The occurrence and biological significance of the α-keto-sugars pseudaminic acid and legionaminic acid within pathogenic bacteria
Matthew Zunk†
and
Milton J. Kiefel
*
Institute for Glycomics, Griffith University Gold Coast, Parklands Drive, Southport, Queensland 4222, Australia. E-mail: m.kiefel@griffith.edu.au; Fax: +61 7 5552 8098; Tel: +61 7 5552 7021
First published on 25th November 2013
Abstract
This article describes the occurrence of the acidic sugars based on pseudaminic acid and legionaminic acid within pathogenic Gram-negative bacteria. In addition to presenting the structural variations found in these sugars, this article also discusses the potential biological role of these unusual sugars.
 Matthew Zunk | Matthew Zunk graduated from Griffith University with a Bachelor of Pharmaceutical Science, first class honours in 2008; being awarded academic excellence in 2004, 2006 and 2008 and receiving multiple scholarships for his academic achievements. Currently he is finalising his PhD at the Institute for Glycomics (Griffith University) under the supervision of Dr Milton J. Kiefel. Since 2012, Mr Zunk has held an academic position within the School of Pharmacy at Griffith University, Gold Coast Campus, teaching undergraduate Pharmaceutical Science and Drug Discovery and Development. His main research interests cover synthetic chemistry and the synthesis of bacterial carbohydrates. |
 Milton J. Kiefel | Milton J. Kiefel completed his PhD in organic chemistry at the University of Melbourne in 1990, before undertaking a postdoctoral position with Professor Gerald Pattenden (University of Nottingham, UK). In 1993 he joined the carbohydrate chemistry group of Prof Mark von Itzstein at Monash University, and in 2000 moved to Griffith University to take up a lecturing position within the Institute for Glycomics. In 2005 he was promoted to Senior Lecturer. His research interests involve the synthesis of novel carbohydrates that can be used as probes to elucidate the function of various carbohydrate-utilising proteins that are related to human diseases. |
Nonulosonic acids
Nonulosonic acids are a structural class of 9-carbon carbohydrate monomers that are α-keto acids. The most abundant naturally occurring nonulosonic acids are the sialic acids (or neuraminic acids) and their derivatives (Fig. 1).1,2 The sialic acid family is widely distributed throughout nature with over 50 naturally occurring derivatives having been identified to date within all domains of cellular organisms.1,3 Sialic acids have been found in nearly all mammalian tissues including embryonic tissues, and play an important role in a number of diseases. For example, overexpression of terminal sialylation mechanisms is directly associated with cancer malignancies.4 The natural occurrence of sialic acids is not limited to mammalian cells, with this structurally diverse family of compounds having been isolated from invertebrate and non-invertebrate species alike. Sialic acids have been isolated from insects, found in fish eggs, and they are increasingly being discovered within pathogenic and non-pathogenic bacteria and archaeal organisms.2,5,6 The importance of sialic acids means they have been discussed in detail elsewhere. For a detailed review of the diversity of sialic acids see articles by Varki,2,5 for an account of the structure, function and metabolism of sialic acids see Traving & Schauer,1 whilst the structural diversity and phylogenetic relationship of organisms encoding for sialic acids has been reviewed by Angata & Varki.3 | ||
| Fig. 1 Naturally occurring sialic acids N-acetylneuraminic acid (Neu5Ac (1)) and keto-deoxynonulosonic acid (KDN (2)). | ||
Our increased understanding of the crucial role that sialic acids play in the lifecycle of various organisms has driven the search for new nonulosonic acid derivatives. Through this process an important sub-class of nonulosonic acid derivative has been found that appear to be unique to bacterial species. These recently identified nonulosonic acids are structurally characterised as 5,7-diamino-3,5,7,9-tetradeoxy-nonulosonate derivatives of which two important parent compounds are known, pseudaminic acid (3) and legionaminic acid (4) (Fig. 2).
 | ||
| Fig. 2 Pseudaminic acid (3) and Legionaminic acid (4). | ||
The biosynthesis of all nonulosonic acids has a number of unique evolutionary aspects.3,5,6 The first is that a nucleotide-activated hexose sugar serves as a precursor to nine carbon α-keto acids. In the case of sialic acid derivatives, this hexose sugar is a D-mannose derivative, whilst compounds such as the pseudaminic and legionaminic acids are synthesised from a nucleotide activated N-acetylglucosamine. Secondly all known nine carbon α-keto acids are synthesised by an enzyme catalysed condensation reaction, whereby a six carbon unit is condensed with a 3 carbon unit (typically a pyruvate derivative) to form the desired nine carbon acidic sugar. Biosynthetically, certain chemical transformations must occur to the nucleotide-activated hexose sugar in order for the appropriate 6 carbon unit to be obtained, since it is the stereochemical and functional group detail that this hexose possesses that directly determines the structural characteristics of the nine carbon nonulosonate that will be generated. Lastly, all nonulosonic acids must be nucleotide activated (usually CMP-activated) in order to be incorporated into the varying structural glycans where they are found.7
Structural diversity and occurrence of 5,7-diamino-3,5,7,9-tetradeoxy-nonulosonates
In 1984, Knirel and colleagues characterised a new type of “sialic acid-like” sugar, which was extracted and purified from the O-chain polysaccharide of the LPS of Pseudomonas aeruginosa O7 and O9 and Shigella boydii type 7.8 This new compound was identified at the time as 5,7-diacetamido-3,5,7,9-tetradeoxy-L-glycero-L-manno-non-2-ulosonic acid, which Knirel generically named pseudaminic acid.8 Since 1984 there have been a number of naturally occurring pseudaminic acid derivatives discovered, which all have an L-glycero-L-manno configuration but vary in the types of N-acyl substitution they contain at C-5 and C-7. Following from this initial discovery, these derivatives are proving to be wide spread within bacterial glycan structures having been so far identified and isolated from a growing number of bacterial species (Table 1).| Nonulosonic acid | Bacterial source | Reference |
|---|---|---|
| L-glycero-L-manno (pseudaminic acid) | ||
 |
Escherichia coli O136 | 62 |
| Proteus vulgaris O39 | 63 | |
| Campylobacter jejuni 81-176 | 13 and 33 | |
| Campylobacter jejuni 11168 | 19 | |
| Campylobacter coli VC167 | 34 | |
| Pseudoateromonas atlantica T9 | 9 | |
| Helicobacter pylori 1061 | 12 and 33 | |
| Pseudoalteromonas atlantica IAM 14165 | 64 | |
| Aeromonas caviae UU51 | 33 | |
| Rhizobium sp. NGR234 | 65 | |
| Sinorhizobium meliloti Rm1021 | 66 and 67 | |
| Cellulophaga fucicola | 68 | |
| Vibrio vulnificus 27562 | 69 | |
| Piscirickettsia salmonis | 70 | |
 |
Pseudomonas aeruginosa O10a | 8 |
| Shigella boydii type 7 | 8, 71 and 72 | |
| Pseudomonas aeruginosa O9a, 9b | 21 and 71 | |
| Sinorhizobium fredii HH103 | 73 | |
| Kribbella spp. VKM | 74 and 75 | |
| Actinoplanes utahensis VKM Ac-674 | 74 and 76 | |
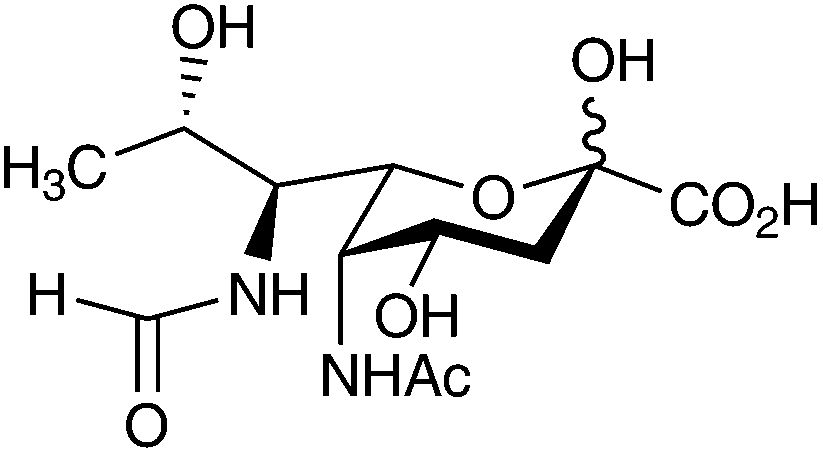 |
Pseudomonas aeruginosa O7a, 7b, 7d and O7a, 7d (immunotype 6) | 77 and 78 |
| Pseudoalteromonas distincta KMM 638 | 79 | |
 |
Vibrio cholera O:2 | 80 |
| Campylobacter jejuni 81-176 | 13 and 33 | |
| Campylobacter coli VC167 | 34 | |
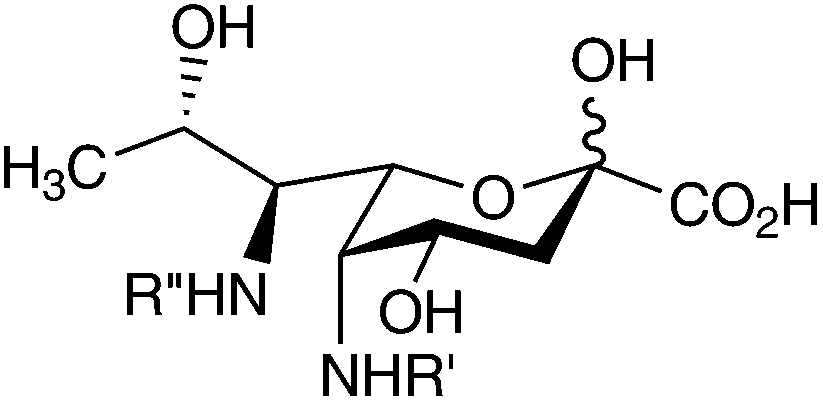 |
||
| R′ = Ac; R′′ = glycerate | Vibrio vulnificus 27562 | 69 |
R′ = C(![[double bond, length as m-dash]](https://www.rsc.org/images/entities/char_e001.gif) O)CH2CH(OH)CH2; R′′ = C(O)H O)CH2CH(OH)CH2; R′′ = C(O)H |
Pseudomonas aeruginosa O7a, 7b, 7c | 78 |
| R′ = R′′ = C(O)CH(OH)CH2OH |
Campylobacter jejuni 81-176 | 13 |
| R′ = Ac; R′′ = C(NH)CH3 |
Campylobacter jejuni 11168 | 19 |
| R′ = C(O)CH(OCH3)CH2OCH3; R′′ = C(NH)CH3 |
Campylobacter jejuni 11168 | 19 |
| R′ = C(O)CH(OCH3)CH2OCH3; R′′ = Ac |
Campylobacter jejuni 11168 | 19 |
| D-glycero-D-galacto (legionaminic acid) | ||
 |
Vibrio alginolyticus 945-80 | 22 and 24 |
| Acinetobacter baumannii O24 | 22 and 28 | |
| Vibrio parahaemolyticus O2 | 26 and 27 | |
| Campylobacter jejuni 11168 | 19 | |
| Escherichia coli O161 | 29 | |
 |
Legionella pneumophila serogroup 1 | 17 and 22 |
| Pseudomonas fluorescens ATCC 49271 | 22 and 23 | |
| Vibrio salmonicida NCMB 2262 | 22 and 25 | |
| Campylobacter coli VC167 | 20 | |
| Campylobacter jejuni 11168 | 19 | |
 |
||
| R′ = C(O)CH2CH(OH)CH3; R′′ = Ac |
Acinetobacter baumannii O24 | 22 and 28 |
| R′ = Ac; R′′ = C(O)CH(NHAc)CH3 |
Vibrio parahaemolyticus KX-V212 | 26 |
| Escherichia coli O161 | 29 | |
| R′ = CH(NHCH3)CH3; R′′ = Ac | Campylobacter coli VC167 | 20 |
| Campylobacter jejuni 11168 | 19 | |
| R′ = C(O)CH2CH2CH(CO2H)NHCH3; R′′ = Ac |
Clostridium botulinum | 81 |
| D-glycero-D-talo (4-epi-legionaminic acid) | ||
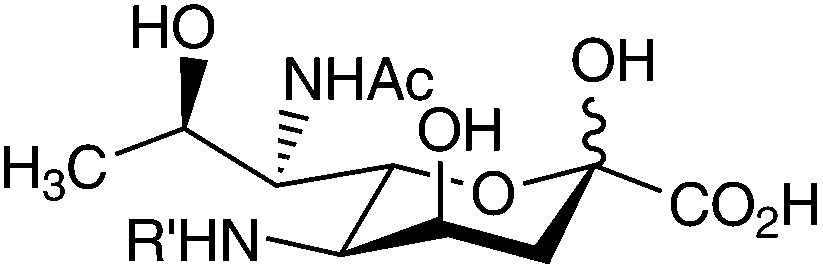 |
||
| R′ = Ac | Legionella pneumophila serogroup 1 | 22 and 39 |
| R′ = Ac | Legionella pneumophila serogroups 3, 4, 5,6,8,9,10,11,12,14, Lansing 3, 16453-92 | 40 |
| R′ = Ac | Shewanella japonica KMM 3601 | 41 |
| R′ = C(NH)CH3 |
Legionella pneumophila serogroup 2 | 40 |
| L-glycero-D-galacto (8-epi-legionaminic acid) | ||
 |
Pseudomonas aeruginosa O12 | 21 |
| Providencia stuartii O20 | 38 | |
| Escherichia coli O108 | 30 | |
 |
||
| R′ = C(NH)CH3; R′′ = Ac |
Morganella morganii KF 1676 (RK 4222) | 36 |
| R′ = Ac; R′′ = C(NH)CH3 |
Shewanella putrefaciens A6 | 37 |
| R′ = Ac; R′′ = C(O)CH(NHAc)CH3 |
Escherichia coli O108 | 31 |
| R′ = C(O)CH2CH(OH)CH3; R′′ = Ac |
Salmonella arizonae O61 | 32 |
| R′ = C(O)CH2CH2CH2OH; R′′ = Ac |
Yersinia ruckeri O1 | 35 |
Within these bacterial species pseudaminic acid and its derivatives are found as components of important cell surface glycans (oligosaccharides and glycoproteins), such as LPS O-antigens,9 capsular polysaccharides,10 pili11 and flagellin12,13 all of which are essential for bacterial virulence.14,15 Interestingly, unlike the sialic acids, pseudaminic acids are found within bacterial glycans as both α and β linked carbohydrates, being less commonly found at the terminal end of glycans. To date most of the glycans that have been characterised contain pseudaminic acid, and its naturally occurring derivatives, as internally linked sugars, often including linkages through the Nitrogen-based functionalities at C-5 and C-7, such as the C-7 linked pseudaminic acid derivative found in Kribbella spp. (Fig. 3).16
 | ||
| Fig. 3 An example of a teichulosonic acid fragment containing C-4 linked and C-7 nitrogen linked pseudaminic acid residue from Kribbella spp.16 | ||
Legionaminic acid (4) (5,7-diacetamido-3,5,7,9-tetradeoxy-D-glycero-D-galacto-non-2-ulosonic acid) was first discovered in 1994 as α-(2,4)-linked homopolymer component of the Legionella pneumophila serotype 1, O-chain LPS.17 L. pneumophila is the causative agent of Legionnaires' disease, which replicates within the alveolar macrophages within human lung tissues, generally leading to a chronic and often fatal pneumonia.18 Studies have shown that legionaminic acid appears to be important in the virulence capabilities of L. pneumophila as it is believed to be the LPS of serotype 1 that is the key determinant for the development of the disease.18 Thus far legionaminic acid and its naturally occurring derivatives have been isolated from other pathogenic species, including Campylobacter,19,20 Pseudomonas,21–23 Vibrio,22,24–27 Acinetobacter,22,28 Escherichia29–31 and Salmonella32 (Table 1). Most notably Campylobacter strains which are gastrointestinal pathogens in numerous mammalian species heavily decorate their flagellin via O-linked serine and threonine residues with this important carbohydrate moiety and also utilise pseudaminic acid pathways in the same manner.13,19,33,34
To date, as with the pseudaminic acids numerous naturally occurring derivatives of legionaminic acid have been identified, possessing various N-substitutions at C-5 and C-7 (Table 1) and numerous linkages to other sugars. Interestingly, there have been two natural structurally distinct isomers of legionaminic acid (4) identified to date.22 The 4-epi (5) and 8-epi (6) isomers of legionaminic acid (Fig. 4) have been fully characterised and isolated from a number of important Gram-negative pathogens (Table 1).22 The 8-epi-legionaminic acid (6) isomer which has an L-glycero-D-galacto configuration has been identified in a number of Gram-negative bacterial strains (Table 1).21,30–32,35–38 The 4-epi-legionaminic acid (5) isomer having a D-glycero-D-talo configuration has so far been isolated from L. pneumophila strains,22,39,40 and only one other species, the marine anaerobic Proteobacteria Shewanella japonica41 (Table 1).
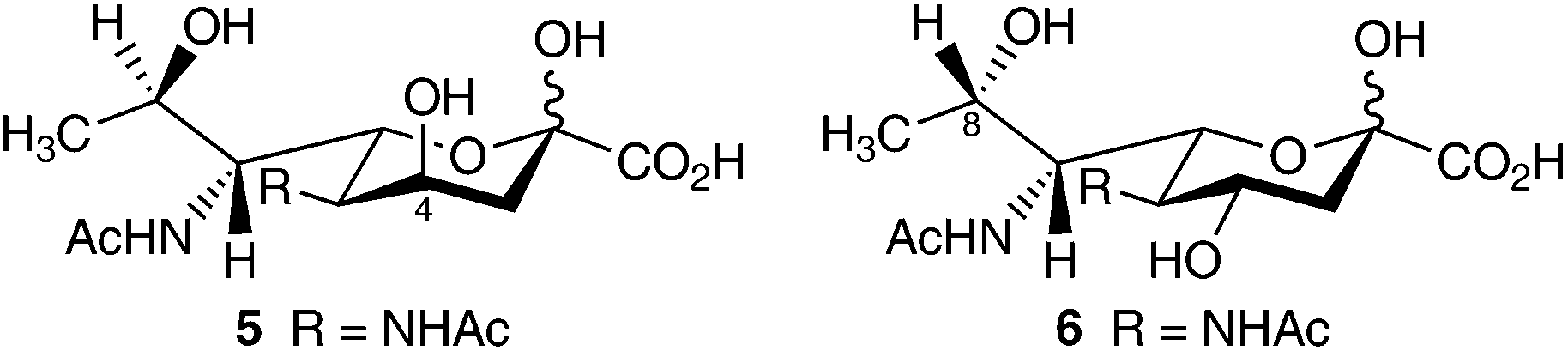 | ||
| Fig. 4 The D-glycero-D-talo isomer 4-epi-legionaminic acid (5), and the L-glycero-D-galacto isomer 8-epi-legionaminic acid (6). | ||
More recently Kandiba et al.42 have identified a 5-N-formyl derivative of legionaminic acid as a terminal residue within the N-glycosylated VP4-derived (structural protein) pentasaccharide glycan of the haloarchaeal pleomorphic virus (HRPV-1).42 This discovery is the first known occurrence of this typically O-linked sugar being isolated from an N-linked glycoprotein and also the first time a legionaminic acid derivative has been identified in an archaeal-derived glycan structure. This group has subsequently analysed the putative nonulosonic acid biosynthetic pathway in Archaea.6 This work highlights the ever increasing knowledge that the legionaminic acids are widespread within nature and have important metabolic and virulent capabilities.
Biosynthesis of 5,7-diamino-3,5,7,9-tetradeoxy-nonulosonates
Biosynthetically both pseudaminic acid (3) and legionaminic acid (4) are synthesised from a nucleotide activated N-acetyl-glucosamine. The complete biosynthesis of pseudaminic acid has been characterised within Campylobacter jejuni and Helicobacter pylori (Scheme 1).20,43–45 Within these bacteria pseudaminic acid is generated from UDP-N-acetylglucosamine (7) in an efficient five step pathway (Scheme 1).20,43–45 The first step in the pathway is catalysed by PseB an enzyme that has a dual function as a NAD(P)-dependant dehydratase and C-5 epimerase, converting UDP-α-D-GlcNAc (7) to UDP-2-acetamido-2,6-dideoxy-β-D-arabino-hexos-4-ulose, 8.45–48 Compound 8 is converted by PseC into UDP-4-amino-4,6-dideoxy-β-L-AltNAc 9 which, as can be seen in Scheme 1, rearranges the entire molecule converting 8 from a D-sugar to an L-sugar.44,45,49 The intermediate 9 is then shuttled through a further two enzyme catalysed reactions, involving PseH and PseG,50,51 being converted into 2,4-diacetamindo-2,4,6-trideoxy-β-L-altropyranose 10. Compound 10 is the substrate for PseI, the enzyme that converts it into pseudaminic acid (3) via an aldol condensation with phosphoenolpyruvate (PEP) (Scheme 1).45,46,52 In order to be incorporated into O-antigens, or utilised in O-linked protein glycosylation pathways, pseudaminic acid (3) must be activated as its CMP-linked derivative.45,46 This reaction is catalysed by PseF, a CMP-pseudaminic acid synthetase, which utilises CTP, producing CMP-pseudaminic acid (11) and pyrophosphate (Scheme 1).45,46 | ||
| Scheme 1 Biosynthetic pathways of pseudaminic acid (3) and legionaminic acid (4). | ||
The characterisation of this pathway was made possible largely by the fact that within nature all nonulosonates such as pseudaminic acid (3) and sialic acids are synthesised via enzymes that have a highly conserved phylogenetic ancestry.3,5 To date genetic studies have shown that all bacteria that encode for nonulosonic acids share a high degree of common genetic homology in regards to the enzymes they produce in order to create the varying 9 carbon sugars they utilise within their life cycles.3,44
Many of the enzymes involved in the biosynthesis of pseudaminic acid (3) have now been extensively studied and characterised. Interestingly, PseB (first enzyme in the pathway) has been found to be inhibited at a micromolar level by CMP-pseudaminic acid (11) (Ki(app) = 18.7μM). This suggests a possible negative feedback mechanism operating within the pathway controlling the forward direction of the biosynthesis, since even high concentrations of the natural substrate UDP-GlcNAc (7) were unable to competitively override the inhibition by 11.45,46 PseG the fourth enzyme in the biosynthetic pathway was fully characterised in 2009 (ref. 51) via X-ray crystallography with both the apo-form and with a UDP-product bound to the active site being published at 1.8 Å resolution.51 This data provided the proof that PseG was indeed a UDP-hydrolase enzyme which belongs to a superfamily of inverting glycosyltransferases known as the metal-independent GT-B family (Family 28).50,51 However, even though PseG belonged to this glycosyltransferase superfamily, Liu and Tanner50 ruled out the possibility that PseG could be the missing CMP-pseudaminyltransferase that would be responsible for incorporating CMP-activated pseudaminic acid (11) into glycan structures.50
Synonymously the legionaminic acid biosynthetic pathway, which in the case of C. jejuni is an alternate pathway that operates at the same time as the pseudaminic acid pathway, has also been fully characterised.53 In C. jejuni legionaminic acid (4) is generated from a nucleotide activated N-acetylglucosamine derivative, although the starting molecule is GDP-rather than UDP-N-acetylglucosamine (Scheme 1).53 The first step in the biosynthesis of legionaminic acid (4) is catalysed by LegB an NAD-dependent 4,6-dehydratase, which converts GDP-N-acetyl-glucosamine (12) to GDP-2-acetamido-2,6-dideoxy-α-D-xylo-hexos-4-ulose (13).53 LegC a PLP-dependent aminotransferase then catalysis the next step converting 13 into GDP-4-amino-4,6-dideoxy-α-D-GlcNAc, which is further processed by an N-acetyltransferase (LegH) to give GDP-2,4-diacetamido-2,4,6-trideoxy-α-D-glucopyranose (14). Intermediate 14 is then converted into 2,4-diacetamido-2,4,6-trideoxy-D-mannopyranose (15) via the catalysis of an enzyme known as LegG, which acts as an NDP-sugar hydrolase and 2-epimerase.53 Compound 15 is a 6-deoxy-mannose derivative or D-rhamnose sugar which is then transformed into legionaminic acid (4) by condensation with a 3-carbon unit (PEP) by the enzyme LegI a legionaminic acid synthase (Scheme 1).53 As within all nonulosonate biosynthetic pathways the resulting 9 carbon α-keto sugar must be nucleotide activated before being able to be incorporated into glycans.3,44 In the case of legionaminic acid it is CMP-activated by the enzyme LegF a CMP-legionaminic acid synthetase producing CMP-legionaminic acid (16) (Scheme 1).53
Remarkably, despite a detailed knowledge of the role of key enzymes in the biosynthesis of nucleotide-activated CMP-pseudaminic acid (11) and CMP-legionaminic acid (16), along with the isolation and characterisation of numerous naturally occurring derivatives (Table 1), the isolation and characterization of possible glycosyltransferase(s) responsible for the incorporation of these CMP-linked nonulosonates into glycan structures remains elusive. However, recently it has been reported that the glycosyltransferase responsible for the transfer of CMP-activated pseudaminic acid onto the flagellin of Aeromonas caviae has been found.54 Whilst these authors did not show direct evidence that the motility-associated factor maf gene provides the enzyme responsible for adding pseudaminic acid onto the flagellin, their studies provide strong evidence that maf1 is indeed a pseudaminyl transferase.54 This is the first report describing the identification of a probable pseudaminyl transferase, and will perhaps provide important information for further studies aimed at identifying this glycosyltransferase enzyme in other bacteria.
The task of finding all enzymes involved in the biosynthesis and incorporation of these nonulosonic acids into bacterial glycans is clearly an area of interest. Shoenhofen et al.45 remarked that after the full characterisation of the pseudaminic acid biosynthetic pathway, the entire pathway could now be screened for inhibitors in vitro, rather than just screening specific enzymes. This screening could potentially be accomplished by combining all five of the biosynthetic enzymes, UDP-GlcNAc (7), and potential inhibitors, thus screening for phosphate release, which only occurs during the last biosynthetic step.45
Biological significance of 5,7-diamino-3,5,7,9-tetradeoxy-nonulosonates in Gram-negative bacteria
Opportunistic bacterial species achieve their pathogenicity via virulence factors which are generally proteins or molecules that are associated with key structural components of the bacterium.13,55 Whilst the outer membrane is the major component contributing to the virulence of all pathogenic Gram-negative bacteria, individual bacterial species also contain other components that contribute to their virulence.14 Structures such as flagellin and pili provide a bacterium motility, which aids in colonisation, invasive and evasive mechanisms.13,56 Motility is a key factor in the adaptation and virulence capacity of many bacterial pathogens that colonize mucosal surfaces. For example the gastroenteritis causing Campylobacter and Helicobacter species demonstrate highly efficient flagellin motility under conditions of elevated viscosity such as that found in the gastric lumen.13 Studies have shown that although flagellin are a major structural determinate for bacterial virulence, it is in fact the glycosylated ends of these flagellin that facilitate bacterial host interactions and therefore largely confer immune responses.12,13Within Campylobacter and Helicobacter strains the carbohydrate modifications of their flagellin glycans are largely based on pseudaminic acid (3) and legionaminic acid (4).12,13,16 The major modifications identified on flagellin from C. jejuni are O-linked pseudaminic acid, or changes to the N-acyl groups (see examples in Table 1). Structural investigations of flagellin from isogenic mutants of Campylobacter and Helicobacter strains13,16,34,57–59 have provided sufficient proof that pseudaminic acid (3) is an essential glycosylation molecule involved in flagellin assembly and function, and therefore these carbohydrates have an important role in the pathogenicity of these bacteria.34,56 Indeed knock out mutants of key pseudaminic acid biosynthetic genes in several strains of Campylobacter are non-motile and accumulate intracellular flagellin subunits of reduced molecular mass, which relates to the bacterium becoming non-viable pathogens.34,59 Analysis of the flagellin from these knock out mutants by mass spectroscopy confirmed the lack of any glycosylation, providing specific proof of a functional role for the glycosylation process in flagellin assembly.34,59 Flagellin glycosylation with pseudaminic acid in Aeromonas hydrophila AH-3 has also been shown to be essential for motility.60
Further studies have shown that pseudaminic acid (3) is a potential substrate and signalling molecule for many proteins essential for flagellin assembly not only in C. jejuni but also H. pylori, with nonulosonates accounting for up to 10% of the total mass of flagellin glycoprotein in certain strains.13,16,34,57–59 Flagellin from C. jejuni and H. pylori strains have been shown to contain 19 amino acids (Ser/Thr residues) that are O-linked to the monosaccharides pseudaminic acid (3) and legionaminic acid (4) and several of their analogues, dependent on the strain.13,16,34,57–59 Discoveries such as this have produced great interest, since directly O-linked pseudaminic acid was the first report of an acidic monosaccharide being linked to a protein within a bacterium.61
Although the exact biological function of both pseudaminic acid (3) and legionaminic acid (4) and their naturally occurring derivatives is not fully understood,34,54,56 it is known that the presence of these post-translational glycolytic modifications of flagellin subunits is absolutely necessary for the assembly of a functional flagellin. Since motility is required for colonisation, the presence of these nonulosonates can be considered a virulence factor in these bacteria.13,16,34,57–59 It has also been noted that due to the structural similarity of these bacterial nonulosonic acids to that of eukaryote sialic acids, this may contribute to bacterial virulence by dampening the immune response to invading bacterium that utilise these important carbohydrates.9,16 Furthermore the relative structural similarities these sugars (especially the legionaminic acids) have with the sialic acids may also enable a potential interaction with host sialic acid-specific lectins, which could explain their possible influence in adhesion and infection.53
Conclusions
To date, the exact biological significance of pseudaminic and legionaminic acid along with their naturally occurring derivatives remains to be fully determined. However, the presence of these complex sugars as components of cell surface-associated glycans in numerous clinically important pathogenic Gram-negative bacteria suggests a potential role in host–bacterial interactions and therefore virulence. The increasing discovery of these carbohydrates within important bacterial structural elements demonstrates that these unique higher sugars are more common to bacteria than previously believed. Moreover, at present due to the importance of their role in the pathogenic capabilities of many clinically important drug-resistant Gram-negative bacteria, there is an opportunity for exploring possible new therapeutic targets that focus on proteins or enzymes associated with the biosynthesis or incorporation of these complex sugars into bacterial glycans. It is expected that the coming decade will see a significant increase in research activity on this area, especially if new improved synthetic strategies emerge that allow the efficient synthesis of pseudaminic acid or legionaminic acid analogues.Notes and references
- C. Traving and R. Schauer, Cell. Mol. Life Sci., 1998, 54, 1330–1349 CrossRef CAS.
- A. Varki, Glycobiology, 1992, 2, 25–40 CrossRef CAS.
- T. Angata and A. Varki, Chem. Rev., 2002, 102, 439–469 CrossRef CAS PubMed.
- P.-H. Wang, J. Cancer Mol., 2005, 1, 73–81 CAS.
- X. Chen and A. Varki, ACS Chem. Biol., 2010, 5, 163–176 CrossRef CAS PubMed.
- L. Kandiba and J. Eichler, FEMS Microbiol. Lett., 2013, 345, 110–120 CrossRef CAS PubMed.
- A. L. Lewis, N. Desa, E. E. Hansen, Y. A. Knirel, J. I. Gordon, P. Gagneux, V. Nizet and A. Varki, Proc. Natl. Acad. Sci. U. S. A., 2009, 106, 13552–13557 CrossRef CAS PubMed.
- Y. A. Knirel, E. V. Vinogradov, V. L. L'vov, N. A. Kocharova, A. S. Shashkov, B. A. Dmitriev and N. K. Kochetkov, Carbohydr. Res., 1984, 133, C5–C8 CrossRef CAS.
- Y. A. Knirel, A. S. Shashkov, Y. E. Tsvetkov, P.-E. Jansson and U. Zähringer, Adv. Carbohydr. Chem. Biochem., 2003, 58, 371–417 CrossRef CAS.
- E. Kiss, B. L. Reuhs, J. S. Kim, A. Kereszt, G. Petrovics, P. Putnoky, I. Dusha, R. W. Carlson and A. Kondorosi, J. Bacteriol., 1997, 179, 2132–2140 CAS.
- J. Horzempa1, C. R. Dean, J. B. Goldberg and P. Castric, J. Bacteriol., 2006, 188, 4244–4252 CrossRef PubMed.
- M. Schirm, E. C. Soo, A. J. Aubry, J. Austin, P. Thibault and S. M. Logan, Mol. Microbiol., 2003, 48, 1579–1592 CrossRef CAS.
- P. Thibault, S. M. Logan, J. F. Kelly, J.-R. Brisson, C. P. Ewing, T. J. Trust and P. Guerry, J. Biol. Chem., 2001, 276, 34862–34870 CrossRef CAS PubMed.
- D. Mokady, U. Gophna and E. Z. Ron, Int. J. Microbiol., 2005, 295, 455–462 CAS.
- P. G. Hitchen and A. Dell, Microbiology, 2006, 152, 1575–1580 CrossRef CAS PubMed.
- Y. A. Knirel, S. D. Shevelev and A. V. Perepelov, Mendeleev Commun., 2011, 21, 173–182 CrossRef CAS PubMed.
- Y. A. Knirel, E. T. Rietschel, R. Marre and U. Zähringer, Eur. J. Biochem., 1994, 221, 239–245 CrossRef CAS.
- C. Cazalet, S. Jarraud, Y. Ghavi-Helm, F. Kunst, P. Glaser, J. Etienne and C. Buchrieser, Genome Res., 2008, 18, 431–441 CrossRef CAS PubMed.
- S. M. Logan, J. P. M. Hui, E. Vinogradov, A. J. Aubry, J. E. Melanson, J. F. Kelly, H. Nothaft and E. C. Soo, FEBS J., 2009, 276, 1014–1023 CrossRef CAS PubMed.
- D. J. McNally, A. J. Aubry, J. P. M. Hui, N. H. Khieu, D. Whitfield, C. P. Ewing, P. Guerry, J.-R. Brisson, S. M. Logan and E. C. Soo, J. Biol. Chem., 2007, 282, 14463–14475 CrossRef CAS PubMed.
- Y. A. Knirel, E. V. Vinogradov, A. S. Shashkov, B. A. Dmitriev, N. K. Kochetkov, E. S. Stanislavsky and G. M. Mashilova, Eur. J. Biochem., 1986, 157, 129–138 CrossRef CAS.
- Y. E. Tsvetkov, A. S. Shashkov, Y. A. Knirel and U. Zähringer, Carbohydr. Res., 2001, 331, 233–237 CrossRef CAS.
- Y. A. Knirel, J. H. Helbig and U. Zähringer, Carbohydr. Res., 1996, 283, 129–139 CrossRef CAS.
- E. L. Nazarenko, A. S. Shashkov, Y. A. Knirel, E. P. Ivanova and Y. S. Ovodov, Bioorg. Khim., 1990, 16, 1426–1429 CAS.
- P. Edebrink, P.-E. Jansson, J. Bøgwald and J. Hoffman, Carbohydr. Res., 1996, 287, 225–245 CrossRef CAS.
- N. Hashii, Y. Isshiki, T. Iguchi, K. Hisatsune and S. Kondo, Carbohydr. Res., 2003, 338, 1055–1062 CrossRef CAS.
- N. Hashii, Y. Isshiki, T. Iguchi and S. Kondo, Carbohydr. Res., 2003, 338, 1063–1071 CrossRef CAS.
- S. R. Haseley and S. G. Wilkinson, Eur. J. Biochem., 1997, 250, 617–623 CAS.
- X. Li, A. V. Perepelov, Q. Wang, S. N. Senchenkova, B. Liu, S. D. Shevelev, X. Guo, A. S. Shashkov, W. Chen, L. Wang and Y. A. Knirel, Carbohydr. Res., 2010, 345, 1581–1587 CrossRef CAS PubMed.
- Y. A. Knirel, S. N. Senchenkova, A. S. Shashkov, S. D. Shevelev, A. V. Perepelov, L. Bin, L. Feng and L. Wang, Adv. Sci. Lett., 2009, 2, 384–387 CrossRef CAS PubMed.
- A. V. Perepelov, B. Liu, S. N. Senchenkova, A. S. Shashkov, S. D. Shevelev, L. Feng, L. Wang and Y. A. Knirel, Biochemistry, 2010, 75, 19–24 CrossRef CAS.
- E. V. Vinogradov, A. S. Shashkov, Y. A. Knirel, N. K. Kochetkov, J. Dabrowski, H. Grosskurth, E. S. Stanislavsky and E. V. Kholodkova, Carbohydr. Res., 1992, 231, 1–11 CrossRef CAS.
- M. Schirm, I. C. Schoenhofen, S. M. Logan, K. C. Waldron and P. Thibault, Anal. Chem., 2005, 77, 7774–7782 CrossRef CAS PubMed.
- S. M. Logan, J. F. Kelly, P. Thibault, C. P. Ewing and P. Guerry, Mol. Microbiol., 2002, 46, 587–597 CrossRef CAS.
- L. M. Beynon, J. C. Richards and M. B. Perry, Carbohydr. Res., 1994, 256, 303–317 CrossRef CAS.
- M. Kilcoyne, A. S. Shashkov, S. N. Senchenkova, Y. A. Knirel, E. V. Vinogradov, J. Radziejewska-Lebrecht, R. Galimska-Stypa and A. V. Savage, Carbohydr. Res., 2002, 337, 1697–1702 CrossRef CAS.
- A. S. Shashkov, V. I. Torgov, E. L. Nazarenko, V. A. Zubkov, N. M. Gorshkova, R. P. Gorshkova and G. Widmalm, Carbohydr. Res., 2002, 337, 1119–1127 CrossRef CAS.
- A. S. Shashkov, N. A. Kocharova, G. V. Zatonsky, A. Błaszczyk, Y. A. Knirel and A. Rozalski, Carbohydr. Res., 2007, 342, 653–658 CrossRef CAS PubMed.
- Y. A. Knirel, H. Moll, J. H. Helbig and U. Zähringer, Carbohydr. Res., 1997, 304, 77–79 CrossRef CAS.
- Y. A. Knirel, S. N. Senchenkova, N. A. Kocharova, A. S. Shashkov, J. H. Helbig and U. Zähringer, Biochemistry, 2001, 66, 1035–1041 CrossRef CAS.
- E. L. Nazarenko1, A. V. Perepelov, L. S. Shevchenko1, E. D. Daeva, E. P. Ivanova1, A. S. Shashkov and G. Widmalm, Biochemistry, 2011, 76, 791–796 CrossRef PubMed.
- L. Kandiba, O. Aitio, J. Helin, Z. Guan, P. Permi, D. H. Bamford, J. Eichler and E. Roine, Mol. Microbiol., 2012, 84, 578–593 CrossRef CAS PubMed.
- D. J. McNally, J. P. M. Hui, A. J. Aubry, K. K. K. Mui, P. Guerry, J.-R. Brisson, S. M. Logan and E. C. Soo, J. Biol. Chem., 2006, 281, 18489–18498 CrossRef CAS PubMed.
- I. C. Schoenhofen, D. J. McNally, E. Vinogradov, D. Whitfield, N. M. Young, S. Dick, W. W. Wakarchuk, J.-R. Brisson and S. M. Logan, J. Biol. Chem., 2006, 281, 723–732 CrossRef CAS PubMed.
- I. C. Schoenhofen, D. J. McNally, J.-R. Brisson and S. M. Logan, Glycobiology, 2006, 16, 8C–14C CrossRef CAS PubMed.
- D. J. McNally, I. C. Schoenhofen, R. S. Houliston, N. H. Khieu, D. M. Whitfield, S. M. Logan, H. C. Jarrell and J.-R. Brisson, ChemMedChem, 2008, 3, 55–59 CrossRef CAS PubMed.
- P. Guerry, C. P. Ewing, M. Schirm, M. Lorenzo, J. Kelly, D. Pattarini, G. Majam, P. Thibault and S. M. Logan, Mol. Microbiol., 2006, 60, 299–311 CrossRef CAS PubMed.
- J. P. Morrison, I. C. Schoenhofen and M. E. Tanner, Bioorg. Chem., 2008, 36, 312–320 CrossRef CAS PubMed.
- I. C. Schoenhofen, V. V. Lunin, J.-P. Julien, Y. Li, E. Ajamian, A. Matte, M. Cygler, J.-R. Brisson, A. Aubry, S. M. Logan, S. Bhatia, W. W. Wakarchuk and N. M. Young, J. Biol. Chem., 2006, 281, 8907–8916 CrossRef CAS PubMed.
- F. Liu and M. E. Tanner, J. Biol. Chem., 2006, 281, 20902–20909 CrossRef CAS PubMed.
- E. S. Rangarajan, A. Proteau, Q. Cui, S. M. Logan, Z. Potetinova, D. Whitfield, E. O. Purisima, M. Cygler, A. Matte, T. Sulea and I. C. Schoenhofen, J. Biol. Chem., 2009, 284, 20989–21000 CrossRef CAS PubMed.
- W. K. Chou, S. Dick, W. W. Wakarchuk and M. E. Tanner, J. Biol. Chem., 2005, 280, 35922–35928 CrossRef CAS PubMed.
- I. C. Schoenhofen, E. Vinogradov, D. M. Whitfield, J.-R. Brisson and S. M. Logan, Glycobiology, 2009, 19, 715–725 CrossRef CAS PubMed.
- J. L. Parker, M. J. Day-Williams, J. M. Tomas, G. P. Stafford and J. G. Shaw, MicrobiologyOpen, 2012, 1, 149–160 CrossRef CAS PubMed.
- M. Zilbauer, N. Dorrell, B. W. Wren and M. Bajaj-Elliott, Trans. R. Soc. Trop. Med. Hyg., 2008, 102, 123–129 CrossRef CAS PubMed.
- S. M. Logan, Microbiology, 2006, 152, 1249–1262 CrossRef CAS PubMed.
- C. Creuzenet, FEBS Lett., 2004, 559, 136–140 CrossRef CAS.
- R. K. Obhi and C. Creuzenet, J. Biol. Chem., 2005, 280, 20902–20908 CrossRef CAS PubMed.
- S. Goon, J. F. Kelly, S. M. Logan, C. P. Ewing and P. Guerry, Mol. Microbiol., 2003, 50, 659–671 CrossRef CAS.
- M. Wilhelms, K. M. Fulton, S. M. Twine, J. M. Tomás and S. Merino, J. Biol. Chem., 2012, 287, 27851–27862 CrossRef CAS PubMed.
- R. G. Spiro, Glycobiology, 2002, 12, 43R–56R CrossRef CAS PubMed.
- M. Staaf, A. Weintraub and G. Widmalm, Eur. J. Biochem., 1999, 263, 656–661 CrossRef CAS.
- A. N. Kondakova, A. V. Perepelov, B. Bartodziejska, A. S. Shashkov, S. N. Senchenkova, M. Wykrota, Y. A. Knirel and A. Rozalski, Carbohydr. Res., 2001, 333, 241–249 CrossRef CAS.
- A. V. Perepelov, A. S. Shashkov, V. I. Torgov, E. L. Nazarenko, R. P. Gorshkova, E. P. Ivanova, N. M. Gorshkovac and G. Widmalma, Carbohydr. Res., 2005, 340, 69–74 CrossRef CAS PubMed.
- A. J.-L. Le Quéré, W. J. Deakin, C. Schmeisser, R. W. Carlson, W. R. Streit, W. J. Broughton and L. S. Forsberg, J. Biol. Chem., 2006, 281, 28981–28992 CrossRef PubMed.
- L. A. Sharypova, G. Chataigné, N. Fraysse, A. Becker and V. Poinsot, Glycobiology, 2006, 16, 1181–1193 CrossRef CAS PubMed.
- M. G. Müller, L. S. Forsberg and D. H. Keating, J. Bacteriol., 2009, 191, 6988–7000 CrossRef PubMed.
- A. V. Perepelov, A. S. Shashkov, S. V. Tomshich, N. A. Komandrovab and O. I. Nedashkovskaya, Carbohydr. Res., 2007, 342, 1378–1381 CrossRef CAS PubMed.
- E. Vinogradov, C. Wilde, E. M. Anderson, A. Nakhamchik, J. S. Lam and D. A. Rowe-Magnus, Carbohydr. Res., 2009, 344, 484–490 CrossRef CAS PubMed.
- E. Vinogradov, M. Frimmelova and R. Toman, Carbohydr. Res., 2013, 378, 108–113 CrossRef CAS PubMed.
- Y. A. Knirel, E. V. Vinogradov, A. S. Shashkov, N. K. Kochetkov, V. L. L'vov and B. A. Dmitriev, Carbohydr. Res., 1985, 141, C1–C3 CrossRef CAS.
- V. L. L'vov, A. S. Shashkov and B. A. Dmitriev, Bioorg. Khim., 1987, 13, 223–233 CAS.
- A. M. Gil-Serrano, M. A. Rodríguez-Carvajal, P. Tejero-Mateo, J. L. Espartero, M. Menendez, J. Corzo, J. E. Ruiz-Sainz and A. M. Buendía-Clavería, Biochem. J., 1999, 342, 527–535 CrossRef CAS.
- A. S. Shashkov, E. M. Tul'skaya, G. M. Streshinskaya, S. N. Senchenkova, A. N. Avtukh and L. I. Evtushenko, Carbohydr. Res., 2009, 344, 2255–2262 CrossRef CAS PubMed.
- E. M. Tul'skaya, A. S. Shashkov, G. M. Streshinskaya, S. N. Senchenkova, N. V. Potekhina, Y. I. Kozlova and L. I. Evtushenko, Biochemistry, 2011, 76, 736–744 CrossRef CAS PubMed.
- A. S. Shashkov, G. M. Streshinskaya, Y. I. Kozlova, E. M. Tul'skaya, S. N. Senchenkova, N. P. Arbatskii, O. V. Bueva and L. I. Evtushenko, Biochemistry, 2012, 77, 511–517 CrossRef CAS PubMed.
- Y. A. Knirel, N. A. Kocharova, A. S. Shashkov and N. K. Kochetkov, Carbohydr. Res., 1986, 145, C1–C4 CrossRef CAS.
- Y. A. Knirel, N. A. Kocharova, A. S. Shashkov, B. A. Dmitriev, N. K. Kochetkov, E. S. Stanislavsky and G. M. Mashilova, Eur. J. Biochem., 1987, 163, 639–652 CrossRef CAS.
- J. Muldoon, A. S. Shashkov, S. N. Senchenkova, S. V. Tomshich, N. A. Komandrova, L. A. Romanenko, Y. A. Knirel and A. V. Savage, Carbohydr. Res., 2001, 330, 231–239 CrossRef CAS.
- L. Kenne, B. Lindberg, E. Schweda, B. Gustafsson and T. Holme, Carbohydr. Res., 1988, 180, 285–294 CrossRef CAS.
- S. M. Twine, C. J. Paul, E. Vinogradov, D. J. McNally, J.-R. Brisson, J. A. Mullen, D. R. McMullin, H. C. Jarrell, J. W. Austin, J. F. Kelly and S. M. Logan, FEBS J., 2008, 275, 4428–4444 CrossRef CAS PubMed.
Footnote |
| † Current address: School of Pharmacy, Griffith University Gold Coast, Parklands Drive, Southport, Queensland, 4222, Australia. E-mail: E-mail: m.zunk@griffith.edu.au, Fax: +61 7 5552 8804, Tel: +61 7 5552 8344 |
| This journal is © The Royal Society of Chemistry 2014 |