
Collagen fibril strain, recruitment and orientation for pericardium under tension and the effect of cross links
Hanan R. Kayeda,
Nigel Kirbyb,
Adrian Hawleyb,
Stephen T. Mudieb and
Richard G. Haverkamp*a
aSchool of Engineering and Advanced Technology, Massey University, Private Bag 11222, Palmerston North, New Zealand. E-mail: r.haverkamp@massey.ac.nz
bAustralian Synchrotron, 800 Blackburn Road, Clayton, Melbourne, Australia
First published on 30th November 2015
Abstract
The structural response of collagen fibrils in pericardium and other tissues when subjected to strain and the effect of cross linking on those structural changes are not well understood. Specifically, there is uncertainty about whether natural cross links of glycosaminoglycan (GAG) and synthetic cross links of glutaraldehyde have a mechanical function. Bovine pericardium was treated either with chondroitinase ABC to remove natural cross links or with glutaraldehyde to form synthetic cross links. The collagen fibril orientation index (OI) and D-spacing was measured on pericardium subjected to strain using synchrotron-based small angle X-ray scattering (SAXS). Under strain the collagen fibrils become much more oriented in the direction of the strain, with OI increasing from 0.25 to 0.89 in chondroitinase ABC-treated material, 0.22 to 0.93 in native material, and 0.22 to 0.77 in the glutaraldehyde-treated material. The proportion of fibrils that are recruited during stress varies from 36% in chondroitinase ABC-treated material, 12% in native material, to 45% in the glutaraldehyde-treated material. The increase in D-spacing shows the individual fibrils are strained in chondroitinase ABC-treated material by 2.4% on average or 4.6% for those in the direction of applied strain, in native material, 2.7% and 4.1%, respectively, and in the glutaraldehyde-treated material, 3.2% and 6.4%, respectively. Glutaraldehyde cross links are, therefore, shown to constrain the collagen fibrils and link them together mechanically. GAGs do not have such a marked mechanical effect; contrarily, the nature of internal structural responses to strain suggests that GAGs may have a lubricating rather than a binding effect.
1. Introduction
The collagen I molecule assembles with a complex hierarchical structure and is a major extracellular matrix component in a multitude of animal structural tissues including pericardium, tendon, cornea, lung and skin. The responses to stresses imposed on collagen materials has been widely studied with the aim of understanding what and how components of collagen play a role in its mechanical properties, both at the macroscopic and microscopic levels.Collagen materials can be stiff, flexible or extensible, depending on the required function in the body. For example, tendons and ligaments are crucial to joint movement, enabling force transmission and are therefore required to be flexible and have high tensile strength. The mechanical properties of collagen are due in part to its highly fibrillar nature1,2 and its ability to respond to stresses.3 However, it has also been suggested that the cross links between collagen fibrils contribute to the mechanical properties of collagen materials.
Proteoglycan bridges are considered to form shapemodules, and are found in the gap regions of collagen fibril D-spacing, linking fibrils together in natural tissue.4–7 These proteoglycan bridges are elastic and predominantly of decoran, containing the glycosaminoglycan (GAG) dermochondan sulfate.8,9
The role of GAGs in the mechanical response of collagen tissues to stress is contested. Among those who consider the role of GAGs to be significant, there are divided opinions on how these cross links function under tension. Some believe GAGs act as force-sharing elements, transferring shear forces via their connections to the collagen fibrils, allowing fibril stretching and restricting sliding,10 whilst others propose the hydrophilic nature of GAGs facilitates fibril sliding.11 How GAGs might transmit forces between fibrils so that the fibrils resist the sliding forces has been modelled.12–16 The energy absorbed by enthalpic transformations in specific GAGs such as dermochondan sulfate can be significant.9,17
Experimental studies of the changes in collagen's mechanical properties resulting from the depletion of the natural GAG content by the application of chondroitinase ABC have highlighted the differences in results and opinions. On one hand the tensile elastic modulus of mouse tendon was found to be reduced over much of the stress–strain curve when GAG content was lowered, while the ultimate tensile force and ultimate stress for the tendon were relatively unchanged.11,18 However, other work has found no altered mechanical properties in tendon resulting from the removal of GAGs.15,19,20 Natural cross linking of collagen in connective tissues increases with age due to glycation and older tissues have been shown to have higher stiffness; therefore, a causal link has been proposed between cross links and stiffness.21
Synthetic cross links can also be introduced; glutaraldehyde is commonly used as a cross-linking agent, particularly in materials for biological heart valve replacements, forming both inter- and intra-molecular cross links between collagen fibrils.22–24 As with GAG cross links, there is debate as to the resulting mechanical properties of collagen tissue treated with glutaraldehyde. Such treatment of bovine pericardium has been reported to result in a less extensible and stiffer material which is stronger than the untreated material.25,26 Contrary to these findings, others have observed an increase in extensibility upon glutaraldehyde treatment,22,27 reduced ultimate tensile strength,27 no changes to ultimate tensile strength22 and increases in elastic modulus.25,28
Cross linking of collagen may affect the arrangement of the collagen fibrils. Glutaraldehyde treatment has been shown to result in a less highly oriented material whereas the removal of GAGs does not have a significant modifying effect.29 The arrangement of collagen fibrils, particularly the extent to which the fibrils are well oriented, is an important determinant of the strength of collagen materials. The structure–function relationship between collagen orientation and its mechanical properties has been determined for a range of tissue types.30–32 The orientation of collagen measured in-plane has been shown in a range of mammal skins processed to leather to be correlated with strength.33,34 A useful method of quantitatively measuring this structural arrangement of collagen fibrils is small angle X-ray scattering (SAXS).30,31,35,36
Here, we investigate the structural response of collagen cross linked by glutaraldehyde or GAGs to applied strain and stress to add to the understanding of the mechanical function, or lack of mechanical function, of these cross links. Bovine pericardium is used as a suitable model material for this study. Bovine pericardium has an established use for heart valve leaflet replacement37 and the effect on the mechanical properties of this material by the removal of GAGs has been investigated.29,38
2. Methods
2.1 Fresh pericardium samples
Fresh bovine pericardium was obtained from Southern Lights Biomaterials and stored in phosphate-buffered saline (PBS) solution (Lorne Laboratories Ltd). The tissue was rinsed briefly in PBS solution, and then cut into rectangles approximately 45–50 × 15 mm, with the long axis taken from the long axis of the heart. The pericardium was washed for 24 h in a 1% octylphenol ethylene oxide condensate (Triton X-100, Sigma), 0.02% EDTA solution made up in PBS. This washing step was conducted under constant agitation at 4 °C. The samples were then rinsed in PBS buffer and stored in PBS. Samples in this state are referred to as “native”. Subsequent processing of this material produced glutaraldehyde-treated or chondroitinase ABC-treated material. All samples were taken from one pericardium and randomly assigned to each treatment method.2.2 Glutaraldehyde treatment
The native pericardium was incubated in a 0.6% glutaraldehyde solution made up in PBS buffer at 4 °C for 24 h with constant agitation.39 It was then stored in a sealed container in a solution of the same composition for 12 days, before being rinsed and stored in PBS until SAXS measurements were performed. The total time in storage was approximately 18 days.2.3 Chondroitinase ABC treatment
Removal of GAG cross links was based on the method described by Schmidt et al. (1990). The native pericardium was incubated in 0.125 units of chondroitinase ABC per ml of buffer solution comprising of 0.05 M tris–HCl, 0.06 M sodium acetate and EDTA-free protease inhibitors (complete, EDTA-free protease inhibitor tablets, Roche Diagnostics, Mannheim, Germany) at approximately 27 °C for 24 h before rinsing and storing in 0.05 M tris–HCl and 0.06 M sodium acetate buffer in a sealed container at 4 °C for 12 days. The samples were then rinsed and stored in PBS until SAXS measurements were performed. The total time in storage was approximately 18 days.Care was taken with all handling, cutting and treatment of the samples not to stretch the material as this might cause fibril alignment to change.
The data here represents a duplication of this experiment (on a different pericardium) with additional care taken in sample selection and handling in the second set of samples and SAXS analysis informed by the experience from the initial experiments. However, one portion of the initial data, that for fibril recruitment, is included (in Section 3.5).
2.4 GAG assay
An assay for sulfated GAGs was performed in triplicate for each of the sample treatments. GAGs were extracted with 1 ml extraction reagent consisting of a 0.2 M sodium phosphate buffer at pH 6.4, containing 8 mg ml−1 sodium acetate, 4 mg ml−1 EDTA, 0.8 mg ml−1 cysteine HCl and 0.1 mg ml−1 papain enzyme (Carica papaya, Sigma, Biochemika, Enzyme no. 3.4.22.2). Each pericardium sample was incubated at 65 °C for 26 h. These samples were centrifuged and the supernatant containing the extracted GAGs collected. The concentration of GAGs in solution was determined with a Blyscan Sulfated Glyscosaminoglycan Assay kit (Bicolor, Carrickfergus, UK). GAGs were precipitated with 1 ml of dye reagent to 20 or 40 μl of supernatant diluted to 100 μl, mechanically inverted for 30 minutes, and then centrifuged. The unbound dye was drained off and 1 ml of Blyscan dissociation reagent was added to the precipitate, left to dissolve for approximately 10 minutes, and centrifuged. Absorbance was measured at a wavelength of 656 nm and compared with a standard curve of chondroitin 4-sulfate GAG reference standard (Biocolor, UK).2.5 SAXS analysis
The rectangles of pericardium were removed from the PBS solutions in which they were stored, mounted and their diffraction patterns recorded while the pericardium was wet. All diffraction patterns were recorded at room temperature.Diffraction patterns were recorded on the Australian Synchrotron SAXS/WAXS beamline, utilizing a high-intensity undulator source. Energy resolution of 10−4 (e.g. 1 × 10−4 Å for 1 Å radiation) was obtained from a cryo-cooled Si(111) double-crystal monochromator and the beam size (FWHM focused at the sample) was 250 × 80 μm, with a total photon flux of about 2 × 1012 ph s−1. All diffraction patterns were recorded with an X-ray energy of 12 keV using a Pilatus 1M detector with an active area of 170 × 170 mm and a sample-to-detector distance of 3371 mm. Exposure time for diffraction patterns was 1 s and data processing was carried out using the software scatterBrainAnalysis V2.71.40
Orientation index (OI) is used to give a measure of the uniformity of fibril orientation (an OI of 1 indicates the fibrils are parallel to each other; an OI of 0 indicates the fibrils are randomly oriented). OI is defined as (90° – OA)/90°, where OA is the minimum azimuthal angle range that contains 50% of the fibrils, based on the method of Sacks for light scattering41 but converted to an index,33 using the spread in azimuthal angle of one or more D-spacing diffraction peaks. The peak area is measured, above a fitted baseline, at each azimuthal angle.
One chondroitinase ABC-treated sample and two of each native and glutaraldehyde-treated samples were tested. Six diffraction patterns were recorded at different positions across the samples following every stretch. From each scattering pattern the OI and D-spacing were calculated from the azimuthal spread of the 5th and 9th order collagen diffraction peaks (at around 0.05 Å−1 and 0.09 Å−1, respectively) and averaged where there were single diffraction peaks. For those diffraction patterns displaying peak splitting and/or double peaks, the 9th order diffraction peak was used to calculate OI and D-spacing. For some of the glutaraldehyde-treated samples, the 9th order peak became very complex at higher strains and background intensity could not be identified and subtracted; in such cases, the 5th order diffraction peak was used to determine OI and D-spacing. The portion of fibrils experiencing higher stresses (larger D-spacing) was determined using the intensities of these split/double diffraction peaks as intensity is proportional to the quantity of collagen fibrils involved in diffraction.
3. Results
3.1 GAG removal by chondroitinase ABC treatment
Chondroitinase ABC treatment removed 81% of GAGs initially present in native pericardium (from 0.90 μg GAGs per mg tissue to 0.17 μg mg−1). Pericardium treated with glutaraldehyde had a similar GAG content to the native material (0.85 μg mg−1). The GAG content of the chondroitinase ABC treated material was statistically significantly lower than the other two materials (Fig. 1).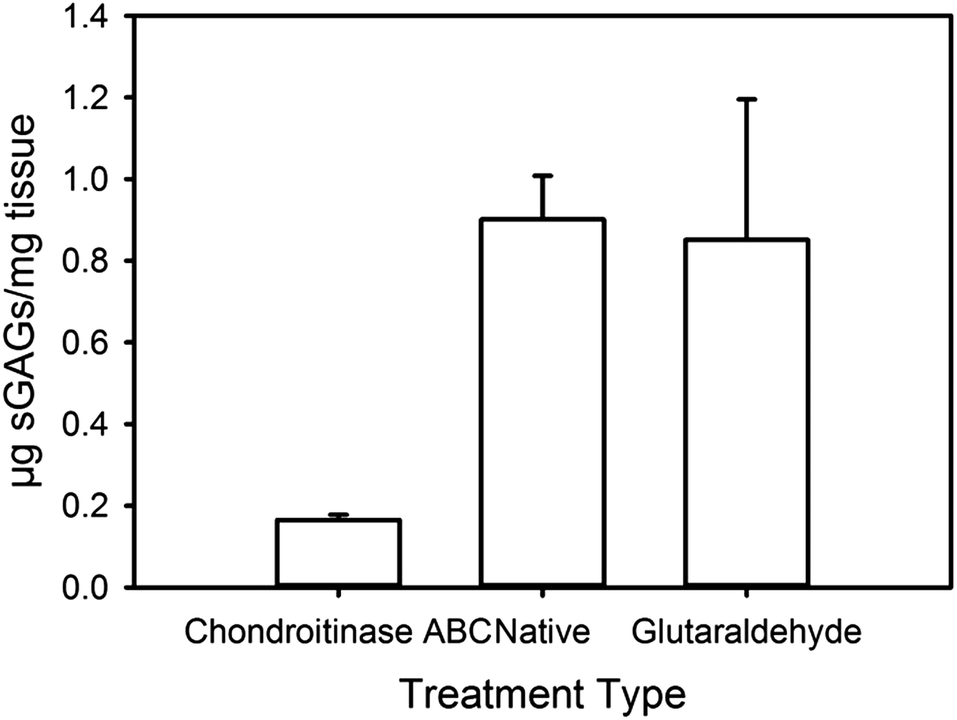 | ||
| Fig. 1 Assay results for the GAG content of the three materials studied here. | ||
3.2 Tensile properties
Stress–strain curves were recorded on the samples being progressively strained during the SAXS measurements (Fig. 2). The stress–strain curves obtained are typical for collagen, showing a foot region and linear region.25 While the stress–strain curves are on small samples, and may therefore not give a representative measure of the mechanical properties to be expected of bulk samples of these materials, these curves are important in the context of the structural analysis during strain of these tissues presented in the body of this work. However, in a previous report we found the ultimate tensile stress was highest for glutaraldehyde-treated pericardium.29 Other studies of glutaraldehyde treatment of bovine pericardium have reported that this material is stronger than the untreated material25,26 although there is a wide variety of opinions regarding changes to other mechanical properties with glutaraldehyde treatment.22,27,28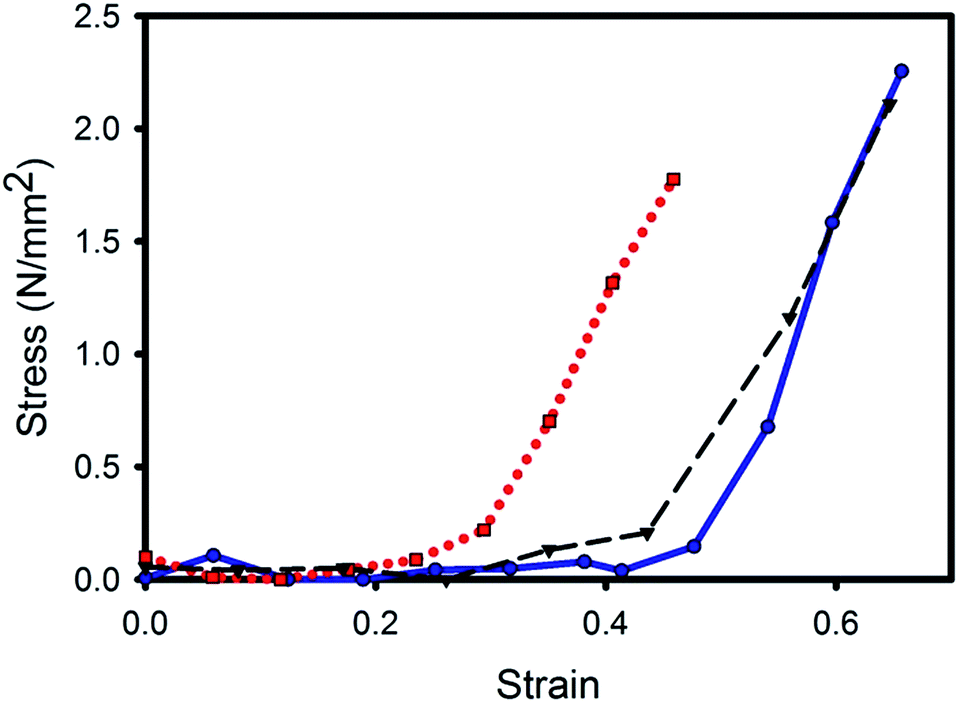 | ||
Fig. 2 Stress–strain curves for the pericardium after three different treatments, while under increasing tension during the SAXS measurements: ( , ,  , blue) chondroitinase ABC treated; ( , blue) chondroitinase ABC treated; ( , ,  , black) native; ( , black) native; ( , ,  , red) glutaraldehyde treated. , red) glutaraldehyde treated. | ||
3.3 SAXS
The pericardium scattering patterns had clearly defined diffraction rings due to the D-spacing periodicity (Fig. 3). When the tissue was not under strain, these rings were of almost uniform intensity around the circle (Fig. 3a), but as the samples were subjected to more strain, the rings subtended a smaller azimuthal angle (Fig. 3b and c). This was a result of the fibrils aligning in the direction of strain. Also, the central region of the pattern elongated at 90° to the direction of the diffraction rings. (This central region is the low q part of the pattern and represents scattering from the collagen fibril diameter, which is at right angles to the D-spacing.) Another feature of the scattering patterns of pericardium under strain was the shifting of the D-spacing scattering angle, particularly of those fibrils aligned in the direction of the strain. This shift is seen as a protuberance on the inside of the diffraction ring but is more readily seen on the integrated intensity plots (Fig. 4). In these, the splitting of the D-spacing into multiple peaks or a broad peak is more apparent at higher diffraction orders. The odd-numbered peaks have a much higher intensity than the even-numbered peaks, a characteristic attributed to a fully hydrated sample.42 | ||
| Fig. 3 A series of typical scattering patterns of native pericardium subjected to (a) no strain; (b) strain of 0.18; (c) strain of 0.45. | ||
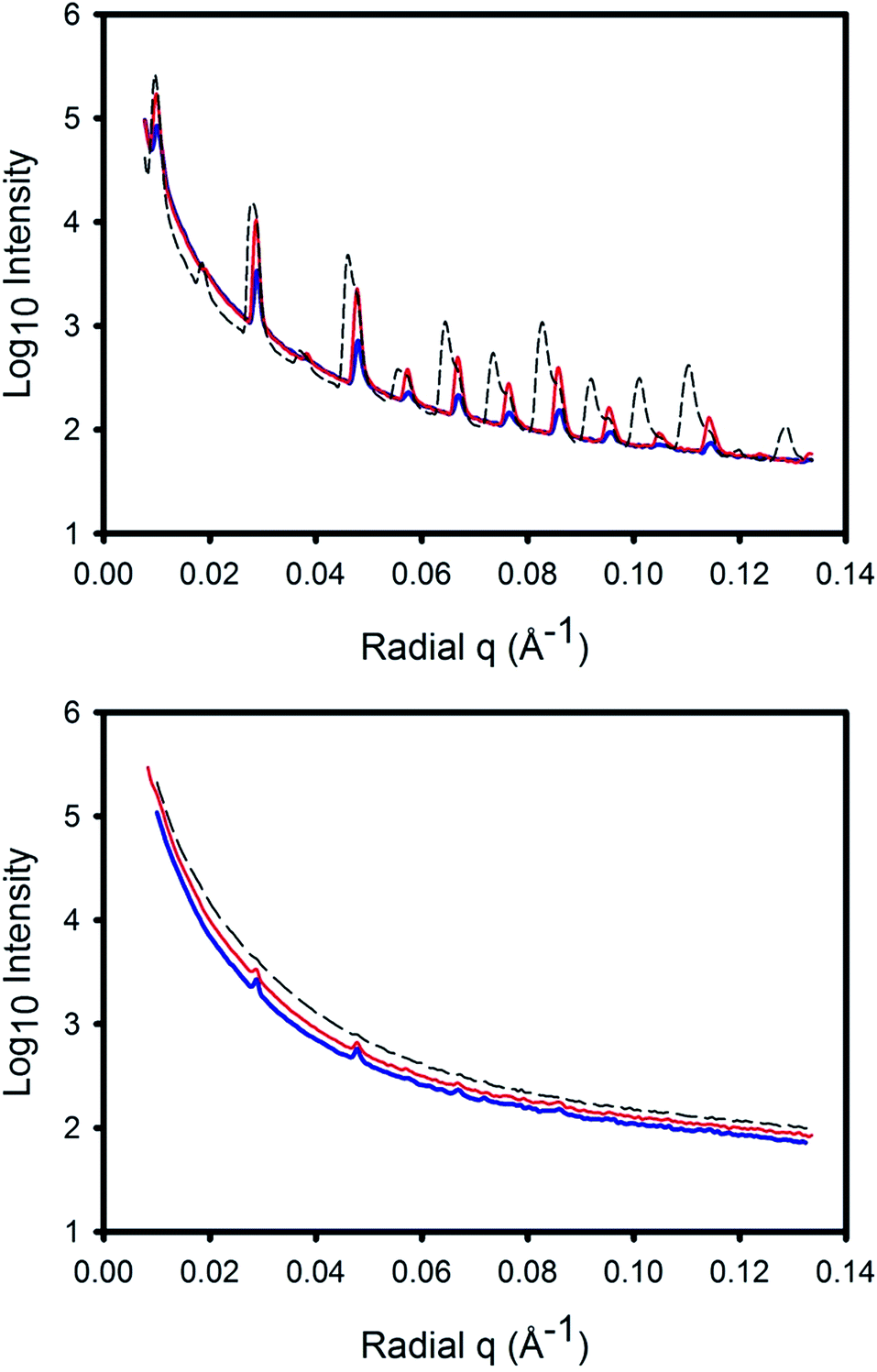 | ||
Fig. 4 Representative integrated scattering patterns of pericardium subjected to varying levels of strain: no strain ( , blue); 18% strain ( , blue); 18% strain ( , red); 45% strain ( , red); 45% strain ( , black). The sharp peaks are due to diffraction of the D-spacing (at different orders) and the peaks split at higher strain. The top image is for a 5° azimuthal angle segment in the direction of strain, the bottom image is for a 5° azimuthal angle segment normal to the direction of strain. , black). The sharp peaks are due to diffraction of the D-spacing (at different orders) and the peaks split at higher strain. The top image is for a 5° azimuthal angle segment in the direction of strain, the bottom image is for a 5° azimuthal angle segment normal to the direction of strain. | ||
3.4 OI
The distribution of fibril orientation is measured from a plot of the intensity (we use the peak area) of any of the collagen diffraction peaks (Fig. 5). A narrow peak in this plot is indicative of more highly aligned collagen fibrils whereas a broader peak indicates a more isotropic arrangement. This can be quantified as the orientation index, OI. | ||
Fig. 5 Representative integrated scattering intensity at the 5th order D-spacing diffraction peak verses azimuthal angle for pericardium subjected to: no strain( , blue); 18% strain ( , blue); 18% strain ( , red); 45% strain ( , red); 45% strain ( , black). , black). | ||
Both the OI and the D-spacing can be visualized in a three-dimensional plot (Fig. 6), where the D-spacing of fibrils in a given direction can be seen more clearly.
 | ||
| Fig. 6 Three-dimensional representation of an example scattering pattern of (a) native pericardium at a strain of 0.45; (b) chondroitinase ABC-treated pericardium at a strain of 0.69, where both the fibril orientation (from the azimuthal angle axis) and the D-spacing shift (from the radial q axis) can be visualized. Only the azimuthal range −90° to 90° is represented as the remaining range is a duplication of this information and only a small portion of the radial angle is displayed representing one D-spacing diffraction peak. | ||
3.5 Changes to structure during strain
Initially, the pericardium exhibited little physical resistance to the strain (the stress was low), and there was little change in the D-spacing (Fig. 7). During this period, however, there was a large change in the OI. The change in OI at this stage of the tensile testing can be attributed to a combination of crimp straightening and re-orientation of the collagen fibrils towards the direction of strain. The D-spacing increase represents the stress on individual fibrils and this increased in tandem with the increasing stress on the whole tissue. (Note that the increase in D-spacing can arise either from the direct stretching of the collagen molecules in a fibril, so that the relative lengths of the overlap and gap regions remain constant, or from the sliding of the collagen molecules past one another, so that the relative length of the gap and overlap regions change. Or the increase can arise from a combination of the two. The mechanism of D-spacing change was not measured here, so D-spacing in this work includes the possibilities of both mechanisms.) Not all fibrils experienced the same stress: the fibrils in line with the direction of applied strain experienced greater stress and underwent a greater change in D-spacing until there were two distinct diffraction peaks for D-spacing at different angles. At the point when these two distinct D-spacings could be identified, the split diffraction peaks were fitted individually to obtain both OI and D-spacing, and were plotted in Fig. 7 in the high-strain portion of the plot. | ||
Fig. 7 Changes in OI and D-spacing as pericardium was subjected to increasing strain for each of the treatment types: (a and b) chondroitinase ABC treated; (c and d) native; (e and f) glutaraldehyde treated, where stress ( , ,  , black); weighted sum OI or D-spacing ( , black); weighted sum OI or D-spacing ( , ,  , blue); non-recruited fibril OI or D-spacing ( , blue); non-recruited fibril OI or D-spacing ( , ,  , red); recruited fibril OI or D-spacing ( , red); recruited fibril OI or D-spacing ( ,· ,· , green). , green). | ||
While the behaviour for all three samples types was broadly similar there were some differences. All three materials have a portion of fibrils that are highly strained and highly oriented. In the glutaraldehyde-treated material, this portion (which can be called “recruited fibrils”, i.e. fibrils that participate in absorbing the stresses) was higher (Table 1) and the stress experienced by these fibrils was greater (evidenced in the D-spacing in Fig. 7), followed by chondroitinase ABC-treated material and finally native material.
| Pericardium treatment | % Fibrils recruited to stretching | ||
|---|---|---|---|
| (Duplicate pericardium measurements) | (From Fig. 6) | Average | |
| Glutaraldehyde | 37.9 | 52.1 | 45.0 |
| Native | 13.1 | 10.9 | 12.0 |
| Chondroitinase ABC | 37.4 | 33.7 | 35.6 |
3.6 Comparison of OI between treatments with increasing strain
As the pericardium was strained, the fibrils reoriented to line up in the direction of strain, as reflected in the lowering of the OI. However, changes in sample OI varied among the treatments (Fig. 8), with the maximum OI achieved being highest for native pericardium, slightly less for the GAG-depleted pericardium and lowest for the glutaraldehyde-cross linked pericardium. The recruited fibrils were all highly oriented as expected3 whilst the remaining non-recruited fibrils in the collagen matrix were less aligned in the glutaraldehyde-treated pericardium than in the native or chondroitinase ABC-treated tissue (Fig. 8b and c, respectively).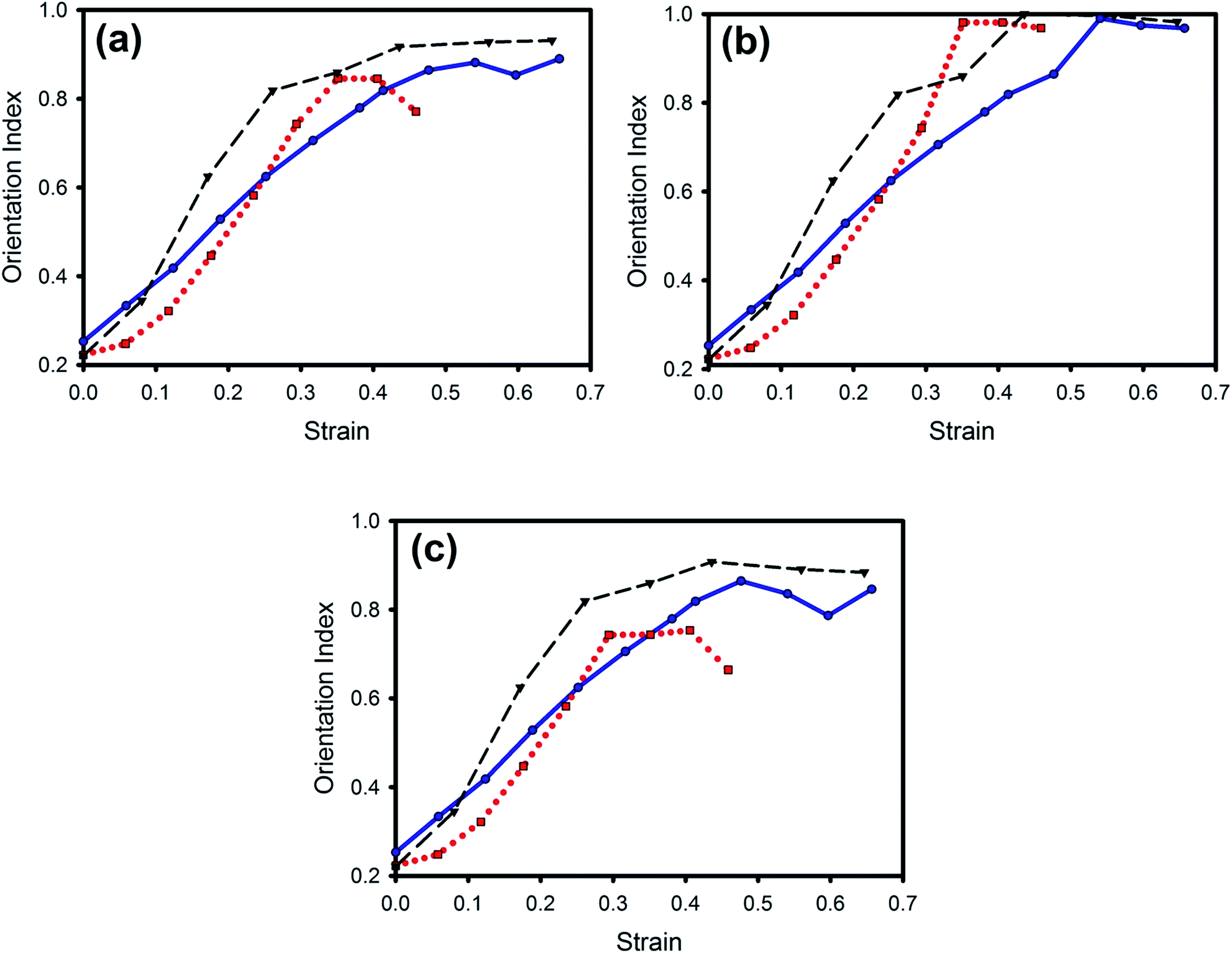 | ||
Fig. 8 Comparison of change in OI with increasing strain for the three treatments: (a) average of all fibrils; (b) recruited fibrils; (c) non-recruited fibrils. Chondroitinase ABC-treated pericardium ( , ,  , blue); native pericardium ( , blue); native pericardium ( , ,  , black); glutaraldehyde-treated pericardium ( , black); glutaraldehyde-treated pericardium ( , ,  , red). , red). | ||
3.7 Comparison of fibril strain with stress
As stress is applied to pericardium, this stress is transmitted to the individual collagen fibrils. This results in an extension of length of the fibrils which can be directly measured by the D-spacing change. The D-spacing change, therefore, acts as an internal strain gauge.2,43 There was a significant variation in the internal strain placed on the collagen fibrils between treatments. Collagen fibrils in the glutaraldehyde-treated materials experienced the greatest strain for a given sample stress, with the chondroitinase ABC-treated material experiencing the least, and native pericardium intermediate between these for the analysis of all the fibrils (Fig. 9). Of those fibrils recruited into stretching, those of native and chondroitinase ABC-treated pericardium experienced similar strains (slightly higher in the latter) whilst those in the glutaraldehyde-treated pericardium experienced significantly higher strains (Fig. 9b). | ||
Fig. 9 Comparison of D-spacing change (indicating fibril strain) with increasing sample stress for the three treatments: chondroitinase ABC treated ( , ,  , blue); native ( , blue); native ( , ,  , black); glutaraldehyde treated ( , black); glutaraldehyde treated ( , ,  , red). (a) Average of all fibrils; (b) recruited fibrils only. , red). (a) Average of all fibrils; (b) recruited fibrils only. | ||
3.8 Fibril strain and tissue strain
The ratios of fibril strain (measured by D-spacing increase) to tissue strain (measured by the extension of the pericardium sample; calculated from data in Fig. 7b, d and f) were the following for the overall tissue for each treatment type: chondroitinase ABC-treated samples 0.12; native 0.12; glutaraldehyde-treated samples 0.16. These ratios are similar to that reported elsewhere of 0.18 for a different sample of native pericardium.2 Of those fibrils taking up the stress, the ratio of fibril strain to tissue strain was 0.24, 0.2 and 0.37 for chondroitinase ABC-treated, native and glutaraldehyde-treated pericardium, respectively.4. Discussion
The internal structural response and stresses on pericardium collagen fibrils subjected to strain can be interpreted in terms of the contribution that the GAG or glutaraldehyde cross linking has on the rearrangement and stress on the individual collagen fibrils.We propose that there are three ways in which this tissue can accommodate the strain and the stresses: (1) by removal of crimp and re-orienting of fibrils (manifested as an OI increase); (2) by stretching of fibrils (D-spacing increase); (3) by sliding of the fibrils (not directly measured here).
The data, and our analysis of it, yield information about the nanostructural behaviours of pericardium tissue being stretched. In turn, these behaviours – strain-induced recruitment of fibrils; strain-induced changes in OI (from crimp removal and rearrangement); and a D-spacing increase from fibril stress – can provide insights into the mechanical structure of the tissue.
4.1 Recruitment
Recruitment is one aspect of re-orientation, and is a measure of the proportion of collagen fibrils that take up the stress that the tissue experiences (Table 1). Fewer fibrils were recruited in the native pericardium (12%) than in the glutaraldehyde-treated (45%) or the chondroitinase ABC-treated pericardium (36%). This suggests that the collagen fibrils in glutaraldehyde treated tissue are mechanically locked together by cross links and, therefore, when collagen fibrils in one direction are subjected to strain, other fibrils also take part in this strain. In native tissue, the minimal recruitment suggests that GAGs do not provide a strong mechanical connection between collagen fibrils. However, if this were so, one would expect the chondroitinase ABC-treated pericardium to behave similarly to the native pericardium, which it does not. So, the evidence from recruitment does not provide evidence for a mechanical cross linking effect of GAGs in tissue.4.2 Reorientation with strain
The amount of strain that a tissue must undergo before reaching maximum OI was similar for native pericardium (0.35) and glutaraldehyde-treated pericardium (0.35) but higher for chondroitinase ABC-treated (GAG depleted) pericardium (0.5). This suggests that in chondroitinase ABC-treated pericardium, mechanical cross links have been removed, allowing more sliding of the collagen fibrils to occur compared with native and glutaraldehyde-treated pericardium. This is consistent with GAGs having a mechanical cross linking action. However, we note the variability in the length of the toe region of the stress–strain curves within treatment groups29 could have been influenced by the initial placement of the samples.4.3 Ratio of fibril strain (stress) to tissue strain
For all three treatment types, the macroscopic tissue strain was larger than the fibril strain, even after the OI reached a plateau, showing that either strain was taken up by other tissue components or that the fibrils had slid axially relative to one another as well as stretching. Therefore, a ratio of fibril strain to tissue strain may indicate the level of fibril sliding.For overall tissue, that ratio of fibril strain to tissue strain was highest in glutaraldehyde-treated pericardium (0.16), with ratios for native tissue and GAG-depleted tissue being the same (0.12). Of the fibrils experiencing the most stress, glutaraldehyde-treated pericardium again had the highest ratio (0.37), whilst native had the lowest (0.20) and chondroitinase ABC-treated material intermediate (0.24). These findings suggest that in glutaraldehyde-treated pericardium the collagen fibrils are held in place, and less able to slide past each other, by the more highly networked structure resulting from the action of cross links. That the ratio of recruited fibril strain to tissue strain for chondroitinase ABC-treated pericardium was slightly higher than that of native pericardium implies that the presence of GAGs encourages more fibril sliding. This strain ratio data, therefore, does not support the theory of GAGs as mechanical cross linkers.
4.4 Ratio of fibril stress to tissue stress
For a given tissue stress, the individual collagen fibrils in glutaraldehyde-cross linked tissue experienced more stress (reflected by the D-spacing increase) than did the native tissue (Fig. 9). The collagen fibrils in GAG-depleted (chondroitinase ABC-treated) tissue experienced less stress than did the native or glutaraldehyde-treated tissue when considering the behaviour of all fibrils. This could be interpreted to mean that in the glutaraldehyde-treated and native pericardia, there are mechanical cross links that hold the fibrils together so that when the tissue is stressed, this stress is transmitted to a greater proportion of the collagen fibrils in the material. In light of this interpretation, the ratio of fibril stress to tissue stress supports a mechanical action of GAGs. However, for those fibrils aligned in the direction of stress, the fibril strain at a given stress was highest for glutaraldehyde-linked pericardium, but was slightly lower for native than for chondroitinase ABC-treated pericardium. From the behaviour of these fibrils, it appears that GAGs do not transmit stresses to individual fibrils.4.5 Alternative explanations
The above interpretations of the analyses of fibril stress and orientation change data can be explained by the mechanical action of glutaraldehyde but do not all support GAGs as mechanical cross linking agents. A radically alternative explanation is that GAGs promote fibril sliding perhaps by acting as lubricants to the collagen fibrils. The sliding forces in fibrous protein systems are typically rather large.44 Lubrication to reduce these forces does not mean weaker material, and it is well known in the leather industry that lubricating processed leather with oil components is necessary to achieve high strength in leather.45Such a role for GAGs acting as lubricants and glutaraldehyde acting as a mechanical cross link are consistent with almost all observations in this work.
The lower fibril recruitment in native material compared to that in chondroitinase ABC-treated material, the ratio of fibril strain to tissue strain (particularly the ratio of the portion of fibrils that take up the most stress), and the ratio of recruited fibril stress to tissue stress are all consistent with GAGs acting as lubricants in the native material, such that the fibrils slide past each other rather than co-opting other fibrils into taking up the applied stress. Unlike glutaraldehyde cross links, which covalently bond to amino acid sidechains on the collagen fibril, GAGs are thought to aggregate through hydrophobic and hydrophilic interactions which can dissociate and reform;46,47 GAG bridges may, therefore, not be as strong as glutaraldehyde cross links, and rather than pulling fibrils into stretch or directly transferring forces, these GAG bridges may skew, dissociate and possibly re-associate as fibrils slide past one another. There is only one set of observations that is not compatible with GAGs acting as lubricants: the reorientation of fibrils with strain (OI change, maximum) although the differences are small.
5. Conclusions
An investigation into the mechanical nature of glycosaminoglycan present in collagen tissue by comparing native pericardium, GAG-depleted pericardium (using chondroitinase ABC) and robustly cross linked pericardium (using glutaraldehyde) identified significant differences in the nanostructural behaviour of the different tissues during mechanical loading. The evidence suggests both GAG and glutaraldehyde cross links have a role in response to applied tension forces, however, the mechanisms by which they respond vary vastly; glutaraldehyde had a clear role in producing a constrained network structure, involving more fibrils in the mechanical response and experiencing higher fibril strains. In contrast, fewer fibrils in native tissue partake in stretching with the fibrils experiencing lower strains. We propose GAGs may act as a lubricant, resulting in more fibril sliding relative to fibril stretching.Acknowledgements
This work was supported by the Massey Doctoral Scholarship (HRK). This research was undertaken on the SAXS/WAXS beamline at the Australian Synchrotron, Victoria, Australia. The Australian synchrotron is acknowledged for travel funding. Southern Lights Biomaterials supplied the pericardium. Sue Hallas assisted with editing.References
- M. A. Meyers, J. McKittrick and P. Y. Chen, Science, 2013, 339, 773–779 CrossRef CAS PubMed.
- H. C. Wells, K. H. Sizeland, H. R. Kayed, N. Kirby, A. Hawley, S. T. Mudie and R. G. Haverkamp, J. Appl. Phys., 2015, 117, 044701 CrossRef.
- W. Yang, V. R. Sherman, B. Gludovatz, E. Schaible, P. Stewart, R. O. Ritchie and M. A. Meyers, Nat. Commun., 2015, 6, 1–9 Search PubMed.
- J. E. Scott and R. A. Stockwell, J. Physiol., 2006, 574, 643–650 CrossRef CAS PubMed.
- A. M. Cribb and J. E. Scott, J. Anat., 1995, 187, 423–428 Search PubMed.
- J. E. Scott and C. R. Orford, Biochem. J., 1981, 197, 213–216 CrossRef CAS PubMed.
- J. E. Scott, Biochem. J., 1980, 187, 887–891 CrossRef CAS PubMed.
- J. E. Scott, J. Physiol., 2003, 553, 335–343 CrossRef CAS PubMed.
- R. G. Haverkamp, M. A. K. Williams and J. E. Scott, Biomacromolecules, 2005, 6, 1816–1818 CrossRef CAS PubMed.
- J. Liao and I. Vesely, J. Biomech., 2007, 40, 390–398 CrossRef PubMed.
- S. Rigozzi, R. Mueller, A. Stemmer and J. G. Snedeker, J. Biomech., 2013, 46, 813–818 CrossRef CAS PubMed.
- Y. Chan, G. M. Cox, R. G. Haverkamp and J. M. Hill, Eur. Biophys. J., 2009, 38, 487–493 CrossRef CAS PubMed.
- R. Puxkandl, I. Zizak, O. Paris, J. Keckes, W. Tesch, S. Bernstorff, P. Purslow and P. Fratzl, Philos. Trans. R. Soc., B, 2002, 357, 191–197 CrossRef CAS PubMed.
- S. W. Cranford and M. J. Buehler, Soft Matter, 2013, 9, 1076–1090 RSC.
- G. Fessel and J. G. Snedeker, J. Theor. Biol., 2011, 268, 77–83 CrossRef CAS PubMed.
- A. Redaelli, S. Vesentini, M. Soncini, P. Vena, S. Mantero and F. M. Montevecchi, J. Biomech., 2003, 36, 1555–1569 CrossRef CAS PubMed.
- R. G. Haverkamp, A. T. Marshall and M. A. K. Williams, J. Phys. Chem. B, 2007, 111, 13653–13657 CrossRef CAS PubMed.
- S. Rigozzi, A. Stemmer, R. Muller and J. G. Snedeker, J. Struct. Biol., 2011, 176, 9–15 CrossRef CAS PubMed.
- G. Fessel and J. G. Snedeker, Matrix Biol., 2009, 28, 503–510 CrossRef CAS PubMed.
- R. B. Svensson, T. Hassenkam, P. Hansen, M. Kjaer and S. P. Magnusson, Connect. Tissue Res., 2011, 52, 415–421 CrossRef CAS PubMed.
- A. J. Bailey, Mech. Ageing Dev., 2001, 122, 735–755 CrossRef CAS PubMed.
- L. Damink, P. J. Dijkstra, M. J. A. Vanluyn, P. B. Vanwachem, P. Nieuwenhuis and J. Feijen, J. Mater. Sci.: Mater. Med., 1995, 6, 460–472 CrossRef.
- D. T. Cheung and M. E. Nimni, Connect. Tissue Res., 1982, 10, 201–216 CrossRef CAS PubMed.
- D. T. Cheung, N. Perelman, E. C. Ko and M. E. Nimni, Connect. Tissue Res., 1985, 13, 109–115 CrossRef CAS PubMed.
- I. J. Reece, R. Vannoort, T. R. P. Martin and M. M. Black, Ann. Thorac. Surg., 1982, 33, 480–485 CrossRef CAS PubMed.
- S. E. Langdon, R. Chernecky, C. A. Pereira, D. Abdulla and J. M. Lee, Biomaterials, 1999, 20, 137–153 CrossRef CAS PubMed.
- H. W. Sung, Y. Chang, C. T. Chiu, C. N. Chen and H. C. Liang, J. Biomed. Mater. Res., 1999, 47, 116–126 CrossRef CAS PubMed.
- A. Mirnajafi, J. Raymer, M. J. Scott and M. S. Sacks, Biomaterials, 2005, 26, 795–804 CrossRef CAS PubMed.
- H. R. Kayed, K. H. Sizeland, N. Kirby, A. Hawley, S. T. Mudie and R. G. Haverkamp, RSC Adv., 2015, 5, 3611–3618 RSC.
- P. P. Purslow, T. J. Wess and D. W. L. Hukins, J. Exp. Biol., 1998, 201, 135–142 CAS.
- J. Liao, L. Yang, J. Grashow and M. S. Sacks, Acta Biomater., 2005, 1, 45–54 CrossRef PubMed.
- T. W. Gilbert, S. Wognum, E. M. Joyce, D. O. Freytes, M. S. Sacks and S. F. Badylak, Biomaterials, 2008, 29, 4775–4782 CrossRef CAS PubMed.
- M. M. Basil-Jones, R. L. Edmonds, S. M. Cooper and R. G. Haverkamp, J. Agric. Food Chem., 2011, 59, 9972–9979 CrossRef CAS PubMed.
- K. H. Sizeland, M. M. Basil-Jones, R. L. Edmonds, S. M. Cooper, N. Kirby, A. Hawley and R. G. Haverkamp, J. Agric. Food Chem., 2013, 61, 887–892 CrossRef CAS PubMed.
- M. M. Basil-Jones, R. L. Edmonds, G. E. Norris and R. G. Haverkamp, J. Agric. Food Chem., 2012, 60, 1201–1208 CrossRef CAS PubMed.
- H. C. Wells, K. H. Sizeland, N. Kirby, A. Hawley and S. Mudie, ACS Biomater. Sci. Eng., 2015, 1, 1026–1038 CrossRef CAS.
- N. Nwaejike and R. Ascione, J. Card. Surg., 2011, 26, 31–33 CrossRef PubMed.
- D. Mavrilas, E. A. Sinouris, D. H. Vynios and N. Papageorgakopoulou, J. Biomech., 2005, 38, 761–768 CrossRef CAS PubMed.
- P. R. Umashankar, T. Arun and T. V. Kumari, J. Biomed. Mater. Res., Part A, 2011, 97, 311–320 CrossRef CAS PubMed.
- D. Cookson, N. Kirby, R. Knott, M. Lee and D. Schultz, J. Synchrotron Radiat., 2006, 13, 440–444 CrossRef CAS PubMed.
- M. S. Sacks, D. B. Smith and E. D. Hiester, Ann. Biomed. Eng., 1997, 25, 678–689 CrossRef CAS PubMed.
- R. H. Stinson and P. R. Sweeny, Biochim. Biophys. Acta, 1980, 621, 158–161 CrossRef CAS.
- R. G. Haverkamp, Chem. N. Z., 2013, 77, 12–17 CAS.
- A. Ward, F. Hilitski, W. Schwenger, D. Welch, A. W. C. Lau, V. Vitelli, L. Mahadevan and Z. Dogic, Nat. Mater., 2015, 14, 583–588 CrossRef CAS PubMed.
- K. H. Sizeland, H. C. Wells, G. E. Norris, R. L. Edmonds, N. Kirby, A. Hawley, S. Mudie and R. G. Haverkamp, J. Am. Leather Chem. Assoc., 2015, 110, 66–71 CAS.
- J. E. Scott and A. M. Thomlinson, J. Anat., 1998, 192, 391–405 CAS.
- J. E. Scott, FASEB J., 1992, 6, 2639–2645 CAS.
| This journal is © The Royal Society of Chemistry 2015 |