
Advances in exploring alternative Taxol sources
Abstract
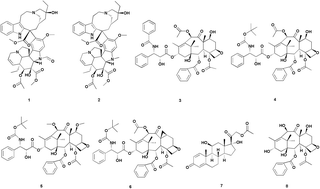
The protection and sustainable utilization of natural resources are among the most pressing global problems of the 21st century. Taxol is a well-known anticancer agent with a unique active mechanism, mainly isolated from the slow-growing yew tree, which has achieved the status of “blockbuster drug” due to its popularity in the treatment of cancers. However, the source of Taxol has always been a primary concern, because its content in the plant is extremely low. In this review, we introduce the advances in exploring alternative Taxol sources aside from total synthesis, including Taxol from nursery cultivated Taxus, semi-synthesis of Taxol, Taxol from Taxus cell culture, production of Taxol by synthetic biology, Taxol from the endophytic fungi and Taxol from other non-Taxus plants. The future perspective of Taxol production is also discussed.
 Please wait while we load your content...
Please wait while we load your content...

