
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceBioaugmentation with an aerobic denitrifying bacterium with quorum quenching activity for improved nitrogen removal and reduced membrane fouling in anoxic/oxic membrane bioreactor†
Pengyuan Weia,
Jun Li *a,
Jing Zhang*b,
Yi Zhanga,
Rong Luoa,
Chaofan Xiea and
Zhaozhao Wangc
*a,
Jing Zhang*b,
Yi Zhanga,
Rong Luoa,
Chaofan Xiea and
Zhaozhao Wangc
aNational Engineering Laboratory for Advanced Municipal Wastewater Treatment and Reuse Technology, Engineering Research Center of Beijing, Beijing University of Technology, Beijing 100124, PR China. E-mail: ljbjut@126.com
bBeijing City Drainage Refco Group Ltd, Beijing 100124, PR China
cThe College of Energy and Environmental Engineering, Hebei University of Engineering, Handan 056038, PR China
First published on 16th January 2023
Abstract
In this study, an aerobic denitrifying bacterium with quorum quenching activity, Acinetobacter sp. WZL728, was inoculated into the anoxic/oxic membrane bioreactor (A/O-MBR) to study its effects on A/O-MBR performance. The pollutant removal and membrane fouling between A/O-MBR with WZL728 (EMBR) and A/O-MBR without WZL728 (CMBR) were compared. WZL728 increased the total nitrogen removal efficiency from 75.05% in CMBR to 91.03% in EMBR and extended the filtration cycle from 5.44 days in CMBR to 9.57 days in EMBR, which indicated that WZL728 improved the pollutant removal performance and mitigated membrane fouling of A/O-MBR. The concentration of N-acyl-homoserine lactones in the biocake of EMBR (EMBRB) was 11.23% of that in the biocake of CMBR (CMBRB). The content of extracellular polymeric substances (EPS) in EMBRB was 69.00% of that in CMBRB. The abundance of bacteria associated with EPS secretion (Alpthaproteobacteria) decreased and the abundance of bacteria associated with EPS degradation (Clostridia) increased in EMBRB. Valine, alanine and uridine diphosphate-N-acetylgalactosamine associated with protein and polysaccharide synthesis were significantly lower in EMBRB than those in CMBRB, which revealed the reason for the decrease of protein and polysaccharide content of EPS within EMBRB. This study provides useful information for improving A/O-MBR performance by probiotics.
1. Introduction
In recent years, eutrophication has caused serious environmental problems.1 Nitrogen is one of the main nutrients causing eutrophication.2 To solve the problem of nitrogen pollution, wastewater treatment plants usually use biological nitrogen removal due to its high efficiency and low cost.1 Among them, the anoxic/oxic (A/O) process has become one of the most widely used technologies in engineering due to its simple operation and stable pollutant removal. However, with increasingly stringent wastewater discharge standards, the effluent from the single A/O process no longer met the existing wastewater discharge standards.The membrane bioreactor (MBR) is an activated sludge treatment process combined with membrane filtration, which combines the biodegradation of activated sludge and the efficient separation of membrane, so it has the advantages of both, such as good solid–liquid separation, excellent effluent quality and small footprint.3,4 The combination of A/O and MBR (A/O-MBR), a process that the secondary sedimentation tank of A/O is replaced with membrane module for solid–liquid separation, can further improve the effluent quality. However, membrane fouling in practical applications severely limited the further spread and application of the technology.4,5 Many methods had been tried to reduce membrane fouling, including physical methods (e.g., backwashing and air spraying), chemical methods (e.g., chemical cleaning), and biological methods (e.g., phage).4,6 Whereas these methods did not fundamentally mitigate membrane fouling. The biocake was the important factor contributing to membrane fouling and EPS was the main component of the biocake.3,4 Therefore, EPS played a key role in the membrane fouling.4 EPS secretion was usually attributed to the communication mechanism called quorum sensing (QS) in which bacteria secreted EPS through signaling molecules.3,7,8 Generally the abundance of Gram-negative bacteria in MBR was significantly higher than that of Gram-positive bacteria.9 The N-acyl-homoserine lactones (AHLs) was the main signaling molecules used by Gram-negative bacteria to secreted EPS.8 Therefore, AHLs had been reported as the main signaling molecules responsible for EPS secretion in MBR.9,10 Degradation of AHLs, quorum quenching (QQ), is an important way to mitigate membrane biofouling. QQ enzymes had been successfully applied in MBR to reduce biofouling.9,10 However, due to the stability and cost of QQ enzymes, the practical QQ approach was needed. Considering that the membrane of A/O-MBR had high efficiency retention effect, the functional bacteria in the reactor were not easily lost. Therefore, inoculation of QQ bacteria (bacteria that could produce QQ enzymes) into the reactor might be a simpler and more economical way to mitigate biofouling. Previous studies had shown that the inoculation with QQ bacteria could significantly reduce biofouling in MBRs.9,11 In addition, the A/O-MBR process had internal circulation, which made it difficult to maintain the ideal anoxic condition in the anoxic tank and affected the denitrification efficiency. It was reported that simultaneous nitrification denitrification was achieved by inoculating an aerobic denitrifying bacterium into the oxic tank of A/O-MBR, which improved the total nitrogen (TN) removal efficiency of A/O-MBR.12
Therefore, in this study, an aerobic denitrifying bacterium with QQ activity (Acinetobacter sp. WZL728, OM967252), which was previously screened,13 was inoculated into the oxic tank of A/O-MBR to improve denitrification performance and reduce membrane biofouling. This study intended to: (i) evaluate the effect of strain WZL728 on the nitrogen and chemical oxygen demand (COD) removal; (ii) investigate the effect of strain WZL728 on membrane fouling by observing filtration cycles, transmembrane pressure (TMP) and fouling resistance; (iii) analyze the changes in the content and composition of AHLs and EPS in the biocake; (iv) assess the effect of strain WZL728 on microbial community structure in the biocake; (v) explore the effect of strain WZL728 on microbial metabolism through differential metabolites. This study revealed that strain WZL728 improved nitrogen removal performance and mitigated membrane biofouling of A/O-MBR through microbial community and metabolomics, which was beneficial for further application of strain WZL728 in A/O-MBR.
2. Materials and methods
2.1. A/O-MBR setup and operation
Two identical laboratory-scale A/O-MBRs were run in parallel (Fig. 1), one for the A/O-MBR with the addition of strain WZL728 (EMBR) and the other for the A/O-MBR without the addition of strain WZL728 (CMBR). The volume of the reactors was 9 L, including 3 L of anoxic tank and 6 L of oxic tank. The synthetic wastewater was pumped into the anoxic tank by a peristaltic pump and then stirred by a mechanical stirrer. The detailed composition of synthetic wastewater was shown in ESI.† The nitrogen source and carbon source of synthetic wastewater were provided by NH4Cl and CH3COONa, respectively. The main parameters of influent water were as follows: NH4+-N 41.44 ± 1.71 mg L−1, TN 42.24 ± 1.80 mg L−1, COD 422.49 ± 17.51 mg L−1. The C/N ratio of the influent water was about 10. The oxic tank was equipped with a polyvinylidene fluoride (PVDF) hollow fiber membrane (average pore size 0.01 μm, surface area 0.06 m2) and a perforated aeration device. CMBR and EMBR were operated at a constant filtration flux of 15 L (m−2 h−1). A peristaltic pump was used to extract the filtrate from the membrane module with a 10 min extraction cycle (9 min extraction and 1 min rest). According to the previous study,14 a recirculation ratio of 300% facilitated the removal of pollutant and reduced energy consumption. Therefore, a peristaltic pump was set up to return the nitrification solution from the oxic tank to the anoxic tank with a recirculation ratio of 300%. The dissolved oxygen (DO) was kept at about 0.4 mg L−1 in the anoxic tank and 3 mg L−1 in the oxic tank. The mixed liquor suspended solids (MLSS) concentration and mixed liquor volatile suspended solids (MLVSS) concentration were approximately 7500 mg L−1 and 4500 mg L−1, respectively. The sludge load was 0.13 kg COD per (kg MLVSS d). The hydraulic retention time (HRT) of CMBR and EMBR was 12 h, including 4 h in the anoxic tank and 8 h in the oxic tank. The SRT was controlled at 30 d by discharging a certain volume of sludge daily. The temperature in the reactors was maintained at about 24 °C and the pH was maintained at about 7.3. A pressure sensor was installed to detect the TMP. When the TMP rose to 40 kPa, the membrane module was taken out for cleaning (tap water rinse, 1% NaClO solution wash and 10% citric acid soak for 8 hours). | ||
| Fig. 1 The schematic diagram of CMBR and EMBR. | ||
The seed sludge from the aeration tank of a wastewater treatment plant (Beijing, China) was inoculated in CMBR and EMBR. From day 0 to day 30 (Start-up phase), the both reactors were acclimated to the synthetic wastewater. Thereafter, the A/O-MBRs underwent a relatively stable phase and ran for 30 days (Phase I). An aerobic denitrifying bacterium with QQ activity (Acinetobacter sp. WZL728, OM967252), which had been previously isolated,13 was inoculated into LB medium for enrichment. When the strain was in logarithmic phase, the aerobic denitrification performance, QQ activity and growth rate of the strain were at a high level.13 The bacterial solution of WZL728 with OD600 of 1.0 was used as the inoculum. According to previous bioaugmentation studies,1,11 the inoculum volume was 20 mL per day. During the phase II, the inoculum of WZL728 (OD600 = 1.0, 20 mL) was inoculated daily into the oxic tank of EMBR. Bioaugmentation was considered successful when the pollutant concentration in the effluent and the filtration cycles were stable. During the whole operation, the samples of influent and effluent were taken to determine NH4+–N, NO2−–N, NO3−–N, TN and COD in daily.
2.2. Determination of fouling resistance, AHLs and EPS
During the last four cycles of each of CMBR and EMBR, the membrane modules on day 4 were removed, and the fouling resistance, AHLs and EPS were measured. The fouling resistance (Rt), including membrane resistance (Rm), biocake resistance (Rc) and pore blocking resistance (Rp), was determined by referring to He et al.4 The content of AHLs and EPS in the biocake was determined by the following method.Specifically, the biocake was scraped off the membrane module and placed in a centrifuge tube containing 100 mL of PBS. The centrifuge tube was shaken to distribute biocake evenly. To ensure the accuracy of the experimental results, the biocake samples from the last four cycles of EMBR and CMBR were separately mixed for the subsequent experiments. The AHLs was extracted by the revised method.9 Specifically, 40 mL of the suspension was centrifuged at 11![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000g for 8 min. The upper liquid obtained by centrifugation was mixed with an equal volume of ethyl acetate and the mixture was then shook at 200 rpm for 1.5 hours. The upper organic phase was separated and dried in a rotary evaporator. Finally, 1 mL of formic acidified acetonitrile (0.1%) was used to dissolve the residue. The resulting solution was filtered through a syringe filter. The content of AHLs in the filtrate was determined by liquid chromatograph-mass spectrometer (LC-MS), and the detailed procedure was referred to Tabraiz et al.9 The EPS was obtained by referring to Shao et al.15 The EPS was considered to consist of protein (PN) and polysaccharide (PS), and the contents of PN and PS were determined by Lowry method and anthraquinone-sulfuric acid method, respectively.15,16
000g for 8 min. The upper liquid obtained by centrifugation was mixed with an equal volume of ethyl acetate and the mixture was then shook at 200 rpm for 1.5 hours. The upper organic phase was separated and dried in a rotary evaporator. Finally, 1 mL of formic acidified acetonitrile (0.1%) was used to dissolve the residue. The resulting solution was filtered through a syringe filter. The content of AHLs in the filtrate was determined by liquid chromatograph-mass spectrometer (LC-MS), and the detailed procedure was referred to Tabraiz et al.9 The EPS was obtained by referring to Shao et al.15 The EPS was considered to consist of protein (PN) and polysaccharide (PS), and the contents of PN and PS were determined by Lowry method and anthraquinone-sulfuric acid method, respectively.15,16
2.3. Microbial community analysis
The 16s rRNA gene high-throughput sequencing was applied to study the microbial community in CMBRB and EMBRB. The biomass samples used for community analysis were as described in Section 2.2 and stored at −20 °C before analysis. The DNA was collected from CMBRB and EMBRB by Mag-Bind Soil DNA Kit (OMEGA, USA). The polymerase chain reaction (PCR) amplification of 16S rDNA was performed with bacterial universal primers 341F (5′-CCTACGGGAGGCAGCAG-3′) and 805R (5′-GACTACHVGGGTATCTAATCC-3′) and the amplified DNA sequences were sequenced by Illumina MiSeq platform. The obtained results were clustered by OTU at 97% similarity using Usearch (version 9.2.64). The relative abundance of a taxon was set as the number of sequences associated with the taxon divided by the total number of sequences in the sample.2.4. Metabolomics analysis
Metabolomics was used to explore the differences in microbial metabolism between CMBRB and EMBRB, and the biological replicates were three for each group. Samples for metabolite analysis was described in Section 2.2. The extraction and detection of metabolites and data processing were performed by Songon Co. Ltd. (Shanghai, China). Specifically, 500 μL of solution (methanol:acetonitrile:water = 2:2:1 (V/V)) was dosed into 25 mg of sludge sample. The mixture was then grinded at 40 Hz for 3 min and sonicated in ice-water bath for 5 min and the above procedure was replicated 3 times. Afterwards, the sample was placed at – 40 °C for 1 h. Finally, the sample was centrifuged at 13800g for 15 min. The upper liquid was gathered for LC-MS detection. The original LC-MS data was transformed into mzXML format by ProteoWizard (version 3), treated with an in-house XCMS-based program and annotated by an in-house database (BiotreeDB).
2.5. Analysis methods
The concentration of nitrogen, COD, MLSS and MLVSS were determined by standard methods.17 Metabolic differences between CMBRB and EMBRB were observed by partial least squares discriminant analysis (PLS-DA). Significance of metabolite differences between EMBRB and CMBRB was determined by two-tailed t-test. Fold change (FC) meant that the average level of a metabolite in EMBRB divided by its average level in the CMBRB. All tests were performed in three independent replications and the results were presented as mean ± standard deviation (SD).3. Results and discussion
3.1. Pollutant removal
The Fig. 2 depicts the removal of nitrogen and organic matter in the EMBR and CMBR. During the start-up phase (0–30 days) and phase I (30–60 days), the above reactors performed similarly for pollutant removal. During the start-up phase, the NH4+–N concentration in the effluent of both reactors gradually decreased and kept at about 1.20 mg L−1 on day 8 (Fig. 2). Interestingly, the accumulation of NO2−–N and NO3−–N occurred in both A/O-MBRs during this phase. This might be due to the slower growth of nitrite oxidizing bacteria and denitrifying bacteria than ammonia oxidizing bacteria. Subsequently, the NO2−–N concentration in the effluent of the both A/O-MBRs gradually decreased until it was almost undetectable, while the NO3−–N concentration gradually decreased and reached a stable level on the 18 day (11.03 mg L−1). In addition, the COD removal efficiency gradually increased and stabilized at approximately 91.26% on day 9. During the phase I, the two reactors operated stably. The NH4+–N and NO3−–N concentrations in the effluent were kept at about 0.76 mg L−1 and 9.48 mg L−1, respectively, while NO2−–N was almost undetectable in the effluent. The COD and TN removal efficiency were maintained at approximately 92.66% and 75.05%, respectively. During phase II (60–150 days), strain WZL728 was inoculated to the EMBR, and the NH4+–N concentration in the effluent remained stable. The EMBR and CMBR showed no difference in the NH4+–N removal. Although the inoculation of WZL728 had no influence on NH4+–N removal by EMBR, the NO3−–N concentration in effluent decreased significantly and finally remained at about 2.75 mg L−1, which was clearly lower than the NO3−–N concentration in the effluent of CMBR (9.50 mg L−1). According to the nitrogen mass balance (ESI†), 57.56% of nitrogen is removed by denitrification in CMBR and 73.29% of nitrogen is removed by denitrification in EMBR. These were attributed to the aerobic denitrification function of strain WZL728. Accordingly, the TN removal efficiency increased significantly and finally stabilized at approximately 91.03% (Fig. 2). In addition, the COD removal efficiency in EMBR increased slightly and finally stabilized at about 99.05%, which was due to the fact that organic carbon source was required as electron donor for the denitrification. Thus, the inoculation of Acinetobacter sp. WZL728 could improve the degradation of NO3−–N and enhance the TN removal efficiency of the A/O-MBR.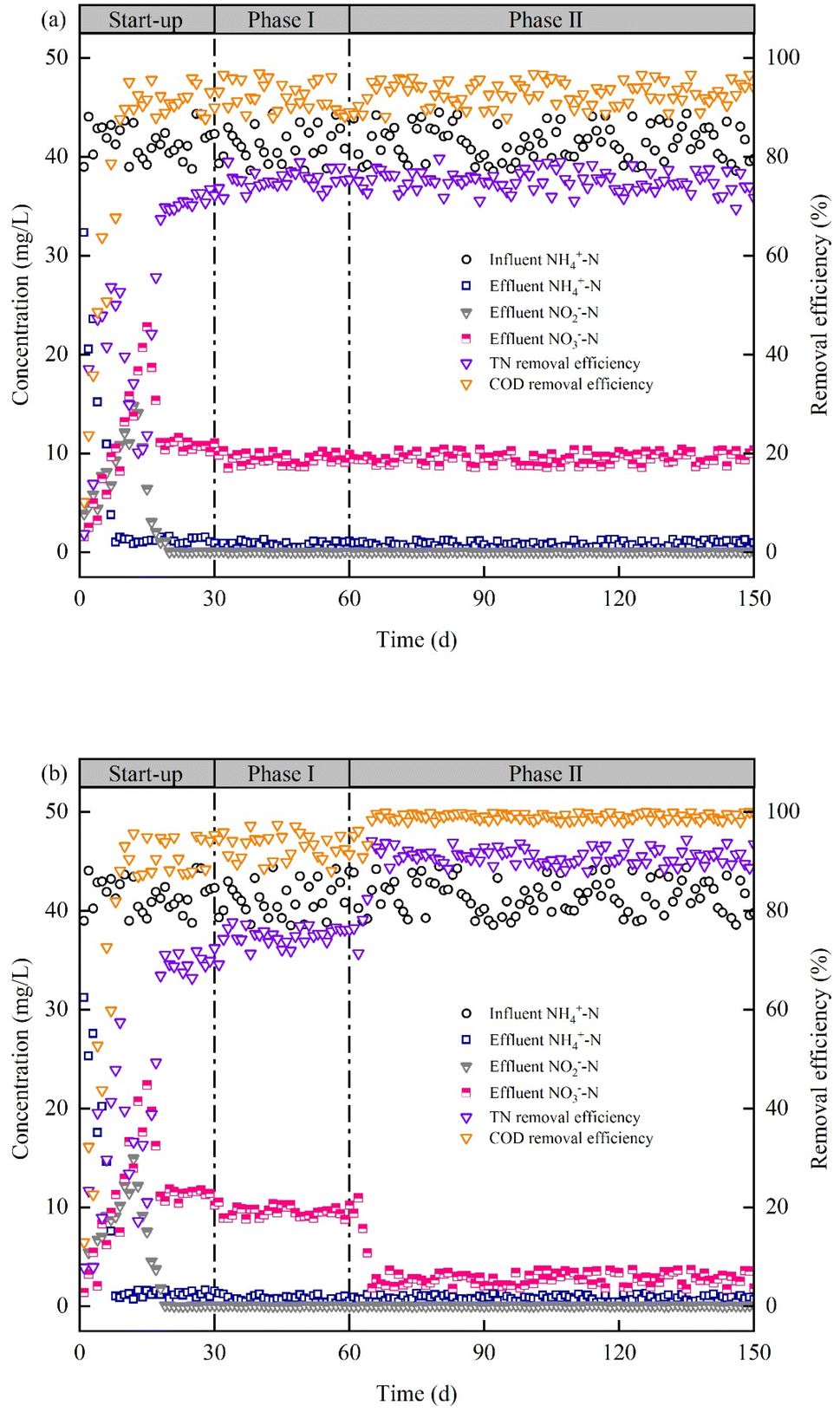 | ||
| Fig. 2 The performance of (a) CMBR and (b) EMBR for pollutant removal at different phases. | ||
3.2. Membrane fouling
During A/O-MBRs operation, the magnitude of the TMP reflected the degree of membrane fouling.16 When the TMP rose to 40 kPa, the membrane module was taken out for cleaning. The profile of filtration cycles for EMBR and CMBR during the whole operation period is shown in Fig. 3a. The average filtration cycle of the CMBR during phase I and II was 5.44 day. After the addition of strain WZL728, the average filtration cycle of EMBR was extended from 5.37 day during phase I to 9.57 day during phase II, and the EMBR was able to operate stably during phase II. In order to explore the reason for the prolonged EMBR filtration cycle during phase II, the TMP was investigated for each reactor from day 92 to day 112. The increase of TMP in the reactors was divided into two phases: slow increase at the beginning and rapid increase at the end (Fig. 3b). The main reason for the slow increase of TMP at the beginning was that the biocake was forming on the membrane. The rapid increase of TMP at the end could be the result of the complete formation of the biocake on the membrane leading to the loss of membrane connectivity.18 It was noteworthy that the TMP underwent a short period of slow increase followed by dramatic increase in each filtration cycle of CMBR, while the period of slow increase for TMP in each filtration cycle of EMBR was significantly longer than that of CMBR, which resulted in the prolongation of each filtration cycle of EMBR (Fig. 3b). In addition, Rt was 13.65 × 1012 m−1 in CMBR, of which Rc was 10.76 × 1012 m−1 accounted for 78.83%, however, Rt was 7.35 × 1012 in EMBR, of which Rc was 4.53 × 1012 accounted for 61.63% (Table 1). These results indicated that the biocake resistance was the primary factor in membrane resistance and WZL728 reduced the fouling resistance mainly by reducing the biocake resistance. Therefore, the addition of strain WZL728 was effective in slowing down the rise of TMP and prolonging the filtration cycle, which was mainly achieved by mitigating the formation of the biocake and extending the TMP slow increase phase. | ||
| Fig. 3 The profile of (a) filtration cycles during the whole operation and (b) TMP at 92–112 d of CMBR and EMBR. | ||
| Resistance distribution | CMBR | EMBR | ||
|---|---|---|---|---|
| m−1 | Percent of Rt | m−1 | Percent of Rt | |
| Rt | 13.65 × 1012 | 100% | 7.35 × 1012 | 100% |
| Rm | 1.17 × 1012 | 8.57% | 1.17 × 1012 | 15.92% |
| Rc | 10.76 × 1012 | 78.83% | 4.53 × 1012 | 61.63% |
| Rp | 1.72 × 1012 | 12.60% | 1.65 × 1012 | 22.45% |
According to past studies, many aerobic denitrifying bacteria were added to MBRs to improve total nitrogen removal, such as Acinetobacter sp. TSH1.1 However, in this study, the WZL728 (an aerobic denitrifying bacterium with quorum quenching activity) was added to the A/O-MBR, which not only improved its the total nitrogen removal but also reduced its membrane fouling. This indicated that strain WZL728 had great potential for bioaugmentation of MBR.
3.3. Effect of QQ bacteria on AHLs in the biocake
It was reported that QQ bacteria mitigated membrane fouling by degrading AHLs.7 To explore the reason for the prolongation of the TMP slow increase phase of EMBR during phase II, the concentration of AHLs was determined. As shown in Fig. 4, the concentrations of C8-HSL, 3-oxo-C8-HSL, C10-HSL, C12-HSL and 3-oxo-C12-HSL in CMBRB were 0.17, 1.05, 0.23, 1.24 and 0.07 ng mg−1 VSS, respectively. The concentrations of 3-oxo-C8-HSL and C12-HSL were significantly higher than those of C8-HSL, C10-HSL and 3-oxo-C12-HSL, which indicated that 3-oxo-C8-HSL and C12-HSL might be the major AHLs during the formation of the bioacake. The concentrations of C8-HSL, 3-oxo-C8-HSL, C10-HSL, C12-HSL and 3-oxo-C12-HSL in EMBRB were only 0.04, 0.11, 0.03, 0.12 and 0.01 ng mg−1 VSS, respectively. This indicated that the concentration of various AHLs in EMBRB were lower than that in CMBRB, especially 3-oxo-C8-HSL and C12-HSL. The contents of 3-oxo-C8-HSL and C12-HSL in EMBRB were only 10.48% and 9.68% of those in CMBRB, respectively. These should be due to the addition of strain WZL728, which was capable of degrading AHLs with long acyl chains (Fig. S1†). In addition, the concentration of total AHLs (C8-HSL, 3-oxo-C8-HSL, C10-HSL, C12-HSL and 3-oxo-C12-HSL) in EMBRB was only 11.23% of that in CMBRB, indicating that the concentration of AHLs in EMBRB was much lower than that in CMBRB. Therefore, strain WZL728 reduced the content of AHLs in EMBRB by 88.77%, which was significantly higher than the previously reported degradation efficiency of AHLs by Acinetobacter guillouiae ST01 (78.06%).9 This led to a lower likelihood of QS occurring in EMBRB, slowing down the formation of the biocake and extending the TMP slow increase phase, and ultimately mitigating membrane fouling in EMBR. | ||
| Fig. 4 The concentration of AHLs in CMBRB and EMBRB. | ||
3.4. Effect of QQ bacteria on EPS in biocake
According to previous studies, EPS was the key component of the biocake.3 Therefore, in this study, the content and composition of EPS in CMBRB and EMBRB were compared to reveal the reasons for the alleviation of membrane fouling by strain WZL728.As shown in Fig. 5, the content of soluble microbial products (SMP) in CMBRB and EMBRB were 63.84 and 38.63 mg g−1 VSS, respectively. High levels of SMP in CMBRB exacerbated membrane fouling by forming a gel layer,5 which was consistent with the rapid increase of TMP in CMBR (Fig. 3b). In addition, the SMP in CMBRB contained 36.56 mg g−1 VSS of PN and 27.28 mg g−1 VSS of PS with PN/PS of 1.34, while the SMP in EMBRB contained 25.81 mg g−1 VSS of PN and 12.82 mg g−1 VSS of PS with PN/PS of 2.01. The relatively lower PS content of SMP in EMBRB resulted in significantly higher PN/PS of SMP in EMBRB. Importantly, PS played a key role in bacterial adhesion to membrane in the early phases of membrane fouling.3 Therefore, the addition of strain WZL728 could not only effectively reduce the content of SMP, but also relatively reduce the content of PS and change the composition structure of SMP. In addition, the loosely bound extracellular polymeric substances (LB-EPS) content in EMBRB was only 57.13% of the LB-EPS content in CMBRB, which showed that the content of LB-EPS in EMBRB was significantly lower than that in CMBRB. However, the PN/PS of LB-EPS in EMBRB was 1.95, which was similar to that in CMBRB (1.89). Therefore, the addition of strain WZL728 reduced the level of LB-EPS in biocake and did not change the structure of LB-EPS. Interestingly, there was no significant difference in the content and structure of tightly bound extracellular polymeric substances (TB-EPS) between EMBRB and CMBRB, which was inconsistent with the significant difference in TMP between CMBR and EMBR. The structure of TB-EPS became dense during the continuous filtration process, so the changes in external conditions did not easily change the property of TB-EPS.8 Overall, the levels of SMP and LB-EPS in EMBRB were significantly lower than those in CMBRB, which was consistent with the low concentration of AHLs in EMBRB (Fig. 4). It was reported that AHLs was essential for the regulation and secretion of EPS.8 Thus, strain WZL728 slowed down the formation of the biocake and mitigated membrane fouling by degrading AHLs to reduce SMP and LB-EPS content in the biocake.
 | ||
| Fig. 5 The concentration of PS and PN in CMBRB and EMBRB. | ||
3.5. Microbial community
During the formation of biocake, microorganisms played an important role.3 Therefore, the 16S rRNA gene high-throughput sequencing was applied to identify microorganisms in CMBRB and EMBRB.At the phylum level, the dominant phylum in EMBRB and CMBRB were both Proteobacteria, Bacteroidetes and Chloroflexi (Fig. 6a). It was reported that Proteobacteria was the main phylum in the formation of biocake leading to membrane fouling.19 The abundance of Proteobacteria in EMBRB (34.74%) was significantly lower than that in CMBRB (42.32%). Proteobacteria belonging to Gram-negative bacteria used AHLs-mediated QS to participate in biocake formation. Strain WZL728 limited the involvement of Proteobacteria in the formation of biocake by QQ, which contributed to slow down membrane fouling of EMBR.19 Bacteroidetes was another widespread phylum in wastewater biological treatment systems and main microorganisms contributing to membrane biofouling, which could accelerate the membrane fouling process by attaching to the membrane through its pilus.7 Similarly, Bacteroidetes also belonged to Gram-negative bacteria. Strain WZL728 reduced its abundance in EMBRB by 4.45% through QQ. In addition, Chloroflexi, which normally lived under anaerobic conditions, grew inside biofilms and was not susceptible to environmental influence.4 Thus the addition of strain WZL728 had no significant effect on its abundance. Notably, the abundance of Firmicutes which belonged to Gram-positive bacteria was upregulated from 5.56% in CMBRB to 11.34% in EMBRB, suggesting that strain WZL728 favored the growth of Gram-positive bacteria by inhibiting the growth of Gram-negative bacteria. In addition, Firmicutes was able to degrade organic matter causing membrane fouling, which might be a significant reason for the decrease of EPS in EMBRB.16
 | ||
| Fig. 6 The distribution of the microbial community at (a) phylum and (b) class in CMBRB and EMBRB. | ||
At the class level, Alpthaproteobacteria belonging to Proteobacteria and Sphingobacteriia belonging to Bacteroidetes were dominant class in CMBRB and EMBRB (Fig. 6b). The abundance of Alpthaproteobacteria in CMBRB was 23.65%, while in EMBRB the abundance of Alpthaproteobacteria was 18.76%. Previous studies had reported that Alpthaproteobacteria could secret EPS, in which it relies on the AHLs-mediated QS system.19 The addition of strain WZL728 reduced the concentration of AHLs in EMBRB, inhibited the EPS secretion of Alpthaproteobacteria. Sphingobacteriia belonging to Bacteroidetes was able to further aggravate membrane fouling.4 The strain WZL728 inhibited the growth of Sphingobacteriia. The abundance of Sphingobacteriia decreased from 15.23% in CMBRB to 12.58% in EMBRB, which was consistent with the slowing of membrane fouling in the EMBR (Fig. 3). In addition, the abundance of Gammaproteobacteria in EMBRB (10.35%) was significantly higher than that in CMBRB (6.68%) due to the inoculation of Acinetobacter sp. WZL728 belonging to Gammaproteobacteria. Clostridia belonging to Firmicutes was able to hydrolyze PS20 and its abundance increased by 5.48% in EMBRB. This was consistent with the relatively low PS content of EPS in EMBRB and was beneficial to mitigate membrane biofouling in EMBR.
Overall, the decrease in the abundance of the Gram-negative bacteria associated with EPS secretion (Alpthaproteobacteria) and the increase in the abundance of Gram-positive bacteria associated with EPS degradation (Clostridia) in EMBRB revealed the reason for its reduced EPS content. This suggested that strain WLZ728 influenced the membrane fouling by altering the concentration of AHLs and thus regulating the abundance of bacteria associated with EPS secretion and degradation in the biocake. In addition, the abundance of Gammaproteobacteria increased accordingly in EMBRB due to the addition of strain WZL728, which was consistent with the enhanced denitrification performance in EMBR.
3.6. Metabolomics
The activities in microorganisms are carried out by numerous macromolecules, and the changes in macromolecules are ultimately reflected at the metabolic level. Metabolomics studies the changes in the metabolites of microorganisms between different environments. Therefore, in order to investigate the differences in microorganism metabolism between CMBRB and EMBRB, metabolomics was performed.As shown in PLS-DA (Fig. 7a), there was a clear separation between CMBRB and EMBRB, indicating that the metabolic state in the biocake between the two reactors was significantly different. This indicated that the inoculation of strain WZL728 leaded to completely different metabolic response of the microorganisms in EMBRB. Specifically, 303 metabolites were detected and 97 metabolites were significantly altered, of which 43 were upregulated (p < 0.05; >2-FC) and 54 were downregulated (p < 0.05; <0.5-FC) in EMBRB (Fig. 7b). This indicated that strain WZL728 interfered with the level of metabolites. The variable importance in projection (VIP) values derived from the weighting coefficients of PLS-DA were able to be used to rank the segregation contribution of metabolites to CMBRB and EMBRB. Therefore, the differential metabolites between CMBRB and EMBRB were screened based on VIP values (>1), FC (>2 or <0.5) and p-values (<0.05), and the results are shown in Fig. 7c. The main amino acids affecting extracellular PN were alanine (Ala) and valine (Val).21 The levels of Ala and Val in EMBRB were only 0.26 and 0.28 of those in CMBRB, respectively. This might be an important reason why the PN level of EPS in EMBRB was significantly lower than that in CMBRB. PS of EPS produced by bacteria played a key role in aggravating membrane fouling. Uridine diphosphate-N-acetylgalactosamine (UDP-GlcNAc), which was an important metabolite involved in PS synthesis, was detected at significantly lower level in EMBRB than that in CMBRB (0.21-FC, p < 0.01).21 It was reported that AHLs mediated extracellular PS and PN production through the regulation of PS and PN synthesis-related metabolites.21 Strain WZL728 mainly degraded 3-oxo-C8-HSL and C12-HSL in EMBRB. Thus, the reason for the differences in PN and PS of EPS between CMBRB and EMBRB may be that strain WZL728 downregulated the levels of metabolites associated with PN and PS synthesis by hydrolyzing 3-oxo-C8-HSL and C12-HSL in EMBRB. Furthermore, UDP-GlcNAc has a higher VIP value compared to Val and Ala (Fig. 7c), which might be an important reason for the relatively lower PS content of EPS in EMBRB (Fig. 3). Thus, strain WZL728 could reduce EPS content by hydrolyzing AHLs and thereby reducing metabolites associated with the synthesis of PN and PS. This revealed the reason for reduced EMBR membrane fouling from the metabolic perspective. Nicotinamide adenine dinucleotides (NAD+), which was involved in oxidation–reduction reactions, played an important role in the conversion of nitrate to gaseous nitrogen.22 Notably, NAD+ was significantly upregulated (3.70-FC) in EMBRB. This was consistent with the improved denitrification performance in EMBR (Fig. 2).
 | ||
| Fig. 7 (a) PLS-DA; (b) volcano plot based on FC and p-value. Red dots indicated that the level of these metabolites was significantly higher in EMBRB than that in CMBRB (FC > 2, p < 0.05). Blue dots indicated that the level of these metabolites was significantly lower in EMBRB than that in CMBRB (FC < 0.5, p < 0.05); (c) differential metabolites screened according to VIP value (>1), FC (>2 or <0.5) and p-value (<0.05). The differential metabolites with VIP value in top 15 were listed. “**” indicated p < 0.01. | ||
The influent water used in this study was synthetic wastewater, which still had a large gap with the real wastewater. The real wastewater not only contained a wide variety of microorganisms but also had complex water quality, which might have an impact on the microbial community and metabolism in the reactor. If the influent water of A/O-MBR is real wastewater, whether inoculation of WZL728 can effectively improve the total nitrogen removal and reduce membrane fouling deserves investigation.
4. Conclusions
This study showed that an aerobic denitrifying bacterium with QQ activity (strain WZL728) was able to improve the performance of A/O-MBR by enhancing nitrogen removal performance and reducing membrane biofouling. Due to the aerobic denitrification function of WZL728, the denitrification performance improved and the TN removal efficiency increased to 91.03% in EMBR. Compared to CMBR, the filtration cycle in EMBR was extended by 1.76 times with the addition of the strain, which was mainly achieved by extending the TMP slow increase phase. During the slow increase phase of TMP, the main AHLs (3-oxo-C8-HSL and C12-HSL) in EMBRB were substantially degraded and the concentration of total AHLs in EMBRB was only 11.23% of that in CMBRB due to the QQ activity of WZL728. The decrease of AHLs reduced the content of SMP and LB-EPS in EMBRB by 39.49% and 42.87%, respectively, by adjusting the microbial community structure and microbial metabolism. On the one hand, decreased abundance of Gram-negative bacteria (Alpthaproteobacteria) associated with EPS secretion and increased abundance of Gram-positive bacteria (Clostridia) associated with EPS degradation led to the decrease of EPS content in EMBRB. On the other hand, decreased levels of metabolites (Ala, Val and UDP-GlcNAc) associated with PS and PN synthesis led to the decrease of EPS in EMBRB. Therefore, this study revealed how WZL728 improved the nitrogen removal performance and mitigated membrane fouling, which was beneficial for further application of the probiotics in A/O-MBR.Author contributions
Conceptualization, Pengyuan Wei; data curation, Chaofan Xie; funding acquisition, Jing Zhang; investigation, Pengyuan Wei; methodology, Pengyuan Wei; project administration, Jun Li and Jing Zhang; resources, Jun Li; software, Yi Zhang and Rong Luo; writing – review & editing, Jing Zhang.Conflicts of interest
The authors declare no conflict of interest.Acknowledgements
This research was supported by Beijing Natural Science Foundation (8222039).References
- J. L. Ren, X. W. Cheng, H. J. Ma and X. N. Ma, Characteristics of a novel heterotrophic nitrification and aerobic denitrification bacterium and its bioaugmentation performance in a membrane bioreactor, Bioresour. Technol., 2021, 342, 125908 CrossRef CAS PubMed.
- P. P. Chen, F. P. Zhang, L. J. Zhang, H. Liu, Q. Zhang, Z. L. Xing and T. T. Zhao, Characterization of a novel salt-tolerant strain Sphingopyxis sp. CY-10 capable of heterotrophic nitrification and aerobic denitrification, Bioresour. Technol., 2022, 358, 127353 CrossRef CAS PubMed.
- B. Jiang, Q. Z. Zeng, Y. Hou, H. X. Li, J. X. Liu, J. Xu, S. N. Shi and F. Ma, Impacts of long-term electric field applied on the membrane fouling mitigation and shifts of microbial communities in EMBR for treating phenol wastewater, Sci. Total Environ., 2020, 716, 137139 CrossRef CAS PubMed.
- H. Y. He, X. D. Xin, W. Qiu, D. Li, Z. C. Liu and J. Ma, Role of nano-Fe3O4 particle on improving membrane bioreactor (MBR) performance: alleviating membrane fouling and microbial mechanism, Water Res., 2022, 209, 117897 CrossRef CAS PubMed.
- L. J. Deng, W. S. Guo, H. H. Ngo, X. C. C. Wang, Y. S. Hu, R. Chen, D. L. Cheng, S. Q. Guo and Y. Y. Cao, Application of a specific membrane fouling control enhancer in membrane bioreactor for real municipal wastewater treatment: sludge characteristics and microbial community, Bioresour. Technol., 2020, 312, 123612 CrossRef CAS PubMed.
- M. T. Xue, H. Gao, X. A. Dong, M. J. Zhan, G. P. Yang and R. Yu, Promotion and mechanisms of Bdellovibrio sp. Y38 on membrane fouling alleviation in membrane bioreactor, Environ. Res., 2022, 212, 113593 CrossRef CAS PubMed.
- H. Waheed, Y. Y. Xiao, I. Hashmi and Y. Zhou, The selective pressure of quorum quenching on microbial communities in membrane bioreactors, Chemosphere, 2020, 247, 125953 CrossRef CAS PubMed.
- J. H. Huang, Y. L. Gu, G. M. Zeng, Y. Yang, Y. C. Ouyang, L. X. Shi, Y. H. Shi and K. X. Yi, Control of indigenous quorum quenching bacteria on membrane biofouling in a short-period MBR, Bioresour. Technol., 2019, 283, 261–269 CrossRef CAS PubMed.
- S. Tabraiz, B. Shamurad, E. Petropoulos, M. Quintela-Buluja, A. Charlton, J. Dolfing and P. J. Sallis, Mitigation of membrane biofouling in membrane bioreactor treating sewage by novel quorum quenching strain of Acinetobacter originating from a full-scale membrane bioreactor, Bioresour. Technol., 2021, 334, 125242 CrossRef CAS PubMed.
- S. Y. Ham, H. S. Kim, E. Cha, J. H. Park and H. D. Park, Mitigation of membrane biofouling by a quorum quenching bacterium for membrane bioreactors, Bioresour. Technol., 2018, 258, 220–226 CrossRef CAS PubMed.
- N. N. Shu, H. Park, S. S. A. Shah, N. Maneda, H. J. Yoo, J. Min, I. Angelidaki and K. H. Choo, Probiotic strategy for biofouling control through direct injection of quorum-quenching bacteria into membrane bioreactors, Chem. Eng. J., 2022, 438, 135572 CrossRef CAS.
- J. F. Guo, P. Guo, S. M. Li, X. J. Li, C. L. Luo, H. Wang and Q. Jiang, Screening an effective aerobic denitrification strain and its performance in MBR, Pol. J. Environ. Stud., 2017, 26, 1089–1096 CrossRef CAS.
- P. Y. Wei, J. Zhang, J. Li, Z. M. Zheng, Z. Z. Wang, H. Han, Y. Zhang and R. Luo, Isolation and identification of a novel aerobic denitrifying bacterium with quorum quenching activity and the effect of environmental factors on its performance, Environ. Technol. Innovat., 2022, 28, 102913 CrossRef CAS.
- X. H. Du, F. Zhao, S. H. Lin and D. D. Wang, Study on the Effects of Influencing Factors on the Treatment of Domestic Sewage by A/O-MBR, Guangdong Chem. Ind., 2013, 40, 139–141 Search PubMed.
- Y. J. Shao, Z. Zhou, J. Jiang, L. M. Jiang, J. P. Huang, Y. Zuo, Y. Q. Ren and X. D. Zhao, Membrane fouling in anoxic/oxic membrane reactors coupled with carrier-enhanced anaerobic side-stream reactor: effects of anaerobic hydraulic retention time and mechanism insights, J. Membr. Sci., 2021, 637, 119657 CrossRef CAS.
- Z. H. Niu, H. Z. Guo, Y. Zhou and S. Q. Xia, Unraveling membrane fouling in anoxic/oxic membrane bioreactors treating anaerobically digested piggery wastewater, J. Environ. Chem. Eng., 2021, 9, 104985 CrossRef CAS.
- APHA, Standard Methods for the Examination of Water and Wastewater, American Public Health Association, Washington, DC, USA, 21st edn, 2006 Search PubMed.
- J. B. Liu, C. Y. Eng, J. S. Ho, T. H. Chong, L. Wang, P. Y. Zhang and Y. Zhou, Quorum quenching in anaerobic membrane bioreactor for fouling control, Water Res., 2019, 156, 159–167 CrossRef CAS PubMed.
- X. Xiao, S. J. You, H. J. Guo, F. Ma, J. N. Zhang, R. Y. Zhang and X. T. Bao, Mycelial pellets for alleviation of membrane fouling in membrane bioreactor, J. Membr. Sci., 2021, 635, 119545 CrossRef CAS.
- M. More, A. M. Veijalainen and H. Heinonen-Tanski, Performic Acid Controls Better Clostridium tyrobutyricum Related Bacteria than Peracetic Acid, Sustainability, 2018, 10, 4116 CrossRef.
- X. Tang, Y. Z. Guo, S. S. Wu, L. M. Chen, H. C. Tao and S. T. Liu, Metabolomics Uncovers the Regulatory Pathway of Acyl-homoserine Lactones Based Quorum Sensing in Anammox Consortia, Environ. Sci. Technol., 2018, 54, 2206–2216 CrossRef PubMed.
- X. T. Yang, J. H. Zhou, T. R. Huo, Y. F. Lv, J. J. Pan, L. M. Chen, X. Tang, Y. P. Zhao, H. L. Liu, Q. Gao and S. T. Liu, Metabolic insights into the enhanced nitrogen removal of anammox by montmorillonite at reduced temperature, Chem. Eng. J., 2021, 410, 128290 CrossRef CAS.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d2ra06676a |
| This journal is © The Royal Society of Chemistry 2023 |