
Borage oil supplementation decreases lipopolysaccharide-induced inflammation and skeletal muscle wasting in mice
Abstract
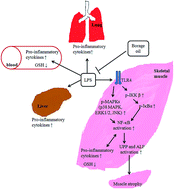
Because in vitro data have shown gamma-linolenic acid (GLA) to be protective in LPS-induced macrophage inflammation and myotube atrophy, we explored the therapeutic value of borage oil (BO), a GLA rich oil, in LPS-induced inflammation and muscle wasting in C57BL/6JNarl mice. Supplementation with BO was more potent than supplementation with soybean oil (SO) in decreasing LPS-induced expression of pro-inflammatory cytokines and glutathione in serum and tissues. Notably, GLA did not reverse LPS-induced inflammatory cytokine expression in C2C12 myotubes transfected with a constitutively active mutant IκB kinase-β plasmid, which suggested the importance of the inhibition of nuclear factor-κB (NF-κB) activation by GLA. Moreover, BO prevented LPS-induced skeletal muscle weight loss as well as molecule expression of ubiquitin-proteasome pathway and the autophagy-lysosomal pathway which played a key role in skeletal muscle protein degradation. BO but not SO reduced the LPS-induced increase in toll-like receptor 4 (TLR4) expression and activation of mitogen-activated protein kinases (MAPKs) and NF-κB in gastrocnemius muscle. In summary, supplementation with BO is more effective than supplementation with SO in preventing LPS-induced inflammation and muscle wasting. Blockade of the TLR4/MAPKs/NF-κB pathway is crucial in the action of BO on LPS-induced inflammation and wasting in skeletal muscle.
 Please wait while we load your content...
Please wait while we load your content...

