
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicencePreservation of heparin-binding EGF-like growth factor activity on heparin-modified poly(N-isopropylacrylamide)-grafted surfaces†
Jun Kobayashi *a,
Yoshinori Arisakab,
Nobuhiko Yuib,
Masayuki Yamatoa and
Teruo Okanoac
*a,
Yoshinori Arisakab,
Nobuhiko Yuib,
Masayuki Yamatoa and
Teruo Okanoac
aInstitute of Advanced Biomedical Engineering and Science, Tokyo Women's Medical University, TWIns, 8-1 Kawadacho, Shinjuku-ku, Tokyo 162-8666, Japan. E-mail: kobayashi.jun@twmu.ac.jp
bDepartment of Organic Biomaterials, Institute of Biomaterials and Bioengineering, Tokyo Medical and Dental University, 2-3-10 Kanda-Surugadai, Chiyoda-ku, Tokyo 101-0062, Japan
cCell Sheet Tissue Engineering Center, Department of Pharmaceutics and Pharmaceutical Chemistry, Health Sciences, University of Utah, 30 South 2000 East, Salt Lake City, UT 84112, USA
First published on 18th November 2021
Abstract
A heparin-modified poly(N-isopropylacrylamide) (PIPAAm)-grafted surface bound with heparin-binding epidermal growth factor-like growth factor (HB-EGF) was able to culture hepatocytes maintaining high albumin secretion and high expression of hepatocyte-specific genes. However, the activity of HB-EGF on the surface and its binding effects on hepatocytes remain unclear. In this study, we investigated the temperature-dependent interactions of HB-EGF and EGF receptor (EGFR) with heparin-modified PIPAAm to evaluate the activity of HB-EGF on the surface. Quartz crystal microbalance (QCM) measurements revealed that the amounts of adsorbed HB-EGF on either the heparin-modified PIPAAm-grafted surface (heparin-IC1) or PIPAAm-grafted surfaces were almost the same regardless of swelling/deswelling of grafted PIPAAm chains. The heparin-IC1 surface bound to HB-EGF at 37 °C had the ability to bind to hepatocytes through specific affinity interaction with EGFR, whose activation was confirmed by western blotting. However, the physisorbed HB-EGF on the PIPAAm surface greatly diminished its activity. Taken together, the introduction of heparin into grafted PIPAAm chains on the surface plays a pivotal role in holding HB-EGF while preserving its activity. Hydration and swelling of surface-grafted PIPAAm chains at 20 °C greatly diminished the attachment of hepatocytes with HB-EGF bound to heparin-IC1, whereas hepatocytes were able to bind to HB-EGF bound to heparin-IC1 at 37 °C. Thus, the equilibrated affinity interaction between EGFRs and surface-bound HB-EGF was considered to be attenuated by steric hindrance due to hydration and/or swelling of grafted PIPAAm chains.
Introduction
Creation of hepatic tissues is an attractive approach for the treatment of liver diseases, such as congenital enzyme deficiencies1,2 and haemophilia,3 and for in vitro models to investigate liver disease,4,5 drug metabolism,6 drug screening,7 and drug toxicity.8 Primary hepatocytes isolated from rodent liver have often been used as a building block for these tissues because of the well-known hepatic functions of terminally differentiated mature hepatocytes. Typically, the isolated hepatocytes were utilized for the formation of three-dimensional tissues such as spheroids formed by self-aggregation,9,10 and scaffold-based tissue using biodegradable polymer.11,12 In our laboratory, a unique method to create scaffold-free and transplantable hepatic tissues was developed using a temperature-responsive culture dish.1,2 The temperature-responsive poly(N-isopropylacrylamide) (PIPAAm)-grafted surface exhibited hydrophilic/hydrophobic alterations across the lower critical solution temperature (LCST) of 32 °C, resulting in reversible cell attachment/detachment.13,14 Primary hepatocytes adhered to and formed a monolayer on the hydrophobized PIPAAm-grafted surface at 37 °C. Cultured hepatocyte sheets were recovered from the hydrated surface as a single layer when the temperature was decreased to 20 °C. Transplantation of hepatocyte sheets was performed at a pre-vascularized subcutaneous site, resulting in long-term secretion of transplanted hepatocyte-specific enzymes.1To create transplantable hepatocyte sheets while maintaining their functions, a heparin-modified PIPAAm-grafted surface was designed to stimulate the receptors of the hepatocytes and recover non-enzymatically cultured cells.15,16 In general, the viability and phenotypic functions of hepatocytes are impaired when isolated from the native in vivo microenvironment of the liver.17 Immobilised heparin molecules on grafted PIPAAm chains were able to capture heparin-binding epidermal growth factor (EGF)-like growth factor (HB-EGF). Hepatocytes cultured on HB-EGF bound to the heparin-modified PIPAAm-grafted surfaces exhibited higher albumin secretion, and higher expression of hepatocyte-specific genes (albumin, hepatocyte nuclear factor 4 alpha, coagulation factor VII, and coagulation factor IX) compared with those on the PIPAAm surface with soluble HB-EGF. EGF tethered through terminal amino groups to a solid surface continuously stimulates cultured hepatocytes, whereas physisorbed EGF on the solid surface exhibits no activity.18 Additional enzymatic treatments, such as trypsin, are required for the liberation of the cultured hepatocytes from the substrate, resulting in the suspension of individual cells. In contrast, the cultured hepatocytes on the HB-EGF bound with the heparin-modified PIPAAm-grafted surfaces were detachable as a single sheet by decreasing the temperature below LCST at 20 °C.16 Although cultured hepatocytes on the HB-EGF-bound surface exhibited high albumin secretion and high expression of hepatocyte-specific genes, the activity of HB-EGF on the surface and their binding effects on hepatocytes remain unclear.
We hypothesised that bound HB-EGF on heparin-modified PIPAAm-grafted surfaces was preserved compared to physisorbed HB-EGF on PIPAAm-grafted surfaces. In previous studies,15,16,18 the activity of bound growth factors was judged by indirect information such as cell growth, spreading, and functions. In this study, we investigated direct and quantitative evidences regarding the preservation of bound growth factor, HB-EGF. Prior to investigating the activity, the amount of bound HB-EGF on PIPAAm-grafted surfaces that were modified with and without heparin was quantified using quartz crystal microbalance (QCM) measurements. In addition, the dynamic adsorption/desorption of HB-EGF during stepwise temperature change on the surfaces was monitored. For elucidating the binding of EGF receptor (EGFR) and surface-bound HB-EGF, the attachment of hepatocytes on the surfaces was performed during short-term incubation, 1.5 h. Temperature-dependent attachment of hepatocytes to HB-EGF bound with heparin-modified PIPAAm-grafted surface and their inhibition with soluble EGF were investigated to confirm EGFR-mediated attachment of hepatocytes, and stimulation of hepatocyte EGFR with the bound HB-EGF was also investigated using western blotting. Taken together, we verified that the introduction of heparin into grafted PIPAAm chains on the surface was significant for holding HB-EGF while preserving its activity.
Experimental
Materials
N-Isopropylacrylamide (IPAAm) was kindly gifted by KJ Chemicals (Tokyo, Japan) and purified by recrystallisation in n-hexane. The compound 2-carboxyisopropylacrylamide (CIPAAm) was synthesised according to a method described by a previous report.19 In addition, 2-propanol, 1-ethyl-3-(3-dimetylaminopropyl)-carbodiimide hydrochloride (EDC), N-hydroxysuccinimide (NHS), high-glucose Dulbecco's modified Eagle medium (DMEM), Dulbecco's phosphate buffered saline (PBS), and penicillin-streptomycin solution (100×) were obtained from Fujifilm Wako Pure Chemicals (Osaka, Japan), and 2-morpholinoethanesulfonic acid monohydrate (MES) and O,O′-bis(2-aminoethyl)ethyleneglycol-N,N,N′,N′-tetraacetic acid (EGTA) were purchased from Dojindo (Kumamoto, Japan). Heparin sodium salt from porcine intestinal mucosa (grade I-A, 180 USP units per mg), bovine serum albumin (BSA), trypsin inhibitor, Ca2+, Mg2+-free Hanks' balanced salt solution, Hanks' balanced salt solution 10×, protease inhibitor cocktail, phosphatase inhibitor cocktail 2, and phosphatase inhibitor cocktail 3 were obtained from Sigma-Aldrich (St. Louis, MO, USA). Hanks' balanced salt solution (Nacalai Tesque, Kyoto, Japan), recombinant human HB-EGF (R&D Systems, Minneapolis, MN, USA), recombinant human EGF (PeproTech, Cranbury, NJ, USA), 50% glucose, 7% NaHCO3, 2% CaCl2 (Otsuka Pharmaceutical, Tokyo, Japan), collagenase type IV (Worthington Biochemical, Lakewood, NJ, USA), 4% paraformaldehyde fixative (Muto Pure Chemical, Tokyo, Japan), and Western Marker Neo (high) (Beacle, Kyoto, Japan) were used as received. Furthermore, 10× Tris-buffered saline, 10% Tween 20, and 7 × 8.4 cm nitrocellulose membrane with 0.2 μm pores were obtained from Bio-Rad Laboratories (Hercules, CA, USA), and 1 M N-2-hydroxyethylpiperazine-N-2-ethane sulfonic acid (HEPES), Hoechst 33258, TCEP-HCl, NuPAGE LDS sample buffer, 4–12% gradient NuPAGE™ gel, NuPAGE™ MOPS SDS running buffer (20×), NuPAGE™ Transfer Buffer (20×), NuPAGE™ Antioxidant, and Novex reversible membrane protein stain kit were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Non-fat dry milk, anti-phospho-EGFR monoclonal rabbit antibody #3777, and anti-EGFR polyclonal rabbit antibody #2232 were obtained from Cell Signalling Technology (Danvers, MA, USA). Percoll, horseradish peroxidase-conjugated donkey anti-rabbit antibody (NA934VS), and ECL™ Prime Western Blotting System were obtained from Cytiva (Marlborough, MA, USA). A protein quantification assay kit was obtained from Macherey-Nagel (Düren, Germany).Preparation of heparin-modified temperature-responsive polymer-grafted surfaces
Poly(IPAAm-co-CIPAAm) (IC1) and heparin-modified IC1 surfaces (heparin-IC1) on QCM sensor chips coated with O2 plasma-treated polystyrene, commercially available tissue culture polystyrene dishes were prepared according to previous studies.15,20 The amount of grafted temperature-responsive polymers on TCPS was determined using an attenuated total reflection Fourier transform infrared spectrometer according to a previous report.15 The grafted amount of temperature-responsive polymer on IC1 and PIPAAm surfaces on 35 mm dishes was 1.64 ± 0.11 and 1.98 ± 0.37 μg cm−2 (mean ± SD), respectively. The difference between the means was not significant (P = 0.25).QCM measurement for quantification of HB-EGF adsorption/desorption
The mass of adsorbed HB-EGF on the PIPAAm and heparin-IC1 surfaces of the QCM sensor chips was determined using a QCM system, according to a previous report.20 All of the solutions to be injected into the QCM sensor were kept at 40 °C to eliminate the formation of bubbles within the tubing and the QCM sensor. At a constant temperature of 20 or 37 °C, a solution of 440 ng mL−1 HB-EGF in PBS, which corresponded to a surface concentration of 100 ng cm−2 HB-EGF on the surface area of 8.8 cm2 in 2 mL, was injected into the flow module at a flow rate of 50 μL min−1 for 60 min. Next, the surface was washed with PBS for 15 min. The adsorbed areal mass was calculated according to the previous report.20Hepatocyte isolation
Male inbred F344/Nslc rats from 8 to 12 weeks of age (Japan SLC, Shizuoka, Japan) were used to prepare isolated primary hepatocytes. The rats were housed under temperature-controlled conditions with a 12 h light/dark cycle and had ad libitum access to rat chow and water. The pre-perfusion solution was Ca2+, Mg2+-free Hanks' balanced salt solution containing 0.5 mM EGTA, 10 mM HEPES, 0.8 g L−1 glucose, 100 U mL−1 penicillin, and 100 μg mL−1 streptomycin. Collagenase solution was Hanks' balanced salt solution containing 0.5 g L−1 collagenase type IV, 0.1 g L−1 trypsin inhibitor, 0.4 g L−1 CaCl2, 1.75 g L−1 NaHCO3, 10 mM HEPES, 0.8 g L−1 glucose, 100 U mL−1 penicillin, and 100 μg mL−1 streptomycin. The washing solution was high-glucose DMEM containing 10% FBS, 10 mM HEPES, 100 U mL−1 penicillin, and 100 μg mL−1 streptomycin.Isolation of hepatocytes from rat liver was carried out using a modified two-step collagenase perfusion method, as previously described.21,22 After the rat was laparotomized under anaesthesia using isoflurane inhalation, a cannula was inserted through the portal vein using a 22G × 1′′ indwelling needle (Terumo, Tokyo, Japan) and tied to the vein using a surgical suture. Tubing from a peristaltic pump with the pre-perfusion solution in a water bath at 39 °C was connected to the inserted cannula at a flow rate of 10 mL min−1. Immediately after the inferior vena cava was cut, first-step perfusion was performed for 5 min at a flow rate of 25 mL min−1. After switching to the collagenase solution, the second-step perfusion was carried out at a flow rate of 25 mL min−1 for 5–8 min. The digested liver was liberated from the body and scratched with a scalpel in the washing solution on ice. The suspension was filtered through a stainless cell strainer with a 60 μm mesh and stored on ice until purification.
To recover the vial cells, purification was performed using the Percoll centrifugation method.23 After centrifugation at 4 °C and 50 × g for 3 min, the cells were resuspended in fresh washing solution to obtain 25 mL of suspension. The suspension was mixed with Percoll solution containing 21.6 mL Percoll and 2.4 mL Hanks' balanced salt solution 10×, followed by centrifugation at 4 °C and 50 × g for 20 min. After the dead cells and supernatant were removed, the cells were resuspended in fresh washing solution and stored on ice until use.
All animal experiments were approved by the Animal Care and Use Committee of Tokyo Women's Medical University and performed in accordance with the institutional guidelines of the Tokyo Women's Medical University on Animal Use.
Initial attachment to HB-EGF-bound temperature-responsive surfaces
Isolated hepatocytes, as described in the previous section, were used to investigate attachment to the surfaces. Heparin-IC1 and PIPAAm surfaces bound to HB-EGF were prepared by incubating 2 mL of 440 ng mL−1 HB-EGF solution in PBS, which corresponded to 100 ng cm−2 on a surface area of 8.8 cm2, at 37 °C for 24 h. The surfaces were washed three times with 2 mL of PBS at 37 °C. The suspension of hepatocytes from the previous section was centrifuged at 4 °C at 50 × g for 3 min and resuspended in high-glucose DMEM containing 2 mg mL−1 BSA, 100 U mL−1 penicillin, and 100 μg mL−1 streptomycin. The hepatocytes were seeded at a density of 5 × 104 cells per cm2 on heparin-IC and PIPAAm surfaces bound with HB-EGF or PIPAAm as a control and incubated in a 5% CO2 incubator at 37 °C for 1.5 h. The incubated hepatocytes were gently rinsed with PBS, fixed with 4% paraformaldehyde fixative for 15 min, and washed twice with PBS. Fluorescent images of the fixed hepatocytes, where the nuclei were stained with 2 μg mL−1 Hoechst 33258 in PBS for 15 min, were obtained using an AxioCam HRc colour CCD (Carl Zeiss, Jena, Germany) controlled with AxioVision 4.8.2.0 software (Carl Zeiss) under a fluorescence microscope (Eclipse TE2000-U; Nikon, Tokyo, Japan) with a 10× objective lens. The number of attached hepatocytes (cells per cm2) was determined by counting the stained nuclei in five images on each surface in triplicate using the analyse particle command of ImageJ software (Bethesda, Maryland, USA).24Inhibitory attachment to HB-EGF-bound temperature-responsive surfaces
Inhibition of the attachment of hepatocytes with soluble EGF was examined on HB-EGF-bound surfaces. Preparation of heparin-IC1 and PIPAAm surfaces bound with HB-EGF was the same as described in the previous section. The suspension of hepatocytes in high-glucose DMEM containing 2 mg mL−1 BSA, 1–10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 ng mL−1 EGF, 100 U mL−1 penicillin, and 100 μg mL−1 streptomycin was seeded at a density of 5 × 104 cells per cm2 on the surfaces, which were incubated in a 5% CO2 incubator at 20 °C or 37 °C for 1.5 h. Determination of the number of attached hepatocytes (cells per cm2) was the same as that described in the previous section.
000 ng mL−1 EGF, 100 U mL−1 penicillin, and 100 μg mL−1 streptomycin was seeded at a density of 5 × 104 cells per cm2 on the surfaces, which were incubated in a 5% CO2 incubator at 20 °C or 37 °C for 1.5 h. Determination of the number of attached hepatocytes (cells per cm2) was the same as that described in the previous section.
Western blotting
Phosphorylated EGFR stimulated with HB-EGF-bound heparin-IC1 surfaces was analysed using western blotting. Hepatocytes suspended in high-glucose DMEM containing 2 mg mL−1 BSA, 100 U mL−1 penicillin, and 100 μg mL−1 streptomycin were incubated in a 5% CO2 incubator at 37 °C with a seeding density of 1 × 105 cells per cm2 on the heparin-IC1 surfaces that were bound with 100 ng cm−2 HB-EGF on a 60 mm dish. The grafted amount of temperature-responsive polymer on heparin-IC1 surfaces on 60 mm dishes was 1.77 ± 0.01 μg cm−2 (mean ± SD). After incubation for 1.5 h, the surfaces were gently washed with warmed PBS twice on a thermostat plate at 38 °C. Immediately, the attached hepatocytes were lysed on ice with 200 μL NuPAGE™ LDS sample buffer containing 50 mM TCEP, protease inhibitor cocktail, and phosphatase inhibitor cocktails 2 and 3. Hepatocytes (1 × 106 cells), which were centrifuged at 4 °C at 50 × g for 3 min and washed twice with 1 mL PBS, were lysed with 100 μL of NuPAGE™ LDS sample buffer and used as a negative control. Lysis of hepatocytes (1 × 106 cells) incubated with 1 μg HB-EGF in 1 mL medium at 37 °C for 15 min was used as a positive control. The lysates were passed through a syringe with a 26G needle (Terumo) to cut genomic DNA, centrifuged at 4 °C at 13000 rpm for 10 min, and the supernatants were stored at −80 °C.
Polyacrylamide gel electrophoresis of the lysates was performed using a Novex™ NuPAGE™ SDS-PAGE Gel system (Thermo Fisher Scientific). The concentration of proteins in the lysates was determined using a protein quantification assay (Macherey-Nagel) after denaturation at 70 °C for 10 min. As a standard, Western Marker Neo (high), which is a mixture of blue-stained proteins and proteins with IgG binding sites, was used for both visual confirmation after blotting and chemiluminescence detection. Lysates (20 μg) in triplicate and a standard (2 μL) were separated on a 4–12% gradient NuPAGE™ gel in NuPAGE™ MOPS SDS running buffer at a constant voltage of 200 V for 50 min on ice. Blotting using the XCell II™ Blot Module was carried out in NuPAGE™ Transfer Buffer with NuPAGE™ Antioxidant onto a nitrocellulose membrane at a constant voltage of 30 V for 1 h. Blotted proteins on the membranes were reversibly stained with Novex reversible membrane protein stain kit. Images of stained total proteins on the membrane were captured using the ChemiDoc Touch Imaging System (Bio-Rad Laboratories).
After destaining, the membrane was blocked with 5% non-fat dry milk in Tris-buffered saline (pH 7.4) composed of 20 mM Tris, 500 mM NaCl, 1% casein, and 0.1% Tween 20 (TBST) at room temperature for 1 h, followed by incubation with anti-phospho-EGFR monoclonal rabbit antibody or anti-EGFR polyclonal rabbit antibody (1:1000 diluted in TBST containing 5% BSA) at 4 °C overnight. The membrane was further incubated with horseradish peroxidase-conjugated donkey anti-rabbit antibody (1:1000 diluted in TBST containing 5% BSA) at room temperature for 1 h and treated with the ECL™ Prime Western Blotting System for 5 min. Chemiluminescence from the membranes was immediately captured using a ChemiDoc Touch Imaging System (Bio-Rad Laboratories).
Densitometry of western blots
Densitometry of western blots was performed for comparative analysis of the amounts of phosphorylated EGFR and total EGFR. The signal intensity of the target protein for each sample was determined by using the “lane and bands” command of Image Lab™ software ver. 6.0.1 (Bio-Rad Laboratories). The signal intensity of total proteins on each sample lane was determined by using the “volume tools” command of Image Lab™ software. Signal intensities were normalised by dividing the signal intensity of the target protein by the signal intensity of the total protein. To compare relative protein levels across the samples, the fold change was calculated using the following equation:25
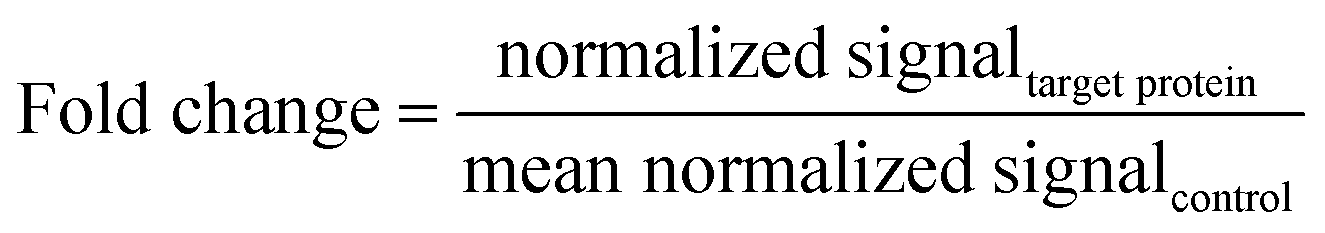 | (1) |
Statistical analyses
All statistical analyses were conducted using RStudio software ver. 1.2.1335 (RStudio Inc., Boston, MA, USA). Comparisons between two groups were carried out using Welch's t test. One-way analysis of variance (ANOVA) was used to compare more than two groups. If the results of one-way ANOVA were significant, multiple comparisons between groups were made using the Tukey honest significant difference (HSD) test. Statistical significance was set at P < 0.05.Results and discussion
Adsorption of HB-EGF on heparin-IC1 and PIPAAm surfaces
The adsorption of HB-EGF on PIPAAm and heparin-IC1 surfaces was quantified using QCM measurements (Fig. 1). To examine the effects of the state of grafted PIPAAm chains, the measurements were conducted with flowing HB-EGF solution at 20 °C in a hydrated state below the LCST and at 37 °C in a dehydrated state above the LCST. Estimated amounts (mean ± SD) of HB-EGF bound on heparin-IC1 surfaces at 20 °C and 37 °C were 177.7 ± 34.6 and 183.0 ± 27.6 ng cm−2, respectively. The amounts on PIPAAm surfaces were estimated to be 161.3 ± 53.7 ng cm−2 at 20 °C and 170.0 ± 56.6 ng cm−2 at 37 °C. ANOVA revealed that the amount of adsorbed HB-EGF was not significantly different between the groups with different surfaces and temperatures. This indicated that the state of the grafted PIPAAm chains and the presence of immobilised heparin on the surface did not affect the adsorption of HB-EGF. | ||
| Fig. 1 The amounts of HB-EGF adsorbed onto PIPAAm and heparin-IC1 surfaces at 20 and 37 °C. The amounts were calculated from QCM measurements using the Sauerbrey equation under flowing 440 ng mL−1 HB-EGF in PBS for 1 h. The bars represent the mean of triplicate experiments. N.S., not statistically significant as determined by one-way ANOVA. | ||
In a previous study, QCM measurements revealed that the adsorption of BSA and fibronectin (molecular weight: 66300 and 440000 g mol−1, respectively) on PIPAAm and heparin-IC1 surfaces was temperature dependent and reduced at temperatures below the LCST. This reduction in protein adsorption was considered to be because of hydration and steric hindrance of the swollen grafted PIPAAm chains.20 In contrast, the amounts of adsorbed HB-EGF were almost the same regardless of the presence of immobilised heparin and the state of grafted PIPAAm chains (Fig. 1), indicating that steric hindrance of the swollen/shrunken PIPAAm chains had little effect on HB-EGF adsorption. Similarly, other groups reported the effects of the size of proteins on the adhesiveness to PIPAAm brush surfaces.26–28 Yu et al. reported that smaller protein lysozyme (14.7 kDa) exhibited adhesion on PIPAAm brushes at 23 °C, whereas the larger proteins, human serum albumin (66.5 kDa) and fibrinogen (341 kDa) weakly adhered to the PIPAAm brushes at the same temperature. When the temperature was increased from 23 °C to 37 °C, the increased rate in the amount of adsorbed protein was decreased as the molecular weight was decreased.26 To explain the effects of the protein size theoretically, three models of protein adsorption behavior on PIPAAm brush surfaces was assumed as follows:29,30 adsorption close to the basal substrate between the collapsed brushes (primary adsorption), adsorption on the outermost layer of the brushes (secondary adsorption), and penetration within the brushes (ternary adsorption). Based on these models, small HB-EGF molecules (predicted molecular weight 9.7 kDa according to the manufacturer's instructions) were presumed to exhibit primary and/or ternary adsorption through diffusing inside the grafted PIPAAm chains, regardless of the presence of swollen/shrunken PIPAAm chains.
Dynamic adsorption/desorption of HB-EGF on heparin-IC1 and PIPAAm surfaces
Fig. 2 shows the monitoring of dynamic changes in Δf during the adsorption of HB-EGF at 37 °C and desorption with a stepwise decrease in the temperature to 20 °C on the heparin-IC1 and PIPAAm surfaces according to the previous report.20 A steep increase in Δf was observed on both surfaces when the temperature was increased from 20 °C to 37 °C. This increase was mainly due to the reduction of viscosity and volumetric mass density of water,31,32 and the release of bound water through dehydration of the grafted PIPAAm chains.20,32 The gentle slope of Δf after the injection of HB-EGF at 40 min was attributed to HB-EGF adsorption on both surfaces. | ||
| Fig. 2 Temperature-dependent Δf for the fifth overtone in PBS to analyse the dynamic process of HB-EGF adsorption on PIPAAm (red) and heparin-IC1 surfaces (blue) during a stepwise temperature change from 20 to 37 °C. Flow rate, 50 μL min−1. | ||
During the temperature change from 37 °C to 20 °C, HB-EGF adsorbed at 37 °C remained on both surfaces. The amount of residual HB-EGF was estimated from Δf between the baselines at 20 °C and the plateau after changing the temperature to 20 °C. The residual HB-EGF on the heparin-IC1 and PIPAAm surfaces was estimated to be 151.0 ± 103.8 and 133.0 ± 28.0 ng cm−2 (mean ± SD) respectively. The difference between the means was not significant (P = 0.80). Therefore, the adsorbed HB-EGF at 37 °C was unable to desorb from both the heparin-IC1 and PIPAAm surfaces even with the swelling of PIPAAm chains with decreasing temperature to 20 °C.
Previous work revealed that cultured hepatocyte sheets on heparin-IC1 surface bound with HB-EGF were detached together with ECMs (e.g., fibronectin) and HB-EGF when the culture temperature was reduced from 37 °C to 20 °C.16 In Fig. 2, however, HB-EGF was found as a residue on the heparin-IC1 surface during dynamic changes in temperature from 37 °C to 20 °C. These results suggest that the release of HB-EGF from the heparin-IC1 surface was accompanied by a low-temperature-induced detachment of hepatocytes. When hepatocytes were incubated on heparin-IC1 surface bound with HB-EGF at 37 °C, EGFRs were accessible on the HB-EGF, which was associated with tethered heparin and compacted PIPAAm chains, resulting in the formation of a complex with EGFR/HB-EGF/heparin.33 Upon lowering the temperature to 20 °C, the complex was considered to be deformed due to the steric hindrance of the swollen grafted PIPAAm chains. In addition, hydration of the PIPAAm chains initiated the detachment of cell sheets, followed by generation of the pulling force through dynamic cytoskeletal contractions within the sheets.34,35 Eventually, HB-EGF was released from the surface, presumably due to its association with EGFR and/or ECMs beneath the cell sheets.
Temperature-dependent attachment of hepatocytes to HB-EGF-bound surfaces
To assess the activity of bound HB-EGF, the attachment of hepatocytes was investigated during short-term incubation. Heparin-IC1 and PIPAAm surfaces bound with HB-EGF were denoted as “HB-EGF/heparin-IC1” and “HB-EGF/PIPAAm,” respectively. Two milligrams per millilitre of BSA in serum-free suspension of hepatocytes was added to quantify the binding between bound HB-EGF and EGFR of hepatocytes under minimising nonspecific binding conditions.36 Fig. 3 shows the percentage of attached hepatocytes after 1.5 h incubation. Microscopic images revealed that the morphology of hepatocytes on HB-EGF/heparin-IC1 and collagen I-coated dish was round-shaped, and actin fibres did not well organised (Fig. S1†). HB-EGF/heparin-IC1 at 37 °C exhibited the highest adhesiveness (6.3 ± 2.9%), which was not significantly different from that in serum-contained DMEM, 8.0%.16 The other surfaces exhibited low adhesiveness (<1%). Almost none of the hepatocytes were attached to HB-EGF/PIPAAm at 37 °C despite the same amount of adsorbed HB-EGF on PIPAAm and heparin-IC1 surfaces, as shown in Fig. 1. This implied that the activity of HB-EGF on heparin-IC1 was preserved due to specific binding between heparin and the heparin-binding domain, leading to activation33 and outward orientation of the EGF-like domain. In contrast, the activity of physisorbed HB-EGF on the surface of PIPAAm was greatly reduced through non-specific adsorption, which resulted in the random orientation of the EGF-like domain and/or denaturation on hydrophobic polystyrene.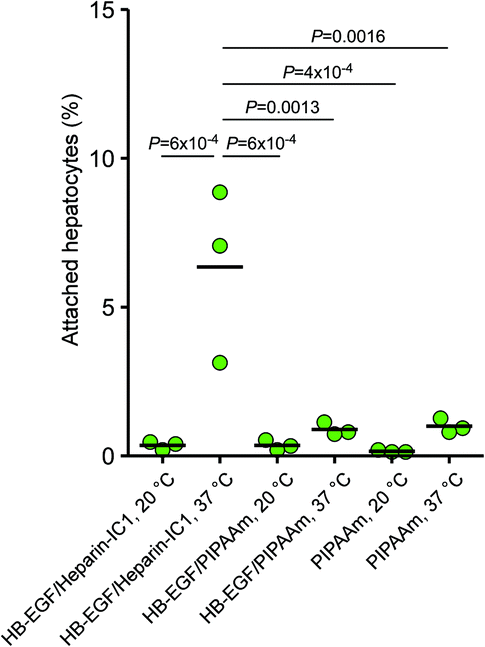 | ||
| Fig. 3 Initial attachment of rat hepatocytes to PIPAAm and heparin-IC1 surfaces after 1.5 h incubation in serum-free DMEM containing 2 mg mL−1 BSA. Seeding density, 5 × 104 cells per cm2. The bars represent the mean of triplicate experiments. P values were determined by one-way ANOVA with Tukey HSD tests. | ||
In addition, hepatocyte attachment was greatly diminished on HB-EGF/heparin-IC1 at 20 °C. It is believed that heparin-modified PIPAAm chains at 20 °C were hydrated and swollen. Thus, it was considered that the attachment of hepatocytes to HB-EGF/heparin-IC1 was attenuated by steric hindrance due to hydration and/or swelling of the grafted PIPAAm chains.
The inhibitory attachment of hepatocytes with soluble EGF was conducted on HB-EGF-immobilized surfaces at 20 °C and 37 °C, as shown in Fig. 4. Almost none of the attached hepatocytes were observed on HB-EGF/PIPAAm, regardless of soluble EGF and/or temperature (percentage of adhered <1%). This indicated that physisorbed HB-EGF on PIPAAm surfaces had no activity toward EGFRs. In contrast, the attachment of hepatocytes to HB-EGF/heparin-IC1 at 37 °C was gradually inhibited by increasing soluble EGF. These data suggested that HB-EGF bound to immobilised heparin held hepatocytes through specific affinity interactions with EGFR. In contrast, at 20 °C, hepatocytes hardly attached to HB-EGF/heparin-IC1 regardless of soluble EGF, indicating that there was no specific affinity between immobilised HB-EGF and EGFR. In the static state, the equilibrated affinity interaction between EGFRs and surface-bound HB-EGF was considered to be attenuated by steric hindrance due to hydration and/or swelling of the grafted PIPAAm chains. This was presumably due to the difficulty of EGFR access due to steric hindrance of the cellular membrane.
 | ||
| Fig. 4 Inhibitory attachment of rat hepatocytes to HB-EGF/heparin-IC1 and HB-EGF/PIPAAm surfaces after 1.5 h incubation with soluble EGF in serum-free DMEM containing 2 mg mL−1 BSA. Seeding density, 5 × 104 cells per cm2. The plots and error bars represent the mean and SD of triplicate experiments. | ||
EGFR stimulation on HB-EGF bound surfaces
The activity of HB-EGF/heparin-IC1 was also investigated by quantifying the stimulation of the EGFRs of hepatocytes. Western blotting for phosphorylated EGFR (pEGFR) and total EGFR was carried out after the cultivation of hepatocytes in serum-free medium on HB-EGF/heparin-IC1 surfaces. Fig. 5 shows western blots for hepatocyte lysates in triplicate with (a) anti-pEGFR and (b) anti-EGFR antibodies. Phosphorylation of EGFR for immobilised HB-EGF on heparin-IC1 surfaces was clearly observed, while that for the negative control was not detected (Fig. 5a). In contrast, residual EGFR on HB-EGF/heparin-IC1 was decreased compared to that of the negative control (Fig. 5b).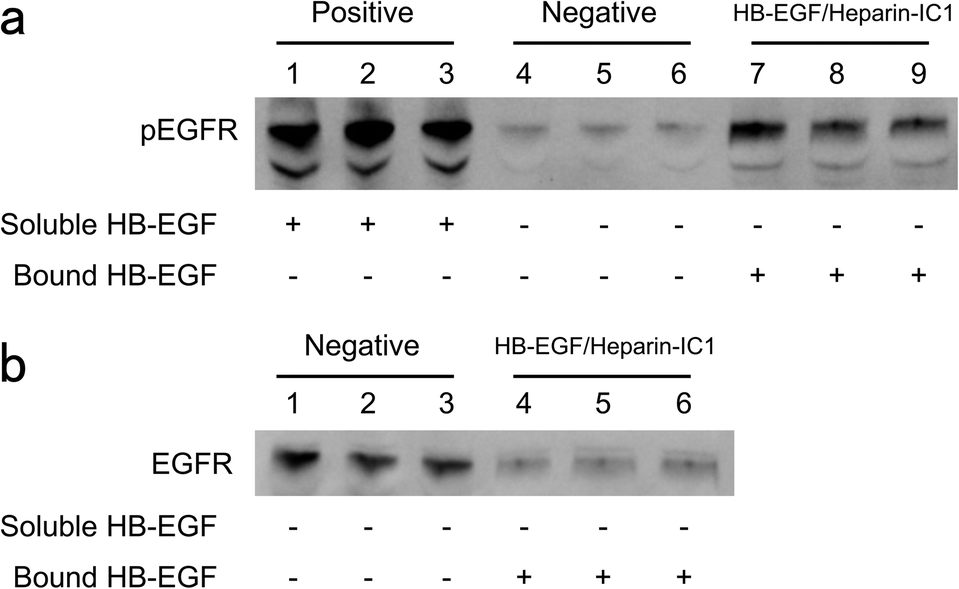 | ||
| Fig. 5 Phosphorylated EGFR of rat hepatocytes cultured on HB-EGF/heparin-IC1 surfaces after 1.5 h incubation in serum-free DMEM containing 2 mg mL−1 BSA. Hepatocyte lysates were analysed in triplicate by western blotting with (a) anti-phospho EGFR and (b) anti-EGFR antibodies. Negative and positive represented the suspension of hepatocytes stimulated without or with 1 μg mL−1 soluble HB-EGF for 15 min at 37 °C, respectively. | ||
Fig. 6 shows the results of densitometric analyses of the western blot membranes in Fig. 5. The values of the fold change in phosphorylated EGFR stimulated with soluble HB-EGF (Fig. 6a) and total EGFR without stimulation (Fig. 6b) were set to 1. The mean fold change in pEGFR on HB-EGF/heparin-IC1 was larger (0.30 ± 0.10) than that of the negative control (0.09 ± 0.01), while the difference between the two means was not significant (P = 0.36). However, the fold change in total EGFR binding with HB-EGF/heparin-IC1 was 0.37 ± 0.06, which was smaller than that of the negative control. This was presumably due to partial internalisation of EGFR through stimulation by HB-EGF/heparin-IC1. Taken together, the activity of immobilised HB-EGF on heparin-IC1 surfaces was preserved.
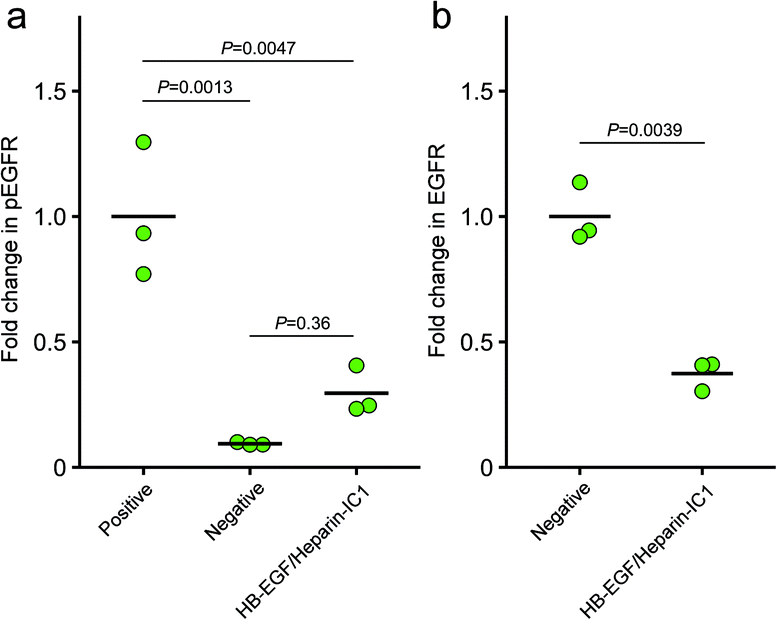 | ||
| Fig. 6 Fold change in phosphorylated EGFR (a) and total EGFR (b). The fold change was calculated from normalized signals by total protein on blotted membranes in Fig. 5. The bars represent the mean of triplicate experiments. P values were determined by one-way ANOVA with Tukey HSD test (a) and Welch's t test (b). | ||
Conclusions
In this study, the temperature-dependent affinity binding of EGFR in hepatocytes with immobilised HB-EGF on heparin-modified, PIPAAm-grafted surfaces was investigated. Hydration and swelling of surface-grafted PIPAAm chains at 20 °C greatly diminished the interaction of EGFR in hepatocytes with HB-EGF/heparin-IC1, while EGFR on hepatocytes was able to bind to HB-EGF/heparin-IC1 with compacted PIPAAm chains at 37 °C. HB-EGF on a heparin-immobilized PIPAAm-grafted surface at 37 °C had the activity to specifically bind to and stimulate EGFR, whereas the physisorbed HB-EGF on the PIPAAm surface greatly diminished their activity. Therefore, the introduction of heparin into grafted PIPAAm chains on the surface plays a pivotal role in preserving the activity of HB-EGF.Author contributions
J. K. conceived the study and designed the experiments. J. K. conducted all experiments and data analyses. J. K. wrote the initial draft of the manuscript. Y. A., N. Y., M. Y., and T. O. critically reviewed and edited the manuscript.Conflicts of interest
T. O. is a founder of CellSeed, Inc. (Tokyo, Japan), which has licenses for certain cell sheet-related technologies and patents from Tokyo Women's Medical University. T. O. and M. Y. are stockholders in CellSeed, Inc. T. O. is the Representative Director of the Organization of Cell Sheet Tissue Engineering and Regenerative Medicine Initiatives (CSTERM), Tokyo, Japan. Tokyo Women's Medical University receives research funds from CellSeed, Inc. and CSTERM.Acknowledgements
Part of this research is based on the Cooperative Research Project of the Research Centre for Biomedical Engineering, Ministry of Education, Culture, Sports, Science, and Technology. This work was supported by JSPS KAKENHI grant numbers JP18H03537 and JP18KK0415.Notes and references
- K. Ohashi, T. Yokoyama, M. Yamato, H. Kuge, H. Kanehiro, M. Tsutsumi, T. Amanuma, H. Iwata, J. Yang, T. Okano and Y. Nakajima, Nat. Med., 2007, 13, 880–885 CrossRef CAS PubMed.
- K. Ohashi and T. Okano, Anat. Rec., 2014, 297, 73–82 CrossRef CAS PubMed.
- K. Tatsumi, M. Sugimoto, D. Lillicrap, M. Shima, K. Ohashi, T. Okano and H. Matsui, PLoS One, 2013, 8, e83280 CrossRef PubMed.
- D. Huang, S. B. Gibeley, C. Xu, Y. Xiao, O. Celik, H. N. Ginsberg and K. W. Leong, Adv. Funct. Mater., 2020, 30, 1909553 CrossRef CAS PubMed.
- S. N. Bhatia, G. H. Underhill, K. S. Zaret and I. J. Fox, Sci. Transl. Med., 2014, 6, 245sr242 Search PubMed.
- S. S. Bale, L. Moore, M. Yarmush and R. Jindal, Tissue Eng., Part B, 2016, 22, 383–394 CrossRef PubMed.
- O. B. Usta, W. J. McCarty, S. Bale, M. Hegde, R. Jindal, A. Bhushan, I. Golberg and M. L. Yarmush, Technology, 2015, 3, 1–26 CrossRef CAS PubMed.
- S. S. Bale, L. Vernetti, N. Senutovitch, R. Jindal, M. Hegde, A. Gough, W. J. McCarty, A. Bakan, A. Bhushan, T. Y. Shun, I. Golberg, R. DeBiasio, B. O. Usta, D. L. Taylor and M. L. Yarmush, Exp. Biol. Med., 2014, 239, 1180–1191 CrossRef PubMed.
- K. Shibuya, M. Watanabe, R. Goto, M. Zaitsu, Y. Ganchiku and A. Taketomi, Cell Transplant., 2021, 30, 9636897211000014 Search PubMed.
- C. T. Nicolas, R. D. Hickey, K. L. Allen, Z. Du, R. M. Guthman, R. A. Kaiser, B. Amiot, A. Bansal, M. K. Pandey, L. Suksanpaisan, T. R. DeGrado, S. L. Nyberg and J. B. Lillegard, Surgery, 2018, 164, 473–481 CrossRef PubMed.
- E. Torok, M. Lutgehetmann, J. Bierwolf, S. Melbeck, J. Dullmann, B. Nashan, P. X. Ma and J. M. Pollok, Liver Transplant., 2011, 17, 104–114 CrossRef PubMed.
- H. Lee, R. A. Cusick, H. Utsunomiya, P. X. Ma, R. Langer and J. P. Vacanti, Tissue Eng., 2003, 9, 1227–1232 CrossRef CAS PubMed.
- N. Yamada, T. Okano, H. Sakai, F. Karikusa, Y. Sawasaki and Y. Sakurai, Makromol. Chem., Rapid Commun., 1990, 11, 571–576 CrossRef CAS.
- T. Okano, N. Yamada, H. Sakai and Y. Sakurai, J. Biomed. Mater. Res., 1993, 27, 1243–1251 CrossRef CAS PubMed.
- Y. Arisaka, J. Kobayashi, M. Yamato, Y. Akiyama and T. Okano, Biomaterials, 2013, 34, 4214–4222 CrossRef CAS PubMed.
- Y. Arisaka, J. Kobayashi, K. Ohashi, K. Tatsumi, K. Kim, Y. Akiyama, M. Yamato and T. Okano, Regener. Ther., 2016, 3, 97–106 CrossRef PubMed.
- K. Kim, K. Ohashi, R. Utoh, K. Kano and T. Okano, Biomaterials, 2012, 33, 1406–1413 CrossRef CAS PubMed.
- P. R. Kuhl and L. G. Griffith-Cima, Nat. Med., 1996, 2, 1022–1027 CrossRef CAS PubMed.
- T. Aoyagi, M. Ebara, K. Sakai, Y. Sakurai and T. Okano, J. Biomater. Sci., Polym. Ed., 2000, 11, 101–110 CrossRef CAS PubMed.
- J. Kobayashi, Y. Arisaka, N. Yui, Y. Akiyama, M. Yamato and T. Okano, Int. J. Mol. Sci., 2018, 19, 1516 CrossRef PubMed.
- P. O. Seglen, in Methods in cell biology, ed. M. P. David, Academic Press, 1976, vol. 13, pp. 29–83 Search PubMed.
- J. Aiken, L. Cima, B. Schloo, D. Mooney, L. Johnson, R. Langer and J. P. Vacanti, J. Pediatr. Surg., 1990, 25, 140–145 CrossRef CAS PubMed.
- T. Mitaka, T. Kojima, T. Mizuguchi and Y. Mochizuki, In Vitro Cell. Dev. Biol.: Anim., 1996, 32, 469–477 CrossRef CAS PubMed.
- C. A. Schneider, W. S. Rasband and K. W. Eliceiri, Nat. Methods, 2012, 9, 671–675 CrossRef CAS PubMed.
- L. Pillai-Kastoori, A. R. Schutz-Geschwender and J. A. Harford, Anal. Biochem., 2020, 593, 113608 CrossRef CAS PubMed.
- Q. Yu, Y. Zhang, H. Chen, Z. Wu, H. Huang and C. Cheng, Colloids Surf., B, 2010, 76, 468–474 CrossRef CAS PubMed.
- C. Xue, N. Yonet-Tanyeri, N. Brouette, M. Sferrazza, P. V. Braun and D. E. Leckband, Langmuir, 2011, 27, 8810–8818 CrossRef CAS PubMed.
- C. Xue, B.-C. Choi, S. Choi, P. V. Braun and D. E. Leckband, Adv. Funct. Mater., 2012, 22, 2394–2401 CrossRef CAS.
- A. Halperin, Langmuir, 1999, 15, 2525–2533 CrossRef CAS.
- A. Halperin and M. Kröger, Macromolecules, 2011, 44, 6986–7005 CrossRef CAS.
- K. K. Kanazawa and J. G. Gordon, Anal. Chem., 1985, 57, 1770–1771 CrossRef CAS.
- G. Z. Zhang, Macromolecules, 2004, 37, 6553–6557 CrossRef CAS.
- R. Takazaki, Y. Shishido, R. Iwamoto and E. Mekada, J. Biol. Chem., 2004, 279, 47335–47343 CrossRef CAS PubMed.
- A. Kushida, M. Yamato, C. Konno, A. Kikuchi, Y. Sakurai and T. Okano, J. Biomed. Mater. Res., 1999, 45, 355–362 CrossRef CAS PubMed.
- M. Yamato, M. Okuhara, F. Karikusa, A. Kikuchi, Y. Sakurai and T. Okano, J. Biomed. Mater. Res., 1999, 44, 44–52 CrossRef CAS PubMed.
- M. D. Pierschbacher and E. Ruoslahti, Proc. Natl. Acad. Sci. U. S. A., 1984, 81, 5985–5988 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/d1ra07317f |
| This journal is © The Royal Society of Chemistry 2021 |