Strategies for the consolidation of biologically mediated events in the conversion of pre-treated lignocellulose into ethanol
Abdul Waheed Bhutto
*ab,
Khadija Qureshi
a,
Khanji Harijan
c,
Gholamreza Zahedi
d and
Alireza Bahadori
e
aDepartment of Chemical Engineering, Mehran University of Engineering and Technology, Jamshoro, Sindh, Pakistan
bDepartment of Chemical Engineering, Dawood University of Engineering & Technology, Karachi, Pakistan. E-mail: abdulwaheed27@hotmail.com; Fax: +922199231195; Tel: +922199231197
cDepartment of Mechanical Engineering, Mehran University of Engineering and Technology, Jamshoro, Sindh, Pakistan
dDepartment of Chemical and Biochemical Engineering, Missouri University of Science & Technology, Rolla, MO 65409, USA. E-mail: zahedig@mst.edu; grzahedi@yahoo.com; Fax: +1573-341-4377; Tel: +1573-341-4423
eSouthern Cross University, School of Environment, Science and Engineering, Lismore, NSW, Australia
First published on 21st October 2013
Abstract
Notwithstanding the plentiful published work on the production of ethanol from lignocellulosic materials this comprehensive review relates how the basic research of a commercially viable industrial production strategy is still lacking. The objective of this review is to compile information on the different strategies and to consolidate the biologically mediated events involved in the conversion of pre-treated lignocellulose to ethanol and associated expenses so that some generalized information can be developed that could help policy makers and other stakeholders in designing a policy framework to promote second generation biofuels. The review also discusses the potential of process integration, its cost competitiveness and role in establishing commercial facilities for the production of ethanol from lignocellulosic biomass.
 Abdul Waheed Bhutto | Abdul Waheed Bhutto is an Assistant Professor at Dawood University of Engineering & Technology, Karachi. He is currently a Ph.D. candidate at Mehran University-Jamshoro. Under the guidance of Khadija Qureshi he is concentrating his efforts on improving the efficiency of processes for biofuel production from lignocellulosic material. He received his Master's Degree from NED University Karachi in 2001. He earned his Bachelor’s degree in chemical engineering from Dawood Engineering College-Karachi in 1998. His research work has focused on alternate energy sources for sustainable development. |
 Khadija Qureshi | Khadija Qureshi is a Professor at Mehran University-Jamshoro. She has an extensive research and teaching background in chemical and environmental engineering. For 15 years she has supervised research projects on the reuse of waste materials and agricultural waste, the conversion of agricultural waste into useful chemicals and liquid fuels. She is currently a Post doctorate fellow at the University of Arizona working on the US Science and Technology Program. She is also a key trainer in gender and mainstreaming women in leadership and management. |
 Khanji Harijan | Khanji Harijan is a Professor at Mehran University-Jamshoro. He received his Ph.D. from Mehran University-Jamshoro in 2008 where he worked on the modeling and analysis of the potential demand for renewable sources of energy in Pakistan. He has an extensive research and teaching background in renewable energy. He is the author of over 100 articles and book chapters. His current research focuses on identifying and addressing barriers to renewable energy development. |
 Gholamreza Zahedi | Gholamreza Zahedi is currently a visiting professor at the Missouri University of Science and Technology, Rolla, USA. He was an Associate Professor at UTM, Malaysia. He is a chartered engineer from IChemE UK and a certified energy manger. Dr Zahedi is an author of 81 journal papers, 4 books and 3 book chapters. Dr Zahedi is a researcher and consultant in engineering processes related to sustainability. Recently Dr Zahedi has started reviewing and analysing governmental policies around the world as a key decision maker for their societies. |
 Alireza Bahadori | Alireza Bahadori, is a researcher at Southern Cross University, NSW, Australia. He received his Ph.D. from Curtin University, Australia. He is an author of over 250 articles and 12 books. His books have been published by prestigious publishers such as John Wiley & Sons, Springer, Taylor & Francis and Elsevier. Dr Bahadori is the recipient of the prestigious Australian Government's Endeavors International Postgraduate award and an award from the State Government of Western Australia through the Western Australia Energy research Alliance (WA:ERA). |
Introduction
Research is in progress worldwide to find economically viable and environmentally friendly alternatives for the quickly depleting fossil fuels.1 Biomass based fuels currently seem to be the most suitable alternative for sustainable development in the context of economic and environmental considerations.2,3 Biomass based resources are evenly distributed and easily available throughout the world. Lignocellulosic biomass can also grow on marginal or non-arable lands without displacing food crops.The utilization of agricultural waste in the production of valuable materials also simultaneously allows the disposal of agricultural waste. However the transformation of agricultural waste into efficient liquid fuel is a very expensive process, which hinders the large scale commercial production.4–6 Rational work is required to find out methods to improve the economics of the conversion process.
The production of fuel ethanol from sugars present in lignocellulosic biomass is still a challenge with many opportunities for progress. Robust microorganisms with higher rates of conversion and yield are required to allow process simplification through consolidating process steps. Such developments are likely to reduce both capital and operating costs to make the process economically viable. Abundant biomass has the potential to serve as a source of carbon based feedstock for fuel and chemical production in much the same way as crude oil presently serves as the carbon feedstock in petrochemical refineries.
It is estimated that for each billion of ethanol-equivalent litres of fuel produced and burned in the US, the combined climate-change and health costs are $123.89 million for gasoline, $124.6–251.49 million for corn ethanol, but only $32.49–55 million for cellulosic ethanol.7,8
The world is working to identify and optimize strategies for the economic conversion of lignocellulosic biomass into fuels, which will subsequently lead to bio based chemicals. In general, lignocellulosic feedstocks are converted into second generation biofuels through two distinct conversion routes, namely the thermo chemical and the biochemical one.9 Biotransformation mediated by microorganisms or enzymes has the advantages of low energy consumption, modest reaction conditions, high selectivity, and the low costs of a bioreactor.10 The literature also suggests the biochemical route has more potential for cost reduction due to its most recent developments and the constant effort for optimization.11–15 This article only discusses the biochemical route.
The current technology for the biochemical conversion of biomass to ethanol consists of four major steps: pre-treatment, hydrolysis, fermentation, and product separation/purification. The pretreatment step opens up the structure of the lignocellulose to improve the accessibility to cellulose and hemicelluloses. These cellulose and hemicelluloses are cleaved to the monomers (sugars) during hydrolyses. The fermentation of sugar produces ethanol whose concentration is enhanced to the commercial level during product separation/purification steps.
The type of pretreatment defines the suitable hydrolysis step and composition of the hydrolysis product. Four biologically mediated events occur during the conversion of pre-treated lignocellulose to ethanol via processes featuring (i) enzymatic hydrolysis; (ii) the production of depolymerizing enzymes (cellulases and hemicellulases); (iii) hydrolysis of the polysaccharide constituents of pre-treated biomass; (iv) fermentation of the hexose and pentose sugars.
Improvements in biomass conversion technology generally entails the consolidation of two or more of these steps. Simultaneous saccharification and fermentation (SSF) combines the hydrolysis and fermentation of hexoses. Simultaneous saccharification and co-fermentation (SSCF) combines the hydrolysis and fermentation of both hexoses and pentoses. The ultimate objective may be a one-step consolidated bioprocessing (CBP) of lignocellulose to bioethanol, in which all four biologically mediated events occur in a single reactor where a single microorganism or microbial consortium converts the pre-treated biomass to ethanol. CBP may represent a breakthrough for low-cost biomass processing, owing to the economic benefits of process integration and avoiding the high costs of enzymes that make the biochemical conversion route unattractive. This article identifies areas of improvement and the way forward by a step involving the conversion of pretreated biomass into ethanol, with its commercial viability. The review also compiles information on different biochemical strategies for the production of ethanol and the associated costs to develop some generalized information that could help policy makers and other stakeholders in designing a policy framework to promote second-generation biofuels. The article focuses on optimizing the process parameters and process integration to improve economic performance, which is followed by a section that discusses the commercial cellulosic ethanol production. Finally, we draw some key conclusions that have implications for policy recommendations.
Pretreatment
Pretreatment is the initial and crucial step for lignocellulose biomass processing. There are several types of pre-treatment methods reported in the literature. These methods bring physical and/or chemical changes in the plant biomass in order to achieve a desired result.Typical goals of pre-treatment include (1) the production of highly digestible solids that enhance sugar yields during enzyme hydrolysis, (2) avoiding the degradation of sugars (mainly pentoses) including those derived from hemicellulose, (3) minimizing the formation of inhibitors for subsequent fermentation steps, (4) the recovery of lignin for conversion into valuable co-products, and (5) cost effectiveness by operating in reactors of moderate size and by minimizing heat and power requirements.16
Pre-treatment methods are selected based on the overall yield and economics, rather than the performance of an individual step. The final selection may vary from country to country based upon feedstock characteristics, their availability and the cost of pretreatment, hydrolysis, fermentation and rectification. The results of pretreatment are balanced against their impact on the cost of the downstream processing steps and the trade-off between operating costs, capital costs and biomass costs.
There are many excellent recent review papers on the pre-treatment of lignocellulose to provide basic knowledge about how biomass composition/structure affects various pretreatment methods.17–25 Table 1 summarizes a discussion of pretreatment methods in the literature.
| Pre-treatment process | Summary |
|---|---|
| Biological | The combination of a biological pre-treatment with a mild alkali pre-treatment significantly improves ethanol production, without producing significant inhibitors that hinder downstream processes. |
| Mechanical comminution | Careful selection of the equipment and final particle size reduces initial investment and energy requirements to improve the process economics. |
| Radiation | Microwave radiation. The microwave-assisted alkali pre-treatment removes more lignin and hemicellulose from wheat straw with a shorter pre-treatment time compared with the conventional alkali one. Likewise microwave heating significantly accelerates the dissolution of cellulose with ionic liquids. |
| Gamma radiation. Gamma irradiation after sulfuric acid/NaOH treatment has a substantial synergistic effect on enzymatic hydrolysis for bioethanol production. | |
| Electron beam irradiation. Electron beam irradiation significantly increases the enzyme digestibility of rice straw, while the crystallinity and enzyme digestibility were also found to be strongly correlated. | |
| The efficiency of radiation methods is very low. In addition these methods are expensive and there are difficulties in industrial application. A combination of radiation with other methods, such as acid treatment, can further accelerate enzymatic hydrolysis, but not without additional cost. However such processes are very helpful for laboratory analysis. | |
| Wet oxidation | During wet oxidation biomass not only undergoes chemical reactions but also undergoes physical rupture. However, the amount of by-products formed is almost always higher with pre-treatment by oxidation than by steam explosion. However the process can be combined with other pre-treatment methods to further increase the yield of sugars. |
| Liquid hot water | The processing does not require any addition of chemicals. LHW pre-treatment reduces the need for neutralization and conditioning chemicals since the processing does not require any addition of chemicals. Size reduction of the incoming biomass is also not needed since the lignocellulose particles break apart when heated in water. Other advantages of LHW pretreatment includes limited corrosion problems, no sludge generation, low capital and operational costs and negligible loss of cellulose under normal operating conditions. The main component of the operating cost is the energy required to feed the water as saturated liquid. |
| Steam explosion | The advantages of steam explosion pretreatment include the low energy requirement and that there are no recycling or environmental costs. Limitations of steam explosion include destruction of a portion of the xylan fraction, incomplete disruption of the lignin–carbohydrate matrix, and the generation of compounds that may be inhibitory to microorganisms used in downstream processes. |
| Supercritical fluid (SCF) | It was found that a combination of supercritical CO2 and steam gave the best overall yield of sugars. The high energy cost to maintain high pressures makes the process uneconomically viable. |
| Ionic liquids | ILs are expensive to synthesize. Further work is required to synthesize ILs on a commercial scale at lower cost. Research is also required to integrate IL pre-treatment with subsequent hydrolysis and fermentation. |
| Organosolvent | The principal costs are the energy consumption and the energy required to recover the solvent. Another challenge is the volatility and flammability of organic solvents, which necessities rigorous procedures for their safe confinement. The carry-through of solvents to fermentation may also result in the inhibition of biofuel production from biomass hydrolysis. |
| Ammonia fiber explosion (AFEX) | High energy costs and liquid loadings, along with the many disadvantages associated with the AFEX process, are still some of the major concerns that need to be addressed before this process is proven to be economically viable. An efficient ammonia recovery process is an important factor to make this pre-treatment method economically feasible. |
| Alkaline extraction | Neutralization after the alkaline pre-treatment of lignocellulosic biomass is problematic because of the gradual release of alkali from the solid portion during washing without the addition of some acid. Neutralizing the lime with CO2 eliminates the solid–liquid separation step. However such a process makes it difficult to separate CaCO3 from other solid particles after hydrolysis and fermentation. The lime recovery cost, or the lime cost, will be another important economic factor for this process. |
Enzymatic hydrolysis
Enzymes are naturally occurring proteins that cause certain chemical reactions to occur. A variety of microorganisms including bacteria and fungi contain enzymes with the ability to degrade cellulosic biomass to glucose monomers.26 A mixture of hydrolytic enzymes collectively known as cellulases carry out the natural biodegradation of cellulosic biomass. These cellulases include both endo-acting (endoglucanases) and exo-acting (cellobiohydrolases) enzymes.27 Since during enzymatic hydrolysis the utilization of both the cellulose and the hemicellulose part is desirable, the complete hydrolysis of cellulose and hemicellulose requires a well-designed cocktail of enzymes consisting of endoglucanases, cellobiohydrolases, β-glucosidases, xylanases, mannanases and various enzymes acting on the side chains of xylans and mannans.28,29 Nevertheless there is no organism with the ability to directly hydrolyse polysaccharides and ferment the liberated sugar to ethanol at the rate and titer required for economically viable operation.30,31 Organisms with a broad substrate range and cellulolytic and/or hemicellulolytic abilities generally suffer from poor growth characteristics or poor product-producing characteristics. These include poor yield, titer and rate or producing mixtures of desirable products along with undesirable ones. In comparison, organisms with only desirable product-producing qualities often suffer from a limited substrate range, including lack of cellulolytic ability, poor fermentation qualities and sensitivity to the inhibitors present in pre-treated lignocellulosic biomass. Strain development is therefore the most important technical obstacle towards the conversion of lignocellulose to commodity products in a consolidation configuration. Cellulases are relatively costly enzymes and to increase sugar production at reduced enzyme loadings is reliant on reduced production costs and an increase in the specific activities for enzymatic mixtures.32Where individual, purified enzymes are used, the individual properties of enzymes are an important factor in the selection of enzymes.33 However, it is also possible to improve the composition of crude mixtures of enzymes by modifying the growth substrate, considering that different combinations of enzymes will be induced by the specific characteristics of the substrate.34 Howard and co-workers have listed data for the specific activities of cellulases from bacteria and fungi that could be used for the selection of a cocktail of enzymes.35 Various parameters remain under consideration for the selecting enzyme mixture. The optimum pH and temperature should be compatible with other enzymes and designed reactor conditions. Enzymes should possess a high stability at the reaction conditions to optimize the enzyme usage and lower costs. Where enzymes are not operating under optimal conditions, higher enzyme loadings may be required in order to achieve the same level of hydrolysis efficiency, which will affect the overall production cost of the process. Enzymes with high specific activity result in lower enzyme dosages which reduces the cost.
The cross-specificity of enzymes (or multi-functionality) also comes under consideration as this may result in enzymes competing for the same sites if they have an overlap of functions. An enzyme with dual functionality may also alleviate the necessity for an additional enzyme and thus reduce costs.29 Other factors that affect optimization of the enzymatic hydrolysis process include enzyme ratios, substrate loadings, enzyme loadings, inhibitors, adsorption and surfactant loading.29
At present enzymes are produced in a dedicated process that involves substantial capital and material expense and represents the second highest contribution to raw material cost after the feedstock itself. The cellulase enzyme cost contribution to the total production cost of lignocellulosic ethanol can be approximated by the following equation:36
| E = (P × L × C/A × Y) × 1000 | (1) |
Typical enzyme loadings for >75% conversion of biomass to sugars are 10–15 Filter Paper Units (FPU) per g glucan (20–30 mg per g glucan) depending on the method of pretreatment and the source of enzyme.37 For high solid loading enzymatic hydrolysis, typical enzyme loadings used are around 30 to 36 mg protein per g glucan. The enzyme cost accounts for 15.7% of the total cost, even if the enzyme loading used is 20 mg per g glucan.38,39
Enzymes are intrinsically expensive and thermodynamically unstable because they are produced by living systems. Due to the chemical and physical recalcitrance of lignocellulose, high enzyme loadings are necessary to obtain reasonable degradation rates.40 In addition, since hemicellulose has a varied composition compared to cellulose, they require a large number of enzymes to hydrolyse different sugar monomers effectively. The on-site enzyme production on part of the lignocellulose substrate already available in the ethanol plant is advantageous.41 The recycling of cellulases is another way of reducing the high cost of enzymes during the bioconversion processes.5 However, it has been difficult to evaluate the reuse or recycling of cellulases primarily because our current knowledge of the characteristics of cellulase adsorption onto lignocellulosic substrates is insufficient.42–44
Enzymes that can degrade both cellulose and hemicellulose are divided into (i) depolymerising enzymes that cleave the backbone and (ii) enzymes that remove substituents that may pose steric hindrances to the depolymerising enzymes.
Table 2 (ref. 29) gives a brief overview of the types of enzymes that are required to degrade complex lignocellulose substrates. However, there are indications that many other proteins may contribute to lignocellulose degradation in ways that are not yet clearly understood.29
| Lignin | Laccase, manganese peroxidase, lignin peroxidase |
| Pectin | Pectin methyl esterase, pectate lyase, polygalacturonase, rhamnogalacturonan lyase |
| Hemicellulose | Endo-xylanase, acetyl xylan esterase, β-xylosidase, endo-mannanase, β-mannosidase, α-L-arabinofuranosidase, α-glucuronidase, ferulic acid esterase, α-galactosidase, p-coumaric acid esterase |
| Cellulose | Cellobiohydrolase, endoglucanase, β-glucosidase |
Microorganisms produce two types of enzyme systems for lignocellulose degradation, namely free and complex systems. Free enzyme systems occur in many aerobic bacteria and fungi, the most studied being Trichoderma reesei and Aspergillus niger.45 When selecting enzymes for the degradation of lignocellulose substrates, the initial choice is made between a customized cocktail of individual enzymes and a commercial crude mixture of enzymes. Either option can be useful. The use of individual enzymes can lead to a greater understanding of the synergy and cooperation between enzymes to degrade a complex substrate, whereas the use of commercial enzymes is a quicker route to commercialization. With respect to the use of individual enzymes, a shortcoming is the lack of commercial availability of pure enzymes for studying interactions between enzymes. This is particularly true for the case of lesser known accessory enzymes whose contributions have not been evaluated.29
The literature estimates that the cost contribution of the enzyme to the production of lignocellulosic ethanol varies significantly, including $0.10 per gal,46 $0.30 per gal,47 $0.32 per gal,38 $0.35 per gal,48 and $0.40 per gal.49 According to Klein-Marcuschamer and coworkers50 an estimation of the cost contribution of enzymes to produce ethanol by the conversion of corn-stover is $0.68 per gal when sugars in the biomass are converted at maximum theoretical yields. The same study suggests the cost contribution of enzymes was $1.47 per gal when the yield was based on saccharification and the fermentation yields reported in the scientific literature. They further suggested that when the enzyme loading was lowered, for instance to 5 FPU per g cellulose, the cost contribution of the enzymes would be in the range of $0.34–0.73 per gal.
Companies such as Novozymes and Genencor, through subsidization by the US government, are working to improve the cost and effectiveness of enzymes for biofuel production by producing crude, commercial mixtures.40 Their two main strategies are (1) an economical improvement in the production of cellulase to reduce the unit cost of the enzyme by process and strain enhancement, e.g., a cheaper medium from lactose to glucose and an alternative inducer system and (2) an improvement in the cellulase enzyme performance to reduce the grams of enzyme for achieving equivalent hydrolysis by cocktails and component improvement.51 Their claim has not been widely accepted because the cellulase mixture they tested was only for a specific pretreated lignocellulosic substrate. The biggest drawback of using commercial enzyme mixtures is the lack of characterization of the enzymes in the mixtures. Some of these mixtures may contain up to 80 proteins and the exact composition is still unknown. In addition to this, US funding to enzyme companies was tasked to improve the enzyme cost with respect to the degradation of acid-pretreated corn stover, therefore these commercial mixtures are therefore not optimized for other types of biomass or other types of pretreatment and further enzymes may have to be added to achieve optimized combinations.40,52 As pretreatments are moving away from acid pretreatments in the use of these commercial enzymes without necessary adjustment they may not give satisfactory performances. Qing and coworkers53 indicated that an important shortcoming of commercial mixtures is the shortage of xylanase activity. The strategy to make the cellulase-based processing more economical include (i) increasing commercial enzyme volumetric productivity, (ii) producing enzymes using cheaper substrates, (iii) producing enzyme preparations with greater stability for specific processes, and (iv) producing cellulases with higher specific activity on solid substrates. Significant effort is still required to lower the contribution of the cost of enzymes in the total biofuel production cost.
Fermentation
In fermentation, microorganisms use the hydrolytic products produced during the hydrolysis of lignocellulosic material as a substrate to produce ethanol. The yield of fermentation is expressed on either a molar or weight basis. The primary stoichiometric equations for the bioethanol production are as follows.Hydrolysis of pentosan to pentose:
| nC5H8O4 + nH2O → nC5H10O5 | (2) |
| n(132) g + n(18) g → n(150) g | (3) |
| (1) g + (0.136) g → (1.136) g | (4) |
Fermentation of pentose to bioethanol:
| 3nC5H10O5 → 5nC2H5OH + 5nCO2 | (5) |
| 3n(150) g → 5n(46) g + 5n(44) g | (6) |
| n(1.136) g → n(0.58) g + n(0.56) g | (7) |
| Yield(ethanol/pentosan) = 0.58 g of ethanol per g of pentosan | (8) |
Hydrolysis of hexosan to hexose:
| nC6H10O5 + nH2O → nC6H12O6 | (9) |
| n(160) g + n(18) g → n(178) g | (10) |
| (1) g + (0.111) g → (1.111) g | (11) |
Fermentation of hexose to bioethanol:
| nC6H12O6 → 2nC2H5OH + 2nCO2 | (12) |
| n(180) g → 2n(46) g + 2n(44) g | (13) |
| n(1.11) g → n(0.567) g + n(0.54) g | (14) |
| Yield(ethanol/hexosan) = 0.58 g of ethanol per g of hexosan | (15) |
The conversion of other oligosaccharides requires their hydrolysis to either hexose or pentose. This also results in the same stoichiometric gains. According to the reactions, the theoretical maximum yield is 0.51 kg ethanol and 0.49 kg carbon dioxide per kg of xylose and glucose. Reduction in the yield below the theoretical value always occurs since the microorganism requires a portion of the substrate for cell growth and maintenance. For E. coli and S. cerevisiae these values are approximately 0.054 and 0.018 grams of glucose per g dry cell weight per h, respectively.54
Fermenting organism
For the commercial production of ethanol from lignocellulosic material a key goal is to convert all C5 (pentose) and C6 (hexose) sugars into ethanol.55 In this regard the ability of the fermenting microorganisms to utilize both C5 and C6 sugars available from the hydrolysate is a vital factor to improve the economically competitive production of cellulosic ethanol.56–58Currently, there are no known natural organisms that have the ability to convert both C5 and C6 sugars to produce high ethanol yields.59 The lack of industrially suitable microorganisms for converting the C5 of biomass into fuel ethanol has been a major technical roadblock in the production of ethanol from lignocellulosic material.
The ideal microorganism for bioethanol production will produce a single fermentation product i.e. ethanol and possess high substrate utilization and processing capacities, fast and deregulated pathways for sugar transport, good tolerance to inhibitors and products, and high metabolic fluxes.60 Temperature tolerance and tolerance towards low pH-values facilitates adaptation to changing operating conditions for different feedstocks.
Fermentation at a high substrate concentration is required to increase the ethanol yield, however an increase in the ethanol level also acts as an inhibitor of microorganism growth and viability. Since high productivity and high ethanol concentrations keep distillation costs low, the tolerance of the fermenting organism to fermentation inhibitors therefore necessitates the use of robust, industrial strains.61
Synthetic biology and metabolic engineering have been used with numerous microorganisms to enhance their ethanol selectively in fermentation. However, the research is still in the early stages of engineering organisms for biofuel production and many different organisms are being explored concurrently for their potential to be the most relevant strains. Most research focuses on S. cerevisiae, Z. mobilis and E. coli.66,89 Many microorganisms, including bacteria and yeasts produce ethanol as the main fermentation product from carbohydrates. However, yeasts, particularly S. cerevisiae are usually the first choice for industrial ethanol production, because of their good fermentative capacity, high tolerance to ethanol and other inhibitors (either formed during the raw-material pre-treatments or produced during fermentation) and the capacity to grow rapidly under the anaerobic conditions that are characteristically established in large-scale fermentation vessels. S. cerevisiae cannot utilize C5 sugars. Many other microorganisms possess the ability to utilize C5 sugar efficiently, but cannot produce ethanol at a sufficient yield and productivity. The details of the common organisms for ethanol production are given in Table 3 (ref. 59 and 62–88) while the characteristics required for an industrially suitable microorganism are summarized in Table 4.68,89,90
| Organism strain group | Minutiae |
|---|---|
| S. cerevisiae | Most commonly used yeast in industrial fermentation |
| Metabolizes glucose by the Embden–Meyerhof (EM) pathway | |
| Produces over 50 mmol of ethanol per h per g of cell protein | |
| High yield (higher than 0.45 g per g at optimal conditions) and a high specific rate (up to 1.3 g per g cell mass per h (ref. 62)) | |
| S. cerevisiae fermentation can reach final ethanol concentrations of 160 g per l | |
| High ethanol tolerance (over 100 g per l has been reported for some strains and media)63 | |
| Produces ethanol from glucose at a high rate (3.3 g l−1 h−1)64 | |
| Can grow at a low pH64 | |
| It is important to control the pH value in the range of 4.0–5.0. Beyond this range, the formation of by-products, such as acetic acid and butyric acid may consume some of the substrate and reduce the efficiency of ethanol fermentation.65 | |
| The maximum specific ethanol production rates were observed between 30 and 45 °C with different initial glucose concentrations | |
| Generally regarded as safe64 | |
| Many strains are known to be robust in industrial applications64 | |
| Wild-type S. cerevisiae efficiently converts both glucose and mannose into ethanol59 | |
| No known natural organisms have the ability to convert both C5 and C6 sugars to produce high ethanol yields59 | |
| Metabolically engineered strains allow the anaerobic growth on xylose in synthetic media with a maximum specific growth rate μmax of 0.09 h−1(ref. 66) | |
| Genetically engineered recombinant strains can produce enzymes to efficiently co-ferment both C5 and C6 sugars.67,68 Recombinant S. cerevisiae (TMB 3400) successfully ferments both xylose and arabinose in addition to glucose69,70 at an efficiency close to its theoretical value of 0.51 g per g (ref. 71). | |
| Cannot tolerate high temperatures64 | |
| Low molecular weight organic acids, furan derivatives, phenolics and inorganic compounds inhibit fermentation unless the feed to fermentation is detoxified | |
| P. stipitis | Natural xylose-fermenting yeast |
| Fed batch cultures produce almost 50 g per l of ethanol from xylose with yields of 0.35 to 0.44 g per g xylose, and can ferment hydrolysates at 80% of the maximum theoretical yield72–74 | |
| Breaks down wood into monomeric sugars, ferments glucose, xylose, mannose, and galactose75 and also has the ability to produce cell mass from L-arabinose, but not ethanol76 | |
| Relatively low ethanol yield and inhibitor tolerance,77,78 and in addition, a very low and well-controlled supply of oxygen is required for efficient xylose fermentation79 | |
| Maximum ethanol productivity is only around 0.9 g l−1 h−1 (ref. 80) | |
| Detoxifies inhibitors present in pretreated lignocellulosic materials by consuming acetic acid and reduces the furan ring in furfural and hydroxymethylfurfural (HMF)76 | |
| The genome sequence for P. stipitis has been published.75 The sequence shows numerous genes encoding xylanase, endo-1,4-β-glucanase, exo-1,3-β-glucosidase, β-mannosidase, and α-glucosidase.75 The presence of these genes in P. stipitis suggests the presence of useful traits for the simultaneous saccharification and fermentation of cellulose and hemicellulose.81 Some strains like yeast strain Y7 because of their ability to produce ethanol from hydrolysates at high efficiency without detoxification offer potential economic benefits by reducing the need for equipment to detoxify the hydrolysate82 | |
| Jeffries and co-workers reported the complete genome sequence of the lignocellulose-bioconverting and xylose-fermenting yeast Pichia stipitis75 | |
| E. coli | Bacteria that metabolizes all major sugar monomers present in plant biomass83 |
| Characterized as a good consumer of xylose84 | |
| Recombinant E. coli, e.g. the KO11 strain has shown ethanol yields from 86 to close to 100% of the theoretical, and final ethanol concentrations up to 40 g per l of hemicellulose hydrolyzates of bagasse, corn stover and corn hulls85 | |
| Narrow and neutral pH growth range (pH 6.0–8.0) | |
| The end products vary greatly among different microorganisms64 | |
| Z. mobilis | Gram-negative facultative anaerobic bacteria |
| Uses the Entner–Doudoroff (ED) pathway to metabolize glucose, which results in only 1 mole of ATP being produced per mole of glucose86 | |
| High ethanol tolerance of up to 16% (vol/vol)86 | |
| Can produce ethanol from glucose at final concentrations of 130 g per l | |
| Tolerates up to 120 g per l ethanol68 | |
| Ferments sugars at a low pH | |
| Produces less biomass than yeast | |
| Good resistance to the inhibitors found in lignocellulosic hydrolysates | |
| Does not ferment or utilize xylose as a carbon source and cannot hydrolyze polysaccharides64 | |
| Relatively easy to genetically manipulate | |
| Seo and co-workers reported the complete genome sequence of Zymomonas mobilis ZM4 (ATCC31821)86 | |
| Zhang and co-workers87 patented a Z. mobilis strain capable of fermenting both xylose and arabinose, the major pentose sugars present in plant material | |
Co-fermentation of 100 g per l sugar (glucose![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :xylose:arabinose—40:40:20) yielded a final ethanol concentration of 42 g per l in 48 h with the preferential order of utilization of glucose first, then xylose and lastly arabinose88 :xylose:arabinose—40:40:20) yielded a final ethanol concentration of 42 g per l in 48 h with the preferential order of utilization of glucose first, then xylose and lastly arabinose88 |
| Trait | Requirement |
|---|---|
| Ethanol yield | >90% of theoretical |
| Ethanol tolerance | >40 g per l |
| Ethanol productivity | >1 g l−1 h−1 |
| Robust grower and simple growth requirements | Inexpensive medium formulation |
| Able to grow in hydrolysates | Resistance to inhibitors |
| Retard contaminants | |
| Culture growth conditions | Acidic pH or higher temperatures |
Lau and co-workers91 suggested that S. cerevisiae 424A (LNH-ST) is the most relevant strain for commercial production due to its ability to ferment both C5 and C6 from un-detoxified and un-supplemented hydrolysate from AFEX-pretreated corn stover at a high yield. S. cerevisiae 424A (LNH-ST) is highly robust and able to ferment both C5 and C6 to ethanol reasonably well (greater than 85% of ethanol yield) even at a high solids loading. A qualitative summary of the relative fermentation performances of Z. mobilis AX101, S. cerevisiae 424A (LNH-ST) and E. coli (KO11) is given in Table 5.92
| Parameters | AX101 | 424A | KO11 |
|---|---|---|---|
| Glucose consumption (in corn steep liquor (CSL) | Very fast | Very fast | Fast |
| In lignocellulosic hydrolysate | Average | Very fast | Average |
| Xylose consumption (in CSL co-fermentation) | Average | Very slow | Fast |
| Xylose consumption (in lignocellulosic hydrolysate) | Very slow | Average | Very slow |
| Nutrient requirement (glucose-only) | Low | Low | Low |
| Nutrient requirement (co-fermentation) | Low | Low | Low |
| Nutrient requirement (xylose-only) | Average | High | Low |
| Growth robustness | Average | Very high | High |
| Metabolic yield | Very high | High | High |
In the future, genomics and pathway engineering are expected to facilitate the development of organisms with the ability to use the full repertoire of cellulosic and hemicellulosic sugars and tolerate high ethanol concentrations to optimize ethanol yields.93,94 E. coli does not natively produce or tolerate high levels of ethanol. However, it is historically the most amenable organism to genetic modifications and it has successfully demonstrated the use of exogenous genes and hybrid pathways to produce foreign compounds. Research in this field to produce genetically engineered microbes is an important way to enhance the lignocellulose conversion yield.
Fermentation process
Fermentation can be carried out as a batch, fed batch or continuous process. The selection of a suitable process depends upon the properties of hydrolysates, kinetic parameters and the resulting process economics. Fed-batch reactors are widely used in industrial applications because they combine the advantages from both batch and continuous processes. The major advantage of fed-batch, compared to batch, is the ability to increase the maximum viable cell concentration, prolong culture lifetime, and allow product accumulation to a higher concentration. An optimal feed-rate time profile is usually close to exponential, however, the simplified time profiles, such as a constant rate or ramp shape profiles can give process optimization results close to optimal.95 This process allows for the maintenance of critical process variables (e.g., temperature, pH, and dissolved oxygen) at specific levels through feedback control.96,97 The control of the flow rate of medium feed is quite advantageous because through such control the inhibitory effect caused by high substrate or product concentrations in the fermentation broth is neutralized. It was observed that the addition of sucrose in a linear or exponentially decreasing way leads to a 10–14% increase in ethanol productivity.98 The optimization of feeding policy plays a crucial role for increasing both the productivity and ethanol yield of fed-batch fermentations. For fed-batch cultures, Alfenore and coworkers99 showed that higher ethanol concentrations (147 g per l) can be obtained in cultures without oxygen limitation (0.2 volume of air per unit volume of medium per minute-vvm) during only 45 h in comparison to microaerobic conditions.In the cases of multiple or repeated batch fermentation, the use of flocculating yeast strains plays an important role. In this process, after starting a conventional batch, the yeasts decant in the same vessel where they are cultivated by removing the clarified culture broth. For the following batch, an equal amount of fresh culture medium is added. In this way, high cell concentrations are attained and the inhibition effect of ethanol is reduced without adding flocculation aids or using separation or recirculation devices. These repeated batches are produced until the activity and viability of culture is lost due to a high exposition to the fermentation environment. When this occurs, the system may be re-inoculated. Some examples of typical batch, fed-batch and repeated batch fermentations for bioethanol production from sugar cane molasses can be observed in Table 6.100
| Regime | Configuration | Ethanol conc. in broth (g per l) | Productivity (g l−1 h−1) | Yield (% of theor. max.) |
|---|---|---|---|---|
| Batch | Reuse of yeast from previous batches; yeast separation by centrifugation | 80–100 | 1–3 | 85–90 |
| Fed-batch | Stirred tank with variable feeding rate (exponent. dependent on time) | 53.7–98.1 | 9–31 | 73.2–89 |
| Repeated batch | Stirred tank; flocculating yeast; up to 47 stable batches | 89.3–92 | 2.7–5.25 | 79.5–81.7 |
| Continuous | CSTR (continuous stirred-tank reactor); cell recycling using a settler; flocculating yeast; aeration 0.05 vvm | 70–80 | 7–8 | |
| Biostill; residence time 3–6 h; cell recycling by centrifugation; recycled stream from distillation column to fermentor | 30–70 | 5–20 | 94.5 | |
| Continuous removal of ethanol | Removal by vacuum; cell recycling | 50 | 23–26.7 |
The design and development of continuous fermentation systems has allowed the implementation of more cost effective processes. Continuous processes have several advantages compared to conventional batch processes mainly due to the reduced construction costs of the bioreactors, lower maintenance and operation requirements, better process control, and higher productivities (see Table 6). Most of these advantages are due to the high cell concentration found in these processes. Such high densities can be reached by immobilization techniques, recovery and recycling of cell biomass, or control of microbial growth. The major drawback is that yeasts cultivated under anaerobic conditions during a long time diminishes their ability to synthesize ethanol. In addition, at a high dilution rate, the substrate is not completely consumed, which reduces the yield. Aeration also plays an important role during continuous cultivation. Cell concentration, cell yield from glucose, and yeast viability enhances with increasing air supply, whereas ethanol concentration decreases under both microaerobic and aerobic conditions. Cell growth inhibition by ethanol is reduced at microaerobic conditions compared to fully anaerobic cultivation and specific ethanol productivity is stimulated with the increase of oxygen percentage in the feed.99 Continuous processes permit the decrease of the product inhibition effect. Through a cascade of continuous reactors, ethanol obtained in the first reactors is easily transported to the following reactors reducing its inhibitory effect.
One of the strategies used for improving the ethanolic fermentation is the utilization of immobilized cells that allow the implementation of continuous processes with higher yields and productivities (see Table 6), and with increased cell concentration.101
The presence of inhibitors directly influences the course of ethanolic fermentation. In continuous or fed-batch fermentations, a low concentration of inhibitors in the broth can be achieved by avoiding high flow rates of feed to bioreactors. In continuous systems, inhibitors diminish the growth rate and the process productivity directly links to the dilution rate. In systems with cell retention (e.g., by cell recirculation using filtration, sedimentation or centrifugation), the increase of accumulables, including the inhibitors, makes the productivity decrease, imposing the need to implement purge streams.
Product separation
Distillation separates ethanol from the fermentation broth followed by dehydration through a molecular sieve to improve the concentration of ethanol. The effects of ethanol concentration in the fermentation broth on distillation energy spending are shown in Fig. 1. | ||
| Fig. 1 Relationship between the ethanol distillation energy and ethanol concentration.10 | ||
Process integration
Fig. 2 (ref. 102) shows that large numbers of individual processes are involved in the conversion of ligno-cellulosic biomass into ethanol which suggests there is considerable potential for process integration to lower capital and operating costs, as well as ensuring the optimum production of valuable co-products. In order to minimize the initial investment and operational cost some of the individual processes can be integrated. However, as second-generation process development is still at the pre-commercial stage, it may take some time to determine the most efficient process pathway and system. | ||
| Fig. 2 Schematic representation of lignocellulose to ethanol process configurations.102 | ||
Separate hydrolysis and fermentation (SHF)
When enzymatic hydrolysis and fermentation are performed sequentially, it is referred to as SHF. In the SHF configuration, the liquid flow from the hydrolysis process first enters the glucose fermentation reactor. In next step, the mixture is distilled to remove the ethanol leaving the unconverted xylose behind. In a second reactor, xylose is fermented to ethanol, and the ethanol is again distilled.103 In SHF each step is performed at its optimal operating conditions, as shown in Fig. 1. However, since cellulolytic enzymes are end product inhibited the rate of hydrolysis progressively slows down with the accumulation of glucose and cellobiose.30 It takes around 72–168 h to achieve high sugar conversions at high solid loading and another 96–168 h to ferment xylose sugars. Thus the overall biological processing times are well over 5 days, leading to higher capital and operating costs.5In SHF the most important factors for the hydrolysis step are reaction time, temperature, pH, enzyme dosage and substrate load. Although an enzyme dose of 100 FPU per g cellulose gives almost 100% hydrolysis, however this amount of cellulase is not economically viable. Commercial operations are proposed at 40 FPU per g cellulose dose, which results in an only 13% reduction in conversion. Regarding the substrate concentration, solid loads at 10% are considered most adequate keeping in view the mixing difficulties and accumulation of inhibitors in the reaction medium. Saha and co-workers obtained a 96.7% yield of monomeric sugars using an enzymatic cocktail of cellulase, β-glucosidase and xylanase for the hydrolysis of wheat straw pre-treated by an alkaline peroxide method.101
An ethanol concentration of 18.9 g per l and a yield of 0.46 g per g of available sugars were achieved in the subsequent fermentation using a recombinant E. coli strain capable of assimilating both hexoses and pentoses. Iogen Corporation, a major manufacturer of industrial enzymes in Canada has developed an SHF process comprising a dilute-acid-catalyzed steam explosion and the removal of the major part of the acetic acid released during the pre-treatment, the use of S. cerevisiae as a fermenting organism, distillation of broth, bioethanol dehydration and the disposal of stillage in landfill.14
Simultaneous saccharification and fermentation (SSF)
In SSF enzymatic hydrolysis and fermentation are carried out simultaneously in the same vessel. Performing the enzymatic hydrolysis together with the fermentation reduces the end-product inhibition of the enzymatic hydrolysis, and the reduced investment costs. Combining hydrolysis and fermentation decreases the number of process steps needed and thereby capital investment by approximately 20%.104 The principal drawback of SSF is the need to find favourable conditions (e.g. temperature and pH) for both the enzymatic hydrolysis and the fermentation and the difficulty to recycle the fermenting organism and the enzymes. Due to the difficulties in separating the yeast from the lignin after fermentation, it is not possible to reuse the yeast in SSF process. The difficulty to recycle the yeast makes it beneficial to operate at low yeast concentration and high solid loading.104 Still if the yeast is produced from carbohydrates within the process, this results in yield loss or otherwise it increases the operational cost when produced separately. Enzymes are equally difficult to recirculate as the enzymes bind to the substrate. When enzymes are produced within the process this results in a loss of substrate or adds to the chemical costs when they are supplied externally. A partial desorption can be obtained by the addition of surfactants.105Simultaneous saccharification and cofermentation (SSCF)
Simultaneous saccharification and cofermentation (SSCF) involves two process steps: cellulase production followed by cellulose hydrolysis and the fermentation of both cellulose and hemicellulose. SSCF reduces the time for enzymatic hydrolysis and improves productivity. SSCF allows a slow, constant release of glucose throughout the process that is beneficial for xylose uptake by xylose-fermenting strains of S. cerevisiae.106–108 In SSCF higher ethanol yields may be achieved by the alleviation of product inhibition in cellulases, which results in more complete hydrolysis of the cellulose component. However, optimum temperatures for the enzymatic hydrolysis and ethanol fermentation are significantly different which makes the simultaneous optimization of the two unit operations difficult. A lower temperature (30–35 °C) in the SSCF process accommodates both microbial growth and ethanol fermentation. However the rate of the enzymatic hydrolysis is compromised, and a much longer time is required to complete the hydrolysis. Since lignins are not separated from cellulose, the fermentation broth is extremely viscous affecting the mixing, heat and mass transfer performance. The energy consumption is high for the distillation of the fermentation broth with low ethanol concentrations, as well as for the treatment of distillage since the amount of the discharge is much larger. Substrate feeding prevents the viscosity from becoming too high and allows a higher total amount of water-insoluble solids (WIS) in the process. The enzyme feeding helps in keeping the glucose concentration low, which kinetically favours xylose uptake and results in a higher xylose conversion. Olofsson106,108 and co-workers reported a time as long as 96 h for the fed-batch SSCF system to convert pretreated wheat straw with 11% WIS to produce only 3.3% (w/v) ethanol. Careful combination of different feeding strategies have shown that it is possible to increase the final WIS content to 11% in the SSCF and still obtain an ethanol yield of 0.35 g per g of fermentable sugars.108 Sequential fermentation with two different microorganisms in different time periods of the fermentation process for better utilization of sugar have also been employed using S. cerevisiae in the first phase for hexose utilization and C. shehatae in the second phase for pentose utilization, but the ethanol yields achieved are not high.100 Similar ethanol yields were obtained by other researchers, approximately 0.21 g per g (63 gal per ton) (see Table 7 (ref. 92 and 109–115)) despite differences in process complexity. Higher yields were obtained when purified cellulose was used or starch combined with lignocellulose. In summary low ethanol productivity and high enzyme loading are the major concerns of SSCF.| Feedstock | Pre-treatment | Biocatalyst | Ethanol | |
|---|---|---|---|---|
| Titer (g per l) | Yield (g per g of untreated feedstock) | |||
| a Results presented were used to calculate yields on an original biomass basis. | ||||
| Rice straw | AFEX | S. cerevisiae 424A (LNH-ST) | 37 | 0.21 |
| Rice straw | AFEX | P. stipitis FPL-061 | 30 | 0.17 |
| Rice straw | AFEX | P. stipitis DX-26 | 28 | 0.16 |
| Switchgrass | Hydrothermolysis | S. cerevisiae D5A | 22 | 0.17 |
| Switchgrass | Hydrothermolysis | K. marxianus IMB | 19 | 0.15 |
| Barley straw | Steam explosion | K. marxianus CECT 10875 | 22 | 0.17 |
| Distillers grains | Liquid hot water | S. cerevisiae D5A | 14 | 0.09 |
| Distillers grains | AFEX | S. cerevisiae D5A | 14 | 0.09 |
| Corn stover | AFEX | S. cerevisiae 424A (LNH-ST) | 40 | 0.22 |
| Corn stover | AFEX | E. coli KO11 | 31 | 0.17 |
| Corn stover | AFEX | Z. mobilis AX101 | 32 | 0.18 |
| Forage sorghum | AFEX | S. cerevisiae 424A (LNH-ST) | 31 | 0.17 |
| Sweet sorghum bagasse | AFEX | S. cerevisiae 424A (LNH-ST) | 42 | 0.15 |
| Forage sorghum | AFEX | S. cerevisiae 424A (LNH-ST) | 31 | 0.18 |
| Sweet sorghum bagasse | AFEX | S. cerevisiae 424A (LNH-ST) | 29 | 0.18 |
| Corn silage | AFEX | S. cerevisiae 424A (LNH-ST) | 28 | 0.31 |
| Whole corn plant | AFEX | S. cerevisiae 424A (LNH-ST) | 30 | 0.32 |
| Paper sludge | No additional treatments | S. cerevisiae RWB222 | 45 | 0.26 |
| Paper sludge | No additional treatments | Z. mobilis 8b | 46 | 0.27 |
Jin and co-workers have proposed a process to hydrolyze the easily digestible biomass first and avoided the slow-down period by performing enzymatic hydrolysis only for 24 h. The unhydrolysed solids with some enzymes adsorbed were recycled to the next cycle for further hydrolysis. By this approach, a part of the enzymes was recycled which reduces the required enzyme loading. This was followed by fermentation under high inoculum level (OD ¼ 20 instead of 0.5–2) to complete fermentation in 24 h. High inoculum level fermentation bypassed the degradation product inhibition on yeast cell growth and hence improved xylose fermentation. The yeast cells were subsequently recycled. It took 24 h for enzymatic hydrolysis and another 24 h for fermentation to complete one cycle.5
Consolidated bioprocessing (CBP)
In CBP, all bioconversion steps are minimized to one step in a single reactor using one or more microorganisms.27,31 Such consolidation of enzyme production, saccharification, and fermentation is increasingly recognized as having potential for the low-cost production of bioethanol as the high costs of capital investment, raw materials, and equipment associated with microbial enzyme production can be avoided.116 A schematic comparison of SSF, SHF, and CBP is given in Fig. 3.117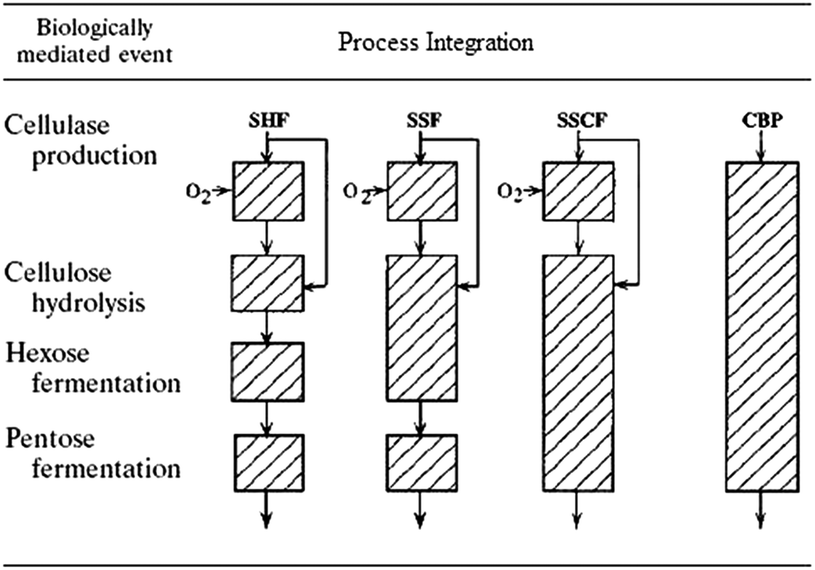 | ||
| Fig. 3 Summary of process integration. Each box represents a bioreactor (not to scale).117 | ||
CBP requires a highly engineered microorganism developed for several different process-specific characteristics.118 Other prerequisites include engineering microbial strains with high cellulolytic activity, high product yield, productivities, and titers, the ability to use many carbon sources, and the resistance to toxic compounds released during the pretreatment of lignocellulosic biomass.119 The ideal CBP organism should be robust with regards to inhibitor tolerance, able to degrade lignocellulose and utilize hexose and pentose sugars at a high efficiency.120
Microorganisms with such a combination of substrate-utilization and product formation properties are not currently available. However, given that the research community is putting in efforts, a breakthrough may be achieved in the near future. Progress in developing CBP-enabling microorganisms is being made through two strategies: engineering naturally occurring cellulolytic microorganisms to improve product-related properties, such as yield and titer, and engineering non-cellulolytic organisms that exhibit high product yields and titers to express a heterologous cellulase system enabling cellulose utilization.31,117
Lynd and co-workers have identified several yeast strains with innate properties that make them attractive as possible CBP organisms.31 The integration of cellulolytic capabilities with metabolic systems specified for targeted chemical production will allow the development of customized CBP microorganisms using advanced gene manipulation technologies.
Barriers to the commercialization of lignocellulosic ethanol include the development of more robust enzymes, reduction of cellulase costs, and the high capital cost associated with a complex process.121 Ethanol yields resulting from most current process configurations are similar, approximately 0.21 g ethanol per g dry cellulosic feedstock. Potentially, this can be increased to at least 0.27 g ethanol per g biomass (83 gal per ton) using simpler processes. A comparison between SSF, SHF and CBP is given in Table 8.
| Fermentation process | Features and advantages | Limitations |
|---|---|---|
| SHF | Each step can be processed at its optimal operating conditions | The rate of hydrolysis progressively slows down with the accumulation of glucose and cellobiose |
| Separate steps minimizes interactions between the steps | End product inhibition minimizes the yield of ethanol | |
| Chance of contamination due to a long period process | ||
| SSF | Performing the enzymatic hydrolysis together with the fermentation reduces the end-product inhibition of the enzymatic hydrolysis, and reduces investment costs | It is difficult to achieve favourable conditions (e.g. temperature and pH) for both the enzymatic hydrolysis and the fermentation |
| The difficulty to recycle the fermenting organism and the enzymes | ||
| SSCF | Involves only two process steps | The process must be operated at lower temperatures to accommodate microbial growth and ethanol fermentation |
| Allows a slow, constant release of glucose throughout the process that is beneficial for xylose uptake by xylose-fermenting strains | The rate of the enzymatic hydrolysis is inevitably compromised, and a much longer time is needed to complete the hydrolysis | |
| Higher ethanol yields can be achieved due to the alleviation of product inhibition in cellulases, which results in more complete hydrolysis of the cellulose component | The fermentation broth is extremely viscous, affecting the mixing, heat and mass transfer performance | |
| The energy consumption is high for the distillation of the fermentation broth with low ethanol concentrations, as well as for the treatment of distillage since the amount of the discharge is much larger | ||
| CBP | Elimination of the enzyme production process | Requires a highly engineered microorganism developed for several different process-specific characteristics |
| Reduction of vessels for saccharification and fermentation | Microorganisms with such a combination of substrate-utilization and product formation properties are not currently available | |
| Reduction of raw substrate materials for reaction | ||
| Reduction of contamination risk | ||
| Reduction of capital investment |
Strain development using metabolically engineered microorganisms
In CBP a single microorganism capable of fermenting pretreated biomass without adding lignocellulolytic enzymes is required, which could potentially result in a lower cost of production of lignocellulosic ethanol.31 S. cerevisiae presently used in bioethanol production ferments glucose, but not xylose. Yeasts engineered to ferment xylose do so slowly, and cannot utilize xylose until the glucose is completely consumed. Producing biofuels in yields high enough to be useful requires engineering of the microorganism's metabolism. Such engineering is not based on just one specific feedstock or host organism. Data-driven and synthetic-biology approaches can be used to optimize both the host and pathways to maximize fuel production.122 The general method employed to develop microbial biofuels is to characterize the enzymes, regulation, and thermodynamics of relevant native metabolic pathways and then to adjust or redesign those pathways around catalytic capabilities, cofactor pools, and other driving forces towards the production of the desired compound.123Ha and co-workers124 have engineered yeasts to coferment mixtures of xylose and cellobiose. In these yeast strains, the hydrolysis of cellobiose takes place inside yeast cells through the action of an intracellular β-glucosidase following import by a high-affinity cellodextrin transporter. For this purpose they introduced a cellodextrin transporter and intracellular β-glucosidase from the cellulolytic fungi Neurospora crassa, into S. cerevisiae strains engineered to ferment xylose.124 Wen and coworkers125 reported the first successful assembly of trifunctional minicellulosomes in S. cerevisiae in vivo with the ability to simultaneously hydrolyse and ferment amorphous cellulose to ethanol. They suggested that coupled with flow cytometry, the yeast surface display provides a more convenient engineering platform, avoiding labor-intensive protein purification steps.125
Sun and co-workers126 have reported the construction of a trifunctional minihemicellulosome on the yeast cell surface. Like a minicellulosome, the engineered minihemicellulosome is also a multienzyme complex including several assembled modules. The chimeric enzymes were assembled in vivo on the miniscaffoldin to hydrolyze arabinoxylan into D-xylose and arabinose. To enable S. cerevisiae to utilize D-xylose, the known D-xylose utilization pathway consisting of xylose reductase (XR), xylitol dehydrogenase (XDH), and D-xylulokinase (XK) from Scheffersomyces stipitis was integrated into the S. cerevisiae L2612 genome. Through an engineered strain they achieved the direct conversion of birchwood xylan to ethanol through the CBP strategy.126 However they suggested further investigations to improve the xylan degradation ability of the CBP strain, since the residual total sugar concentration reached a plateau after 80 hours. For the complete degradation and saccharification of xylan, other critical xylanases might also be needed.126 Kim and coworkers designed a cellulolytic yeast consortium, which was composed of cells displaying mini cellulosomes generated via the random assembly of CelA and CBHII to a mini CipA, and cells displaying BGLI independently.127 They suggested that there is no limitation on the number of enzymes to be incorporated into this cellulosome structure. With such a consortium ethanol production can be easily optimized by simply changing the combination ratio of the different populations.
Liu and coworkers128 while reviewing the range of liquid fuels that can be produced in E. coli suggested E. coli as the most convenient starting point for engineering microbial catalysts for biofuel production. They suggested upstream problems such as converting lignocellulosic biomass into fermentable carbon, and downstream problems such as improving biocatalyst tolerance to high fuel concentrations require attention. Christine and co-workers123 suggested that due to comparatively faster growth rates, a higher potential product yield per cell, genetic tractability, and depth of knowledge of the organism, well-studied bacteria or yeast are the ideal hosts for advanced biofuel production. The Gram-negative bacterium E. coli and the yeast S. cerevisiae are examples of user-friendly hosts that have been genetically engineered to generate a range of biofuels from the simple sugar glucose, from pathways based around the central metabolism of fermentation or respiration.122,123 According to Christine and coworkers123 bacteria provide an ideal metabolic “factory” due to their incredible malleability and resiliency and their toolbox of evolutionarily honed central and peripheral metabolic pathways. Recently Zheng and coworkers have reported the high-specificity production of isoprenol and prenol by microbial fermentation using the heterogenous mevalonate-dependent (MVA) isoprenoid pathway for the synthesis of isopentenyl pyrophosphate (IPP) and dimethylallyl pyrophosphate (DMAPP) intermediates, and then converting IPP and DMAPP to isoprenol and prenol, respectively.129 Several studies have suggested that the full capacity for microbial biofuel production is yet to be realized.123,130–132 Further progress in metabolically engineered microorganisms is needed to improve the strain activity and to overcome the slow catalysis and low fermentation efficiency.
Barriers to the improvement of ethanol yields
Establishing economically feasible fermentation processes requires markedly increasing final product titers due to the high energy demands of subsequent product recovery steps, as well as the capital and production costs associated with biorefinery equipment.119 Microorganisms that are resistant to inhibitors are a prerequisite for the high-titer production of fuels and chemical products.133 Recombinant ethanologenic organisms (E. coli and Z. mobilis) have been created to ferment both glucose and xylose, but they currently produce lower ethanol titers. Improvements in ethanol yields and tolerance are required to increase rates of production from all sugar constituents of lignocellulosic biomass. As titers are increased, rates slow down and eventually cease at ∼6 wt percentage ethanol, the upper limit for wild-type E. coli. By comparison, wild-type yeast and Z. mobilis can reach titers of >15% ethanol from cornstarch glucose, but have failed to achieve these levels for pentose sugars. This also contributes to the cost of lignocellulosic ethanol by increasing capital expenditure, reducing product yields, and increasing water volumes that must be handled as part of relatively dilute product streams.Yield and productivity enhancement may be accomplished through applying concepts and methods of metabolic engineering. Eliminating branches of competing pathways that lead to unwanted products help in yield maximization. This is usually carried out by deleting genes encoding enzymes that catalyze competing reaction pathways. If such pathways are responsible for the synthesis of metabolites essential for cell growth and function, down regulation of these genes may be preferable to complete gene knockout. In all cases, an optimal balancing of enzymatic activities is critical for the satisfactory function of the resulting engineered strain. A strategy for increasing ethanol tolerance allows the microbial process to evolve under the proper selective pressure (in this case, higher ethanol concentrations) to increasingly higher ethanol tolerances.
To increase the ethanol concentration, a high WIS is required. However, high WIS content increases the viscosity of the medium, leading to severe mixing problems. For each feedstock, there is a different upper limit for WIS content that is practically possible to handle. A high WIS content gives a high concentration of inhibitors. A low viscosity of the medium can be maintained in fed-batch systems by the gradual feeding of new material to the reactor, in such a case the viscosity decreases due to enzymatic degradation.134 By controlling both enzyme and substrate feeding, it is possible to improve the xylose conversion in SSCF from 40% to 50%, in comparison to a regular fed-batch process.
Critical parameters and figures of merit needed for a cost-competitive process
The critical considerations of a commercial competitive biomass-to-ethanol process are(i) A high yield with complete sugar utilization; (ii) minimum by-product formation; (iii) minimum loss of carbon into cell mass; (iv) a higher final ethanol titer; (v) a higher overall volumetric productivity, especially under high-solid conditions; (vi) tolerance to inhibitors present in hydrolysates.
To address the above consideration this review suggests the following figures of merit.
(i) Utilizing both hexoses and pentoses to produce ethanol at a yield greater than 95% of theoretical yield; (ii) keeping the final ethanol titers in the range of 10 to 15 wt%; (iii) maintaining overall volumetric productivity at 2–5 grams of ethanol per liter per hour; (iv) selecting an organism with the ability to grow and metabolize effectively on actual hydrolysates (with only minerals as added nutrients).
Strategies to promote the commercial production of lignocellulosic-derived ethanol
The massive utilization of fuel ethanol in the world requires its production technology to be cost-effective and environmentally sustainable. When compared based on the cost of production second-generation biofuels are simply too expensive to produce relative to the fossil fuels they could replace. A recent literature review reported current production costs of second-generation ethanol to be in the range of $0.60–$1.30 per liter.46 Technology models taking into account the actual research results and projecting them to commercial scale, estimate an ethanol cost of $0.64 per liter at present.135 In order to reduce costs, process improvements have to be made in several areas, including pre-treatment, enzymatic hydrolysis, and fermentation. With the on-going research and resulting technological advancement, it is anticipated that the production costs will come down to as low as $0.30–$0.40 per litre by 2020. The cost of raw material varies considerably between different studies (US$22–US$61 per metric ton dry matter) and it accounts for 45–60% of the total production costs for second generation biofuels, depending on the conversion efficiency and applied technology. Unlike the first generation biofuels, a collection of raw materials (e.g., agricultural residues) would be relatively expensive in the case of second-generation biofuels.15,136 With regards to feedstock, major obstacles are cost, supply, harvesting, and handling. The cost of raw materials can be reduced by using the whole crop for producing the main product and co-products. Pre-treatment, is another expensive processing step, with costs as high as US$0.08 per litre of ethanol produced. The contribution of enzymes to production costs is about US$0.04 per litre of ethanol, with some variation depending upon actual ethanol yields resulting from the particular pre-treatment approach. In the hydrolysis process the challenge is to achieve an efficient process for the depolymerization of cellulose and hemicellulose to produce fermentable monomers with high concentration.137 A high ethanol yield requires complete hydrolysis of both cellulose and hemicelluloses with a minimum sugar degradation, followed by efficient fermentation of all sugars in the biomass. In the short term, co-products like the production of fuel, heat and electricity may improve the commercial viability. However, in the long term, bioethanol technology will form the basis for the sustainable production of commodity chemicals and materials in future biorefineries. A high co-product yield means a reduced energy demand for ethanol production. As the biomass industry develops, new fuels and new starting materials are under investigation. Techno economic analysis will play a key role in process development and the targeting of technical and economic barriers for these new fuels and feedstocks.48The world's first commercial cellulosic ethanol production plant from wood wastes, with an annual capacity 1.4 million litres began its operation in 2007 in Osaka (Japan). The technology, licensed by Verenium Corporation, a US-based cellulose company, uses genetically engineered strains of E. coli to co-ferment C6 and C5 sugars. Table 9 gives a list of existing, planned, and under construction facilities to produce cellulosic biofuels.
| Company | Plant location | Feedstock | Technology | Capacity (million l per year) |
|---|---|---|---|---|
| Abengoa | Colwich, Kan., USA | Corn stover, wheat straw, milo (sorghum) stubble, switchgrass, and others | Thermochemical and biochemical processing | 43.15 |
| Abengoa | York, Neb., USA | Corn stover, residual starch | Enzymatic hydrolysis (Chrysosporium lucknowense), integrated with dry mill corn production | 1.8 (0.075 from corn stover) |
| Abengoa | Salamanca, Spain | Wheat straw, cereal | Steam pretreatment, enzymatic hydrolysis | 4.92 |
| ALICO, Inc. | LaBelle, Fla., USA | Yard and citrus wastes | Gasification, fermentation of syngas | 52.6 |
| BioEthanol Japan | Osaka, Japan | Wood construction waste | Enzymatic hydrolysis, fermentation (Klebsiella oxytoca and E. coli) | 1.4 |
| BioFuels Energy Corp. | Raymondville, Texas, USA | Grass and tree trimmings | 15.14 | |
| BlueFire Ethanol | Lancaster, Calif., USA | Green waste | Acid hydrolysis, fermentation | 11.73 |
| Borregaard Industries Ltd | Sarpsborg, Norway | Wood | Wet inceneration | 20 |
| Colusa Biomass Energy Coporation | Colusa, Calif., USA | Rice straw and hulls | Acid hydrolysis, fermentation | 75.7 |
| China Resources Alcohol Corporation | ZhaoDong city, Heilongjiang province | Corn stover | Steam pre-treatment, enzymatic hydrolysis | 6.4 |
| DuPont-BP Biofuels | Wissington, England | Sugar beets | Enzymatic hydrolysis, fermentation to biobutanol | 34 |
| Iogen | Shelley, Idaho, USA | Wheat straw, barley straw, corn stover, switchgrass and rice straw | Enzymatic hydrolysis (Trichoderma reesei, Saccharomyces) | 68 |
| Iogen | Ottawa, Canada | Wheat, oat and barley straw | Enzymatic hydrolysis (Trichoderma reesei, Saccharomyces) | 3 |
| Lignol | Vancouver, Canada | Softwood and hardwood | Pulping liquor pretreatment, enzymatic hydrolysis | 4.92 |
| Mascoma | Rochester, N.Y., USA | Paper sludge, wood chips, switch grass and corn stover. | Enzymatic hydrolysis and fermentation (Thermoanaero-bacterium saccharolyticum) | 1.9 |
| Poet | Emmetsburg, Iowa, USA | Corn fiber, corn stover | Enzymatic hydrolysis, integrated with dry milling | 113.5 |
| Range Fuels | Soperton, Ga., USA | Timber and forest residue | Pyrolysis and catalytic conversion | 151.41 |
| Verenium | Jennings, La., USA | Sugarcane bagasse and specially bred energy cane | Enzymatic hydrolysis, fermentation (Klebsiella oxytoca and E. coli) | 5.3 |
| Western Biomass | Upton, Wyo., USA | Ponderosa pine wood chips, waste | CO2 pretreatment, enzymatic hydrolysis | 3.785 |
Based on the currently available technology the capital investments for cellulose-based ethanol production are estimated to be in the range of $1.06 to $1.48 per litre of ethanol annual capacity.138 The operation costs associated with these plants are between $0.35 and $0.45 per litre depending on the type of feedstock and technologies selected. The on-going research in biofuel conversion technologies are expected to reduce the capital investment needed to $0.95–$1.27 per litre ethanol annual capacity and to reduce the operating cost to $0.11–$0.25 per litre of ethanol.103 However, large and risky investments are required for the technological breakthroughs necessary to achieve the above expected cost reductions.
The current research tendencies for improving fuel ethanol production are linked to the nature of the raw materials, processing steps, and related process engineering issues. For the large-scale commercial production of ethanol from lignocellulosic material, a strong connection between the feedstock selection and downstream processing has to be developed. Table 10 summarizes the different strategies to improve the economics of lignocellulosic-derived bioethanol.
| Attribute | Advantages | Limitations | Way forward |
|---|---|---|---|
| Microorganisms | Numerous microorganisms have been engineered to selectively produce ethanol | Currently, there are no known natural organisms that have the ability to convert both C5 and C6 sugars to produce high ethanol yields | It is clear that we are still in the early stages of engineering organisms for biofuel production, as manifested by the many different organisms being explored concurrently for their potential to be the most relevant strain |
| The lack of industrially suitable microorganisms for converting 5-carbon of biomass into fuel ethanol has traditionally been cited as a major technical roadblock in the production of ethanol from lignocellulosic material | In the future, genomics and pathway engineering should considerably facilitate the development of a variety of organisms able to use the full repertoire of cellulosic and hemicellulosic sugars and tolerate high ethanol concentrations to optimize ethanol yields | ||
| Research in this field to produce genetically engineered microbes is an important way to enhance the lignocellulose conversion yield | |||
| Process integration | This approach benefits in terms of lower capital and operating costs, as well as ensuring the optimum production of valuable co-products | Microorganisms with the combination of substrate-utilization and product formation properties required for CBP are not currently available | Due to the large number of individual processes in the overall conversion of ligno-cellulosic biomass into ethanol, there remains considerable potential for process integration |
| CBP requires a highly engineered microorganism developed for several different process-specific characteristics | Given that second-generation process development is still at the precommercial stage, it may take some time to determine the most efficient process pathway and system | ||
| The use of high-solids loadings in biomass-pre-treatment (in all the unit operations) | Increased sugar and ethanol yields | Lack of free water, the high viscosities, and the increased production of inhibitors must be overcome in order to achieve economically viable sugar and ethanol yields | Researchers are tackling these problems on two fronts: reactor design and pre-treatment optimization |
| More efficient processes compared to the conventional conversion process | High solids concentrations and the recycling of process streams increase the concentration of compounds that are inhibitory to enzymatic hydrolysis and fermentation, necessitating detoxification or fed-batch technology | Horizontal paddle reactors and percolation reactors have both been shown to be possible alternatives to standard reactor designs when it comes to high solids | |
| Decreased capital and production costs | The choice of pretreatment can also affect the effectiveness of the overall conversion process | ||
| Decreased water and power use | |||
| Producing enzymes from lower cost feedstocks | Enzymes are intrinsically expensive because they must be produced by living systems and are thermodynamically unstable | The contribution of enzymes to the total production cost can be lowered by shifting to lower cost feedstocks reducing the fermentation times, and reducing the complexity of the process to drive down capital cost | |
| Due to the chemical and physical recalcitrance of lignocellulose, high enzyme loadings are necessary to obtain reasonable degradation rates | Much is to be gained by achieving high overall biofuel yields at low enzyme loadings. This fact places, indirectly, emphasis on the need for the development of improved pre-treatment and enzyme technologies | ||
| Commercially available enzyme mixtures | Use of commercial enzymes may be a quicker route to commercialisation | The biggest drawback of using commercial enzyme mixtures is the lack of characterization of the enzymes in the mixtures | Cellulase research efforts are focused on developing a cost-effective, highly thermo stable, synergistically acting enzyme mixture that would meet the end user's needs |
| Most of these mixtures have been developed for optimal hydrolysis on acid-pre-treated corn stover and are thus not optimal for other substrates and other pre-treatments | |||
| Reducing the energy demand of the process | High co-product yield requires a reduced energy demand for ethanol production | In the short term, co-products are likely to be used for the production of fuel, heat and electricity | |
| Long term, bioethanol technology will form the basis for the sustainable production of commodity chemicals and materials in future biorefineries | |||
| Commercial cellulosic ethanol production | Bio-fuels provide the opportunity for non-oil-producing countries to be less dependent on oil import | Given the current investments being made to make improvements in the process technologies, some expectations have arisen that 2nd-generation biofuel production will reach full commercialisation in the near future | |
| Bio-ethanol can be produced from any biomass, thus access to raw material is virtually unlimited | |||
| Integrated approach | Promotion of second generation biofuels can help provide solutions to multiple policy drivers including energy security and diversification, rural economic development, and GHG mitigation | The need to accommodate the variability in biomass feedstock and to manipulate ethanol and sugar tolerance to potential inhibitors generated in the pre-saccharification treatment remains a scientific challenge | Policies designed to specifically support the promotion of second generation biofuels must be carefully developed if they are to avoid unwanted consequences and potentially delay commercialisation |
| Policies like a national carbon price or other CO2 reduction incentives could help produce a more competitive market for biofuels | |||
| Funding for 2nd-generation biofuel demonstration plants are needed from both the public and private sector |
Policy framework
Bio-fuels provide the opportunity for non-oil-producing countries to lessen their dependence on oil import. Bio-ethanol can be produced from any biomass, thus access to raw material is virtually unlimited.139 The promotion of second-generation biofuels can help provide solutions to multiple policy drivers. including energy security and diversification, rural economic development, and greenhouse gas mitigation. It can also assist with the reduction of other environmental impacts, at least those relative to the use of other transport fuels. Improving technology performance and process design can help in lowering the environmental impacts in the whole life cycle.24 Policies designed to specifically support the promotion of second generation biofuels must be carefully developed if they are to avoid unwanted consequences and potentially delay commercialization.55 Through the combined efforts of a diverse range of scientists and engineers, we will succeed in making economical and sustainable biofuel production a reality.The capacity of biofuels to simultaneously advance multiple policy goals should be considered while designing incentive mechanisms. Published research has focused on upgrading many different types of feedstocks. To maintain year-round production, feedstock-blending studies could be a topic of future research. Overall, many recent advances in enzyme and microorganism development, optimization, and utilization for lignocellulosic biomass conversion have been reported, and with continued research in the area of process development for lignocellulosic biomass, implementation of renewable biofuels and a corresponding reduction in fossil fuel use can be envisioned.140 The high ethanol yield and utilizing the pentose fraction for ethanol production improves the process economy.141
The National Renewable Energy Laboratory (NREL) of the United States Department of Energy projects that by 2030, technology developments will enable yields of ethanol to approach some 400 liters per dry metric ton of biomass feedstock converted compared with about 270 liters per ton that have been achieved with known technology today. In the longer term both low feedstock costs and large scale production are projected to bring down the production costs.
Conclusion
The production of ethanol from lignocellulosic material at a commercial scale remains a challenging issue. For current technologies employed at a commercial level, the main share in the cost structure corresponds to the feedstock, followed by the processing expenditures. The current research tendencies for improving fuel ethanol production are linked to the nature of the employed raw materials, processing steps, and related process engineering issues. This review has discussed different options for the biochemical conversion of sugar derived from different lignocellulosic materials into ethanol. Based on the above a discussion of the different strategies to improve the economics of lignocellulosic-derived ethanol are summarized in Table 10. More fundamental studies are required to increase the understanding of how the pre-treatment affects the different types of biomass on a structural and molecular level and how that interacts with the subsequent hydrolysis and fermentation for different feedstocks. Research should concentrate on the pre-treatment and hydrolyzing of lignocellulosic material as ways to minimize the formation of fermentation inhibitors. The key to establishing a commercial ethanol production facility is lessening the number of operations and operating costs of each unit of operation. The commercialization of CBP would represent a breakthrough for low-cost biomass processing, due to economic benefits of process integration and avoiding the high costs of enzymes that at present make the biochemical conversion route unattractive. The full capacity for microbial biofuel production is yet to be realized. Further progress in metabolically engineered microorganism has the potential to improve the strain activity to overcome the slow catalysis and low fermentation efficiency.These developments will also facilitate the low cost production of commodities, such as alcohols, fatty acids, isoprenoid and organic acids.
Nomenclature
| A | Enzyme activity in FPU per g protein |
| C | Cellulose content of the biomass (%) |
| E | Price of enzyme in $ per l ethanol |
| P | Price of protein in $ per kg |
| L | Enzyme loading in FPU per g cellulose |
| Y | Ethanol yield in l bdt−1(litre per dry biomass ton). |
Notes and references
- P. S. Nigam and A. Singh, Prog. Energy Combust. Sci., 2011, 37, 52–68 CrossRef CAS PubMed.
- M. J. Groom, E. M. Gray and P. A. Townsend, Conserv. Biol., 2008, 22, 602–609 CrossRef PubMed.
- A. Muller, Clim. Change, 2009, 94, 319–331 CrossRef CAS PubMed.
- G. W. Huber, S. Iborra and A. Corma, Chem. Rev., 2006, 106, 4044–4098 CrossRef CAS PubMed.
- M. Jin, C. Gunawan, N. Uppugundla, V. Balan and B. E. Dale, Energy Environ. Sci., 2012, 5, 7168–7175 CAS.
- R. Luque, L. Herrero-Davila, J. M. Campelo, J. H. Clark, J. M. Hidalgo, D. Luna, J. M. Marinas and A. A. Romero, Energy Environ. Sci., 2008, 1, 542–564 CAS.
- I. P. Samayam and C. A. Schall, Bioresour. Technol., 2010, 101, 3561–3566 CrossRef CAS PubMed.
- P. Langan, S. Gnanakaran, K. D. Rector, N. Pawley, D. T. Fox, D. W. Cho and K. E. Hammel, Energy Environ. Sci., 2011, 4, 3820–3833 CAS.
- T. Damartzis and A. Zabaniotou, Renewable Sustainable Energy Rev., 2011, 15, 366–378 CrossRef CAS PubMed.
- W.-D. Huang and Y. H. P. Zhang, Energy Environ. Sci., 2011, 4, 784–792 CAS.
- Y. Sun and J. Cheng, Bioresour. Technol., 2002, 83, 1–11 CrossRef CAS.
- N. Mosier, C. Wyman, B. Dale, R. Elander, Y. Y. Lee, M. Holtzapple and M. Ladisch, Bioresour. Technol., 2005, 96, 673–686 CrossRef CAS PubMed.
- C. A. Cardona and Ó. J. Sánchez, Bioresour. Technol., 2007, 98, 2415–2457 CrossRef CAS PubMed.
- C. A. Cardona, Ó. J. Sánchez and L. F. Gutiérrez, in Process Synthesis for Fuel Ethanol Production, CRC Press, 2009, pp. 221–283 Search PubMed.
- W. E. Mabee, P. N. McFarlane and J. N. Saddler, Biomass Bioenergy, 2011, 35, 4519–4529 CrossRef PubMed.
- G. Brodeur, E. Yau, K. Badal, J. Collier, K. B. Ramachandran and S. Ramakrishnan, Enzyme Res., 2011, 2011, 787532 Search PubMed.
- G. Rødsrud, M. Lersch and A. Sjöde, Biomass Bioenergy, 2012, 46, 46–59 CrossRef PubMed.
- N. Xu, W. Zhang, S. Ren, F. Liu, C. Zhao, H. Liao, Z. Xu, J. Huang, Q. Li, Y. Tu, B. Yu, Y. Wang, J. Jiang, J. Qin and L. Peng, Biotechnol. Biofuels, 2012, 5, 58 CrossRef CAS PubMed.
- W. Zhang, Z. Yi, J. Huang, F. Li, B. Hao, M. Li, S. Hong, Y. Lv, W. Sun, A. Ragauskas, F. Hu, J. Peng and L. Peng, Bioresour. Technol., 2013, 130, 30–37 CrossRef CAS PubMed.
- M. Galbe and G. Zacchi, Biomass Bioenergy, 2012, 46, 70–78 CrossRef CAS PubMed.
- A. Limayem and S. C. Ricke, Prog. Energy Combust. Sci., 2012, 38, 449–467 CrossRef CAS PubMed.
- L. Viikari, J. Vehmaanperä and A. Koivula, Biomass Bioenergy, 2012, 46, 13–24 CrossRef CAS PubMed.
- D. Chiaramonti, M. Prussi, S. Ferrero, L. Oriani, P. Ottonello, P. Torre and F. Cherchi, Biomass Bioenergy, 2012, 46, 25–35 CrossRef CAS PubMed.
- A. L. Borrion, M. C. McManus and G. P. Hammond, Renewable Sustainable Energy Rev., 2012, 16, 4638–4650 CrossRef PubMed.
- F. Hu and A. Ragauskas, BioEnergy Res., 2012, 5, 1043–1066 CrossRef CAS.
- B. Joshi, M. R. Bhatt, D. Sharma, J. Joshi, R. Malla and L. Sreerama, Biotechnol. Mol. Biol. Rev., 2011, 172–182 CAS.
- M. Dashtban, H. Schraft and W. Qin, Int. J. Biol. Sci., 2009, 5, 578–595 CAS.
- P. Harnpicharnchai, V. Champreda, W. Sornlake and L. Eurwilaichitr, Protein Expression Purif., 2009, 67, 61–69 CrossRef CAS PubMed.
- J. S. Van Dyk and B. I. Pletschke, Biotechnol. Adv., 2012, 30, 1458–1480 CrossRef CAS PubMed.
- B. Hahn-Hägerdal, M. Galbe, M. F. Gorwa-Grauslund, G. Lidén and G. Zacchi, Trends Biotechnol., 2006, 24, 549–556 CrossRef PubMed.
- L. R. Lynd, W. H. v. Zyl, J. E. McBride and M. Laser, Curr. Opin. Biotechnol., 2005, 16, 577–583 CrossRef CAS PubMed.
- Z. Zhang, A. A. Donaldson and X. Ma, Biotechnol. Adv., 2012, 30, 913–919 CrossRef CAS PubMed.
- Z.-X. Lin, H.-M. Zhang, X.-J. Ji, J.-W. Chen and H. Huang, Appl. Biochem. Biotechnol., 2011, 164, 23–33 CrossRef CAS PubMed.
- B. Sipos, Z. Benkő, D. Dienes, K. Réczey, L. Viikari and M. Siika-aho, Appl. Biochem. Biotechnol., 2010, 161, 347–364 CrossRef CAS PubMed.
- R. L. Howard, E. Abotsi, E. L. Jansen van Rensburg and S. Howard, Afr. J. Biotechnol., 2003, 2, 602–661 CAS.
- J. D. Stephen, W. E. Mabee and J. N. Saddler, Biofuels, Bioprod. Biorefin., 2011, 6, 159–176 Search PubMed.
- S. Decker, W. Adney, E. Jennings, T. Vinzant and M. Himmel, Appl. Biochem. Biotechnol., 2003, 107, 689–703 CrossRef.
- A. Dutta, N. Dowe, K. N. Ibsen, D. J. Schell and A. Aden, Biotechnol. Prog., 2010, 26, 64–72 CAS.
- A. Dutta, M. Talmadge, J. Hensley, M. Worley, D. Dudgeon, D. Barton, P. Groendijk, D. Ferrari, B. Stears, E. M. Searcy, C. T. Wright and J. R. Hess, Process Design and Economics for Conversion of Lignocellulosic Biomass to Ethanol: Thermochemical Pathway by Indirect Gasification and Mixed Alcohol Synthesis, NREL, Oak Ridge, TN 37831-0062, 2011 Search PubMed.
- G. Banerjee, J. Scott-Craig and J. Walton, BioEnergy Res., 2010, 3, 82–92 CrossRef PubMed.
- K. Kadam, in Handbook on Bioethanol: Production and Utilization, ed. C. E. Wyman, Taylor & Francis, Washington, DC, USA, 1996, pp. 213–252 Search PubMed.
- M. Tu, R. P. Chandra and J. N. Saddler, Biotechnol. Prog., 2007, 23, 1130–1137 CrossRef CAS PubMed.
- Y. Lu, B. Yang, D. Gregg, J. Saddler and S. Mansfield, Appl. Biochem. Biotechnol., 2002, 98–100, 641–654 CrossRef CAS.
- B. Qi, X. Chen, Y. Su and Y. Wan, Bioresour. Technol., 2011, 102, 2881–2889 CrossRef CAS PubMed.
- Y.-H. P. Zhang and L. R. Lynd, Biotechnol. Bioeng., 2004, 88, 797–824 CrossRef CAS PubMed.
- A. Aden and T. Foust, Cellulose, 2009, 16, 535–545 CrossRef CAS.
- L. R. Lynd, M. S. Laser, D. Bransby, B. E. Dale, B. Davison, R. Hamilton, M. Himmel, M. Keller, J. D. McMillan, J. Sheehan and C. E. Wyman, Nat. Biotechnol., 2008, 26, 169–172 CrossRef CAS PubMed.
- D. Klein-Marcuschamer, P. Oleskowicz-Popiel, B. A. Simmons and H. W. Blanch, Biomass Bioenergy, 2010, 34, 1914–1921 CrossRef CAS PubMed.
- F. K. Kazi, J. A. Fortman, R. P. Anex, D. D. Hsu, A. Aden, A. Dutta and G. Kothandaraman, Fuel, 2010, 89(suppl. 1), S20–S28 CrossRef CAS PubMed.
- D. Klein-Marcuschamer, P. Oleskowicz-Popiel, B. A. Simmons and H. W. Blanch, Biotechnol. Bioeng., 2012, 109, 1083–1087 CrossRef CAS PubMed.
- M. Knauf and M. Moniruzzaman, Int. Sugar J., 2004, 106, 147–150 CAS.
- S. Merino and J. Cherry, ed. L. Olsson, Biofuels, Springer, Berlin/Heidelberg, 2007, vol. 108, pp. 95–120 Search PubMed.
- Q. Qing and C. E. Wyman, Bioresour. Technol., 2011, 102, 1359–1366 CrossRef CAS PubMed.
- J. A. Roels and N. W. F. Kossen, in Progress in Industrial Microbiology, ed. M. J. Bull, Elsevier, 1978, p. 294 Search PubMed.
- R. E. H. Sims, W. Mabee, J. N. Saddler and M. Taylor, Bioresour. Technol., 2010, 101, 1570–1580 CrossRef CAS PubMed.
- G. Stephanopoulos, Science, 2007, 315, 801–804 CrossRef CAS PubMed.
- S. Sakai, Y. Tsuchida, S. Okino, O. Ichihashi, H. Kawaguchi, T. Watanabe, M. Inui and H. Yukawa, Appl. Environ. Microbiol., 2007, 73, 2349–2353 CrossRef CAS PubMed.
- M. von Sivers and G. Zacchi, Bioresour. Technol., 1996, 56, 131–140 CrossRef CAS.
- A. J. Ragauskas, C. K. Williams, B. H. Davison, G. Britovsek, J. Cairney, C. A. Eckert, W. J. Frederick, J. P. Hallett, D. J. Leak, C. L. Liotta, J. R. Mielenz, R. Murphy, R. Templer and T. Tschaplinski, Science, 2006, 311, 484–489 CrossRef CAS PubMed.
- H. Alper and G. Stephanopoulos, Nat. Rev. Microbiol., 2009, 7, 715–723 CrossRef CAS PubMed.
- B. Hahn-Hägerdal, K. Karhumaa, C. Fonseca, I. Spencer-Martins and M. Gorwa-Grauslund, Appl. Microbiol. Biotechnol., 2007, 74, 937–953 CrossRef PubMed.
- C. Verduyn, E. Postma, W. A. Scheffers and J. P. van Dijken, J. Gen. Microbiol., 1990, 136, 395–403 CrossRef CAS PubMed.
- G. P. Casey and W. M. M. Ingledew, Crit. Rev. Microbiol., 1986, 13, 219–280 CrossRef CAS PubMed.
- D. la Grange, R. den Haan and W. van Zyl, Appl. Microbiol. Biotechnol., 2010, 87, 1195–1208 CrossRef PubMed.
- Y. Lin, W. Zhang, C. Li, K. Sakakibara, S. Tanaka and H. Kong, Biomass Bioenergy, 2012, 47, 395–401 CrossRef CAS PubMed.
- M. Kuyper, A. A. Winkler, J. P. van Dijken and J. T. Pronk, FEMS Yeast Res., 2004, 4, 655–664 CrossRef CAS PubMed.
- M. Zhang, C. Eddy, K. Deanda, M. Finkelstein and S. Picataggio, Science, 1995, 267, 240–243 CrossRef CAS PubMed.
- B. S. Dien, M. A. Cotta and T. W. Jeffries, Appl. Microbiol. Biotechnol., 2003, 63, 258–266 CrossRef CAS PubMed.
- K. Karhumaa, B. Wiedemann, B. Hahn-Hagerdal, E. Boles and M.-F. Gorwa-Grauslund, Microb. Cell Fact., 2006, 5, 18 CrossRef PubMed.
- H. Danner and R. Braun, Chem. Soc. Rev., 1999, 28, 395–405 RSC.
- B. C. H. Chu and H. Lee, Biotechnol. Adv., 2007, 25, 425–441 CrossRef CAS PubMed.
- J. C. Preez, B. Driessel and B. A. Prior, Appl. Microbiol. Biotechnol., 1989, 30, 53–58 CrossRef.
- B. Hahn-Hägerdal and N. Pamment, Appl. Biochem. Biotechnol., 2004, 116, 1207–1209 CrossRef.
- J. N. Nigam, J. Biotechnol., 2001, 87, 17–27 CrossRef CAS.
- T. W. Jeffries, I. V. Grigoriev, J. Grimwood, J. M. Laplaza, A. Aerts, A. Salamov, J. Schmutz, E. Lindquist, P. Dehal, H. Shapiro, Y.-S. Jin, V. Passoth and P. M. Richardson, Nat. Biotechnol., 2007, 25, 319–326 CrossRef CAS PubMed.
- F. Agbogbo and G. Coward-Kelly, Biotechnol. Lett., 2008, 30, 1515–1524 CrossRef CAS PubMed.
- C. Van Zyl, B. Prior and J. Du Preez, Appl. Biochem. Biotechnol., 1988, 17, 357–369 CrossRef CAS.
- I. C. Roberto, L. S. Lacis, M. F. S. Barbosa and I. M. de Mancilha, Process Biochem., 1991, 26, 15–21 CrossRef CAS.
- M. E. Ligthelm, B. A. Prior and J. C. Preez, Appl. Microbiol. Biotechnol., 1988, 28, 63–68 CAS.
- T. W. Jeffries, Curr. Opin. Biotechnol., 1996, 7, 337–342 CrossRef CAS.
- R. Berson, J. Young, S. Kamer and T. Hanley, Appl. Biochem. Biotechnol., 2005, 124, 923–934 CrossRef.
- S. Tian, G. Zhou, F. Yan, Y. Yu and X. Yang, Biotechnol. Adv., 2009, 27, 656–660 CrossRef CAS PubMed.
- F. Alterthum and L. O. Ingram, Appl. Environ. Microbiol., 1989, 55, 1943–1948 CAS.
- Z. Zheng, T. Chen, M. Zhao, Z. Wang and X. Zhao, Microb. Cell Fact., 2012, 11, 37 CrossRef CAS PubMed.
- A. Asghari, R. J. Bothast, J. B. Doran and L. O. Ingram, J. Ind. Microbiol. Biotechnol., 1996, 16, 42–47 CrossRef CAS.
- J.-S. Seo, H. Chong, H. S. Park, K.-O. Yoon, C. Jung, J. J. Kim, J. H. Hong, H. Kim, J.-H. Kim, J.-I. Kil, C. J. Park, H.-M. Oh, J.-S. Lee, S.-J. Jin, H.-W. Um, H.-J. Lee, S.-J. Oh, J. Y. Kim, H. L. Kang, S. Y. Lee, K. J. Lee and H. S. Kang, Nat. Biotechnol., 2005, 23, 63–68 CrossRef CAS PubMed.
- M. Zhang, Y. C. Chou, S. K. Picataggio, M. Finkelstein, US Pat., 5843760, 1997.
- A. Mohagheghi, K. Evans, Y.-C. Chou and M. Zhang, Appl. Biochem. Biotechnol., 2002, 98–100, 885–898 CrossRef CAS.
- J. Zaldivar, J. Nielsen and L. Olsson, Appl. Microbiol. Biotechnol., 2001, 56, 17–34 CrossRef CAS.
- T. W. Jeffries, Trends Biotechnol., 1985, 3, 208–212 CrossRef CAS.
- M. W. Lau and B. E. Dale, Proc. Natl. Acad. Sci. U. S. A., 2009, 106, 1368–1373 CrossRef CAS PubMed.
- M. Lau, C. Gunawan, V. Balan and B. Dale, Biotechnol. Biofuels, 2010, 3, 11 CrossRef PubMed.
- E. M. Rubin, Nature, 2008, 454, 841–845 CrossRef CAS PubMed.
- C. Dellomonaco, F. Fava and R. Gonzalez, Microb. Cell Fact., 2010, 9, 3 CrossRef PubMed.
- D. Levišauskas and T. Tekorius, Inform. Tech. Contr., 2005, 34, 231–236 Search PubMed.
- J. C. Gunther, J. S. Conner and D. E. Seborg, Biotechnol. Prog., 2007, 23, 851–857 CAS.
- J. C. Gunther, D. E. Seborg and J. Baclaski, Fault detection and diagnosis in industrial fed-batch fermentation, Minneapolis, MN, 2006 Search PubMed.
- O. F. Echegaray, J. C. M. Carvalho, A. N. R. Fernandes, S. Sato, E. Aquarone and M. Vitolo, Biomass Bioenergy, 2000, 19, 39–50 CrossRef CAS.
- S. Alfenore, X. Cameleyre, L. Benbadis, C. Bideaux, J. L. Uribelarrea, G. Goma, C. Molina-Jouve and S. E. Guillouet, Appl. Microbiol. Biotechnol., 2004, 63, 537–542 CrossRef CAS PubMed.
- Ó. J. Sánchez and C. A. Cardona, Bioresour. Technol., 2008, 99, 5270–5295 CrossRef PubMed.
- B. C. Saha, L. B. Iten, M. A. Cotta and Y. V. Wu, Biotechnol. Prog., 2005, 21, 816–822 CrossRef CAS PubMed.
- F. M. Gírio, C. Fonseca, F. Carvalheiro, L. C. Duarte, S. Marques and R. Bogel-Łukasik, Bioresour. Technol., 2010, 101, 4775–4800 CrossRef PubMed.
- C. N. Hamelinck, G. v. Hooijdonk and A. P. C. Faaij, Biomass Bioenergy, 2005, 28, 384–410 CrossRef CAS PubMed.
- K. Olofsson, M. Bertilsson and G. Liden, Biotechnol. Biofuels, 2008, 1, 7 CrossRef PubMed.
- F. Xu, H. Ding, D. Osborn, A. Tejirian, K. Brown, W. Albano, N. Sheehy and J. Langston, J. Mol. Catal. B: Enzym., 2008, 51, 42–48 CrossRef CAS PubMed.
- K. Olofsson, A. Rudolf and G. Liden, J. Biotechnol., 2008, 134, 112–120 CrossRef CAS PubMed.
- K. Öhgren, O. Bengtsson, M. F. Gorwa-Grauslund, M. Galbe, B. Hahn-Hägerdal and G. Zacchi, J. Biotechnol., 2006, 126, 488–498 CrossRef PubMed.
- K. Olofsson, M. Wiman and G. Liden, J. Biotechnol., 2010, 145, 168–175 CrossRef CAS PubMed.
- B. A. Faga, M. R. Wilkins and I. M. Banat, Bioresour. Technol., 2010, 101, 2273–2279 CrossRef CAS PubMed.
- M. P. García-Aparicio, J. M. Oliva, P. Manzanares, M. Ballesteros, I. Ballesteros, A. González and M. J. Negro, Fuel, 2011, 90, 1624–1630 CrossRef PubMed.
- Y. Kim, R. Hendrickson, N. S. Mosier, M. R. Ladisch, B. Bals, V. Balan, B. E. Dale, B. S. Dien and M. A. Cotta, Bioresour. Technol., 2010, 101, 5385–5393 CrossRef CAS PubMed.
- B.-Z. Li, V. Balan, Y.-J. Yuan and B. E. Dale, Bioresour. Technol., 2010, 101, 1285–1292 CrossRef CAS PubMed.
- Q. Shao, S. Chundawat, C. Krishnan, B. Bals, L. Sousa, K. Thelen, B. Dale and V. Balan, Biotechnol. Biofuels, 2010, 3, 12 CrossRef PubMed.
- J. Zhang and L. R. Lynd, Biotechnol. Bioeng., 2010, 107, 235–244 CrossRef CAS PubMed.
- C. Zhong, M. Lau, V. Balan, B. Dale and Y.-J. Yuan, Appl. Microbiol. Biotechnol., 2009, 84, 667–676 CrossRef CAS PubMed.
- S. I. Mussatto, G. Dragone, P. M. R. Guimarães, J. P. A. Silva, L. M. Carneiro, I. C. Roberto, A. Vicente, L. Domingues and J. A. Teixeira, Biotechnol. Adv., 2010, 28, 817–830 CrossRef CAS PubMed.
- L. R. Lynd, P. J. Weimer, W. H. van Zyl and I. S. Pretorius, Microbiol. Mol. Biol. Rev., 2002, 66, 506–577 CrossRef CAS.
- T. Hasunuma and A. Kondo, Biotechnol. Adv., 2012, 30, 1207–1218 CrossRef CAS PubMed.
- T. Hasunuma, F. Okazaki, N. Okai, K. Y. Hara, J. Ishii and A. Kondo, Bioresour. Technol., 2013, 135, 513–522 CrossRef CAS PubMed.
- V. Menon and M. Rao, Prog. Energy Combust. Sci., 2012, 38, 522–550 CrossRef CAS PubMed.
- C. C. Geddes, I. U. Nieves and L. O. Ingram, Curr. Opin. Biotechnol., 2011, 22, 312–319 CrossRef CAS PubMed.
- P. P. Peralta-Yahya, F. Zhang, S. B. del Cardayre and J. D. Keasling, Nature, 2012, 488, 320–328 CrossRef CAS PubMed.
- C. A. Rabinovitch-Deere, J. W. K. Oliver, G. M. Rodriguez and S. Atsumi, Chem. Rev., 2013, 113, 4611–4632 CrossRef CAS PubMed.
- S.-J. Ha, J. M. Galazka, S. Rin Kim, J.-H. Choi, X. Yang, J.-H. Seo, N. Louise Glass, J. H. D. Cate and Y.-S. Jin, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 504–509 CrossRef CAS PubMed.
- F. Wen, J. Sun and H. Zhao, Appl. Environ. Microbiol., 2010, 76, 1251–1260 CrossRef CAS PubMed.
- J. Sun, F. Wen, T. Si, J.-H. Xu and H. Zhao, Appl. Environ. Microbiol., 2012, 78, 3837–3845 CrossRef CAS PubMed.
- S. Kim, S.-H. Baek, K. Lee and J.-S. Hahn, Microb. Cell Fact., 2013, 12, 14 CrossRef CAS PubMed.
- T. Liu and C. Khosla, Annu. Rev. Genet., 2010, 44, 53–69 CrossRef CAS PubMed.
- Y. Zheng, Q. Liu, L. Li, W. Qin, J. Yang, H. Zhang, X. Jiang, T. Cheng, W. Liu, X. Xu and M. Xian, Biotechnol. Biofuels, 2013, 6, 57 CrossRef CAS PubMed.
- S. Bastian, X. Liu, J. T. Meyerowitz, C. D. Snow, M. M. Y. Chen and F. H. Arnold, Metab. Eng., 2011, 13, 345–352 CrossRef CAS PubMed.
- E. I. Lan and J. C. Liao, Proc. Natl. Acad. Sci. U. S. A., 2012, 109, 6018–6023 CrossRef CAS PubMed.
- T. Liu, H. Vora and C. Khosla, Metab. Eng., 2010, 12, 378–386 CrossRef CAS PubMed.
- T. Hasunuma and A. Kondo, Biotechnol. Adv., 2012, 30, 1207–1218 CrossRef CAS PubMed.
- D. Hodge, M. Karim, D. Schell and J. McMillan, Appl. Biochem. Biotechnol., 2009, 152, 88–107 CrossRef CAS PubMed.
- M. Haque, Doctor of Philosophy, Oklahoma State University, 2010.
- M. A. Carriquiry, X. Du and G. R. Timilsina, Energy Policy, 2011, 39, 4222–4234 CrossRef PubMed.
- N. Sarkar, S. K. Ghosh, S. Bannerjee and K. Aikat, Renewable Energy, 2012, 37, 19–27 CrossRef CAS PubMed.
- M. M. Wright and R. C. Brown, Biofuels, Bioprod. Biorefin., 2007, 1, 49–56 CrossRef CAS.
- C. N. Hamelinck and A. P. C. Faaij, Energy Policy, 2006, 34, 3268–3283 CrossRef PubMed.
- J. C. Hicks, J. Phys. Chem. Lett., 2011, 2, 2280–2287 CAS.
- P. Sassner, M. Galbe and G. Zacchi, Biomass Bioenergy, 2008, 32, 422–430 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2014 |