
Antioxidant and hepatoprotective effects of the food seasoning curry leaves Murraya koenigii (L.) Spreng. (Rutaceae)
Abstract
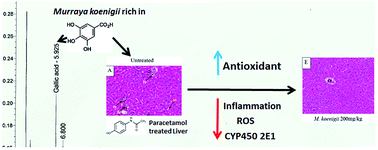
Murraya koenigii (L.) Spreng. (Rutaceae), a common spice, has been traditionally used to reduce inflammation and hepatitis. The present study aimed to reveal the antioxidant and anti-inflammatory activity as well as the regulation of cytochrome P450 levels elicited by aqueous extracts of M. koenigii leaves in response to paracetamol-induced liver toxicity in BALB/c mice. Liver toxicity was induced by an overdose of paracetamol followed by treatment with a M. koenigii leaf aqueous extract. The levels of serum liver markers, liver antioxidants, inflammatory markers and liver cytochrome P450 2E1 were quantified after 14 days of treatment. Histopathological analysis of the liver was also carried out. In vitro antioxidant levels and phenolic acid characterization were also performed. The extracts (50 and 200 mg kg−1 body weight) effectively restored the serum liver profiles (alanine transaminase, aspartate transaminase and alkaline phosphatase), liver antioxidant levels (superoxide dismutase, glutathione and ferric reducing ability of plasma) and inflammatory markers (tumor necrosis factor alpha, inducible nitric oxide synthase, nuclear factor kappa-light-chain-enhancer of activated B cells and nitric oxide) to healthy levels in a dosage dependent manner. The level of liver cytochrome P450 2E1 was also lowered in the extract treated groups. Histopathological assessment showed that treatment with 200 mg kg−1 of the M. koenigii aqueous extract was able to reduce liver necrosis in mice fed paracetamol. Gallic acid concentration was the highest among all the phenolic acids detected in the extract. These results suggested that the M. koenigii aqueous extract, which possessed antioxidant and anti-inflammatory effects, can be used as a potential treatment for liver diseases caused by oxidative stress.
 Please wait while we load your content...
Please wait while we load your content...

