
Effect of glucose tolerance factor (GTF) from high chromium yeast on glucose metabolism in insulin-resistant 3T3-L1 adipocytes
Abstract
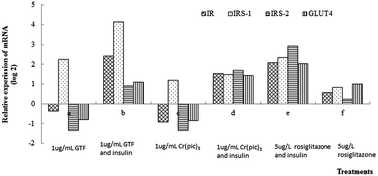
The purpose of this paper was to assess and compare the impact of GTF, CrCl3 and Cr(pic)3 on glucose metabolism and explore the underlying mechanism of GTF in insulin-resistant 3T3-L1 adipocytes. The insulin-resistant 3T3-L1 adipocytes were induced by incubation with insulin for 48 h. Purified GTF from high chromium yeast was used in this study, with a m/z of 769 to 712, and glutamic acid, glycine, and cysteine in an approximate ratio of 1 : 1 : 1. In addition nicotinic acid and Cr(III). GTF, CrCl3, Cr(pic)3 and rosiglitazone (positive control) were applied to the cells. The effective dose of GTF ranged from 0.5 μg mL−1 to 1.5 μg mL−1. GTF decreased cell viability significantly (P < 0.01) at doses of 3 μg mL−1 or higher. Glucose consumption in insulin-resistant 3T3-L1 adipocytes induced by GTF increased significantly (P < 0.05) when incubated with GTF after 12 h. Among GTF, Cr(pic)3 and CrCl3, GTF stimulated glucose consumption is the greatest. In the presence of insulin, the relative expression level of insulin receptor (IR), insulin receptor substrate-1 (IRS-1), insulin receptor substrate-2 (IRS-2) and glucose transporter 4 (GLUT4) mRNA were increased by GTF by 2.4, 4.1, 0.9 and 1.1-fold, respectively, however, only IRS-1 was increased by 2.3-fold in the absence of insulin. GTF affected mRNA levels of IR and IRS-1 significantly (P < 0.01) as compared to the other two. This study not only further demonstrates that chromium containing complexes show promise in reducing insulin resistance in instances of type 2 diabetes, but also that among the chromium complexes, GTF performs the best. Additionally, new mechanistic details of how GTF affects mRNA levels of insulin signalling proteins were revealed.
 Please wait while we load your content...
Please wait while we load your content...

