Self-assembly of Cu(II) with amyloid β19–20 peptide: relevant to Alzheimer's disease†
Xiaofang Shen*,
Xiaorong Deng and
Yuehong Pang
State Key Laboratory of Food Science and Technology, School of Food Science and Technology, Jiangnan University, Wuxi, Jiangsu 214122, P.R. China. E-mail: xfshen@jiangnan.edu.cn
First published on 8th May 2014
Abstract
Amyloid-β (Aβ) peptide Aβ19–20 tends to self-assemble into well-ordered tubular structures under physiological conditions, while in the presence of equimolar Cu(II), they assembled into homogeneous microvesicles; a chelator can competitively bind to Cu(II) from the Cu(II)–Aβ19–20 complex, resulting in reforming of the peptide tubular structure.
Alzheimer's Disease (AD) is an age-related neurodegenerative disorder characterized by a progressive loss of memory and the deterioration of higher cognitive functions.1 More than 26 million people worldwide are afflicted by AD. However, no drugs or therapeutics have thus far been developed to treat AD.2 The self-assembly of amyloid-β (Aβ) peptides into highly ordered amyloid fibril structures represents one of the pathological hallmarks of AD.3 Under normal conditions, about 90% of secreted Aβ peptides are Aβ1–40, species that are highly fibrillogenic and deposited early in individuals with AD.4 The central seven-residue segment Aβ16–22 is thought to be crucial for amyloid fibril formation. While the forepost of such sequences in the full-length peptide do not have that propensity.5 Furthermore, the amyloid fibril formation of other short beta peptide fragments, such as Aβ1–28,6 Aβ11–17,7 Aβ16–20,8 Aβ16–22,9 Aβ25–35,10 Aβ31–35 (ref. 11) and Aβ39–42 (ref. 12) are also reported. Short peptides serve as an excellent model system for exploring amyloid fibrils formation in particular and biological self-assembly processes in general.13
Since its discovery as a core recognition motif of the Alzheimer's Aβ peptide, Aβ19–20 (diphenylalanine, Phe–Phe) has increasingly become the subject of intense investigation in the fields of both medicine and nanotechnology.14 It has been demonstrated that Aβ19–20 tends to self-assemble into well-ordered nanotubes. The formed tubular structures may share some structural properties with the amyloid fibrils as they have similar vibrational spectrum and birefringence as compared to the fibrils.15 This indicates that the dipeptide aromatic motifs contain all the molecular information needed to form wellordered-supramolecular structures at the nano-scale. One striking property of the peptide nanotubes is their remarkable rigidity, with a high Young's modulus of 19–27 GPa using two independent techniques.16 Scientists try to find out why the peptide nanostructures are so rigid.17 However, little is known about the forces underlying their stability and relationship with the AD. The π–π stacking interactions may accelerate amyloid fibril formation by providing geometrical restrictions that promote directionality and orientation of the growing fibril, together with energetic contributions from the stacking itself.18 Indeed, replacement of the phenylalanine residues at the C terminus of Aβ1–42 by hydrophobic residues resulted in somewhat slower aggregation, while a mutant variant containing four phenylalanine residues at the C terminus of Aβ1–42 displayed faster amyloidogenesis.19
In an aging brain, metal ions might have a relevant role in the development and progression of the AD disease.20 The disordering process associated with Aβ aggregation of AD is greatly influenced by the metal ions (i.e. Cu, Al, Fe and Zn). The metals are found in both the core and rim of the AD senile plaques. Analysis of the autopsy of AD patients shows abnormally high levels of Cu (∼393 μM) present in the senile plaques; the normal cerebral extracellular concentration is 0.2–1.7 μM.21 The vitro studies also showed that Cu(II) would influence the formation of amyloid fibrils.20c,22 Cu(II) has been proposed to play a central role in the amyloid cascade process linked to the development of AD. Involvement in both the Aβ aggregation process and reactive oxygen species production has been considered. Researchers have accordingly put forth the “metals hypothesis” of AD, which postulates that compounds designed to inhibit Cu(II)/Aβ interactions and redistribute Cu(II) may offer therapeutic potential for treating AD.23
The involvement of Cu(II) with Aβ in AD and the potential development of metal chelating therapy indicate the importance of elucidating the fundamental mechanisms of Cu(II) effects on short peptide model Aβ19–20. Herein, we investigate the Cu(II) ion effects on self-assembly of Aβ19–20 peptide and the ability of ethylene diamine tetraacetic acid (EDTA) to compete chelating Cu(II) and then recover the structure of Aβ19–20 peptide (Fig. 1A). Actually, we observed the visual change in solutions at the beginning. The addition of Cu(II) to Aβ19–20 made the solution appear dark blue. When equimolar EDTA was added to the above mixed solution, the color of the solution became greenish blue, which was consistent with that of Cu(II) with EDTA (Fig. 1B).
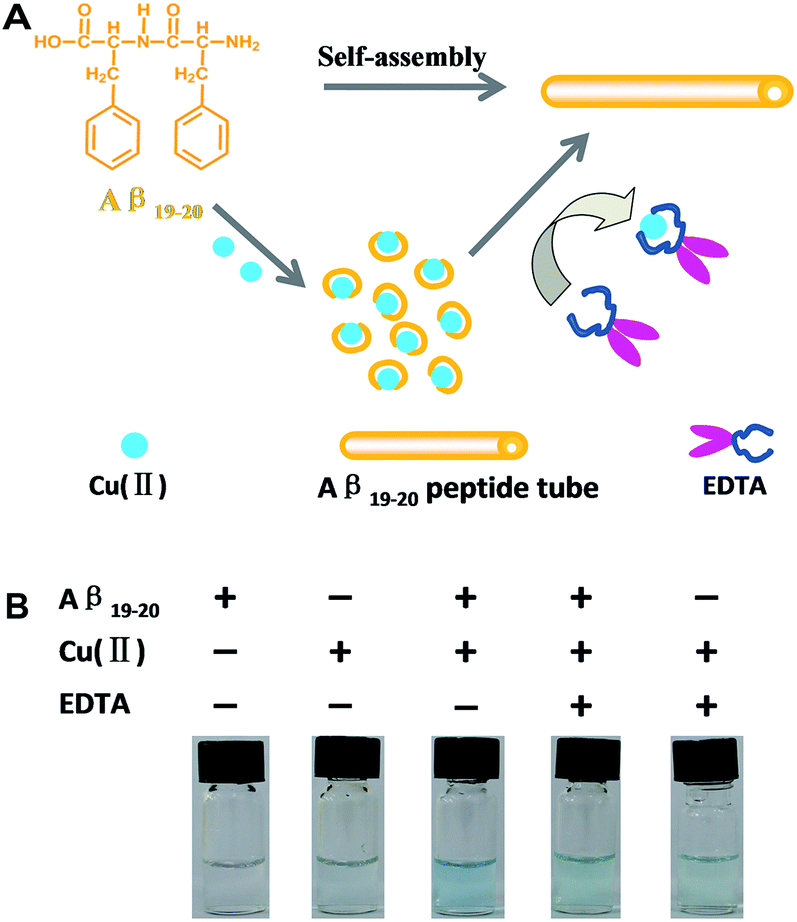 | ||
| Fig. 1 Schematic representation of (A), Aβ19–20 peptide assembly with Cu(II) ions and then EDTA competes to chelate Cu(II), meanwhile (B) the visible change in solutions. | ||
Fig. 2 shows the effect of Cu(II) on the self-assembled Aβ19–20 tubes aggregates overnight. We could observe the Aβ19–20 peptide formed tubes on the solid substrate (Fig. 2 A and C), as reported previously by Gazit group.14a The average diameter of the tubes was about 1.0–2.5 μm with a long persistence length, ranging from approximately 100 μm to millimeter. However, in the presence of Cu(II) as molecule ratio to Aβ19–20 is 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1, we could not observe tubular structure (Fig. 2B and D). Instead, abundant uniform microvesicles with an average diameter about 0.8–5.0 μm were observed. The transition from tubular structures to dense microvesicles evident from the scanning electron microscopy (SEM) images illustrate that Cu(II) inhibited original peptide tube assembly and formed a new structure.
:1, we could not observe tubular structure (Fig. 2B and D). Instead, abundant uniform microvesicles with an average diameter about 0.8–5.0 μm were observed. The transition from tubular structures to dense microvesicles evident from the scanning electron microscopy (SEM) images illustrate that Cu(II) inhibited original peptide tube assembly and formed a new structure.
 | ||
| Fig. 2 (A) Self-assembled structures of Aβ19–20, (B) 1:1 molar ratio of Cu(II) to Aβ19–20. (C) and (D) are SEM enlarged images of (A) and (B), respectively. Scale bar: (A) and (B) are 10 μm, (C) and (D) are 5 μm. | ||
Cu(II) induced π–π stacking changes of Aβ19–20 peptide: We attribute the visual changes in solution and rearrangement of the structures to the π–π interactions of the aromatic rings on phenylalanine. Aβ19–20 peptide features aromatic moieties which play a key role in the fibril self-assembly process and drive a well-ordered stacking of aromatic rings by π–π interactions.14a,e The UV-vis spectra of various molar ratio of Cu(II) to Aβ19–20: 0:1, 1:4, 1:2, 1:1 and 2:1 in pH 7.4 Tris–HCl buffer are shown in Fig. 3. The absorption peak of Aβ19–20 is at 258 nm. It is known that the absorption is attribute to benzene ring characteristic absorption band, which corresponds to the π–π* transition and the vibration of phenyl rings. The addition of the Cu(II) noticeably increased the absorption of Aβ19–20 peptide (Fig. 3A). The maximum response was observed when molar ratio of Cu(II) to Aβ19–20 reached 1:1. The result shows that Cu(II) interacted with Aβ19–20 peptide and, to some extent, influenced the vibration of Aβ19–20 phenyl rings.
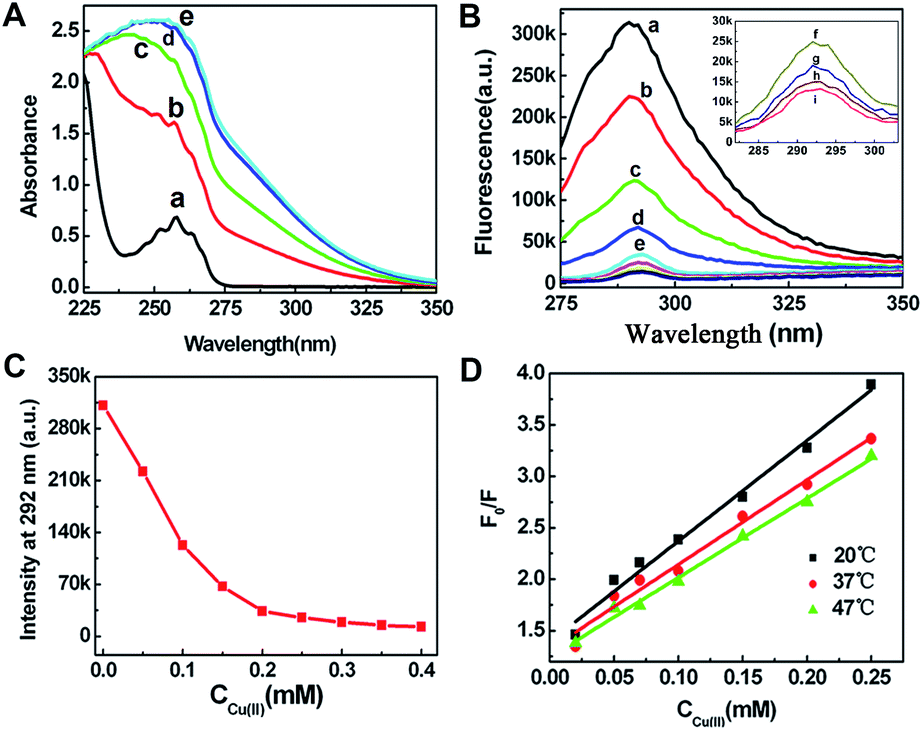 | ||
| Fig. 3 (A) UV-vis spectra of Aβ19–20 at 258 nm with various molar ratio of Cu(II) to Aβ19–20. From a to e: 0:1, 1:4, 1:2, 1:1 and 2:1. (B) Fluorescence titration spectra with different molar ratio of Cu(II) to Aβ19–20. From (a) to (i): 0:1, 1:4, 1:2, 1:1.3, 1:1, 1.25:1, 1.5:1, 1.75:1 and 2:1. Inset is enlarged from (f) to (i). (C) Nonlinear fitting curve of Aβ19–20 peptide binding to Cu(II). (D) The Stern–Volmer plots for the quenching of fluorescence by Cu(II) at 20, 37 and 47 °C. | ||
Further evidence of the involvement of aromatic interactions with Cu(II) was provided by fluorescence spectra, which is the crucial to the phenylalanine dipeptide assembly. Fig. 3B shows the fluorescence titration spectra of Aβ19–20 peptide with different concentrations of Cu(II) solution. Aβ19–20 possesses two phenylalanine residues that emitted fluorescence with the maximum at 292 nm upon excitation at 265 nm, which was belonging to the π–π stacking interactions.24 The decrease in the fluorescence intensity was the most marked change with the successive addition of Cu(II), indicating that the fluorescence of Aβ19–20 was quenched. The curve has a tendency towards invariability at Cu(II)/Aβ19–20 molar ratio of 1:1(Fig. 3C), indicating the Cu(II) influenced the π–π stacking of Aβ19–20.
Intrinsic peptide fluorescence is a useful and sensitive tool to study variations in a peptide conformation and its interactions with other molecules.7 To understand the interaction stoichiometry of Aβ19–20 peptide with Cu(II) ions, fluorescence titration method was used and the nonlinear fitting curve of Aβ19–20 peptide binding to Cu(II) (ESI†). The fluorescence titration data processing can be described as follows:22b,25 Assuming a protein-to-ligand binding reaction (P + nL ↔ PLn), the fraction of bound ligand (γ) can be related to the equilibrium dissociation constant (Kd), as shown in eqn (1).
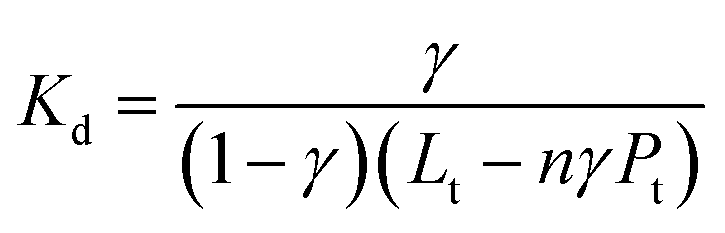 | (1) |
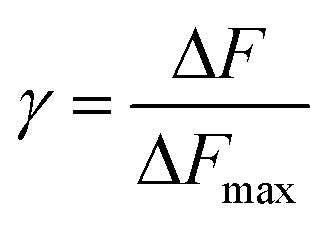 | (2) |
Combining eqn (1) and (2) yields (3):
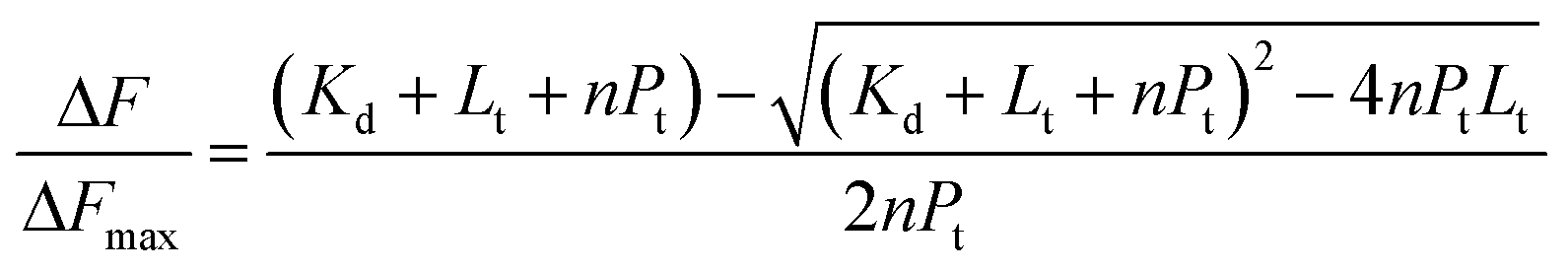 | (3) |
According to eqn (3), Kd can be obtained. The fractions of Aβ19–20 bound with Cu(II) were plotted against the Cu(II) concentration. The data could be fitted to a single ligand binding with the equilibrium dissociation constant, Kd = 4.24 μM, suggesting a high affinity of Aβ19–20 peptide with Cu(II).
Cu(II) ion caused Aβ19–20 peptide endogenous fluorescence quenching regularly, but did not influence the excited fluorescence molecular state, thus altering the absorption spectrum. This may be due to Cu(II) associated with Aβ19–20 and complex form, resulting in peptide fluorescence static quenching. Static quenching and dynamic quenching vary temperature. The quenching rate constants decrease with increasing temperature for static quenching, but the reverse effect is observed for the case of dynamic quenching.25 To further characterize the quenching mold, a possible quenching mechanism was evident from the Stern–Volmer curves of Aβ19–20 with Cu(II) at different temperatures (20, 37 and 47 °C) as shown in Fig. 3D. The Stern–Volmer plots were linear, the slopes of which decreased with increasing temperature. This indicated the occurrence of a static quenching interaction between Aβ19–20 and Cu(II). Quenching data were then fit into the Stern–Volmer eqn (4).
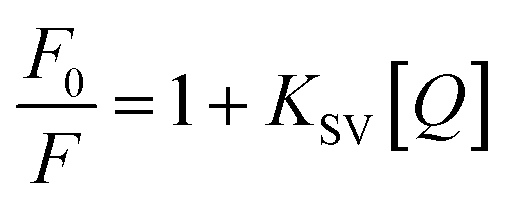 | (4) |
To demonstrate that Cu(II) influenced and mediated the assembly of Aβ19–20 peptides, we performed chelation competition experiments with chelator. EDTA exerting an antagonizing effect on the Cu(II)-induced changes of the intrinsic fluorescence of Aβ19–20 peptide. The intrinsic fluorescence of Aβ19–20 peptide was gradually recovered and reached maximum at 48 h (ESI†). Morphological changes also reveal the competitive chelation of the chelator to Cu(II) from Aβ19–20 (Fig. 4). Co-existed 3.2 mM Aβ19–20 peptide and equimolar Cu(II) self-assembled into homogeneous microvesicles (Fig. 4A). Interestingly, when the equimolar chelating agent EDTA was injected into Cu(II) and Aβ19–20 peptide mixed solution, enabling it to chelate Cu(II) from Cu(II)–Aβ19–20 complex. As a result, microvesicles disappeared and the peptide tubular structures were obtained (Fig. 4B).
 | ||
| Fig. 4 (A) Microvesicles self-assembled by Aβ19–20 peptide with equimolar Cu(II). (B) Tubular structures of Aβ19–20 reformed in the presence of EDTA. Scale bar is 10 μm. | ||
Cu(II) ions interact with Aβ19–20 peptide and form Cu(II)–Aβ19–20 complex, which influenced π–π stacking of Aβ19–20 peptide and induced its morphology from dense tubes to an homogeneous microvesicles. Chelator can completely chelate Cu(II) from Cu(II)–Aβ19–20 complex and reform Aβ19–20 peptide tubular structure. These results facilitate the understanding of the clinically related Cu(II) ions with Aβ in AD and provide a rationalized mechanism for fibrils formation. However, due to the complex etiology of AD, a simple interaction model currently is not possible for fully understanding the disease. On the one hand, high-resolution structural studies of amyloid fibrils have made huge progress in recent years, such as utilizing a variety of 13C/15N-labeling strategies and combined with multidimensional magic angle spinning (MAS) NMR techniques,26 and the combination of electron paramagnetic resonance (EPR) spectroscopy with isotopic labelling.27 These techniques can unambiguously determine the Cu2+/Aβ coordination to provide credible new insights into the assembly. On the other hand, development of novel classes of potential therapeutics using the tools of coordination chemistry to create synthetic ligands for competing metal ions with Aβ, one can hope these will actually derail these diseases in the future.
This work was supported by the National Natural Science Foundation of China (21005032) and the Natural Science Foundation of Jiangsu Province (BK2012107).
Notes and references
- (a) N. C. Berchtold and C. W. Cotman, Neurobiol. Aging, 1998, 19, 173–189 CrossRef CAS; (b) R. D. Terry and R. Katzman, Ann. Neurol., 1983, 14, 497–506 CrossRef CAS PubMed.
- (a) D. Kim, N. H. Kim and S. H. Kim, Angew. Chem., Int. Ed., 2013, 52, 1139–1142 CrossRef CAS PubMed; (b) M. A. Telpoukhovskaia and C. Orvig, Chem. Soc. Rev., 2013, 42, 1836–1846 RSC.
- (a) M. P. Murphy and H. LeVine, J. Alzheimer's Dis., 2010, 19, 311–323 Search PubMed; (b) C. Haass and D. J. Selkoe, Nat. Rev. Mol. Cell Biol., 2007, 8, 101–112 CrossRef CAS PubMed; (c) W. Bondareff, J. Alzheimer's Dis., 2013, 35, 1–6 CAS; (d) I. W. Hamley, Chem. Rev., 2012, 112, 5147–5192 CrossRef CAS PubMed.
- S. Kar, S. P. M. Slowikowski, D. Westaway and H. T. J. Mount, J. Neuropsychiatry Clin. Neurosci., 2004, 29, 427–441 Search PubMed.
- C. SivakamaSundari, S. Rukmani and R. Nagaraj, J. Pept. Sci., 2012, 18, 122–128 CrossRef CAS PubMed.
- T. Wasiak, M. Ionov, K. Nieznanski, H. Nieznanska, O. Klementieva, M. Granell, J. Cladera, J.-P. Majoral, A. M. Caminade and B. Klajnert, Mol. Pharmaceutics, 2012, 9, 458–469 CrossRef CAS PubMed.
- C. He, Y. Han, Y. Fan, M. Deng and Y. Wang, Langmuir, 2012, 28, 3391–3396 CrossRef CAS PubMed.
- S. Dehn, V. Castelletto, I. W. Hamley and S. Perrier, Biomacromolecules, 2012, 13, 2739–2747 CrossRef CAS PubMed.
- N. Chaudhary and R. Nagaraj, J. Pept. Sci., 2011, 17, 115–123 CrossRef CAS PubMed.
- M. Naldi, J. Fiori, M. Pistolozzi, A. F. Drake, C. Bertucci, R. Wu, K. Mlynarczyk, S. Filipek, A. De Simone and V. Andrisano, ACS Chem. Neurosci., 2012, 3, 952–962 CrossRef CAS PubMed.
- Y. Kuroki, Y. Sasaki, D. Kamei, Y. Akitake, M. Takahashi, S. Uematsu, S. Akira, Y. Nakatani, I. Kudo and S. Hara, Biochem. Biophys. Res. Commun., 2012, 424, 409–413 CrossRef CAS PubMed.
- M. M. Gessel, C. Wu, H. Li, G. Bitan, J.-E. Shea and M. T. Bowers, Biochemistry, 2012, 51, 108–117 CrossRef CAS PubMed.
- M. Reches and E. Gazit, Curr. Nanosci., 2006, 2, 105–111 CrossRef CAS.
- (a) M. Reches and E. Gazit, Science, 2003, 300, 625–627 CrossRef CAS PubMed; (b) E. Gazit, Chem. Soc. Rev., 2007, 36, 1263–1269 RSC; (c) X. Yan, P. Zhu and J. Li, Chem. Soc. Rev., 2010, 39, 1877–1890 RSC; (d) I. Cherny and E. Gazit, Angew. Chem., Int. Ed., 2008, 47, 4062–4069 CrossRef CAS PubMed; (e) M. Reches and E. Gazit, Nat. Nanotechnol., 2006, 1, 195–200 CrossRef CAS PubMed.
- E. Gazit, Prion, 2007, 1, 32–35 CrossRef.
- (a) X. H. Yan, P. L. Zhu and J. B. Li, Chem. Soc. Rev., 2010, 39, 1877–1890 RSC; (b) N. Kol, L. A. Abramovich, D. Barlam, R. Z. Shneck, E. Gazit and I. Rousso, Nano Lett., 2005, 5, 1343–1346 CrossRef CAS; (c) L. Niu, X. Chen, S. Allen and S. J. B. Tendler, Langmuir, 2007, 23, 7443–7446 CrossRef CAS PubMed.
- I. Azuri, L. Adler-Abramovich, E. Gazit, O. Hod and L. Kronik, J. Am. Chem. Soc., 2014, 136, 963–969 CrossRef CAS PubMed.
- I. Cherny and E. Gazit, Angew. Chem., Int. Ed., 2008, 47, 4062–4069 CrossRef CAS PubMed.
- W. Kim and M. H. Hecht, Proc. Natl. Acad. Sci. U. S. A., 2006, 103, 15824–15829 CrossRef CAS PubMed.
- (a) P. Zatta, D. Drago, S. Bolognin and S. L. Sensi, Trends Pharmacol. Sci., 2009, 30, 346–355 CrossRef CAS PubMed; (b) M. A. Telpoukhovskaia and C. Orvig, Chem. Soc. Rev., 2013, 42, 1836–1846 RSC; (c) F. Hane, G. Tran, S. J. Attwood and Z. Leonenko, PLoS One, 2013, 8, e59005 CAS; (d) D. Pramanik, C. Ghosh, S. Mukherjee and S. G. Dey, Coord. Chem. Rev., 2013, 257, 81–92 CrossRef CAS PubMed.
- (a) C. Bouras, P. Giannakopoulos, P. F. Good, A. Hsu, P. R. Hof and D. P. Perl, Eur. Neurol., 1997, 38, 53–58 CrossRef CAS PubMed; (b) M. A. Lovell, J. D. Robertson, W. J. Teesdale, J. L. Campbell and W. R. Markesbery, J. Neurol. Sci., 1998, 158, 47–52 CrossRef CAS.
- (a) S. Bolognin, L. Messori, D. Drago, C. Gabbiani, L. Cendron and P. Zatta, Int. J. Biochem. Cell Biol., 2011, 43, 877–885 CrossRef CAS PubMed; (b) W. T. Chen, Y. H. Liao, H. M. Yu, I. H. Cheng and Y. R. Chen, J. Biol. Chem., 2011, 286, 9646–9656 CrossRef CAS PubMed; (c) B. Alies, E. Renaglia, M. Rozga, W. Bal, P. Faller and C. Hureau, Anal. Chem., 2013, 85, 1501–1508 CrossRef CAS PubMed.
- (a) Y. Jiao and P. Yang, J. Phys. Chem. B, 2007, 111, 7646–7655 CrossRef CAS PubMed; (b) S. C. Drew, C. J. Noble, C. L. Masters, G. R. Hanson and K. J. Barnham, J. Am. Chem. Soc., 2009, 131, 1195–1207 CrossRef CAS PubMed; (c) C. Hureau, Coord. Chem. Rev., 2012, 256, 2164–2174 CrossRef CAS PubMed; (d) S. C. Drew and K. J. Barnham, Acc. Chem. Res., 2011, 44, 1146–1155 CrossRef CAS PubMed; (e) H. A. Feaga, R. C. Maduka, M. N. Foster and V. A. Szalai, Inorg. Chem., 2011, 50, 1614–1618 CrossRef CAS PubMed.
- P. Zhu, X. Yan, Y. Su, Y. Yang and J. Li, Chem.–Eur. J., 2010, 16, 3176–3183 CrossRef CAS PubMed.
- Y. R. Chen and A. C. Clark, Protein Sci., 2004, 13, 2196–2206 CrossRef CAS PubMed.
- Y. C. Su, C. J. Sarell, M. T. Eddy, G. T. Debelouchina, L. B. Andreas, C. L. Pashley, S. E. Radford and R. G. Griffin, J. Am. Chem. Soc., 2014, 136, 6313–6325 CrossRef CAS PubMed.
- S. C. Drew and K. J. Barnham, Acc. Chem. Res., 2011, 44, 1146–1155 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c4ra02758b |
| This journal is © The Royal Society of Chemistry 2014 |