
Self-assembly of Cu(ii) with amyloid β19–20 peptide: relevant to Alzheimer's disease†
Abstract
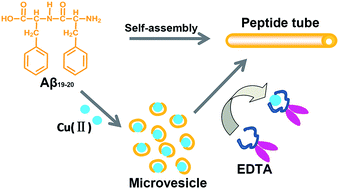
Amyloid-β (Aβ) peptide Aβ19–20 tends to self-assemble into well-ordered tubular structures under physiological conditions, while in the presence of equimolar Cu(II), they assembled into homogeneous microvesicles; a chelator can competitively bind to Cu(II) from the Cu(II)–Aβ19–20 complex, resulting in reforming of the peptide tubular structure.
- This article is part of the themed collection: Towards understanding and treating Alzheimer’s disease
 Please wait while we load your content...
Please wait while we load your content...

