
Response of intestinal signaling communication between the nucleus and peroxisome to nanopolystyrene at a predicted environmental concentration†
Huimin
Shao
,
Yan
Kong
and
Dayong
Wang
 *
*
Key Laboratory of Environmental Medicine Engineering in Ministry of Education, Medical School, Southeast University, Nanjing 210009, China. E-mail: dayongw@seu.edu.cn
First published on 25th November 2019
Abstract
The molecular responses of organisms to environmental toxicants at environmentally relevant concentrations are still largely unknown. Nematode Caenorhabditis elegans is a sensitive animal model used for environmental exposure assessment. We here determined the role of canonical Wnt/β-catenin signaling, a conserved molecular signaling among different organisms, in regulating the response of nematodes to nanopolystyrene (100 nm) at a predicted environmental concentration (1 μg L−1) and the underlying mechanism. Among the components of the canonical Wnt/β-catenin signaling pathway, nanopolystyrene (1 μg L−1) only decreased GSK-3 expression and increased β-catenin BAR-1 expression. GSK-3 acted upstream of BAR-1 to regulate the response to nanopolystyrene, and the intestine-specific activity of the GSK-3-BAR-1 signaling cascade in regulating the response to nanopolystyrene was observed. Transcriptional factor DAF-16 and peroxisomal protein PRX-5 were identified as downstream targets of both BAR-1 and Wnt effector POP-1 in regulating the response to nanopolystyrene. During the control of the response to nanopolystyrene, DAF-16 and PRX-5 functioned synergistically, suggesting that the intestinal canonical Wnt/β-catenin signaling mediates two different molecular signals to regulate the response to nanopolystyrene. In the peroxisome, KAT-1 and ACOX-1.6 were further identified as downstream targets of PRX-5 to regulate the response to nanopolystyrene. Therefore, exposure to nanopolystyrene could activate the canonical Wnt/β-catenin-mediated signaling communication between the nucleus and peroxisome. Our results suggest the crucial function of intestinal canonical Wnt/β-catenin-mediated nucleus–peroxisome signaling communication in response to nanopolystyrene exposure at a predicted environmental concentration.
Environmental significanceThe responses of organisms to nanoplastic particles in different aspects have received great attention. However, the response at the organelle level to nanoplastic particles is still largely unclear. In this study, using animal model Caenorhabditis elegans, we identified an intestinal signaling communication between the nucleus and peroxisome in response to nanopolystyrene at a predicted environmental concentration. Moreover, this signaling communication was activated by the canonical Wnt/β-catenin signaling. Our data demonstrated the crucial function of signaling communication between the nucleus and peroxisome in intestinal cells required for the response of organisms to nanoplastic particles at a predicted environmental concentration. Our results also highlight the crucial roles of the canonical Wnt/β-catenin signaling and intestinal barrier in response to nanoplastic particles at environmentally relevant concentrations. |
Introduction
In recent years, the potential toxicity of nanoplastic particles to organisms has received great attention. Nanopolystyrene can be potentially used in personal care products, drug delivery, biosensors, and bioimaging.1–4 With nanopolystyrene as an example, exposure to nanoplastics has been found to cause adverse effects on different organisms, such as the monogonont rotifer, Daphnia, zebrafish, sea urchins, and crustaceans.5–10 The observed adverse effects from nanopolystyrene exposure include induction of oxidative stress, reduction in growth, suppression in reproduction, a decrease in locomotion behavior, a deficit in development, reduction in lifespan, and a decrease in the feeding rate.5,7–10 The predicted environmentally relevant concentrations of nanoplastics (100 nm) are in the range of ≤1 μg L−1.11 Nevertheless, the response of organisms to nanoplastics at environmentally relevant concentrations and the underlying mechanisms are still largely unclear.Caenorhabditis elegans is a wonderful model animal for evaluating the response to or the toxicity induction of various toxicants or stresses due to its sensitivity to environmental exposure.12–15 Additionally, the thoroughly described molecular and genetic backgrounds make C. elegans more suitable for elucidating the underlying molecular mechanism for the observed response to certain toxicants or stresses.16 Recently, C. elegans has been used to assess the toxicity in various aspects from exposure to microplastics.17–22 It has been further observed that the potential adverse effects of nanopolystyrene at a predicted environmental concentration (1 μg L−1) could be detected in nematodes.23–26 For example, prolonged exposure (from L1-larvae to adult day-1) to nanopolystyrene (1 μg L−1) could cause severe induction of oxidative stress in the acs-22 mutant with the deficit in the function of the intestinal barrier.23
In organisms, canonical Wnt/β-catenin signaling is evolutionarily conserved, and has important functions during their development. Previous studies have suggested that the Wnt signaling pathway can participate in the control of stress response to environmental toxicants, such as graphene oxide.27–29 However, the role of canonical Wnt/β-catenin signaling in regulating the response to nanopolystyrene at predicted environmental concentrations is still unclear. In nematodes, the major components of the canonical Wnt signaling pathway contain β-catenin BAR-1 (a transcriptional factor), the APC complex containing APR-1/Axin, PRY-1/CK1a, KIN-19, and GSK-3, Dishevelled proteins (MIG-5, DSH-1, and DSH-2), and Frizzled receptors (LIN-17, MOM-5, MIG-1, and CFZ-2).30 With nanopolystyrene (100 nm) as an example, we have identified some molecular signaling pathways (such as insulin and p38 mitogen-activated protein kinase (MAPK) signaling pathways) required for the control of response to nanoplastic particles in nematodes.20,26,31 In this study, we further investigated the role of the canonical Wnt/β-catenin signaling pathway in regulating the response of nematodes to nanopolystyrene (100 nm) at a predicted environmental concentration (1 μg L−1). Moreover, we examined the underlying mechanism for the function of the canonical Wnt/β-catenin signaling pathway in regulating the response to nanopolystyrene. Our results suggested the crucial function of intestinal canonical Wnt/β-catenin signaling in regulating the response to nanopolystyrene at a predicted environmental concentration by affecting the signaling communication between the nucleus and peroxisome.
Experimental
Nanopolystyrene characterization
Nanopolystyrene without dye labeling was from Janus New-Materials Co. (Nanjing, China). Based on analysis by transmission electron microscopy (TEM), the size of the used nanopolystyrene was 102.8 ± 4.5 nm (Fig. 1a). The size of nanopolystyrene particles was further confirmed by dynamic light scattering (DLS) using a Nano Zetasizer (Malvern Instrument Ltd., Malvern, UK), and the detected size of nanopolystyrene was 103.5 ± 4.8 nm. The Raman spectrum of nanopolystyrene is shown in Fig. 1b. The zeta potential of the used nanopolystyrene was −9.213 ± 0.478 mV. The predicted environmentally relevant concentration of nanoplastic particles (100 nm) was selected as 1 μg L−1.11,32 Working solutions of nanopolystyrene were prepared by diluting a stock solution (1 mg mL−1) with K medium. Before use, the particle solutions were sonicated for 30 min (40 kHz, 100 W). Because we only detected the obvious toxicity of leachates from nanopolystyrene (10 mg L−1) (Fig. S1†), the observed toxicity of nanopolystyrene in the range of μg L−1 might not be due to the leachates from nanopolystyrene.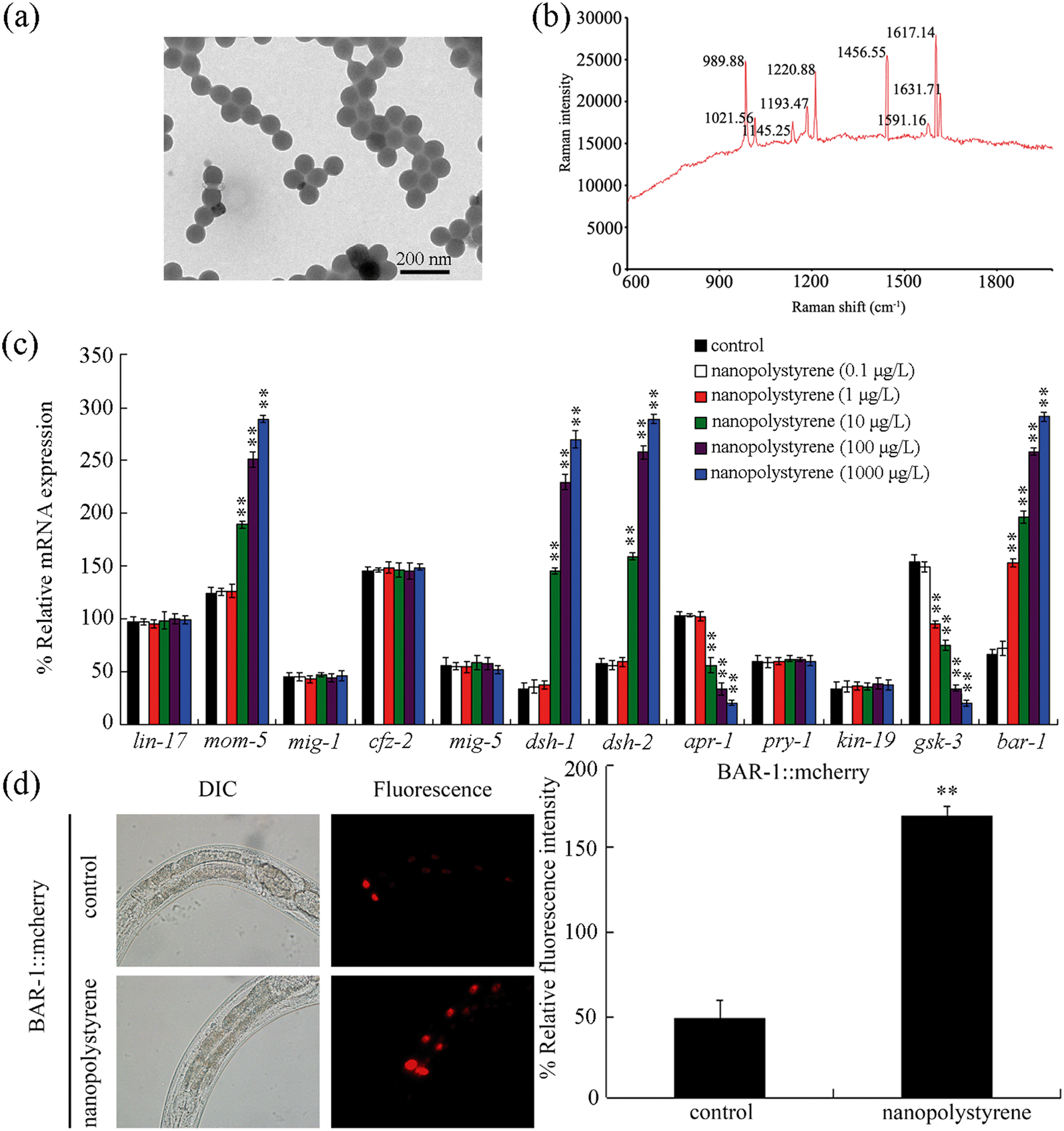 | ||
| Fig. 1 Effect of nanopolystyrene on expressions of genes encoding the canonical Wnt/β-catenin signaling pathway. (a) TEM image of nanopolystyrene in K medium. (b) Raman spectrum of nanopolystyrene. (c) Effect of nanopolystyrene on transcriptional expressions of genes encoding the canonical Wnt/β-catenin signaling pathway. Control, without nanopolystyrene exposure. Bars represent means ± SD. **P < 0.01 vs. control. (d) Effect of nanopolystyrene on expression of intestinal BAR-1::mcherry. Control, without nanopolystyrene exposure. Exposure to nanopolystyrene was performed from L1-larvae to adult day-3. Exposure concentration of nanopolystyrene was 1 μg L−1. Bars represent means ± SD. **P < 0.01 vs. control. | ||
Animal strains and maintenance
The used nematode strains contain wild-type N2, mutants bar-1(ga80), gsk-3(tm1020), and rrf-1(pk1417), and transgenic strains thuIs751[BAR-1::mcherry],25 CF1553/muIs84[SOD-3::GFP], VP303/kbIs7[nhx-2p::rde-1], NR222/rde-1(ne219);kzIs9, TU3401/sid-1(pk3321);[pCFJ90(myo-2p::mCherry)+unc-119p::sid-1], and WM118/rde-1(ne300);neIs9, zIs356[Pdaf-16::daf-16a/b::GFP]. The thuIs751 strain is a gift from Dr. Xiao Liu's lab. The other strains were purchased from the Caenorhabditis Genetic Center. VP303 is a genetic tool for intestine-specific RNA interference (RNAi) knockdown of certain gene(s),33 and NR222, TU3401, WM118, or rrf-1(pk1417) is used for epidermal, neuronal, muscle, or germline RNAi knockdown of certain gene(s).34–37 Animals were normally maintained on nematode growth medium (NGM) plates at 20 °C with Escherichia coli OP50 as the food source.38 The used mutants were out-crossed at least four times. The collected gravid animals were first lysed using a bleaching mixture solution (0.45 M NaOH, 2% HOCl). After that, the released eggs were used to prepare age synchronous L1- or L2-larvae.Exposure and toxicity assessment
The nanopolystyrene solutions (1–10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 μg L−1) were mixed with OP50 (∼4 × 106 colony-forming units (CFUs)). The age synchronous L1 larvae were transferred into these prepared nanopolystyrene solutions. The exposure to nanopolystyrene was performed from L1-larvae to adult day-3. During the exposure, the nanoparticle solutions were refreshed daily. That is, the exposed nematodes were transferred into new nanopolystyrene solutions with OP50 added daily. After the exposure, the nematodes were washed with M9 buffer three times. After that, the nematodes were used for toxicity assessment using different endpoints.
000 μg L−1) were mixed with OP50 (∼4 × 106 colony-forming units (CFUs)). The age synchronous L1 larvae were transferred into these prepared nanopolystyrene solutions. The exposure to nanopolystyrene was performed from L1-larvae to adult day-3. During the exposure, the nanoparticle solutions were refreshed daily. That is, the exposed nematodes were transferred into new nanopolystyrene solutions with OP50 added daily. After the exposure, the nematodes were washed with M9 buffer three times. After that, the nematodes were used for toxicity assessment using different endpoints.
Reactive oxygen species (ROS) production was used to reflect the activation of oxidative stress.13,39 The ROS production was determined as described.40 The examined nematodes were labeled with 1 μM 5′,6′-chloromethyl-2′,7′-dichlorodihydro-fluorescein diacetate (CM-H2DCFDA) for 3 h in the dark. After that, the nematodes were washed with M9 buffer three times. The nematodes were mounted on a 2% agar pad. Fluorescence images were observed under a laser scanning confocal microscope (excitation wavelength, 488 nm; emission filter, 510 nm). The intestinal ROS signals were examined by relative fluorescence units and normalized to the autofluorescence. The nematodes not labeled with CM-H2DCFDA were used as a negative control, and only very weak autofluorescence could be observed under such a condition. For each treatment, fifty animals were examined.
Locomotion behavior reflects the functional state of motor neurons of animals. The observed nanopolystyrene toxicity in decreasing locomotion behavior was largely due to the activation of oxidative stress by nanopolystyrene exposure, because treatment with an antioxidant (10 mM ascorbate) could significantly suppress the nanopolystyrene toxicity in decreasing locomotion behavior (Fig. S2†). Head thrash and body bend were selected to assess the locomotion behavior as described.41 After the exposure, the nematodes were first washed with M9 buffer three times. After that, the nematodes were transferred onto the surface of a NGM plate without feeding of OP50. The nematodes were left on this NGM plate for 1 min before the counting of locomotion behavior under a dissecting microscope. A change in the bending direction at the body mid-region of nematodes is considered as a head thrash, and the frequency in a 1 min period was counted. A change in the posterior bulb direction is considered as a body bend, and the frequency in a 20 s period was counted. The locomotion behavior under normal conditions without nanopolystyrene exposure was used as the control. For each treatment, thirty animals were examined.
Brood size reflecting the reproductive capacity of nematodes was determined as described.42 After nanopolystyrene exposure, the nematodes were first washed with M9 buffer three times. The brood size refers to the number of offspring at all stages beyond the egg.43 For each treatment, ten animals were examined.
Quantitative real-time polymerase chain reaction (qRT-PCR)
The total RNA of animals was prepared using Trizol (Invitrogen, UK) according to manufacturer's protocol. The concentration and purity of the prepared RNAs were evaluated using a spectrophotometer based on a ratio of OD260/280. The cDNA synthesis was performed in a 12.5 μL reaction volume containing 625 ng total RNA, 0.5 mM reverse-transcript primers, 3 mM MgCl2, 75 mM KCl, 50 mM Tris–HCl, 10 mM dithiothreitol, 20 units ribonuclease inhibitor, and 100 U reverse transcriptase (Takara, China). After the cDNA synthesis, gene expressions were analyzed in an ABI 7500 real-time PCR system with Evagreen (Biotium, USA). The relative transcriptional expression ratio between the examined genes and reference gene (tba-1 encoding alpha-tubulin protein) was determined. The biological reactions were examined in triplicate. The related primer information is provided in Table S1.†RNAi
The E. coli HT115 strains expressing double-stranded RNA corresponding to certain gene(s) were from the ORFeome-RNAi v1.1 library. E. coli HT115 (DE3) grown in LB broth containing ampicillin (100 mg mL−1) at 37 °C overnight was plated onto NGM plates containing 5 mM isopropyl 1-thio-β-D-galactopyranoside (IPTG) and 100 mg mL−1 ampicillin. To perform the RNAi, the nematodes were fed with E. coli HT115 strains expressing double-stranded RNA corresponding to certain gene(s) on NGM plates as described.44 The prepared L1-larvae (L2-larvae for elo-5(RNAi)) were grown on RNAi or vector control plates. When they developed into gravid at 20 °C, the adult nematodes were transferred onto a fresh NGM plate to let them lay eggs in order to obtain the second generation for the toxicity assessment. HT115 bacteria harboring empty vector L4440 containing two T7 promoters flanking a polylinker were used as a negative control. RNAi efficiency was confirmed by qRT-PCR (data not shown).Statistical analysis
Statistical analysis was performed using SPSS 12.0 (SPSS Inc., Chicago, USA). Differences between groups were determined using one-way analysis of variance (ANOVA). For the multiple factor comparison, two-way ANOVA analysis was performed. The probability level of 0.01 (**) was considered to be statistically significant.Results
Response of canonical Wnt/β-catenin signaling to nanopolystyrene at predicted environmental concentration
After exposure, nanopolystyrene (0.1 μg L−1) did not obviously alter expressions of all the examined genes (Fig. 1c). Exposure to nanopolystyrene (1 μg L−1) also did not significantly affect the expressions of genes (lin-17, mom-5, mig-1, and cfz-2) encoding Frizzled receptors and genes (mig-5, dsh-1, and dsh-2) encoding Dishevelled proteins (Fig. 1c). Among the genes encoding the APX complex, nanopolystyrene (1 μg L−1) also did not significantly alter the expressions of apr-1, pry-1 and kin-19 (Fig. 1c). Different from these, nanopolystyrene (1 μg L−1) significantly decreased the expression of the gsk-3 gene encoding an ortholog of human GSK3A, a component of the APC complex, and increased the expression of the bar-1 gene encoding the canonical β-catenin (Fig. 1c). Moreover, nanopolystyrene (1 μg L−1) significantly increased the expression of intestinal BAR-1::mcherry (Fig. 1d).Exposure to nanopolystyrene (10–1000 μg L−1) did not affect the expressions of lin-17, mig-1, cfz-2, mig-5, pry-1, and kin-19 (Fig. 1c). In contrast, the expressions of mom-5, dsh-1, and dsh-2 could be increased by nanopolystyrene (10–1000 μg L−1) (Fig. 1c). Expressions of apr-1 and gsk-3 could also be decreased by nanopolystyrene (10–1000 μg L−1) (Fig. 1c).
Mutation of bar-1 induced a susceptibility to nanopolystyrene toxicity
We next employed ROS production and locomotion behavior as endpoints to examine the effect of bar-1 mutation on toxicity induction of nanopolystyrene. Nanopolystyrene exposure induced the significant ROS production and the decrease in locomotion behaviors of head thrash and body bend (Fig. 2). In nanopolystyrene exposed bar-1 mutant nematodes, we observed the more severe induction of ROS production and decrease in locomotion behaviors of head thrash and body bend compared with those in nanopolystyrene exposed wild-type nematodes (Fig. 2). That is, compared with toxicity induction of nanopolystyrene in wild-type nematodes, we observed that a more severe toxicity of nanopolystyrene was formed in bar-1 mutant nematodes. Therefore, bar-1 mutation potentially induces a susceptibility to nanopolystyrene toxicity. | ||
| Fig. 2 Mutation of bar-1 induced a susceptibility to the nanopolystyrene toxicity in inducing intestinal ROS production (a) and in decreasing locomotion behavior (b). Control, without nanopolystyrene exposure. Exposure concentration of nanopolystyrene was 1 μg L−1. Exposure to nanopolystyrene was performed from L1-larvae to adult day-3. Bars represent means ± SD. **P < 0.01 vs. control (if not specifically indicated). | ||
Mutation of gsk-3 induced a resistance to nanopolystyrene toxicity
We further used ROS production and locomotion behavior as endpoints to analyze the effect of gsk-3 mutation on toxicity induction of nanopolystyrene. In nanopolystyrene exposed gsk-3 mutant nematodes, we observed the suppression of ROS production and the increase in locomotion behaviors of head thrash and body bend compared with those in nanopolystyrene exposed wild-type nematodes (Fig. 3a and b). That is, compared with toxicity induction of nanopolystyrene in wild-type nematodes, the toxicity of nanopolystyrene could be obviously suppressed in gsk-3 mutant nematodes. Therefore, different from the bar-1 mutant, the gsk-3 mutation potentially induces a resistance to nanopolystyrene toxicity.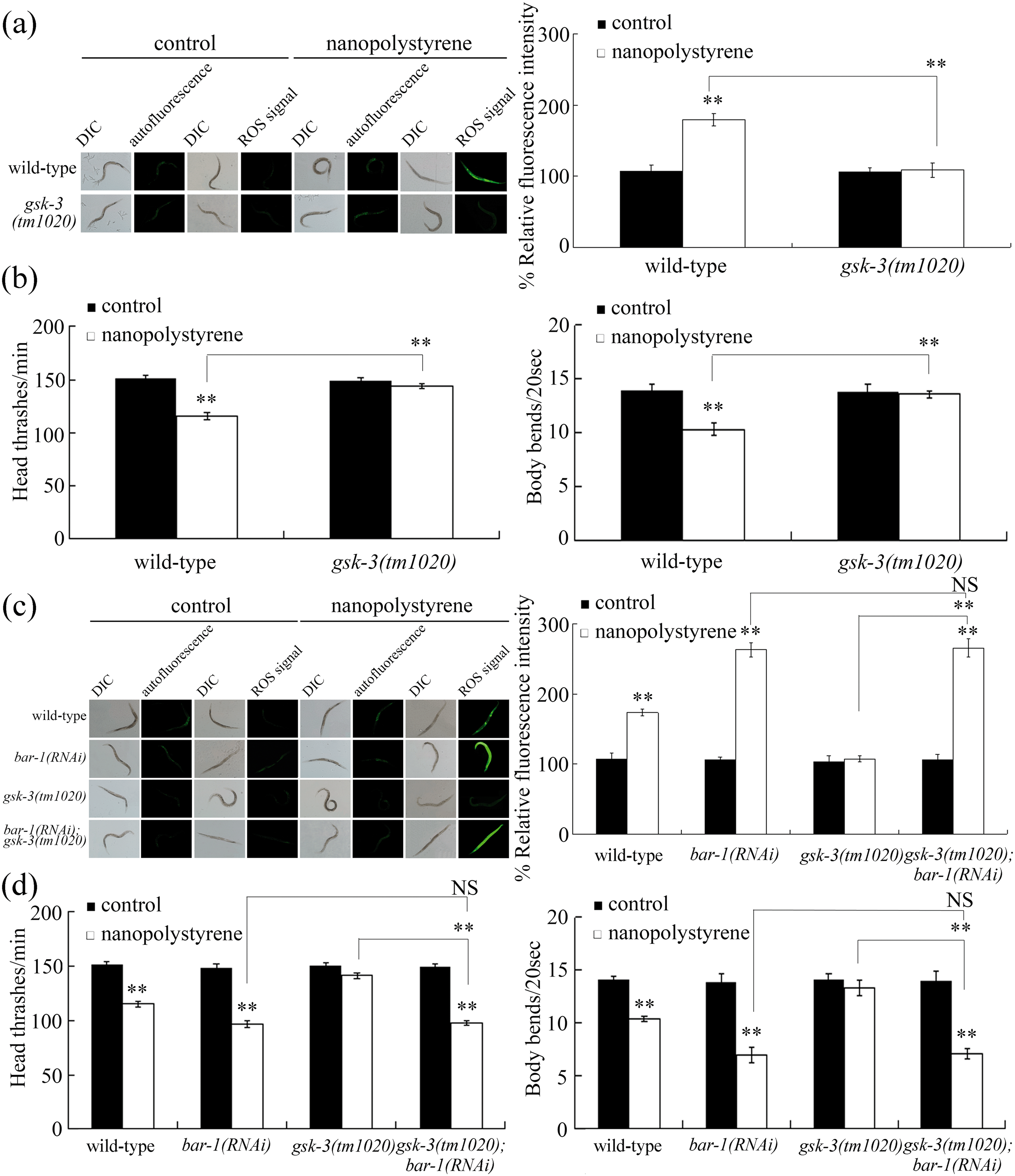 | ||
| Fig. 3 Genetic interaction between BAR-1 and GSK-3 in regulating the response to nanopolystyrene. (a) Mutation of gsk-3 induced a resistance to the nanopolystyrene toxicity in inducing intestinal ROS production. (b) Mutation of gsk-3 induced a resistance to the nanopolystyrene toxicity in decreasing locomotion behavior. (c) Genetic interaction between BAR-1 and GSK-3 in regulating the nanopolystyrene toxicity in inducing intestinal ROS production. (d) Genetic interaction between BAR-1 and GSK-3 in regulating the nanopolystyrene toxicity in decreasing locomotion behavior. Control, without nanopolystyrene exposure. Exposure concentration of nanopolystyrene was 1 μg L−1. Exposure to nanopolystyrene was performed from L1-larvae to adult day-3. Bars represent means ± SD. **P < 0.01 vs. control (if not specifically indicated). NS, no significance. | ||
Genetic interaction between BAR-1 and GSK-3 in regulating the nanopolystyrene toxicity
Using the strain gsk-3(tm1020);bar-1(RNAi), we observed that the RNAi knockdown of bar-1 significantly induced the ROS production and decreased the locomotion behaviors of head thrash and body bend in nanopolystyrene exposed gsk-3 mutant nematodes (Fig. 3c and d). That is, the RNAi knockdown of bar-1 could effectively suppress the resistance of gsk-3(tm1020) mutant nematodes to the nanopolystyrene toxicity in inducing ROS production and in decreasing locomotion behavior. Therefore, genetic interaction analysis demonstrated that GSK-3 acted upstream of BAR-1 to regulate the response of nematodes to nanopolystyrene.Tissue-specific activity of BAR-1 in regulating the response to nanopolystyrene
BAR-1 is expressed in many tissues, including the intestine (https://www.wormbase.org). We further determined the effects of the RNAi knockdown of bar-1 in different tissues on toxicity induction of nanopolystyrene. In nanopolystyrene exposed nematodes with the RNAi knockdown of bar-1 in the epidermis, neurons, muscle, or germline, we observed similar ROS production to that in nanopolystyrene exposed NR222, TU3401, WM118, or rrf-1 nematodes (Fig. 4a). That is, the RNAi knockdown of bar-1 in the epidermis, neurons, muscle, or germline did not significantly affect the nanopolystyrene toxicity in inducing intestinal ROS production. In contrast, we observed the more severe ROS production and brood size in nanopolystyrene exposed nematodes with the intestinal RNAi knockdown of bar-1 compared with those in nanopolystyrene exposed VP303 nematodes (Fig. 4a and S3†). That is, the intestinal RNAi knockdown of bar-1 induced a susceptibility to the nanopolystyrene toxicity in inducing intestinal ROS production and in reducing brood size. Therefore, BAR-1 acted in the intestine to regulate the response to nanopolystyrene. | ||
| Fig. 4 Tissue-specific activity of BAR-1 and GSK-3 in regulating the response to nanopolystyrene. (a) Tissue-specific activity of BAR-1 in regulating the response to nanopolystyrene. (b) GSK-3 acted in the intestine to regulate the response to nanopolystyrene. Control, without nanopolystyrene exposure. L4440, empty vector. Exposure concentration of nanopolystyrene was 1 μg L−1. Exposure to nanopolystyrene was performed from L1-larvae to adult day-3. Bars represent means ± SD. **P < 0.01 vs. control (if not specifically indicated). | ||
Using VP303 as a genetic tool, we further found the suppression of ROS production and the increase in brood size in nanopolystyrene exposed gsk-3(RNAi) nematodes compared with those in nanopolystyrene exposed VP303 nematodes (Fig. 4b and S3†). That is, the intestinal RNAi knockdown of gsk-3 further induced a resistance to the nanopolystyrene toxicity in inducing intestinal ROS production and in reducing the brood size. Meanwhile, in the isolated intact intestine, we also observed that exposure to nanopolystyrene (1 μg L−1) significantly decreased the expression of gsk-3 (data not shown).
Identification of downstream targets of intestinal BAR-1 in regulating the response to nanopolystyrene
Some reports have suggested a series of potential targets for BAR-1 in regulating various biological processes,45–54 and some of them can be expressed in the intestine (https://www.wormbase.org). Among these possible intestinal targeted genes, exposure to nanopolystyrene (1 μg L−1) could significantly increase the expressions of pop-1, daf-16, clec-60, clec-52, F53A9.8, sta-2, grd-12, mrp-5, prx-5, and daf-41 (Fig. 5a). In contrast, exposure to nanopolystyrene (1 μg L−1) did not obviously affect the expressions of ceh-13, mab-5, sptf-2, cgt-2, lin-39, egl-5, ilys-3, cnp-2, hmit-1.1, nas-27, dpy-22, lin-7, ogt-1, and pal-1 (Fig. S4†). Moreover, the intestinal RNAi knockdown of bar-1 could significantly decrease the expressions of pop-1, daf-16, and prx-5 in nematodes exposed to nanopolystyrene (1 μg L−1) (Fig. 5b). The increase in the expression of bar-1, pop-1, or prx-5 was concentration-dependent in nanopolystyrene (1–100 μg L−1) exposed nematodes (data not shown). | ||
| Fig. 5 Identification of downstream targets of BAR-1 in regulating the response to nanopolystyrene. (a) Effect of nanopolystyrene exposure on gene expressions. Bars represent means ± SD. **P < 0.01 vs. control. (b) Effect of the intestinal RNAi knockdown of bar-1 on gene expressions in nematodes exposed to nanopolystyrene. Bars represent means ± SD. **P < 0.01 vs. VP303. (c) Effect of the intestinal RNAi knockdown of pop-1 or prx-5 on nanopolystyrene toxicity in inducing intestinal ROS production. Bars represent means ± SD. **P < 0.01 vs. control (if not specifically indicated). (d) Effect of the intestinal RNAi knockdown of pop-1 or daf-16 on gene expressions. Bars represent means ± SD. **P < 0.01 vs. VP303. (e) Genetic interaction between DAF-16 and PRX-5 in regulating the nanopolystyrene toxicity in inducing intestinal ROS production. Bars represent means ± SD. **P < 0.01 vs. control (if not specifically indicated). Control, without nanopolystyrene exposure. Exposure concentration of nanopolystyrene was 1 μg L−1. Exposure to nanopolystyrene was performed from L1-larvae to adult day-3. | ||
daf-16 encoding a FOXO transcriptional factor in the insulin signaling pathway acts in the intestine to regulate the response of nematodes to nanopolystyrene by affecting the functions of its targeted genes (sod-3, mtl-1, and gpd-2).26 Using VP303 as a genetic tool, the intestinal RNAi knockdown of pop-1 or prx-5 also induced a susceptibility to the nanopolystyrene toxicity in inducing intestinal ROS production (Fig. 5c). Therefore, POP-1, DAf-16, and PRX-5 may act as downstream targets for intestinal BAR-1 in regulating the response to nanopolystyrene.
In nematodes, BAR-1 mediates the canonical Wnt signaling by forming a BAR-1/POP-1 physical complex to activate the expression of its downstream target genes.45 After exposure to nanopolystyrene, it was observed that the intestinal RNAi knockdown of pop-1 significantly decreased the expressions of daf-16 and prx-5 (Fig. 5d), which confirms the role of POP-1 in mediating the function of BAR-1 in regulating the stress response. In nematodes, prx-5 encodes an ortholog of human receptor PEX (peroxisomal biogenesis factor).
Genetic interaction between DAF-16 and PRX-5 in regulating the response to nanopolystyrene
After exposure to nanopolystyrene, the intestinal RNAi knockdown of daf-16 did not obviously alter the expression of prx-5 (Fig. 5d), which implies that PRX-5 and DAF-16 may not act in the same genetic pathway to regulate the response to nanopolystyrene. Meanwhile, we observed the more severe induction of intestinal ROS production in nanopolystyrene exposed daf-16(RNAi);prx-5(RNAi) nematodes compared with that in nanopolystyrene exposed daf-16(RNAi) or prx-5(RNAi) nematodes (Fig. 5e), which suggests that DAF-16 and PRX-5 functioned synergistically to regulate the response to nanopolystyrene.Identification of downstream targets of intestinal PRX-5 in regulating the response to nanopolystyrene
Some reports have also raised some potential targets for PRX-5 in regulating various biological processes,55,56 and at least four of them (ACS-1, PNK-1, ELO-5, and ELO-6) can be expressed in the intestine (https://www.wormbase.org). Receptor PEX5, an ortholog of PRX-5 can interact directly with proteins containing type 1 peroxisomal targeting signal (PTS1) in humans.57 In nematodes, peroxisomal PTS1 proteins contain KAT-1, T02G5.7, ACOX-1.1, ACOX-1.2, ACOX-1.3, ACOX-1.4, ACOX-1.5, ACOX-1.6, and ACOX-3.58Among these possible targeted genes, nanopolystyrene (1 μg L−1) could only significantly decrease the expressions of kat-1 and acox-1.6 (Fig. 6a). Both KAT-1 and ACOX-1.6 can be expressed in the intestine (https://www.wormbase.org). Moreover, the intestinal RNAi knockdown of prx-5 could significantly increase the expression of kat-1 and acox-1.6 in nematodes exposed to nanopolystyrene (1 μg L−1) (Fig. 6b). Meanwhile, the intestinal RNAi knockdown of prx-5 could not obviously affect the expression of daf-16 (Fig. 6b).
 | ||
| Fig. 6 KAT-1 and ACOX-1.6 acted downstream of PRX-5 to regulate the response to nanopolystyrene. (a) Effect of nanopolystyrene exposure on gene expressions. Control, without nanopolystyrene exposure. Exposure concentration of nanopolystyrene was 1 μg L−1. Exposure to nanopolystyrene was performed from L1-larvae to adult day-3. Bars represent means ± SD. **P < 0.01 vs. control. (b) Effect of the intestinal RNAi knockdown of prx-5 on gene expressions in nematodes exposed to nanopolystyrene. Exposure concentration of nanopolystyrene was 1 μg L−1. Exposure to nanopolystyrene was performed from L1-larvae to adult day-3. Bars represent means ± SD. **P < 0.01 vs. VP303. (c) Genetic interaction between PRX-5 and KAT-1 or ACOX-1.6 in regulating the nanopolystyrene toxicity in inducing intestinal ROS production. Control, without nanopolystyrene exposure. Exposure concentration of nanopolystyrene was 1 μg L−1. Exposure to nanopolystyrene was performed from L1-larvae to adult day-3. Bars represent means ± SD. **P < 0.01 vs. control (if not specifically indicated). (d) A diagram showing the molecular basis for intestinal canonical Wnt/β-catenin signaling in regulating the response to nanopolystyrene at a predicted environmental concentration. | ||
Using VP303 as a genetic tool, we further observed that the intestinal RNAi knockdown of kat-1 or acox-1.6 induced a resistance to the nanopolystyrene toxicity in inducing intestinal ROS production (Fig. 6c), suggesting that KAT-1 and ACOX-1.6 may act as potential downstream targets for intestinal PRX-5 in regulating the response to nanopolystyrene.
Genetic interaction between PRX-5 and KAT-1 or ACOX-1.6 in regulating the response to nanopolystyrene
We found that the intestinal RNAi knockdown of kat-1 or acox-1.6 could effectively suppress the susceptibility of prx-5(RNAi) nematodes to the nanopolystyrene toxicity in inducing intestinal ROS production (Fig. 6c). That is, KAT-1 and ACOX-1.6 further acted downstream of intestinal PRX-5 in the peroxisome to regulate the response to nanopolystyrene.Discussion
The canonical Wnt/β-catenin signaling can control the response to various environmental toxicants in nematodes.14 For example, a signaling cascade of MOM-5/CFZ-2-DSH-1/DSH-2-APR-1/GSK-3-BAR-1 in the canonical Wnt/β-catenin signaling pathway has been shown to be involved in the regulation of response to graphene oxide (1 mg L−1).28 Different from this, exposure to nanopolystyrene (1 μg L−1) only significantly altered the expressions of gsk-3, bar-1, and pop-1 (Fig. 1c, d and 5a), suggesting that a signaling cascade of GSK-3-BAR-1-POP-1 in the canonical Wnt/β-catenin signaling pathway was required for the regulation of response to nanopolystyrene (1 μg L−1). Therefore, only a brief signaling cascade in the canonical Wnt/β-catenin signaling pathway can be induced to regulate the response to nanopolystyrene at a predicted environmental concentration.In nematodes, mutation of bar-1 or pop-1 induced a susceptibility to the nanopolystyrene toxicity (Fig. 2 and 5c), and mutation of gsk-3 induced a resistance to the nanopolystyrene toxicity (Fig. 3a and b). Meanwhile, the RNAi knockdown of bar-1 could suppress the resistance of gsk-3 mutant nematodes to the nanopolystyrene toxicity (Fig. 3c and d). These observations demonstrated that the altered expression of the GSK-3-BAR-1-POP-1 signaling cascade mediated a protective function in nematodes exposed to nanopolystyrene. That is, the decreased expression of GSK-3 may mediate a protective function in nematodes exposed to nanopolystyrene by increasing the expressions and the function of BAR-1 and POP-1.
Previous studies have suggested that the TGF-β signaling pathway and let-7 signaling pathway, two important development-related signaling pathways, can function in the intestine to regulate the stress response in nematodes.59,60 In this study, the tissue-specific activity analysis indicated that the signaling cascade of GSK-3-BAR-1 could further act in the intestine to regulate the response to nanopolystyrene (Fig. 4 and S3†). The canonical Wnt/β-catenin signaling pathway is a novel identified development-related signaling pathway with the intestinal function in regulating the stress response in nematodes. Insulin and p38 MAPK signaling pathways were recently identified to act in the intestine to regulate the response to nanopolystyrene.26,31 Here, we identified the canonical Wnt/β-catenin signaling pathway as another intestinal signaling required for the regulation of response to nanopolystyrene.
It has been shown that the physical interaction between BAR-1 and POP-1 exists in the cells of nematodes.45 Our data further identified the FOXO transcriptional factor DAF-16 in the insulin signaling pathway and PRX-5 as the downstream targets of BAR-1/POP-1 complex in the intestine to regulate the response to nanopolystyrene (Fig. 5a–d). That is, the BAR-1-POP-1 signaling cascade in the intestinal nucleus could regulate the response to nanopolystyrene by affecting the activity of another transcriptional factor DAF-16. The functional interaction between the β-catenin and the FOXO transcriptional factor DAF-16 was also found to be required for the control of oxidative stress.48
In nematodes, PRX-5 is an important peroxisomal protein. Peroxisomal proteins KAT-1 and ACOX-1.6 were further identified as the downstream targets of PRX-5 in the regulation of response to nanopolystyrene (Fig. 6a–c). Therefore, in the peroxisome, the signaling cascade of PRX-5-KAT-1/ACOX-1.6 was formed to be required for the regulation of response to nanopolystyrene. In nematodes, KAT-1 is a peroxisomal oxidation enzyme governing the fatty acid oxidation,61 implying the involvement of fatty acid metabolism related signaling in regulating the response to nanopolystyrene. So far, the exact biological function(s) of ACOX-1.6 is still unclear.
Moreover, we observed a synergistic interaction between DAF-16 and PRX-5 in the regulation of response to nanopolystyrene (Fig. 5e). This implies that, in the intestinal cells, the BAR-1-POP-1 signaling cascade may activate two different molecular signals to regulate the response to nanopolystyrene (Fig. 6d). One is mediated by the nuclear protein DAF-16 and its several targets (SOD-3, MTL-1, and GPD-2).26 The RNAi knockdown of bar-1, pop-1, or daf-16 could suppress the induction of SOD-3::GFP, whereas the RNAi knockdown of gsk-3 could increase the SOD-3::GFP expression (Fig. S5†). Another one is the PRX-5-mediated peroxisomal signaling. SOD-3 is a mitochondrial Mn-SOD protein, which implies that the canonical Wnt/β-catenin signaling may also mediate the signaling communication between the nucleus and mitochondria in nanopolystyrene exposed nematodes.
In this study, exposure to nanopolystyrene (1 μg L−1) did not affect the expressions of genes encoding Frizzled receptors (Fig. 1c), suggesting that the activation of Wnt/β-catenin signaling was not through the Frizzled receptors under the conditions of nanopolystyrene (1 μg L−1) exposure. Moreover, we found that mutation of daf-2 encoding the insulin receptor in the insulin signaling pathway could not alter both the gsk-3 expression and the bar-1 expression in nematodes exposed to nanopolystyrene (1 μg L−1) (data not shown), suggesting that the activation of Wnt/β-catenin signaling was also not through the insulin receptor. Therefore, the exact function of G protein coupled receptors (GPCRs) to mediate the activation of canonical Wnt/β-catenin signaling in nematodes exposed to nanopolystyrene at predicted environmental concentrations is still unclear.
Conclusions
Together, we determined the important role of canonical Wnt/β-catenin signaling in regulating the response to nanopolystyrene at a predicted environmental concentration and the underlying mechanisms in nematodes. The dysregulated canonical Wnt/β-catenin signaling induced by nanopolystyrene at a predicted environmental concentration mediated a protective response. Canonical Wnt/β-catenin signaling acted in the intestine to regulate the response to nanopolystyrene, and an intestinal cascade of GSK-3-BAR-1-POP-1 was identified for the canonical Wnt/β-catenin signaling in response to nanopolystyrene. During the control of response to nanopolystyrene, transcriptional factor DAF-16 and peroxisomal protein PRX-5 were identified as downstream targets of intestinal BAR-1 and POP-1. DAF-16 and PRX-5 functioned synergistically, which suggests that intestinal canonical Wnt/β-catenin signaling regulated the response to nanopolystyrene by activating two different molecular signals. In nematodes, nanopolystyrene exposure could activate the canonical Wnt/β-catenin-mediated signaling communication between the nucleus and peroxisome. Our data highlight the crucial function of intestinal canonical Wnt/β-catenin signaling in modulating the response to nanopolystyrene at a predicted environmental concentration in nematodes.Ethical statement
All animal procedures were performed in accordance with the Guidelines for Care and Use of Laboratory Animals of Southeast University and approved by the Animal Care & Welfare Committee of Southeast University.Conflicts of interest
There are no conflicts of interests.References
- B. Han, W. Liu, J. Li, J. Wang, D. Zhao, R. Xu and Z. Lin, Water Res., 2017, 120, 199–210 CrossRef CAS PubMed.
- T. Wang, L. Wang, X. Li, X. Hu, Y. Han, Y. Luo, Z. Wang, Q. Li, A. Aldalbahi, L. Wang, S. Song, C. Fan, Y. Zhao, M. Wang and N. Chen, ACS Appl. Mater. Interfaces, 2017, 22, 18619–18625 CrossRef PubMed.
- M. Cooley, A. Sarode, M. Hoore, D. A. Fedosov, S. Mitragotri and A. Sen Gupta, Nanoscale, 2018, 32, 15350–15350 RSC.
- L. Zheng, G. Cai, S. Wang, M. Liao, Y. Li and J. Lin, Biosens. Bioelectron., 2019, 124–125, 143–149 CrossRef CAS PubMed.
- C. Della Torre, E. Bergami, A. Salvati, C. Faleri, P. Cirino, K. A. Dawson and I. Corsi, Environ. Sci. Technol., 2014, 48, 12302–12311 CrossRef CAS PubMed.
- C. Jeong, E. Won, H. Kang, M. Lee, D. Hwang, U. Hwang, B. Zhou, S. Souissi, S. Lee and J. Lee, Environ. Sci. Technol., 2016, 50, 8849–8857 CrossRef CAS PubMed.
- Y. Ma, A. Huang, S. Cao, F. Sun, L. Wang, H. Guo and R. Ji, Environ. Pollut., 2016, 219, 166–173 CrossRef CAS PubMed.
- S. Rist, A. Baun and N. B. Hartmann, Environ. Pollut., 2017, 228, 398–407 CrossRef CAS PubMed.
- C. Gambardella, S. Morgana, S. Ferrando, M. Bramini, V. Piazza, E. Costa, F. Garaventa and M. Faimali, Ecotoxicol. Environ. Saf., 2017, 145, 250–257 CrossRef CAS PubMed.
- Q. Chen, M. Gundlach, S. Yang, J. Jiang, M. Velki, D. Yin and H. Hollert, Sci. Total Environ., 2017, 584–585, 1022–1031 CrossRef CAS PubMed.
- R. Lenz, K. Enders and T. G. Nielsen, Proc. Natl. Acad. Sci. U. S. A., 2016, 113, E4121–E4122 CrossRef CAS PubMed.
- M. C. K. Leung, P. L. Williams, A. Benedetto, C. Au, K. J. Helmcke, M. Aschner and J. N. Meyer, Toxicol. Sci., 2008, 106, 5–28 CrossRef CAS PubMed.
- D.-Y. Wang, Nanotoxicology in Caenorhabditis elegans, Springer Nature Singapore Pte Ltd., 2018 Search PubMed.
- Y.-L. Zhao, H. Chen, Y.-H. Yang, Q.-L. Wu and D. Y. Wang, Sci. Total Environ., 2019 DOI:10.1016/j.scitotenv.2019.134492.
- H.-L. Liu, D.-Q. Guo, Y. Kong, Q. Rui and D.-Y. Wang, Ecotoxicol. Environ. Saf., 2019 DOI:10.1016/j.ecoenv.2019.109554.
- D.-Y. Wang, Molecular Toxicology in Caenorhabditis elegans, Springer Nature Singapore Pte Ltd., 2019 Search PubMed.
- L. Zhao, M. Qu, G. Wong and D.-Y. Wang, Environ. Sci.: Nano, 2017, 4, 2356–2366 RSC.
- L. Lei, S. Wu, S. Lu, M. Liu, Y. Song, Z. Fu, H. Shi, K. M. Raley-Susman and D. He, Sci. Total Environ., 2018, 619–620, 1–8 CrossRef CAS PubMed.
- S. K. Hanna, A. R. Montoro Bustos, A. W. Peterson, V. Reipa, L. D. Scanlan, S. Hosbas Coskun, T. J. Cho, M. E. Johnson, V. A. Hackley, B. C. Nelson, M. R. Winchester, J. T. Elliott and E. J. Petersen, Environ. Sci. Technol., 2018, 52, 5968–5978 CrossRef CAS PubMed.
- Y.-H. Yang, H.-M. Shao, Q.-L. Wu and D.-Y. Wang, Environ. Pollut., 2019 DOI:10.1016/j.envpol.2019.113439.
- M. Qu, L.-B. Luo, Y.-H. Yang, Y. Kong and D.-Y. Wang, Sci. Total Environ., 2019 DOI:10.1016/j.scitotenv.2019.134131.
- M. Qu, Y.-L. Zhao, Y.-Y. Zhao, Q. Rui, Y. Kong and D.-Y. Wang, Environ. Pollut., 2019 DOI:10.1016/j.envpol.2019.113137.
- M. Qu, K.-N. Xu, Y.-H. Li, G. Wong and D.-Y. Wang, Sci. Total Environ., 2018, 643, 119–126 CrossRef CAS PubMed.
- M. Qu, Y.-X. Qiu, Y. Kong and D.-Y. Wang, Environ. Pollut., 2019 DOI:10.1016/j.envpol.2019.112978.
- M. Qu, A. Nida, Y. Kong, H.-H. Du, G.-S. Xiao and D.-Y. Wang, Ecotoxicol. Environ. Saf., 2019 DOI:10.1016/j.ecoenv.2019.109568.
- H.-M. Shao, Z.-Y. Han, N. Krasteva and D.-Y. Wang, Nanotoxicology, 2019, 13, 174–188 CrossRef CAS PubMed.
- L.-T. Zhi, M.-X. Ren, M. Qu, H.-Y. Zhang and D.-Y. Wang, Sci. Rep., 2016, 6, 39261 CrossRef CAS PubMed.
- L.-T. Zhi, M. Qu, M.-X. Ren, L. Zhao, Y.-H. Li and D.-Y. Wang, Carbon, 2017, 113, 122–131 CrossRef CAS.
- P.-D. Liu, H.-M. Shao, Y. Kong and D.-Y. Wang, J. Environ. Sci., 2020, 88, 200–208 CrossRef.
- D. M. Eisenmann, Wnt signaling, WormBook, 2005, DOI:10.1895/wormbook.1.7.1.
- M. Qu, Y.-Q. Liu, K.-N. Xu and D.-Y. Wang, Adv. Biosyst., 2019, 3, 1800325 CrossRef CAS.
- M. Al-Sid-Cheikh, S. Rowland, K. Stevenson, C. Rouleau, T. B. Henry and R. C. Thompson, Environ. Sci. Technol., 2018, 52, 14480–14486 CrossRef CAS PubMed.
- Y. Li, D. Zhao, T. Horie, G. Chen, H. Bao, S. Chen, W. Liu, R. Horie, T. Liang, B. Dong, Q. Feng, Q. Tao and X. Liu, Proc. Natl. Acad. Sci. U. S. A., 2017, 114, E6352–E6360 CrossRef CAS PubMed.
- M. V. Espelt, A. Y. Estevez, X. Yin and K. Strange, J. Gen. Physiol., 2005, 126, 379–392 CrossRef CAS PubMed.
- T. Sijen, J. Fleenor, F. Simmer, K. L. Thijssen, S. Parrish, L. Timmons, R. H. A. Plasterk and A. Fire, Cell, 2001, 107, 465–476 CrossRef CAS PubMed.
- Y. Shen, L. F. Ng, N. P. W. Low, T. Hagen, J. Gruber and T. Inoue, PLoS One, 2016, 11, e0153233 CrossRef PubMed.
- H. Qadota, M. Inoue, T. Hikita, M. Koppen, J. D. Hardin, M. Amano, D. G. Moerman and K. Kaibuchi, Gene, 2007, 400, 166–173 CrossRef CAS PubMed.
- S. Brenner, Genetics, 1974, 77, 71–94 CAS.
- Y. Kong, H.-L. Liu, W.-J. Li and D.-Y. Wang, Biochem. Biophys. Res. Commun., 2019, 517, 278–284 CrossRef CAS PubMed.
- Y.-L. Zhao, L. Jin, Y. Wang, Y. Kong and D.-Y. Wang, Sci. Rep., 2019, 9, 12144 CrossRef PubMed.
- X.-K. Cheng, S.-S. Dong, Q. Rui, D.-Y. Wang and J.-D. Jiang, Ecotoxicol. Environ. Saf., 2019 DOI:10.1016/j.ecoenv.2019.109848.
- Q. Rui, S.-S. Dong, W.-K. Jiang and D.-Y. Wang, Ecotoxicol. Environ. Saf., 2019 DOI:10.1016/j.ecoenv.2019.109782.
- Y.-L. Zhao, Q.-L. Wu and D.-Y. Wang, Biomaterials, 2016, 79, 15–24 CrossRef CAS PubMed.
- L.-F. Shi, X.-H. Jia, T.-T. Guo, L. Cheng, X.-X. Han, Q.-L. Wu and D.-Y. Wang, RSC Adv., 2019, 9, 13722–13735 RSC.
- H. C. Korswagen, M. A. Herman and H. C. Clevers, Nature, 2000, 406, 527–532 CrossRef CAS PubMed.
- E. F. Hoier, W. A. Mohler, S. K. Kim and A. Hajnal, Genes Dev., 2000, 14, 874–886 CAS.
- H. Zhang and S. W. Emmons, Genes Dev., 2000, 14, 2161–2172 CrossRef CAS PubMed.
- M. A. Essers, L. M. M. de Vries-Smits, N. Barker, P. E. Polderman, B. M. T. Burgering and H. C. Korswagen, Science, 2005, 308, 1181–1184 CrossRef CAS PubMed.
- L. Dreier, M. Burbea and J. M. Kaplan, Neuron, 2005, 46, 51–64 CrossRef CAS PubMed.
- A. Yoda, H. Kouike, H. Okano and H. Sawa, Development, 2005, 132, 1885–1893 CrossRef CAS PubMed.
- A. B. Byrner, M. T. Weirauch, V. Wong, M. Koeva, S. J. Dixon, J. M. Stuart and P. J. Roy, J. Biol., 2007, 6, 8 CrossRef PubMed.
- J. E. Irazoqui, A. Ng, R. J. Xavier and F. M. Ausubel, Proc. Natl. Acad. Sci. U. S. A., 2008, 105, 17469–17474 CrossRef CAS PubMed.
- M. L. Van der Bent, M. G. Sterken, R. J. M. Volkers, J. A. G. Riksen, T. Schmid, A. Hajnal, J. E. Kammenga and L. B. Snoek, Sci. Rep., 2014, 4, 4926 CrossRef PubMed.
- B. M. Jackson, P. Abete-Luzi, M. W. Krause and D. M. Eisenmann, G3: Genes, Genomes, Genet., 2014, 4, 733–747 CrossRef CAS PubMed.
- M. Kniazeva, Q. T. Crawford, M. Seiber, C. Y. Wang and M. Han, PLoS Biol., 2004, 2, e257 CrossRef PubMed.
- R. Wang, M. Kniazeva and M. Han, PLoS One, 2013, 8, e76270 CrossRef CAS PubMed.
- H. Thieringer, B. Moellers, G. Dodt, W. H. Kunau and M. Driscoll, J. Cell Sci., 2003, 116, 1797–1804 CrossRef CAS PubMed.
- A. M. Motley, E. H. Hettema, R. Ketting, R. Plasterk and H. F. Tabak, EMBO Rep., 2000, 1, 40–46 CrossRef CAS PubMed.
- M.-X. Ren, L. Zhao, X. Lv and D.-Y. Wang, Nanotoxicology, 2017, 11, 578–590 CrossRef CAS PubMed.
- L.-T. Zhi, Y.-L. Yu, X.-Y. Li, D.-Y. Wang and D.-Y. Wang, PLoS Pathog., 2017, 13, e1006152 CrossRef PubMed.
- A. Gurvitz, S. Langer, M. Piskacek, B. Hamilton, H. Ruis and A. Hartig, Yeast, 2000, 17, 188–200 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c9en01085h |
| This journal is © The Royal Society of Chemistry 2020 |