
Glucagon-like peptide-1 loaded phospholipid micelles for the treatment of type 2 diabetes: improved pharmacokinetic behaviours and prolonged glucose-lowering effects
Abstract
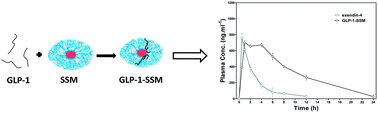
Glucagon-like peptide-1 (GLP-1) and GLP-1 receptor agonists are actively pursued as therapeutic agents for type 2 diabetes mellitus (T2DM). However, the therapeutic utility of GLP-1 is limited due to its rapid inactivation by dipeptidyl peptidase IV, and many GLP-1 receptor agonists suffer from innegligible adverse effects. In the present study, in order to develop long-acting GLP-1 derivatives with improved hypoglycemic activity, native GLP-1 (7-36)-NH2 was loaded into sterically stabilized phospholipid micelles (SSM), affording GLP-1-SSM. In vitro stability test and in vivo pharmacokinetic study demonstrated that the association of GLP-1 with SSM led to enhanced stability and drug utilization without affecting its insulinotropic and glucose-lowering activities. Single and multiple glucose tolerance tests confirmed that GLP-1-SSM was a long-acting antidiabetic agent comparable or even better than exendin-4. More importantly, preclinical studies found out that a chronic twice daily treatment of GLP-1-SSM in type 2 diabetic db/db mice suppressed body weight gain and food uptake, decreased HbA1c value, and restored the glucose tolerance ability. Collectively, our results suggest that GLP-1-SSM is a promising therapy for the treatment of T2DM and deserves further investigation.
 Please wait while we load your content...
Please wait while we load your content...

