
Hyperlipidemia affects the absorption, distribution and excretion of seven catechins in rats following oral administration of tea polyphenols†
Abstract
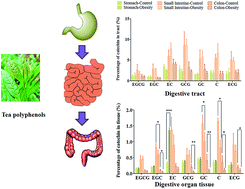
To explore the effects of hyperlipidemia on the pharmacokinetics of tea polyphenols, a comparative pharmacokinetics study of seven catechins between normal and obese rats was conducted. Rats were orally administered tea polyphenols (350 mg kg−1) and plasma, stomach, small intestine and colon samples of rats were obtained at 5, 30, 120, 360 and 720 min post administration. The plasma levels of (−)-gallocatechin from obese rats were significantly lower than those of normal rats. During the digestion of tea polyphenols in vivo, compared to normal rats, the levels of seven catechins within the gastric content and tissue of obese rats were significantly increased, in addition to the small intestinal tissue levels of (−)-epigallocatechin gallate and (−)-gallocatechin gallate. On the contrary, the colonic tissue levels of (−)-epigallocatechin, (−)-gallocatechin gallate, (−)-gallocatechin and (+)-catechin in obese rats were significantly decreased compared to the levels in normal rats. Furthermore, the fecal excretion of the seven catechins in obese rats was highly increased. To sum up, hyperlipidemia changed the pharmacokinetics of catechins by increasing their distribution in the stomach and small intestine, but decreasing their distribution in the colon.
 Please wait while we load your content...
Please wait while we load your content...

