snyder, Appl. Spectrosc., 2008, 62, 1–9 CrossRef CAS PubMed.
- M. Salbreiter, A. Wagenhaus, P. Rösch and J. Popp, Anal. Chem., 2024, 96, 15702–15710 CrossRef CAS.
- E. Bast, Mikrobiologische Methoden: Eine Einführung in grundlegende Arbeitstechniken, Springer, 2014 Search PubMed.
- D. Storozhuk, O. Ryabchykov, J. Popp and T. Bocklitz, arXiv, 2022, preprint, arXiv.2201.07586, DOI:10.48550/arXiv.2201.07586.
- T. Dörfer, T. Bocklitz, N. Tarcea, M. Schmitt and J. Popp, Z. Phys. Chem., 2011, 225, 753–764 CrossRef.
- O. Ryabchykov, W. S. Iwan, J. Popp and T. Bocklitz, Spectroscopy, 2022, 37, 48–50 Search PubMed.
- S. Guo, T. Bocklitz and J. Popp, Analyst, 2016, 141, 2396–2404 RSC.
- S. Stöckel, J. Kirchhoff, U. Neugebauer, P. Rösch and J. Popp, J. Raman Spectrosc., 2016, 47, 89–109 CrossRef.
- K. Czamara, K. Majzner, M. Z. Pacia, K. Kochan, A. Kaczor and M. Baranska, J. Raman Spectrosc., 2015, 46, 4–20 CrossRef CAS.
- J. M. Benevides, S. A. Overman and G. J. Thomas Jr, J. Raman Spectrosc., 2005, 36, 279–299 CrossRef CAS.
- S. Managò, G. Zito and A. C. De Luca, Opt. Laser Technol., 2018, 108, 7–16 CrossRef.
- N. Töpfer, M. M. Müller, M. Dahms, A. Ramoji, J. Popp, H. Slevogt and U. Neugebauer, Integr. Biol., 2019, 11, 87–98 CrossRef PubMed.
- K. Maquelin, C. Kirschner, L. P. Choo-Smith, N. van den Braak, H. P. Endtz, D. Naumann and G. J. Puppels, J. Microbiol. Methods, 2002, 51, 255–271 CrossRef CAS PubMed.
- N. Uzunbajakava, A. Lenferink, Y. Kraan, E. Volokhina, G. Vrensen, J. Greve and C. Otto, Biophys. J., 2003, 84, 3968–3981 CrossRef CAS.
- G. Azemtsop Matanfack, A. Pistiki, P. Rösch and J. Popp, Life, 2021, 11, 1003 CrossRef CAS.
- W. E. Huang, M. Li, R. M. Jarvis, R. Goodacre and S. A. Banwart, Adv. Appl. Microbiol., 2010, 70, 153–186 CAS.
- Y.-S. Huang, T. Karashima, M. Yamamoto, T. Ogura and H.-o. Hamaguchi, J. Raman Spectrosc., 2004, 35, 525–526 CrossRef CAS.
- M. Kakita, V. Kaliaperumal and H.-o. Hamaguchi, J. Biophotonics, 2012, 5, 20–24 CrossRef CAS PubMed.
- T. C. Strekas and T. G. Spiro, Biochim. Biophys. Acta, Protein Struct., 1972, 278, 188–192 CrossRef CAS PubMed.
- P. Carmona, Spectrochim. Acta, Part A, 1980, 36, 705–712 CrossRef.
- W. H. Woodruff, T. G. Spiro and C. Gilvarg, Biochem. Biophys. Res. Commun., 1974, 58, 197–203 CrossRef CAS PubMed.
- L. Chung, K. S. Rajan, E. Merdinger and N. Grecz, Biophys. J., 1971, 11, 469–482 CrossRef CAS PubMed.
- H. R. Curran, B. Brunstetter and A. Myers, J. Bacteriol., 1943, 45, 485–494 CrossRef CAS PubMed.
- B. J. Kolodziej and R. A. Slepecky, J. Bacteriol., 1964, 88, 821–830 CrossRef CAS PubMed.
- M. T. Sinnelä, A. M. Pawluk, Y. H. Jin, D. Kim and J.-H. Mah, Front. Microbiol., 2021, 12, 744953 CrossRef PubMed.
- R. Slepecky and J. Foster, J. Bacteriol., 1959, 78, 117–123 CrossRef CAS PubMed.
- W. Murrell, Spore formation and germination as a microbial reaction to the environment, presented at the Symp. Soc. Gen. Microbiol, 1961, Vol. 11, pp. 100–150.
- T. Tang, K. Rajan and N. Grecz, Biophys. J., 1968, 8, 1458–1474 CrossRef CAS PubMed.
- P. Dürre, Handbook on Clostridia, CRC Press, Boca Raton, 1st edn, 2005 Search PubMed.
- P. Vos, G. Garrity, D. Jones, N. R. Krieg, W. Ludwig, F. A. Rainey, K.-H. Schleifer and W. B. Whitman, Bergey's Manual of Systematic Bacteriology: Volume 3: The Firmicutes, Springer-Verlag, New York, 2011 Search PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d5an00115c |
| This journal is © The Royal Society of Chemistry 2025 |