
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Effect of solid-state fermentation with Pleurotus ostreatus on the protein content and other nutritional components of de-seeded carob fruits
Shahab
Iqbal
,
Özge
Tirpanalan-Staben
and
Knut
Franke
 *
*
Leibniz University Hannover, Institute of One Health, Hannover, Germany. E-mail: franke@foh.uni-hannover.de
First published on 26th July 2024
Abstract
Carob pulp is a currently underutilized by-product of locust bean gum production from carob pods. It is dense in several valuable nutritional components such as carbohydrates, dietary fiber, minerals and some bioactive substances but low in protein. One of the strategies to facilitate its valorization is suggested as enhancing its protein content. Therefore, solid-state fermentation (SSF) of carob pulp was investigated using the edible fungus species Pleurotus ostreatus. For this purpose, carob pulp was first soaked with water, then treated thermally at 121 °C and subsequently subjected to SSF. During the SSF period of 30 days, samples were drawn every 5 days and analyzed with respect to protein, reducing sugars (glucose and fructose) and dietary fiber contents as well as polyphenol content and antioxidative properties. SSF with Pleurotus ostreatus could be proven to triple the protein content as well as the total fiber content of carob pulp. An increase in protein content is likely connected to the production of fungal biomass. However, a decrease in the reduced sugar content as well as in the antioxidant properties was observed. Results confirmed that SSF using Pleurotus ostreatus could be applied to valorize the carob pulp by boosting its protein content, and thereby a contribution to the circular bioeconomy in food production could be suggested.
Sustainability spotlightUp to now, the production of high-quality and safe foods leads to the generation of by-products, which normally leave the food chain and are often under-utilized. The work to be published in ‘Sustainable Food Technology’ contributes to the reduction of food waste by improving the valorization of a by-product from the production of a stabilizer. Due to the increase in nutrition value, especially proteins and fibers, the by-product can be re-integrated into the food chain. Moreover, the results of the study can have a positive influence on protein supply worldwide. Therefore, this study aligns with the SDGs #2 ‘Zero hunger’, #3 ‘Good health and well-being’, and #12 ‘Responsible consumption and production’. |
1 Introduction
Besides the challenge to feed a growing population including the supply of proteins, the world is facing a huge problem of food waste. One of the important sources of food waste is the residuals from the agro-food industry, also known as by-products.1 In the EU, the amount of food waste resulting from by-product accumulation accounts for 19% of the total food that leaves the food supply chain.2 Huge proportions of these by-products are still underutilized as feed ingredients, for fertilizer production, or even landfilled.3Carob pulp, also known as carob kibbles, is an example of such an industrial by-product accruing after the production of locust bean gum (E410), a stabilizer applied in food formulations, which is obtained from carob fruits. So far, the carob pulp is mostly used for animal feed.4,5
Carob fruit (Ceratonia siliqua L.), also known as the locust bean, belongs to the leguminous plant family and is commonly cultivated in the Mediterranean region.6,7 The fruits consist of 90% pulp and 10% seeds, the latter is the raw material of locust bean gum.8,9 Remaining carob pulp could be a reliable source of dietary fibers, sugars, and a range of bioactive compounds such as polyphenols and D-pinitol.10 Therefore, carob pulp could be an interesting component for a wide variety of health-beneficial food products.4,11 Despite several attempts towards its utilization as a food product,4,12 such applications have remained very limited. One of the main reasons for the under-utilization is the low protein content of carob pulp.7,12,13 Because protein is the most important and often limited macronutrient, there is a need to increase the protein content before usage as a food ingredient.
During the last years, fermentation as a biotechnological process has gained rising popularity as a highly effective technique for processing food and upgrading by-products. It has distinct advantages, such as cost-effectiveness, eco-friendliness, and mild reaction conditions, setting it apart from chemical and physical methods to process foods. It can also be applied to add value to by-products from food manufacturing such as nutritional enrichment of crops or crop residues by biotransformation.14 In particular, solid-state fermentation (SSF) could serve as an effective bioprocess for the production of plant-based ingredients exhibiting enhanced nutritional and functional properties.15,16
Various microorganisms are used for SSF, notably those particular species, which are long recognized for their safety in the biological realm. One of them is Pleurotus ostreatus, a white-rot fungus capable of growing on different agri-food wastes. It is able to produce enzymes that can degrade the lignocellulosic structure of the substrate and convert low-cost industrial by-products into value-added food items.17 Furthermore, some studies indicated that this fungus variety can be used to increase protein content in the substrates.16,18
However, the usage of Pleurotus ostreatus in an SSF of the by-product carob pulp to increase its value as a nutritional food component has not been investigated, so far. Previously, carob has been investigated as a substrate for solid-state fermentation to observe the changes in nutritional value using various microfungi such as Monascus ruber,19Rhizopus oligosporus,19,20Aspergillus niger,21 as well as yeasts such as Saccharomyces cerevisiae and Saccharomyces rouxii.20 The latter ones proved to be able to increase the protein content. However, all these fermentations benefited from the relatively high sucrose content in carob. On the other hand, Pleurotus ostreatus is a macrofungi species, that possesses lignolytic activities allowing it to grow directly on lignocellulosic material, such as carob pulp with low sucrose content. Therefore, this study investigates the impact of SSF with Pleurotus ostreatus on carob pulp where the easily accessible sucrose was extracted before fermentation. The changes in some nutritionally relevant parameters of the substrate such as contents of protein and reducing sugars as well as the antioxidative properties of carob pulp were evaluated.
2 Experimental
2.1 Materials
Carob fruits (Ceratonia siliqua L.) were acquired from the local market of Hannover, Germany. The Pleurotus ostreatus oyster mushroom strain MG 1005 and malt extract agar Petri plates (90 × 15 mm) were purchased from MycoGenetics (Everswinkel, Germany). The assay kit ‘Available Carbohydrates and Dietary Fiber’ (K-ACHDF 06/18, lot number 220303-3), later on, called Megazyme kit, which was used for the determination of contents in dietary fibers, glucose and fructose, was supplied by Megazyme International Ltd (Wicklow, Ireland). All the reagents used were of analytical grade.2.2 Solid-state fermentation (SSF)
The carob fruits were first washed to eliminate adherent dust and stored overnight at 30 °C in a hot air oven (Heratherm OGS100, Thermo Electron GmbH, Langenselbold, Germany) to remove the surface water. Afterwards, carob fruits were manually broken into kibble shapes using pestle and mortar and the seeds were removed manually. The resulting size of the carob kibbles was between 18 to 22 mm.Agar plates were inoculated with a Pleurotus ostreatus inoculum syringe using a 2 mL inoculum/Petri plate and incubated at 30 °C for 10 days. These plates were used as the starter culture for the SSF.
Kibbles were subjected to fungal SSF using the following method. 20 g of the material was soaked in 80 mL of distilled water overnight in conical flasks (100 mL) to reduce the sucrose. The following day, water was removed manually and the soaked kibbles were washed two times using 80 mL of distilled water at 100 °C. Afterwards, the amount of water in the samples was adjusted to 45% (w/w) related to the initial mass of kibbles (20 g) and the samples were thermally treated at 121 °C for 30 min (iCombi Classic LM200BE, Rational AG, Landsberg, Germany). Subsequently, carob kibbles in the flasks were inoculated with one portion of the starter culture containing 0.05 g of mycelium in a sterilized laminar bench (SAVVY, Lamsystems GmbH, Wustermark, Germany). Finally, the flasks with the kibbles were incubated at 30 °C for different time intervals. Samples were analyzed at 0 (before fermentation), 5, 15, 20, 25, and 30 days of fermentation. For each sample, the entire flask content was dried at 55 °C overnight in the hot air oven, ground manually using pestle mortar, and then sealed in zipper bags. Dried samples were stored in a fridge until further analyses. These drying conditions were sufficient to increase the dry matter content to up to 99% as found in the preliminary trials (data not shown).
2.3 Chemical analyses
As mentioned above, because of the very low residual moisture of <1%, all results of the chemical analyses presented later in the tables were related to the dry matter of the carob pulp.Duplicate samples weighing 1 g each combined with 40 mL of MES-TRIS buffer solution were added in Duran bottles (250 mL) and stirred thoroughly. The samples were firstly incubated with 50 μL of α-amylase (bottle 3 of the kit) for 35 min at 80 °C and subsequently incubated with 100 μL of protease (bottle 4 of the kit) at 60 °C for 30 min. For pH adjustment after protease treatment, 5 mL of 3 M acetic acid solution was added. The last treatment involved incubation with 200 μL amyloglucosidase (bottle 5 of the kit) at 60 °C for 30 min.
Following these enzymatic reactions, 1 mL of each sample solution was removed for the determination of the carbohydrate content and transferred to a 50 mL volumetric flask. The volume was then adjusted to 50 mL with 100 mM sodium maleate buffer. The remaining samples in the Duran bottles underwent filtration through a 50 mL Celite-in-bed crucible with porosity 2 (DWK Life Sciences, Mainz, Germany). Four volumes of 95% v/v ethanol with a temperature of 60 °C were added to the filtrate to precipitate the SDF. The mix was filtered after 1 h and both residues (IDF as filter residue and SDF as filtrate) were washed with 78% v/v ethanol, 95% v/v ethanol, and acetone, and then dried at 103 °C overnight in the same hot air oven as mentioned above. After drying, the fractions were weighed. Protein and ash contents of the dried fractions were determined as described above. Dietary fiber content was calculated from the weight after drying and corrected with respect to its protein and ash content. The total dietary fiber content (TDF) was calculated as the sum of IDF and SDF.
After incubation, 2.3 mL of distilled water was added to the blank cuvette and 2 mL was added to the sample cuvette. Subsequently, 0.1 mL buffer (bottle 1 of the kit) and 0.1 mL NADP+ and ATP solution (bottle 2 of the kit) were added to both cuvettes. The mixtures were thoroughly mixed and the absorbance (A1) at 340 nm was measured after 3 min using the Spectrometer SPECORD 205 (Analytik Jena AG, Jena, Germany). Afterwards, 0.02 mL of hexokinase and glucose-6-phosphate dehydrogenase (bottle 7 of the kit) were added and mixed with the other content. The absorbance (A2) at 340 nm was read after 5 min. Finally, 0.02 mL of phosphoglucose isomerase solution (bottle 8 of the kit) was added and the absorbance (A3) at 340 nm was recorded after 9 min.
The concentration C in g L−1 of D-glucose and D-fructose was calculated as follows:
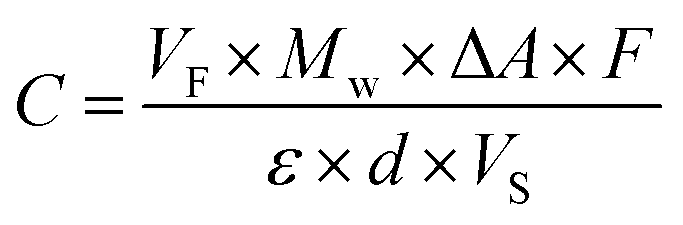
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :80:10, v/v/v) was added. This mixture was held at 24 °C for 2.5 h in a shaker water bath (H 20 S, Lauda, Königshofen, Germany). Afterwards, samples were centrifuged at 1448×g for 10 min.
:80:10, v/v/v) was added. This mixture was held at 24 °C for 2.5 h in a shaker water bath (H 20 S, Lauda, Königshofen, Germany). Afterwards, samples were centrifuged at 1448×g for 10 min.
The total polyphenolic content of carob samples was determined using the Folin–Ciocalteu method.25 Initially, 0.1 mL extract was mixed with 0.5 mL of Folin–Ciocalteu reagent (10%, v/v, in water) and 1.5 mL of sodium carbonate solution (20%, m/v, in water) in a test tube of 15 mL volume. Then, the mixture was incubated for 2 h in the darkness at 24 ± 1 °C and the absorbance was measured using a spectrophotometer (SPECORD® 205 Analytik Jena, Germany) at a wavelength of 760 nm after pouring the mixture into 1.6 mL cuvettes. The extraction solution was used as the blank. Methanolic solutions of gallic acid concentrations in the range of 5 to 800 ppm were used to establish the calibration curve for quantification. Results are reported as mg gallic acid equivalents (GAE) per g of sample related to dry matter (DM).
Antioxidant properties of the samples were determined as DPPH (2,2-diphenyl-1-picrylhydrazyl) radical scavenging activity according to the method proposed by Tontul and Topuz (2017).26 An amount of 50 μL extract together with 950 μL of freshly prepared DPPH solution (60 μM in 80% methanol) was added to the 1.6 mL cuvettes and mixed thoroughly. The mixture was incubated at room temperature in the dark for 30 min and absorbance was recorded against blank (50 μL of the extraction solution instead of sample) at 516 nm. Results were reported as Trolox equivalents (TE) in mg per g of sample related to DM using a calibration curve, where methanolic solutions of Trolox with different concentrations between 2 and 64 ppm were applied.
3 Results and discussion
3.1 Protein and ash contents
The growth of Pleurotus ostreatus and the development of fungal biomass on the de-sugared carob kibbles during fermentation can be observed from the photo in Fig. 1 taken after 30 days of fermentation. | ||
| Fig. 1 Photo of the carob kibbles after 30 days of SSF in the Erlenmeyer flask. | ||
The protein and ash contents of the carob kibbles before treatment (control), after thermal treatment (before fermentation) and during the SSF are summarized in Table 1. All contents are related to DM as mentioned in Chapter 2.3 and the results are presented as mean values ± standard deviations.
| Sample | Contents in % w/w DM* | |
|---|---|---|
| Protein | Ash | |
| a *DM dry matter, ** results are expressed as mean value ± standard deviation, different lowercases indicate significantly different protein or ash contents, respectively, with p < 0.05. | ||
| Control (kibbles) | 3.54 ± 0.06g* | 3.50 ± 0.14b |
| After thermal treatment | 4.64 ± 0.12f | 3.38 ± 0.01b |
| After 5 days SSF | 6.18 ± 0.06e | 1.70 ± 0.14c |
| After 10 days SSF | 6.96 ± 0.06d | 3.30 ± 0.14b |
| After 15 days SSF | 8.10 ± 0.06c | 3.30 ± 0.14b |
| After 20 days SSF04 | 9.50 ± 0.06b | 3.40 ± 0.01b |
| After 25 days SSF | 9.68 ± 0.06b | 4.08 ± 0.14a |
| After 30 days SSF | 12.39 ± 0.31a | 4.38 ± 0.02a |
The initial protein content in carob powder related to DM was measured at 3.54 ± 0.06%, which is in line with the findings of previous studies. For example, Avallone et al. (1997)25 reported a protein content between 1 and 5% in eight different varieties of carob collected from Italy. An elevation of approximately 30% in the protein content was observed in the sample after the thermal treatment. Similar results were found in a study, where thermal treatment increased the total protein concentration of extracts from black beans by 62.5% after autoclaving at 100 °C for 30 min and by 78.5% after autoclaving two times at 121 °C for 15 min.27 Exposure to higher temperatures can induce denaturation, aggregation, cross-linking, and changes in hydration that can lower protein solubility.28 Therefore, the increase in protein concentration might be attributed to a washout of other more water-soluble substances related to protein, which results in an apparent increase in protein content.
During the following 30 days of SSF, an increase in the protein content of by 250% compared to the initial amount was observed. This can be attributed to an increase in the fungal protein due to the growth and fungal metabolic activity during SSF. With respect to the influence of SSF on protein content in other substrates, Imelda et al. (2008)29 documented a notable augmentation in protein content of 49% in wheat bran after 8 days of SSF using Aspergillus niger. The rise in the protein content observed during fermentation could be attributed to the effective conversion of complex carbohydrates into microbial protein and the generation of various proteinaceous enzymes.29,30
The ash content in the initial carob kibbles (control) was 3.5%, which is consistent with the results reported by Youssef et al. (2013)31 who found 3.0% of ash in carob powder purchased from Aswan Governorate, Egypt. With respect to the effect of SSF, a decrease in ash content was detected at the beginning of SSF (until the 5th day). Later on, ash content slightly increased up to the end of SSF. However, no significant differences were found between the 10th and 20th days of SSF. A slight but significant increase could be detected between days 20 and 25 with no further significant change until day 30. A comparable increase from 2.12% to 3.73% in ash content was also reported by Terefe et al. (2021)32 in maize flour after 48 h of fermentation using L. plantarum and S. cerevisiae co-cultures. Similar to our study, a slight decrease in the ash content was observed during the first fourth of their study. This initial reduction during the start of SSF was attributed to a consumption of minerals (ash) for the metabolic activities of the microorganisms during SSF.33 Imelda et al. (2008)29 suggested that the loss in dry matter during SSF eventually increases the unaltered component such as the ash content. Terefe et al. (2021)32 discussed a loss of organic matter and accumulation of inorganic matter as possible reasons for the increase in the ash content.
3.2 Dietary fiber contents
Table 2 presents the amount of dietary fibers in the carob kibbles before SSF and during the fermentation as mean values ± standard deviations.| Sample | Contents in % w/w DM* | ||
|---|---|---|---|
| IDF* | SDF* | TDF* | |
| a *DM dry matter, IDF insoluble dietary fiber, SDF soluble dietary fiber, TDF total dietary fiber, ** results are expressed as mean value ± standard deviation, different lowercases indicate significantly different protein or ash contents, respectively, with p < 0.05. | |||
| Control (kibbles) | 29.46 ± 1.67f** | 2.70 ± 0.71b | 32.16 ± 2.38e |
| After thermal treatment | 40.04 ± 0.53e | 6.80 ± 1.56a | 46.84 ± 2.08d |
| After 5 days SSF | 58.09 ± 0.43d | 7.60 ± 0.42a | 65.69 ± 0.85c |
| After 10 days SSF | 61.16 ± 0.12d | 7.95 ± 1.20a | 69.12 ± 1.33c |
| After 15 days SSF | 61.77 ± 1.23d | 7.25 ± 0.07a | 69.02 ± 1.16c |
| After 20 days SSF04 | 66.48 ± 0.52c | 6.95 ± 0.92a | 73.43 ± 0.40b |
| After 25 days SSF | 69.08 ± 0.53b | 6.85 ± 0.92a | 75.92 ± 1.45a,b |
| After 30 days SSF | 72.34 ± 0.52a | 6.30 ± 0.42a | 78.64 ± 0.94a |
Initial dietary fiber concentrations were 29.46 ± 1.67%, 2.70 ± 0.71%, and 32.16 ± 2.38% for IDF, SDF and TDF, respectively. Comparable dietary fiber concentrations in carob pulp were reported by Ikram et al. (2023)34 with 27–50% of TDF and by Rodriguez-Solana et al. (2021)35 with concentrations between 9.7 and 37.3%.
Concentrations of dietary fiber increased during the SSF as could be observed from Table 2. An increase of up to 145% in the TDF content and up to 245% in SDF related to the initial contents in the untreated kibbles was measured. TDF concentrations continuously increased during fermentation except during the period between days 10 and 15. The increase in TDF content in the carob kibbles during fermentation is primarily attributed to the rise in IDF content, as a significant increase was not observable for the SDF during the fermentation period. Whereas a significant increase in the SDF content (152%) was determined after the thermal treatment of the carob kibbles.
Thermal treatment of potatoes at 130 °C for 100 min resulted in an increase in TDF concentration from 6.8% to 10.3%.36 Comparable results were observed for the treatment of wheat and rice bran, where IDF and TDF increased after autoclaving at pH values between 3.5 and 4.5 and different holding times between 0.5 and 1.5 h.37 With respect to the SDF concentrations, these authors measured an increase in the rice bran samples, but a decrease in the wheat bran samples.
The changes in SDF concentrations due to thermal treatment can be due to different reasons. Such as the processing can break glycosidic bonds within the complex carbohydrates forming the IDF. This effect would increase their solubility resulting in a higher content of SDF. On the other hand, further breakage of the carbohydrate chains might lead to smaller molecules, which are no longer considered fibers resulting in a decrease in SDF.38
Considering the influence of fermentation, an increase in crude fiber content from 0.82% in control to 2.56% after 48 h of the fermentation was observed in soybean meal using Bacillus coagulans.29 This phenomenon was attributed to the utilization of the soluble and readily digestible carbohydrates by the growing mycelium, resulting in a high level of apparently indigestible fiber content. The IDF and TDF contents in beans Phaseolus vulgaris were enhanced after natural fermentation with naturally present microorganisms on the seeds for 48 h as well as after a lactic acid fermentation using Lactobacillus plantarum.39 On the other hand, SDF contents decreased in both types of fermentation. The authors suggested that the lower SDF contents could be attributed to degradation of SDF into fragments with lower molecular weights during the fermentation process. Therefore, a portion of SDF might not be fully recovered by ethanolic precipitation for the fiber analysis, resulting in an apparent decrease in the fiber contents measured.
3.3 D-Glucose and D-fructose contents
Changes in the contents of D-glucose and D-fructose in the carob kibbles during fermentation are summarized in Table 3 presented as mean values ± standard deviations.| Sample | Contents in % w/w DM* | |
|---|---|---|
| D-Glucose | D-Fructose | |
| a *DM dry matter, ** results are expressed as mean value ± standard deviation, different lowercases indicate significantly different protein or ash contents, respectively, with p < 0.05. | ||
| Control (kibbles) | 5.79 ± 0.31b** | 4.52 ± 0.31a |
| After thermal treatment | 3.25 ± 0.33c | 4.54 ± 0.64a |
| After 5 days SSF | 8.97 ± 1.59a | 4.05 ± 0.98a,b |
| After 10 days SSF | 5.78 ± 1.65a,b,c | 2.65 ± 0.40b |
| After 15 days SSF | 3.19 ± 1.12d | 1.76 ± 0.50b,c |
| After 20 days SSF04 | 3.07 ± 0.90d | 0.98 ± 0.41c |
| After 25 days SSF | 1.28 ± 0.21e | 0.55 ± 0.08c |
| After 30 days SSF | 1.70 ± 0.37e | 0.69 ± 0.21c |
D-Glucose content was boosted in the initial stage of SSF following a decline after the thermal treatment. It peaked on the 5th day of fermentation. Subsequently, a continuous decline was observed until the 25th day of fermentation and a slight and non-significant increase at day 30 of SSF. A different trend was observed for the D-fructose content, where only a slight, but non-significant, increase was observed during the first 5 days of fermentation. Then, it continued to decrease until the 25th day and became constant afterwords. The decrease in the D-glucose content after thermal treatment can be attributed to an extraction of the water-soluble monosaccharides during the soaking time (washout effect). In the case of D-fructose, this drop due to soaking was not found. This might indicate that the D-fructose was less accessible for the water in the untreated carob matrix compared to D-glucose. The contents of reducing sugars in the untreated carob pulp as measured in this study are comparable to data published by Khlifa et al. (2013)40 who reported values between 2.2% and 7.8%.
With respect to the effect of fermentation, a trend comparable to our results was observed in the fermentation of willow herb leaves.41 The authors found an increase in D-glucose and D-fructose content in the first 24 h compared to the non-fermented samples, but a remarkable decrease was observed after 48 and 72 h of aerobic fermentation. The increase of the monosaccharide contents at the beginning of SSF might be related to the hydrolysis of residuals of the disaccharide sucrose into monosaccharides.41 Later on, the molecules were consumed within the metabolic activity of the fungi, which reduced the detectable amount of these components. However, such an increase in content at the beginning of SSF was not observable for D-fructose, which might be related to faster metabolism of this monosaccharide by the fungi from the beginning.
3.4 Phenolic content and antioxidant activity of the carob kibbles
Table 4 shows the phenolic contents in the kibbles and their antioxidant activity measured via the DPPH test as mean values ± standard deviations.| Sample | TPC* (mg GAE* g−1 DM*) | DPPH* (mg TE* g−1 DM) |
|---|---|---|
| a *TPC total phenolic content, GAE gallic acid equivalents, DM dry matter, DPPH 2,2-diphenyl-1-picrylhydrazyl, TE trolox equivalents, ** results are expressed as mean value ± standard deviation, different lowercases indicate significantly different protein or ash contents, respectively, with p < 0.05. | ||
| Control (kibbles) | 3.87 ± 0.12a** | 7.82 ± 0.75a |
| After thermal treatment | 3.81 ± 0.10a | 6.29 ± 0.09a |
| After 5 days SSF | 3.73 ± 0.03a | 3.15 ± 0.08b |
| After 10 days SSF | 3.47 ± 0.12b | 2.80 ± 0.04c |
| After 15 days SSF | 3.01 ± 0.07c | 1.46 ± 0.32d |
| After 20 days SSF04 | 2.80 ± 0.10d | 1.32 ± 0.72b,c,d,e |
| After 25 days SSF | 1.95 ± 0.04e | 0.73 ± 0.13d,e |
| After 30 days SSF | 1.77 ± 0.02f | 0.45 ± 0.04e |
The initial concentration of the phenolic compounds was 3.87 ± 0.12 mg GAE g−1, whereas the concentration decreased to 1.77 ± 0.02 mg GAE g−1 after 30 days of fermentation representing approximately only 46% of the initial value. Processing of the carob kibbles before SSF including thermal treatment seemed to have only negligible effect on TPC because no statistically significant difference (p > 0.05) was observed between initial and thermally treated samples.
Other studies found a significant decrease in TPC during thermal treatment. Sánchez-García et al. (2022)16 reported a significant reduction in TPC after autoclaving lentil grains and flour before subjecting to SSF with Pleurotus ostreatus. Babar et al. (1988)42 found an even higher reduction in polyphenol contents by about 91% and 70% in meal and seeds of jack bean, respectively, after moist heat treatment at 120 °C for 20 min. The reduction in polyphenols after such a thermal treatment was attributed to the interaction of polyphenols with other components of seeds, such as protein, to form insoluble tannin–protein complexes, which are not accessible for analysis.42 This effect might be less pronounced in the carob matrix compared to the other materials because of low protein contents resulting in less distinct changes. With respect to the influence of SSF itself, a decrease in TPC after 14 days was measured in the study mentioned above.16 In another study performed by Xu et al. (2018),43 TPC in sorghum decreased with increasing fermentation time of up to 35 days, where different strains of edible fungi were applied. However, an increase in TPC for other grains under the same conditions was measured, too.43
A similar decreasing effect as for TPC was observed for the DPPH activity in our study. The DPPH activity decreased significantly from 7.82 ± 0.75 mg TE g−1 in the initial state to 0.45 ± 0.04 mg TE g−1 after 30 days of SSF representing a reduction of approximately 94%. Thermal treatment of the samples before SSF resulted in a decrease in DPPH activity compared to the initial state of the carob kibbles. Similar changes in antioxidative activity measured with the ORAC test were found by Sanchiz et al. (2019)44 for pistachio, cashew, and chestnut samples after a heat treatment at 121 °C for 30 min.
Likewise, Sánchez-García et al. (2022)16 reported a similar pattern of DPPH changes in lentil grains (61.6% reduction), lentil flour (61.5%), quinoa seeds (17% reduction) and quinoa flour (23.1% reduction) after 14 days of SSF using Pleurotus ostreatus. This phenomenon might be a result of competitive reactions promoting the formation of either pro-oxidant or antioxidant compounds during SSF. The outcome might be either a decrease or increase in the food's total antioxidant capacity, which is measured by the DPPH test.16
4 Conclusions
The study shows that solid-state fermentation of carob kibbles, a by-product of locust bean gum manufacturing from carob fruits, by using the edible fungal strain of Pleurotus ostreatus can contribute to an improvement of carob valorization by means of significantly increasing its protein content. Thus, especially the SSF with Pleurotus ostreatus for purposes of protein generation from low-value lignocellulosic by-products of food manufacturing shows potential. Since in the case of carob, this study recorded a threefold increase in its protein content. Evidently, these results are also interesting for the circular bioeconomy in food production, which needs to be put into practice in future. Application of the fermented substrate in different food matrices could be the next step in the research activities. In this respect, it should be also considered that this type of fermentation leads to a decrease in the antioxidant capacity of the carob kibbles. Here, more research is required to understand the mechanisms behind this decrease and for possible strategies to overcome this effect. In the meantime, the addition of the antioxidative agents in the final recipes, where fermented carob products are used, could be considered.Data availability
All relevant data for this study is included in the article.Author contributions
Shahab Iqbal developed the methodology, analyzed and validated the data, and wrote the manuscript. Özge Tirpanalan-Staben developed the idea of the study, supervised the methodology and edited the manuscript. Knut Franke was involved in the acquisition of funding and project administration and wrote and edited the manuscript.Conflicts of interest
The authors declare that there are no conflicts of interest.Acknowledgements
The stay of Shahab Iqbal at the Institute of Food and One Health was founded by the German Academic Exchange Service (DAAD) with grant number 91799387.References
- European Commission, Communication from the Commission to the Council and the European Parliament on the Interpretative Communication on Waste and By-Products, European Commission, Brussels, 2007 Search PubMed.
- I. Mateos-Aparicio, A. Matias, C. M. Galanakis and C. M. Galanakis, in The Role of Alternative and Innovative Food Ingredients and Products in Consumer Wellness, Academic Press, New York, San Francisco, London, 2020, pp. 239–281 Search PubMed.
- A. Schieber, F. C. Stintzing and R. Carle, By-products of plant food processing as a source of functional compounds – recent developments, Trends Food Sci. Technol., 2001, 12, 401–413 CrossRef CAS.
- M. E. Brassesco, T. R. S. Brandão, C. L. M. Silva and M. Pintado, Carob bean (Ceratonia siliqua L.): a new perspective for functional food, Trends Food Sci. Technol., 2021, 114, 310–322 CrossRef CAS.
- S. Barak and D. Mudgil, Locust bean gum: processing, properties and food applications – a review, Int. J. Biol. Macromol., 2014, 66, 74–80 CrossRef CAS PubMed.
- H. Bahry, A. Pons, R. Abdallah, G. Pierre, C. Delattre, N. Fayad, S. Taha and C. Vial, Valorization of carob waste: definition of a second-generation bioethanol production process, Bioresour. Technol., 2017, 235, 25–34 CrossRef CAS PubMed.
- I. Turhan, Optimization of extraction of D-pinitol and phenolics from cultivated and wild types of carob pods using response surface methodology, Int. J. Food Eng., 2011, 7, 7 Search PubMed.
- H. Bahry, R. Abdalla, A. Pons, S. Taha and C. Vial, Optimization of lactic acid production using immobilized Lactobacillus rhamnosus and carob pod waste from the Lebanese food industry, J. Biotechnol., 2019, 306, 81–88 CrossRef CAS PubMed.
- I. Turhan, N. Tetik, M. Aksu, M. Karhan and M. Certel, Liquid-solid extraction of soluble solids and total phenolic compounds of carob bean (Ceratonia siliqua L.), J. Food Process Eng., 2006, 29, 498–507 CrossRef.
- H. El Batal, A. Hasib, F. Dehbi, N. Zaki, A. Ouatmane and A. Boulli, Assessment of nutritional composition of carob pulp (Ceratonia Siliqua L.) collected from various locations in Morocco, J. Mater. Environ. Sci., 2016, 7, 3278–3285 CAS.
- F. A. Ayaz, H. Torun, R. H. Glew, Z. D. Bak, L. T. Chuang, J. M. Presley and R. Andrews, Nutrient content of carob pod (Ceratonia siliqua L.) flour prepared commercially and domestically, Plant Foods Hum. Nutr., 2009, 64, 286–292 CrossRef CAS PubMed.
- A. Loullis and E. Pinakoulaki, Carob as cocoa substitute: a review on composition, health benefits and food applications, Eur. Food Res. Technol., 2018, 244, 959–977 CrossRef CAS.
- Y. Ercan, T. Irfan and K. Mustafa, Optimization of ethanol production from carob pod extract using immobilized Saccharomyces cerevisiae cells in a stirred tank bioreactor, Bioresour. Technol., 2013, 135, 365–371 CrossRef CAS PubMed.
- K. S. M. S. Raghavarao, T. V. Ranganathan and N. G. Karanth, Some engineering aspects of solid-state fermentation, Biochem. Eng. J., 2003, 13, 127–135 CrossRef CAS.
- C. Dai, Y. Hou, H. Xu, L. Huang, M. Dabbour, B. K. Mintah, R. He and H. Ma, Effect of solid-state fermentation by three different Bacillus species on composition and protein structure of soybean meal, J. Sci. Food Agric., 2022, 102, 557–566 CrossRef CAS PubMed.
- J. Sánchez-García, A. Asensio-Grau, J. García-Hernández, A. Heredia and A. Andrés, Nutritional and antioxidant changes in lentils and quinoa through fungal solid-state fermentation with Pleurotus ostreatus, Bioresour. Bioprocess., 2022, 9, 51 CrossRef PubMed.
- M. Ritota and P. Manzi, Pleurotus spp. cultivation on different agri-food by-products: example of biotechnological application, Sustainability, 2019, 11, 5049 CrossRef CAS.
- A. Asensio-Grau, J. Calvo-Lerma, A. Heredia and A. Andrés, Enhancing the nutritional profile and digestibility of lentil flour by solid state fermentation with Pleurotus ostreatus, Food Funct., 2020, 11, 7905–7912 RSC.
- R. Kokke, Improvement of carob pods as feed by solid-substrate fermentation, J. Appl. Bacteriol., 1977, 43, 303–307 CrossRef CAS.
- S. Marakis, M. Lambraki, G. Marakis, S. Roussos, S. Roussos, B. K. Lonsane, M. Raimbault and G. Viniegra-Gonzalez, Advances in Solid State Fermentation, Springer, Dordrecht, 1997, pp. 235–244 Search PubMed.
- T. Smail, O. Salhi and J. S. Knapp, Solid-state fermentation of carob pods by Aspergillus niger for protein production: effect of particle size, World J. Microbiol. Biotechnol., 1995, 11, 171–173 CrossRef CAS PubMed.
- AOAC, Official Methods of Analysis of AOAC International, AOAC International, Gaithersburg, 22nd edn, 2023 Search PubMed.
- L. Gao, S. Wang, B. D. Oomah and G. Mazza, Wheat Quality Elucidation, the Bushuk Legacy, ed. P. K. W. Ng and C. W. Wrigley, AACC International, St. Paul, 2002, pp. 219–233 Search PubMed.
- T. Beta, S. Nam, J. E. Dexter and H. D. Sapirstein, Phenolic content and antioxidant activity of pearled wheat and roller-milled fractions, Cereal Chem., 2005, 82, 390–393 CrossRef CAS.
- R. Avallone, M. Plessi, M. Baraldi and A. Monzani, Determination of chemical composition of carob (Ceratonia siliqua): protein, fat, carbohydrates, and tannins, J. Food Compos. Anal., 1997, 10, 166–172 CrossRef CAS.
- I. Tontul and A. Topuz, Effects of different drying methods on the physicochemical properties of pomegranate leather (pestil), LWT--Food Sci. Technol., 2017, 80, 294–303 CrossRef CAS.
- A. Escobedo, G. Loarca-Piña, M. Gaytan-Martínez, I. Orozco-Avila and L. Mojica, Autoclaving and extrusion improve the functional properties and chemical composition of black bean carbohydrate extracts, J. Food Sci., 2020, 85, 2783–2791 CrossRef CAS PubMed.
- S. M. Beck, K. Knoerzer, J. Sellahewa, M. A. Emin and J. Arcot, Effect of different heat-treatment times and applied shear on secondary structure, molecular weight distribution, solubility and rheological properties of pea protein isolate as investigated by capillary rheometry, J. Food Eng., 2017, 208, 66–76 CrossRef CAS.
- J. Imelda, P. R. Raj and D. Bhatnagar, Effect of solid state fermentation on nutrient composition of selected feed, Indian J. Fish., 2008, 55, 327–332 Search PubMed.
- O. O. Olukomaiya, O. Q. Adiamo, W. C. Fernando, R. Mereddy, X. Li and Y. Sultanbawa, Effect of solid-state fermentation on proximate composition, anti-nutritional factor, microbiological and functional properties of lupin flour, Food Chem., 2020, 315, 126238 CrossRef CAS PubMed.
- M. K. E. Youssef, M. M. El-Manfaloty and H. M. Ali, Assessment of proximate chemical composition, nutritional status, fatty acid composition and phenolic compounds of carob (Ceratonia siliqua L.), Food Publ. Health, 2013, 3, 304–308 Search PubMed.
- Z. K. Terefe, M. N. Omwamba and J. M. Nduko, Effect of solid state fermentation on proximate composition, antinutritional factors and in vitro protein digestibility of maize flour, Food Sci. Nutr., 2021, 9, 6343–6352 CrossRef CAS PubMed.
- N. M. Nnam, Evaluation of nutritional quality of fermented cowpea (Vigna unguiculata) flours, Ecol. Food Nutr., 1995, 33, 273–279 CrossRef.
- A. Ikram, W. Khalid, K. Wajeeha Zafar, A. Ali, M. F. Afzal, A. Aziz, I. Faiz ul Rasool, A. Al-Farga, F. Aqlan and H. Koraqi, Nutritional, biochemical, and clinical applications of carob: a review, Food Sci. Nutr., 2023, 11, 3641–3654 CrossRef CAS PubMed.
- R. Rodríguez-Solana, A. Romano and J. M. Moreno-Rojas, Carob pulp: a nutritional and functional by-product worldwide spread in the formulation of different food products and beverages. A review, Processes, 2021, 9, 1146 CrossRef.
- P. Varo, K. Veijalainen and P. Koivistoinen, Effect of heat treatment on the dietary fibre contents of potato and tomato, Int. J. Food Sci. Technol., 1984, 19, 485–492 CrossRef CAS.
- B. Özkaya, S. Turksoy, H. Özkaya and B. Duman, Dephytinization of wheat and rice brans by hydrothermal autoclaving process and the evaluation of consequences for dietary fiber content, antioxidant activity and phenolics, Innovative Food Sci. Emerging Technol., 2017, 39, 209–215 CrossRef.
- S. Iqbal, Ö. Tirpanalan-Staben and K. Franke, Modification of dietary fibers to valorize the by-products of cereal, fruit and vegetable industry – a review on treatment methods, Plants, 2022, 11, 3466 CrossRef CAS PubMed.
- M. A. Martín-Cabrejas, B. Sanfiz, A. Vidal, E. Mollá, R. Esteban and F. J. López-Andréu, Effect of fermentation and autoclaving on dietary fiber fractions and antinutritional factors of beans (Phaseolus vulgaris L.), J. Agric. Food Chem., 2004, 52, 261–266 CrossRef PubMed.
- M. Khlifa, A. Bahloul and S. Kitane, Determination of chemical composition of carob pod (Ceratonia siliqua L.) and its morphological study, J. Mater. Environ. Sci., 2013, 4, 348–353 CAS.
- N. Vaitkeviciene, E. Jariene, J. Kulaitiene, M. Lasinskas, A. Blinstrubiene and E. Hallmann, Effect of solid-state fermentation on vitamin C, photosynthetic pigments and sugars in willow herb (Chamerion angustifolium (L.) Holub.) leaves, Plants, 2022, 11, 3300 CrossRef CAS PubMed.
- V. S. Babar, J. K. Chavan and S. S. Kadam, Effects of heat treatments and germination on trypsin inhibitor activity and polyphenols in jack bean (Canavalia ensiformis L. DC), Plant Foods Hum. Nutr., 1988, 38, 319–324 CrossRef CAS PubMed.
- L.-N. Xu, S. Guo and S. Zhang, Effects of solid-state fermentation with three higher fungi on the total phenol contents and antioxidant properties of diverse cereal grains, FEMS Microbiol. Lett., 2018, 365, fny163 CAS.
- A. Sanchiz, M. M. Pedrosa, E. Guillamón, C. Arribas, B. Cabellos, R. Linacero and C. Cuadrado, Influence of boiling and autoclave processing on the phenolic content, antioxidant activity and functional properties of pistachio, cashew and chestnut flours, LWT--Food Sci. Technol., 2019, 105, 250–256 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2024 |