
Poly(vinyl alcohol): review of its promising applications and insights into biodegradation
Abstract
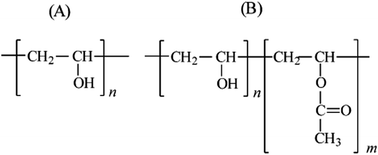
Poly(vinyl alcohol) (PVA) is a water soluble synthetic polymer, with a backbone composed only of carbon atoms and is biodegradable under both aerobic and anaerobic conditions. This polymer can be prepared by the hydrolysis of polyvinylacetate and is one of the most important synthetic polymers used in commercial, industrial, medical and nutraceutical applications. The environmental issues caused by PVA industrial practice have increased globally. Several methods have been used to treat PVA industrial discharge including in particular physicochemical methods such as electrocoagulation. Nowadays, use of bioremediation for PVA release, which has caused serious pollution problems in the natural environment, has attracted much interest. The bioremediation ability of microorganisms and their PVA degrading enzymes, especially PVA oxidases/hydrolases, has long been perceived. These enzymes as well as symbiotic microorganisms could be an effective means for biodegradation of PVA.
 Please wait while we load your content...
Please wait while we load your content...

