Microfluidic evaluation of red cells collected and stored in modified processing solutions used in blood banking
Yimeng
Wang
,
Adam
Giebink
and
Dana M.
Spence
*
Department of Chemistry, Michigan State University, East Lansing, MI 48824, USA. E-mail: dspence@chemistry.msu.edu
First published on 14th November 2013
Abstract
The most recent American Association of Blood Banks survey found that 40![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 units of blood are required daily for general medicine, hematology/oncology, surgery, and for accident and trauma victims. While blood transfusions are an extremely important component of critical healthcare, complications associated with transfusion of blood components still exist. It is well-established that the red blood cell (RBC) undergoes many physical and chemical changes during storage. Increased oxidative stress, formation of advanced glycation endproducts, and microparticle formation are all known to occur during RBC storage. Furthermore, it is also known that patients who receive a transfusion have reduced levels of available nitric oxide (NO), a major determinant in blood flow. However, the origin of this reduced NO bioavailability is not completely understood. Here, we show that a simple modification to the glucose concentration in the solutions used to process whole blood for subsequent RBC storage results in a remarkable change in the ability of these cells to stimulate NO. In a controlled in vitro microflow system, we discovered that storage of RBCs in normoglycemic versions of standard storage solutions resulted in RBC-derived ATP release values 4 weeks into storage that were significantly greater than day 1 release values for those RBCs stored in conventional solutions. During the same storage duration, microfluidic technologies enabled measurements of endothelium-derived NO that were stimulated by the ATP release from the stored RBCs. In comparison to currently accepted processing solutions, the NO production increased by more than 25% in the presence of the RBCs stored in the normoglycemic storage solutions. Control experiments using inhibitors of ATP release from the RBCs, or ATP binding to the endothelium, strongly suggest that the increased NO production by the endothelium is directly related to the ability of the stored RBCs to release ATP. We anticipate these findings to represent a starting point in controlling glucose levels in solutions used for blood component storage, especially considering that current solutions contain glucose at levels that are nearly 20-fold greater than blood glucose levels of a healthy human, and even 10-fold greater than levels found in diabetic bloodstreams.
000 units of blood are required daily for general medicine, hematology/oncology, surgery, and for accident and trauma victims. While blood transfusions are an extremely important component of critical healthcare, complications associated with transfusion of blood components still exist. It is well-established that the red blood cell (RBC) undergoes many physical and chemical changes during storage. Increased oxidative stress, formation of advanced glycation endproducts, and microparticle formation are all known to occur during RBC storage. Furthermore, it is also known that patients who receive a transfusion have reduced levels of available nitric oxide (NO), a major determinant in blood flow. However, the origin of this reduced NO bioavailability is not completely understood. Here, we show that a simple modification to the glucose concentration in the solutions used to process whole blood for subsequent RBC storage results in a remarkable change in the ability of these cells to stimulate NO. In a controlled in vitro microflow system, we discovered that storage of RBCs in normoglycemic versions of standard storage solutions resulted in RBC-derived ATP release values 4 weeks into storage that were significantly greater than day 1 release values for those RBCs stored in conventional solutions. During the same storage duration, microfluidic technologies enabled measurements of endothelium-derived NO that were stimulated by the ATP release from the stored RBCs. In comparison to currently accepted processing solutions, the NO production increased by more than 25% in the presence of the RBCs stored in the normoglycemic storage solutions. Control experiments using inhibitors of ATP release from the RBCs, or ATP binding to the endothelium, strongly suggest that the increased NO production by the endothelium is directly related to the ability of the stored RBCs to release ATP. We anticipate these findings to represent a starting point in controlling glucose levels in solutions used for blood component storage, especially considering that current solutions contain glucose at levels that are nearly 20-fold greater than blood glucose levels of a healthy human, and even 10-fold greater than levels found in diabetic bloodstreams.
Insight, innovation, integrationThe work presented here provides biological insight by demonstrating that modification to the solutions used to process (collect and store) red blood cells (RBCs) for storage prior to transfusion leads to an improvement in these cells’ ability to release ATP and, subsequently, stimulate nitric oxide production in endothelial cells. Efficacy was confirmed with the technological innovation of using a microfluidic device and other microflow techniques to measure ATP release from the stored cells in a flow-based environment. This integration was necessary to prove that insufficient nitric oxide bioavailability (INOBA) after transfusion may be due to reduced cell-to cell-communication between RBCs and the endothelium and this reduction in communication is due to the hyperglycemic conditions of current storage protocols. |
The World Health Organization (WHO) reports that over 100 million units of blood are collected globally each year and are most commonly used in cardiovascular surgery, transplant surgery, massive trauma, and therapy for solid and haematological malignancies in high income countries. In low- and middle-income countries it is used more often for management of pregnancy-related complications, childhood malaria complicated by severe anaemia, and trauma-related injuries.1
In the United States alone, it is estimated by the American Association of Blood Banks that 40000 units of blood are required daily for general medicine, hematology/oncology, surgery, and for accident and trauma victims. While the most recent National Blood Collection and Utilization Survey (NBCUS) suggests that a “shortage” of stored units of red blood cells (RBCs) was not necessarily an issue in the US during the period covered by the survey, the complications associated with receiving a transfusion remain one of the critical barriers to progress in the field.2 Thus, while blood transfusions are an extremely important component of critical healthcare, complications associated with transfusion of blood components still exist.3,4
Recently, it has been reported that people who receive a transfusion suffer from insufficient nitric oxide bioavailability (INOBA), which has a profound effect on blood flow.5 The concept of INOBA associated with the transfusion of stored RBCs is of particular interest to our group and others due to a wealth of literature describing the RBC as a determinant of blood flow.6–8 Specifically, the RBC has been shown to release nitric oxide (NO) through different mechanisms when exposed to hypoxia.9,10 In addition, the RBC is also capable of releasing adenosine-5′-triphosphate (ATP) when exposed to hypoxia,11,12 flow-induced deformation,13,14 and various molecular stimuli.15,16 Importantly, this RBC-derived ATP has been shown to stimulate NO production in various cell types such as the vascular endothelium,17 as well as platelets.18
However, the origin of the INOBA is not completely understood, and when stored RBCs are transfused, it is not clear if the INOBA is due to a reduction in RBC-derived NO, or RBC-stimulated NO. Differentiating these two sources using an in vivo model would be very difficult and complex, if not impossible. Unfortunately, even if an in vitro tool or device that mimics in vivo cellular properties and cell-to-cell communication could be developed, the origins of the RBC properties that may be leading to the INOBA would still not be completely understood.
Investigators in the field of transfusion medicine are well aware of various properties that may affect the stored RBC. Interestingly, many of these stored RBC properties (e.g., oxidative stress19 and advanced glycation endproducts (AGEs)20), collectively known as the red cell storage lesion,21 also occur in the RBCs obtained from people with diabetes.22 It would be premature to assume that the RBC storage lesion and diabetic complications have the same origin; however, an examination of the processing solutions used to prepare RBCs for storage reveals a very interesting environment into which collection and storage occur.
Consider 450 mL of whole blood collected from a donor;23 the number of RBCs in that collection volume can be estimated by assuming that ∼40% of the collected blood is comprised of RBCs and, at a volume of ∼87 fL per RBC, one could estimate that the number of RBCs is approximately 2 × 1012 in the whole blood collection. These RBCs are collected in ∼70 mL of citrate phosphate dextrose (CPD) solution, which contains about 1.6 g of glucose (or 9.0 × 10−3 moles); in other words, these RBCs are being exposed to a solution (the CPD) that has a glucose concentration of ∼129 mM prior to collection of the whole blood. Even after the collection of the ∼450 mL of whole blood, the glucose levels are still >20 mM. After collection into the CPD, the RBCs are separated and added to 100 mL of an additive solution (AS-1) that contains glucose at a concentration of ∼111 mM. After the addition of the RBCs, the final glucose concentration is ∼40 mM. To give this value some perspective, keep in mind that a normoglycemic, healthy individual has a bloodstream glucose level between 4 and 6 mM.24 In short, the glucose concentrations in the collection and storage solutions (nearly an order of magnitude higher than a healthy individual) may adversely affect the stored RBC.
Here, using a microfluidic device, we demonstrate that the solutions used for collection of whole blood, and for storage of purified RBCs, are having a significant impact on flow-induced ATP release from stored RBCs, which in turn is affecting endothelium-derived NO production. Importantly, we also provide evidence that collection and storage in normoglycemic versions of CPD and AS-1 (labeled as CPD-N and AS-1N, respectively), followed by periodic feeding of the stored cells to maintain glucose concentrations near normoglycemic levels, results in ATP release values 4 weeks into storage that are statistically higher than day 1 values for cells processed in typical CPD–AS-1 solutions. We also demonstrate that this RBC-derived ATP is having a significant impact on endothelium-derived NO.
Results
Flow-induced ATP release from stored RBCs
To determine the stored RBC's ability to release flow-induced ATP, 2 micro-units (the small scale storage containers that were used to store RBCs used in the studies reported here) of stored RBCs were evaluated; one of these micro-units contained RBCs collected and stored in CPD and AS-1, respectively, while the other micro-unit contained RBCs collected and stored in CPD with 5.5 mM glucose (CPD-N) and AS-1 with 5.5 mM glucose (AS-1N). The process for preparing these cells is shown in Fig. 1a. The cells in these samples were first evaluated for their ability to release ATP while flowing through a microbore tubing system described previously.25 The results in Fig. 1b show that the RBCs collected and stored in the CPD-N and AS-1N released a significantly higher (p < 0.005) amount of ATP though the first 8 days (day 1, 0.312 ± 0.008 μM to 0.178 ± 0.019 μM at day 8) than those collected and stored in the CPD and AS-1 (day 1, 0.126 ± 0.002 μM to 0.070 ± 0.011 μM at day 8). | ||
| Fig. 1 In the upper sequence, CPD or CPD-N is injected into non-treated collection tubes that are under vacuum. Whole blood (∼7 mL) is then drawn into each tube; after centrifugation, the plasma and buffy layers are separated from the RBCs. Next, ∼2 mL of an additive solution (either AS-1 or AS-1N) are added to concentrated RBCs that has been collected in either CPD or CPD-N, respectively. Aliquots of 1 mL of these RBCs are pipetted into miniature PVC bags. If needed, the bags are opened weekly for periodic feeding or experimentation (a). In (b) the ATP release from the RBCs stored in both protocols is shown; while the cells stored in AS-1N (gray bars) show significantly higher ATP release than those in AS-1 (black bars) for the first 8 days (*p < 0.005), the release gradually decreased as a function of storage duration. This is possibly explained by the data in (c) where lactate measurements from the supernatant of the storage bags indicate that glycolytic metabolism had ceased in the AS-1N. A determination of glucose in these bags verified that glucose had been exhausted from the AS-1N storage solution (not shown). Lactate accumulation and glucose were present in the AS-1 solution. Data represent mean ± s.e.m. (n = 4 for all). | ||
Although the ATP release from the CPD-N–AS-1N cells on day 8 was statistically higher (p < 0.05) than the day 1 release from cells processed in CPD–AS-1, the results in Fig. 1b also show that the ATP release from the RBCs processed in CPD-N–AS-1N was also decreasing rapidly. Results from additional experiments evaluating lactate levels in the supernatant of the storage solutions, which were performed on the same day as the ATP release studies, suggested that the lactate levels in the supernatant of RBCs stored in CPD-N–AS-1N stopped accumulating around day 8 (see Fig. 1c). The decrease in lactate accumulation was verified by a quantitative determination of glucose concentration in the AS-1N supernatant, which was found to be exhausted by day 8 (data not shown). Collectively, the results shown in Fig. 1 suggest that while a normoglycemic collection and storage protocol may be beneficial for maintenance of ATP release from stored RBCs, it is not necessarily conducive for storage periods beyond 1 week.
Flow-induced ATP release from stored RBCs with glucose maintenance
In order to maintain glucose levels at ∼5 mM, a periodic glucose feeding was performed in CPD-N–AS-1N storage strategies (see Fig. 1a). As shown in Fig. 2a, the ATP release from those RBCs processed in CPD-N–AS-1N with feeding (described in detail in the Methods section) was significantly higher (0.285 ± 0.028 μM) than those processed in CPD–AS-1 (0.079 ± 0.021 μM) through 29 days (p < 0.05). In addition, RBCs collected and stored in CPD-N–AS-1N released ATP at a level on day 36 (0.178 ± 0.046 μM) of storage that was statistically equivalent (p = 0.694) to the amount of ATP from RBCs collected and stored in the CPD–AS-1 on day 1 (0.152 ± 0.042 μM). As shown in Fig. 2b, the periodic feeding of the RBCs stored in the CPD-N–AS-1N solutions enabled glycolytic processes, as evident by the continued lactate accumulation throughout the storage period. | ||
| Fig. 2 RBC-derived ATP is shown in (a) for cells processed using the CPD–AS-1 solutions (black bars) or the CPD-N–AS-1N solutions with glucose supplementation occurring weekly beginning at Day 5 of storage (gray bars). Not unlike the data shown in Fig. 1, the cells stored in AS-1N consistently release higher levels of ATP than their AS-1-stored counterparts. However, the weekly maintenance of glucose levels in the storage bags helped maintain increased release of ATP from the RBCs throughout the 29 day storage duration (*p < 0.05). In fact, the release from the AS-1N stored cells on Day 36 is statistically equivalent (p = 0.694) to those cells stored in AS-1 on Day 1. The weekly glucose supplementation resulted in lactate accumulation (b) during the entire storage period, while glucose measurements (not shown) verified that glucose concentrations in the AS-1N were maintained between 5–6 mM. Data represent mean ± s.e.m. (n = 4 for all). | ||
ATP-stimulated endothelial cells NO release on chip
The potential importance of the increased ATP release from RBCs processed in CPD-N–AS-1N, shown above in Fig. 2a, is related to its well-established role as a stimulus of NO production in various cell types. Here, a microfluidic device enabled RBC-derived ATP to diffuse to an endothelial layer and stimulate NO production. Devices similar to that shown in Fig. 3a have been previously employed by our group to investigate the cellular communication mediated by ATP release from RBCs.26 However, this is the first time that such a device has been used to investigate stored RBCs. Fig. 3b represents the molecule-mediated signaling that occurs between the two types of cells. RBCs processed with the different collection and storage solutions were pumped through the micron-scale channels; the resultant flow-induced ATP then diffuses through the pores of a membrane that separates the flowing RBC sample from a layer of immobilized endothelial cells. The NO released from the endothelial cells is measured using 4-amino-5-methylamino-2′,7′-difluorescein (DAF-FM), a fluorogenic probe for NO. The amount of NO production from endothelial cells stimulated by ATP release from flowing RBCs processed in CPD-N–AS-1N was normalized to the NO stimulated by ATP released from RBCs in CPD–AS-1 on each day. As shown in Fig. 3c, the NO production by endothelial cells in the presence of RBCs processed with CPD-N–AS-1N was significantly increased (p < 0.02) by at least 25% throughout first 22 days of storage in comparison to those RBCs processed in CPD–AS-1. | ||
| Fig. 3 A top-down view of the microfluidic device employed to determine NO production from endothelial cells with a standard plate reader is shown in (a). This device, derived from PDMS and described in more detail elsewhere, enables RBCs processed in CPD–AS or CPD-N–AS-1N to flow beneath a layer of endothelial cells. When the cells release ATP due to flow-induced deformation, they release ATP that can diffuse through a porous membrane, stimulating NO production from endothelium via purinergic receptor signaling. A fluorogenic probe facilitates the measurement of NO released by the endothelium (b). In (c) the percentage increase in NO production by the endothelial cells (>25% for the first 3 weeks of storage) is significantly higher when exposed to RBCs processed in CPD-N–AS-1N (*p < 0.02). The NO production is shown in (d) after the RBCs were exposed to GLI, an inhibitor of ATP release from the RBCs. As shown, there is no significant increase in NO production (p = 0.742), regardless of storage solution, when the stored RBCs are exposed to GLI, suggesting that the NO increase is due to endothelial cell stimulation by ATP. The data in (e), NO measurements in the presence and absence of PPADS, confirm that the signals from the endothelial cells are due to NO (p = 0.623). Data represent mean ± s.d. (n = 4 for all). | ||
Two negative control experiments were performed to ensure that this NO release was stimulated by flowing RBC-derived ATP release. First, RBCs in CPD–AS-1 and CPD-N–AS-1N were incubated with glibenclamide (GLI), an inhibitor of ATP release from RBCs.27 The data in Fig. 3d show that inhibition of the RBCs stored in the AS-1N with GLI decreases the NO production to a level that is statistically equal (p = 0.742) to those cells stored in AS-1. This strongly suggests that the increased endothelium-derived NO is not due to stimulation by ATP due to RBC lysis; if ATP due to lysis was the stimulus for the enhanced NO production, GLI inhibition would have no effect on the results.
In addition to inhibition at the RBC level, a second control was performed at the endothelial level. Specifically, endothelial cells cultured on the microfluidic device were incubated with pyridoxalphosphate-6-azophenyl-2′,4′-disulfonic acid (PPADS), which is a recognized inhibitor of the P2Y purinergic receptor for ATP.28 As shown in Fig. 3e, NO production in the presence of CPD-N–AS-1N processed RBCs, which had been significantly higher than when exposed to RBCs processed in CPD–AS-1, were unable to stimulate such increases when the endothelium was inhibited with PPADS. The data in Fig. 3e provide further evidence that the increase in NO production from the endothelial cells is directly a result of ATP stimulation. The combined data in Fig. 3 suggests that endothelium-derived levels of NO are strongly related to the ability of the stored RBC to release ATP.
Determination of intracellular ATP
Intracellular levels of ATP in the RBC are typically in the single-digit millimolar range, and the amount released is sub-micromolar.29 Furthermore, Sprague et al. have shown that intracellular levels of ATP in the RBC do not necessarily explain the decreased release of ATP from stimulated cells.30 Therefore, although ATP release from the stored RBCs was not expected to correlate with intracellular concentrations of ATP, a study was performed to determine these concentrations throughout the storage period. The data in Fig. 4 show that intracellular concentrations of ATP for all three storage protocols are nearly identical for the first week of storage. However, the RBCs stored in CPD-N–AS-1N, whose glucose levels were not maintained by periodic feeding, had intracellular ATP concentrations approach zero by week 4 of storage. The cells processed in CPD–AS-1 or CPD-N–AS-1N with glucose addition had intracellular ATP concentrations that were maintained above 0.5 mM per RBC over the course of 5 weeks of storage. As shown, the RBCs processed in the higher glucose-containing collection and storage solutions maintained slightly higher levels of ATP than those stored in the CPD-N–AS-1N. However, as shown in Fig. 1 and 2, this did not translate to higher ATP release. | ||
| Fig. 4 The intracellular concentrations of ATP are shown for RBCs processed in CPD–AS-1 (black circle), CPD-N–AS-1N with periodic glucose feeding (white circle), and CPD-N–AS-1N without periodic glucose feeding (black triangle). The ATP concentrations are highest for those stored in AS-1 and lowest for AS-1N without feeding. The intracellular ATP concentrations for the AS-1N storage with feeding is slightly lower than AS-1, suggesting that the differences in ATP release from these cells is not necessarily due to the level of ATP within the cell. Data represent mean ± s.e.m. (n = 4 for all). | ||
Determination of hemolysis
In the US, the Food and Drug Administration (FDA) recommends that hemolysis of stored RBCs should not exceed 1%, while in Europe the recommendation is lower at 0.8%.31 Due to the importance of ATP in the maintenance of cell membrane integrity, hemolysis of the RBCs processed in the CPD–AS-1 was compared to cells stored in CPD-N–AS-1N versions with and without periodic feeding with glucose. Fig. 5 shows that the percent hemolysis remained statistically less than 1.0% for the initial 3 weeks of storage and remained statistically equal to 1% through 5 weeks of storage for both CPD–AS-1 and CPD-N–AS-1N with glucose maintenance. Importantly, the hemolysis values for these solutions were statistically equal (p = 0.366), suggesting that the normoglycemic storage solutions with feeding do not increase the rate of lysis in comparison to the currently accepted CPD–AS-1 processing protocol. In the absence of glucose (non-feeding protocol), the CPD-N–AS-1N solutions were not sufficient for maintaining acceptable hemolysis rates.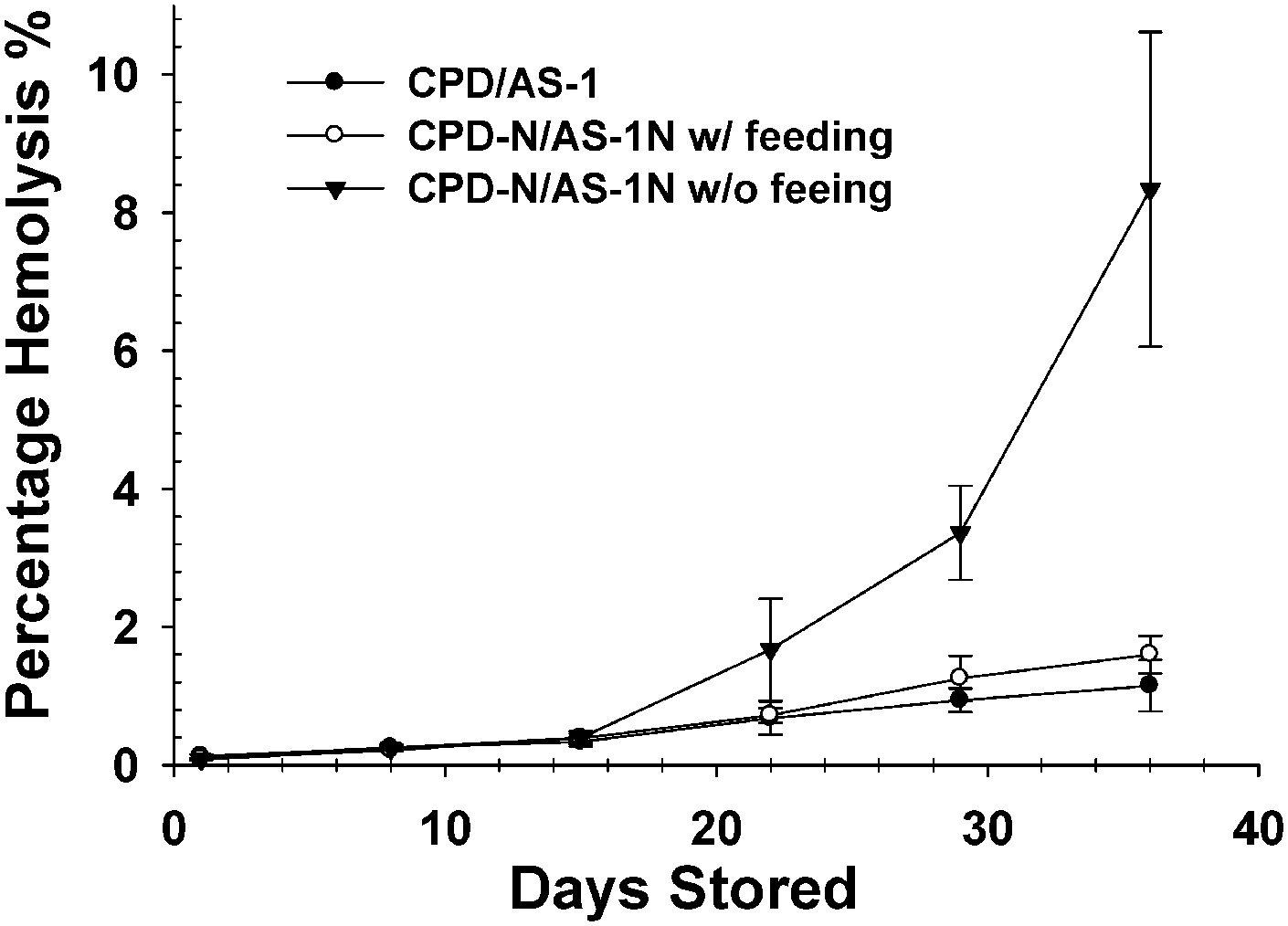 | ||
| Fig. 5 The percentage lysis of RBCs stored in CPD–AS-1 (black circle), CPD-N–AS-1N with periodic glucose feeding (white circle), and CPD-N–AS-1N without periodic glucose feeding (black triangle) is shown. It is obvious from the data shown that AS-1N storage without glucose maintenance is detrimental to RBC lysis. However, for those cells stored in AS-1N with glucose feeding, the extent of lysis is statistically equal to those cells stored in AS-1 (p = 0.366). Data represent mean ± s.e.m. (n = 4 for all). | ||
Discussion
In the United States, the NCBUS reported 15.7 million units were annually donated in its most recent survey, a value exceeding the number of units of red cells transfused during that period by over 1 million.2 However, even though there was not a numerical shortage of stored units of RBCs, according to the NCBUS, post-transfusion complications are still an area of concern in transfusion medicine. Various reports have suggested that, in some instances, people are less likely to survive after receiving a transfusion in comparison to those who did not receive any units of stored blood components.32–35 It is also well-established that people receiving transfused RBCs in storage for more than two weeks are more likely to develop complications than those receiving RBCs stored for less than two weeks.32,36 Whenever possible, neonates should be given the freshest stored cells possible, typically less than 5 days in storage.37 Collectively, these reports suggest that while blood banking centers and hospitals are doing an outstanding job of managing units of stored RBCs, the complications resulting from transfusion are serious, especially as these units age in storage.To be effective, the transfused RBC must be intact, have sufficient survival rates in vivo, and circulate.38 These three properties translate to a required reduction in RBC lysis, the ability of the transfused RBCs to avoid removal due to cell apoptotic or cell senescence mechanisms, and the ability of the transfused RBCs to traverse blood vessels. In combination, satisfying these three properties helps ensure that the RBC will be able to conduct its primary physiological objective of delivering heme-bound oxygen to demanding organs and tissues.
In addition to oxygen delivery, the RBC has also been reported to be a major determinant in the maintenance of vascular caliber.6,7 Multiple mechanisms that describe how the RBC fulfills its role in the control of blood flow have been proposed, although these mechanisms generally fall into one of two categories. One describes the RBC's ability to donate NO directly.6 Importantly, these mechanisms have been shown to play a role in stored RBCs and, in fact, recent work has shown that the ability to reload stored RBCs with components that could replenish NO levels have a beneficial impact on tissue oxygenation in transfused animal models.39
A second category portrays the RBC's ability to stimulate NO production in other cell types by its ability to release ATP (a known stimulus of endothelial nitric oxide synthase (eNOS)).7,40 Similar to mechanisms involving direct release of NO from the RBC, RBC-derived ATP has been shown to be altered during storage. McMahon et al. reported that RBCs stored for longer than one week were incapable of releasing ATP in response to hypoxia, a well-recognized stimulus of ATP release from RBCs.41 This same work also demonstrated that the reduction in ATP release was correlated with an increase in RBC adherence to endothelial cells. It was also concluded that methods to enhance ATP release from the RBCs during storage would be beneficial to patients receiving a transfusion.
Importantly, a reduction in ATP release from RBCs exposed to various stimuli is not limited solely to stored RBCs. Reduced ATP release from RBCs purified from the whole blood of people with primary pulmonary hypertension30 and cystic fibrosis42 has been reported. Furthermore, our group and others have reported that ATP release from the RBCs of people with type 2 diabetes is also significantly lower in comparison to healthy, non-diabetic controls.18,43 These reports involving ATP release from the RBCs of diabetic individuals are potentially important when one considers a hallmark feature of diabetes, namely, the hyperglycemic conditions of the bloodstream, and the glucose levels in the solutions used during the processing of whole blood for RBC storage. The bloodstream glucose concentration of a person with diabetes is typically between 7 and 9 mM, or higher if poorly controlled. As mentioned previously, the concentration of glucose in the current solutions used to collect and store whole blood and RBCs, respectively, are both well above 100 mM, but it is not to be assumed that diabetes and the RBC storage lesion have the same root cause. However, a common feature of both diabetes and the RBC storage lesion is the hyperglycemic conditions to which the RBCs are exposed. Interestingly, the ATP release data for those cells processed in the CPD–AS-1 solutions overlap with those obtained in separate studies by our group and others when studying release from the RBCs of people with type 2 diabetes.18
The data in Fig. 1b provides evidence that the ability of the RBC to release ATP in response to mechanical deformation is compromised when processed using the standard CPD–AS-1 collection and storage solutions. When a separate aliquot of these RBCs were analyzed in normoglycemic versions of these solutions (CPD-N–AS-1N), the ATP release was significantly increased. In fact, our results show that the ATP release on day 16 in the CPD-N–AS-1N system was statistically equal (p = 0.735) to that of the CPD–AS-1 system on day 1. Such results are important considering that complications associated with transfusions generally increase when using RBCs that have been stored for more than 2 weeks.
Unfortunately, storing the RBCs in a normoglycemic solution (AS-1N) resulted in the complete exhaustion of glucose in the storage bag, as indicated by both lactate accumulation measurements (Fig. 1c) and direct determination of glucose. Therefore, a rudimentary feeding protocol was developed that allowed for normoglycemic levels of glucose to be maintained in miniaturized storage bags without affecting overall sample volume. When this protocol was implemented, the ATP release remained steady throughout storage; in fact, the data in Fig. 2b show that the level of ATP release on day 36 is statistically higher than the release on day 1 using standard CPD–AS-1 collection and storage solutions.
The ATP release from RBCs processed in normoglycemic solutions shown in Fig. 1 and 2 are increased in comparison to those processed with current protocols (CPD–AS-1). However, these data alone do not provide requisite evidence that INOBA in transfusion medicine is simply a result of decreased ATP release from RBCs due to high glucose levels in the solutions used to process whole blood. Therefore, a microfluidic device was employed to determine the origin of NO measured above an endothelium that was exposed to the RBCs processed in the different collection and storage solutions. The results from the microfluidic device experiments, shown in Fig. 3, suggest that the NO measured above the endothelium is a result of eNOS stimulation from RBC-derived ATP binding to the endothelium. These data are enhanced by the use of GLI (an inhibitor of ATP release) and PPADS (a puringergic receptor inhibitor that blocks the ATP binding site). NO levels were not statistically increased when these antagonists were employed. Both of these antagonists would affect endothelium-derived NO, but not necessarily RBC-derived NO; thus, the data presented here provides evidence that the NO being measured in Fig. 3c is not RBC-derived, but rather, RBC-stimulated by ATP. Not to be overlooked in this discussion of NO origin is the fact that the normoglycemic collection and storage solutions are keys to enabling the release of ATP from the stored RBCs.
The underlying biochemical reasoning for increased ATP release from the RBCs stored in normoglycemic solutions does not appear to be due to intracellular levels of ATP during storage. The data in Fig. 4 show that intracellular levels of ATP are actually slightly increased in those RBCs stored in the higher glucose-containing AS-1 solutions. This is not completely surprising as Sprague et al. have shown that people with primary pulmonary hypertension (PPH) release less ATP in response to stimulation, even though their RBCs contained levels of intracellular ATP statistically equal to those of non-PPH controls.30 Thus, even when cells have normal levels of intracellular ATP, it does not translate to normal release in response to various stimuli. Interestingly, Sprague et al. have shown that increased glycation of cellular molecules affects key components required for ATP release from the RBC.43,44 Therefore, it may be possible that the high glucose concentrations in CPD and AS-1 may be reducing ATP release by adversely affecting key molecular components in the ATP release pathway. In addition to the work by Sprague et al. showing decreased ATP release from RBCs with increased glycation, others have also shown increased advanced glycation endproducts (AGEs) in stored RBCs.45
The modified collection and storage solutions seem to have a beneficial effect on ATP release and subsequent NO production, which could be advantageous in vivo. However, another key component to successful blood transfusion is that the RBCs remain intact. In fact, regardless of how much NO is being stimulated or produced by the RBC, if RBC lysis is present, the bioavailability of NO will more than likely drop due to NO scavenging by free hemoglobin. While no measurements involving cell survival were performed in this study, the percentage of RBCs that underwent hemolysis did not significantly increase when stored in the normoglycemic versions of the collection and storage solutions, followed by maintenance of glucose levels. When these cells were not maintained with physiological levels of glucose by periodic feeding, the lysis did increase, as to be expected as glucose and ATP are both key components in maintaining proper cellular membrane structure and functionality. It is noteworthy that the hemolysis rates for the RBCs stored during this study would most likely improve, i.e., a reduction in the hemolysis rate, if the collected whole blood had been leukocyte reduced by buffy coat removal and filtration. It has been shown that mean hemolysis rates are significantly lowered when stored products are leukoreduced by filtration.46 This method was avoided here due to the small volume of samples that were collected from donors. However, future studies involving standard collection volumes would most certainly benefit from leukoreduction by filtration methods. Not only has leukoreduction been shown to affect hemolysis rates, but also lipid composition and microvesicles.47,48
Conclusions
Regardless of its origin or fate, INOBA is a recognized problem associated with transfusion of stored RBCs. Integration of microfluidic technologies enabled measurements of endothelium-derived NO to be performed in the presence of RBCs collected and stored in standard CPD–AS-1 solutions and normoglycemic versions of each. It was found that the ATP-stimulated NO production was increased by over 25% in the normoglycemic versions, even after 4 weeks of storage. Importantly, we demonstrated that storage of RBCs in solutions that are maintained at normoglycemic concentrations (∼5–6 mM) result in significantly increased levels of ATP release from the stored cells. In fact, the ATP release from RBCs stored in normoglycemic versions of the processing solutions on day 36 of storage was equal to that of RBCs stored in the current, standard versions of CPD–AS-1 on day 1 of storage. Our results, coupled with those found in the literature involving the RBCs of people with diabetes, strongly suggest that maintenance of normoglycemic levels in collection and storage solutions may help increase NO bioavailability during and after transfusion.Experimental
Blood collection
All reagents were from Sigma-Aldrich (St. Louis, MO) unless indicated otherwise. CPD and AS-1 were prepared according to standard compositions found in the literature.49 Specifically, 50 mL of CPD were prepared containing 101.9 mM sodium citrate, 15.6 mM citric acid, 128.8 mM dextrose, and 16.1 mM monobasic sodium phosphate, pH = 5.6. For AS-1, 200 mL were typically prepared containing 154.0 mM sodium chloride, 41.2 mM mannitol, 1.8 mM adenine, and 111.1 mM dextrose, and adjusted to a pH of 5.8 before autoclaving. All reagents were prepared from solid forms and used as received without further purification. Normoglycemic versions of CPD and AS-1 (CPD-N and AS-1N, respectively) were prepared in a manner identical to CPD and AS-1, but with the glucose level at 5.5 mM.All blood collection procedures from informed and consented donors were approved by the Biomedical and Health Institutional Review Board at Michigan State University. The collection process consisted of preparing 6 non-siliconized and untreated (i.e., no heparin or other anticoagulant) 10 mL glass Vacutainer tubes (BD, Franklin Lakes NJ); 3 of these tubes contained 1 mL of CPD, while the other 3 contained 1 mL of CPD-N. Next, approximately 7 mL of whole blood were collected into each tube, resulting in a total volume of 8 mL. The blood remained in the collection solutions for at least 30 min, but not more than 2 h at room temperature (∼20 °C), prior to processing. Whole blood processing consisted of centrifugation at 2000g for 10 min followed by removal of the plasma and buffy coat layers by aspiration. Importantly, an additional top 2 mm layer of the packed RBCs were also removed to minimize leukocyte presence during subsequent storage in the AS-1 or AS-1N solutions. The purified RBCs from the 3 tubes containing CPD were then combined into a single 15 mL tube, followed by the addition of AS-1 such that the ratio of packed RBC:AS-1 volume was 2:1. The same protocol was followed for RBCs collected in CPD-N and stored in AS-1N. Finally, 2 mL of the RBCs (stored in the AS-1 or AS-1N) were added to PVC bags and stored at 4 °C. Prior to use, PVC bags were sterilized under UV light overnight. The PVC bags were prepared in-house using rolled PVC and a heat sealer. The PVC did not contain any di(2-ethylhexyl) phthalate (DEHP), a common plasticizer used in clinical storage bags. All solutions used in collection and storage were autoclaved at 10 bar and 121 °C prior to use. All blood collection and storage processes were performed under sterile conditions.
The RBCs stored in the normoglycemic AS-1N solutions required periodic feeding. These cells were “fed” by opening the PVC storage bag and adding a 10 μL droplet of 200 mM glucose in saline to the RBCs and then re-sealing the bag. This helped to maintain the glucose concentrations in the stored cells at around 5 mM, while not changing the volume into which the cells were stored.
Determination of hemolysis
Percent hemolysis was determined by the ratio of free hemoglobin (Hbf) released into the surrounding media to the total Hb (Hbt) contained in the sample. Supernatant samples were prepared by centrifuging the RBC samples at 2000g for 10 min and then further centrifuging again at 15000g for 15 min. The resulting supernatant was then diluted 1/10 in Drabkin's solution. Similarly, Hbt samples were prepared by diluting RBC samples 1/1000 in Drabkin's solution so that the final cyanmethemoglobin (HbCN) concentration fell within the range of the calibration curve (0–0.8 g L−1 HbCN). After mixing, samples and standards were incubated at room temperature in the dark for 30 min. Absorbance was read at 550 nm in a commercial plate reader (Spectramax M4, Molecular Devices). The Hb concentration was then calculated from the linear equation of the calibration curve. The sample hematocrit was determined manually by collecting RBCs in microcapillary tubes, spinning in a microhematocrit centrifuge (CritSpin M960-22, Statspin) and visually quantifying the percentage of packed red cells using a microcapillary reader. Hemolysis was then calculated according to the formula:| % Hemolysis = (Supernatant Hb (g L−1) × [100 − hematocrit (%)])/Total Hb (g L−1) |
Determination of lactic acid
Samples for lactic acid accumulation were prepared using the 2-step centrifugation as described above. Next, 5 μL of supernatant were diluted to 100 μL with distilled and deionized water (DDW, 18.0 MΩ); these dilutions resulted in lactic acid concentrations within the range of a calibration curve that was prepared with external lactic acid standards ranging from 0 to 100 μM. To perform the assay, 100 μL of a solution containing lactate dehydrogenase (5.0 U mL−1) and NAD+ (5 mM) in trizma buffer (0.1 M, pH 8.9) were added to the 100 μL samples described above. This mixture was then allowed to react for 15 min at 37 °C prior to the measurement of fluorescence intensity at 460 nm (excitation at 340 nm).Flow-induced ATP release from red blood cells
ATP release from RBCs was determined using the chemiluminescent luciferase assay. A 1000 μM stock solution of ATP was prepared by dissolving 0.0055 g of ATP in 10 mL of DDW. ATP standards (0–1000 nM) were prepared by diluting aliquots of the stock ATP solution in AS-1 or AS-1N. For ATP measurements involving RBCs, all RBC samples were diluted to 7% hematocrit in either AS-1 or AS-1N. To prepare the luciferin–luciferase mixture required for the chemiluminescence determination of ATP, 2 mg of luciferin were dissolved in 5 mL of DDW and then added to a vial containing firefly tail extract, which was used as the source for luciferase.The measurement of flow-induced ATP release has been described previously.50 Briefly, to measure the ATP release, a 500 μL syringe (Hamilton, Fisher Scientific) was filled with the luciferin–luciferase mixture described above. A second syringe was filled with either ATP standards (during calibration) or a 7% RBC solution (for measuring ATP release from the cells). Both solutions were pumped through 30 cm sections of microbore tubing with an internal diameter of 50 μm (Polymicro Technologies, Phoenix, AZ) at a rate of 6.7 μL min−1 using a dual syringe pump (Harvard Apparatus, Boston, MA). The streams containing the luciferin–luciferase mixture and ATP standard or 7% RBCs combined at a mixing T-junction. The combined stream flowed through a segment (∼5 cm) of microbore tubing (i.d., 75 μm) that had its polyimide coating removed, allowing for the detection of resultant chemiluminescence emission from the reaction using a photomultiplier tube (PMT, Hamamatsu Corporation, Hamamatsu, Japan) placed in a light excluding box. The ATP release from RBCs was measured immediately after placement in the storage solutions (AS-1 or AS-1N) and weekly through day 36 of storage. In order to account for any ATP already present in the sample prior to flow, an aliquot of the cells were centrifuged and supernatant analyzed for ATP. This value was subtracted from that obtained during the flow experiments to obtain a true value of the ATP that was released from the cells, as opposed to any ATP that was already present in the extracellular matrix.
Measurement of intracellular ATP
To determine the amount of ATP in RBCs at various points during storage, samples were removed from storage bags, centrifuged and washed with tris buffered saline 4 times to remove any extracellular ATP. Based on the hematocrit of the packed RBCs, a small aliquot of these RBCs (typically between 2–3 μL) were removed and diluted with an appropriate volume of tris buffered saline to create 50 μL of a RBC solution with a hematocrit of 7%. Next, the RBCs in this 50 μL solution were lysed and diluted by the addition of 1000 μL of DDW. After 50 μL of this lysed solution were further diluted with 950 μL tris buffered saline, 100 μL aliquots of the diluted sample were mixed with 100 μL of a luciferin–luciferase mixture, and the resultant chemiluminescence intensity was measured by the PMT in a plate reader. The quantitative amounts of ATP in the RBC samples were determined based on a calibration curve generated with authentic ATP standards (from 0 to 0.5 μM), also prepared in the tris buffered saline.Measurement of ATP stimulated, endothelium-derived NO
Endothelium-derived NO production was determined using a microfluidic device fabricated by soft photolithographic methods.26,51 A poly(dimethylsiloxane) (PDMS) slab was cast by pouring a mixture of PDMS and curing agent (Sylgard 184 silicone elastomer kit, Dow Corning, midland, MI) onto a silicon-based master. This bottom slab of PDMS contained channels with inside diameters approximating those of arterioles in vivo. A second layer of PDMS, representing the top layer of our microfluidic device, is lithographically featureless, except for 1/8′′ holes that are created using a paper-hole punch. To complete the device, a track-etch polycarbonate membrane (with pore diameters of 0.2 μm, Steriltech Inc., Kent, WA) is sealed between the two PDMS layers. In this construct, the membrane represents the bottom of a “well” prepared when portions of the PDMS was removed by the paper-hole punch. Importantly, cells can now be cultured on the polycarbonate membrane surface.Microfluidic devices were prepared for cell culture by coating the polycarbonate membrane in each well in with 10 μL of a 50 μg mL−1 fibronectin solution, and subsequent air-blow drying and exposure to UV light for sterilization. Meanwhile, endothelial cells (from bovine pulmonary arterioles) were cultured in T-25 culture flasks with Dulbecco's modified eagle medium (DMEM) containing 5.5 mM glucose, 10% (v/v) fetal bovine serum, and penicillin/streptomyocin. Cells were subcultured when 80% confluent in the flasks. Cells were detached from the flask, harvested in a test tube by centrifuging at 1500g for 5 min and then resuspended in 600 μL of culture media; 10 μL of this solution were then added into each well. After 1 h, media was changed, and every hour thereafter. The device was prepared ∼12 h before experimentation.
To evaluate NO production in the endothelium, endothelial cells in each well of the device were rinsed with Hank's balanced salt solution (HBSS, to remove culture media) followed by the addition of 10 μL of a 5 μM DAF-FM (Molecular Probes, Carlsbad, CA) solution prepared in HBSS. Next, a solution of 7% RBCs was pumped through the channels of the microfludic device for 30 min at 1 μL min−1 and 37 °C. The ATP released from the flowing RBCs is able to diffuse through the pores of the membrane (the RBCs are too large to diffuse through these pores and continue to flow to a waste reservoir on the device), stimulating NO production through purinergic receptor signaling on the endothelial cells. NO that is produced interacts with the probe and an increase in fluorescence intensity is measured using an excitation wavelength of 488 nm and emission wavelength of 515 nm.
To ensure that NO production is from RBC-derived ATP stimulated endothelial cells, two control experiments were performed. First, the activity of cystic fibrosis transmembrane conductance regulator (CFTR) which plays an important role in ATP release mechanism of RBC can be inhibited by GLI. Specifically, 0.0247 g of GLI were weighed and dissolved into 10 mL of 0.05 M NaOH in a hot water bath (∼50 °C) to generate a 5 mM solution. 20 μL of this solution were added into 1 mL of 7% RBC sample to create a final concentration of 100 μM. Then, samples were incubated for 30 min at room temperature. Secondly, PPADS which is a non-selective P2 purinergic antagonist, was used to block the response of ATP stimulation from endothelial cells. Briefly, 50 μL of 100 mM PPADS stock solution (in DMSO) were diluted with 450 μL of HBSS to create a final solution of 10 mM just prior to use. After removing cell culture media and rinsing with HBSS, 10 μL of this PPADS solution were added into wells and the entire device was incubated at 37 °C for 30 min. Next, wells were rinsed several times (usually 3–4 times) with HBSS to remove excess PPADS solution, which has a dark orange color, before the addition of DAF-FM probe. Both of these two controls were performed as described above. Samples without these treatments were always run with the controls for comparison.
References
- Blood Safety and Availability, World Health Organization Fact Sheet, 2011.
- B. L. Whitaker, 2011 National Blood Collection and Utilization Survey, NBCUS American Association of Blood Banks, 2011 Search PubMed.
- J. E. Squires, Risks of transfusion, South. Med. J., 2011, 104, 762–769 CrossRef PubMed.
- B. D. Spiess, Risks of transfusion: outcome focus, Transfusion, 2004, 44, 4S–14S CrossRef PubMed.
- J. D. Roback, R. B. Neuman, A. Quyyumi and R. Sutliff, Insufficient nitric oxide bioavailability: a hypothesis to explain adverse effects of red blood cell transfusion, Transfusion, 2011, 51, 859–866 CrossRef CAS PubMed.
- B. W. Allen and C. A. Piantadosi, How do red blood cells cause hypoxic vasodilation? The SNO-hemoglobin paradigm, Am. J. Physiol.: Heart Circ. Physiol., 2006, 291, H1507–H1512 CrossRef CAS PubMed.
- M. L. Ellsworth, Red blood cell-derived ATP as a regulator of skeletal muscle perfusion, Med. Sci. Sports Exercise, 2004, 36, 35–41 CrossRef CAS PubMed.
- R. S. Sprague, M. L. Ellsworth, A. H. Stephenson and A. J. Lonigro, ATP: the red blood cell link to NO and local control of the pulmonary circulation, Am. J. Physiol., 1996, 271, H2717–H2722 CAS.
- D. J. Singel and J. S. Stamler, Chemical physiology of blood flow regulation by red blood cells: the role of nitric oxide and S-nitrosohemoglobin, Annu. Rev. Physiol., 2005, 67, 99–145 CrossRef CAS PubMed.
- J. R. Pawloski and J. S. Stamler, Nitric oxide in RBCs, Transfusion, 2002, 42, 1603–1609 CrossRef CAS.
- M. L. Ellsworth, The red blood cell as an oxygen sensor: what is the evidence?, Acta Physiol. Scand., 2000, 168, 551–559 CrossRef CAS PubMed.
- G. R. Bergfeld and T. Forrester, Release of ATP from human erythrocytes in response to a brief period of hypoxia and hypercapnia, Cardiovasc. Res., 1992, 26, 40–47 CrossRef CAS.
- A. K. Price, D. J. Fischer, R. S. Martin and D. M. Spence, Deformation-induced release of ATP from erythrocytes in a poly(dimethylsiloxane)-based microchip with channels that mimic resistance vessels, Anal. Chem., 2004, 76, 4849–4855 CrossRef CAS PubMed.
- J. Edwards, R. Sprung, R. Sprague and D. M. Spence, Chemiluminescence detection of ATP release from red blood cells upon passage through microbore tubing, Analyst, 2001, 126, 1257–1260 RSC.
- M. Raththagala, W. Karunarathne, M. Kryziniak, J. McCracken and D. M. Spence, Hydroxyurea stimulates the release of ATP from rabbit erythrocytes through an increase in calcium and nitric oxide production, Eur. J. Pharmacol., 2010, 645, 32–38 CrossRef CAS PubMed.
- J. A. Meyer, J. M. Froelich, G. E. Reid, W. K. A. Karunarathne and D. M. Spence, Metal-activated C-peptide facilitates glucose clearance and the release of a nitric oxide stimulus via the GLUT1 transporter, Diabetologia, 2008, 51, 175–182 CrossRef CAS PubMed.
- R. G. Bogle, S. B. Coade, S. Moncada, J. D. Pearson and G. E. Mann, Bradykinin and ATP stimulate L-arginine uptake and nitric oxide release in vascular endothelial cells, Biochem. Biophys. Res. Commun., 1991, 180, 926–932 CrossRef CAS.
- J. S. Carroll, C. J. Ku, W. Karunarathne and D. M. Spence, Red blood cell stimulation of platelet nitric oxide production indicated by quantitative monitoring of the communication between cells in the bloodstream, Anal. Chem., 2007, 79, 5133–5138 CrossRef CAS PubMed.
- S. Sharifi, W. H. Dzik and S. M. H. Sadrzadeh, Human plasma and Tirilazad mesylate protect stored human erythrocytes against the oxidative damage of gamma-irradiation, Transfus. Med., 2000, 10, 125–130 CrossRef CAS.
- N. S. Mangalmurti, S. Chatterjee, G. J. Cheng, E. Andersen, A. Mohammed, D. L. Siegel, A. M. Schmidt, S. M. Albelda and J. S. Lee, Advanced glycation end products on stored red blood cells increase endothelial reactive oxygen species generation through interaction with receptor for advanced glycation end products, Transfusion, 2010, 50, 2353–2361 CrossRef CAS PubMed.
- D. J. Kor, C. M. Van Buskirk and O. Gajic, Red Blood Cell Storage Lesion, Bosnian J. Basic Med. Sci., 2009, 9, S21–S27 Search PubMed.
- J. L. Wautier, M. P. Wautier, O. Chappey, C. Zoukourian, P. J. Guillausseau and L. Capron, Diabetic erythrocytes bearing advanced glycation end products induce vascular dysfunctions, Clin. Hemorheol., 1996, 16, 661–667 Search PubMed.
- C. D. Hillyer, L. E. Silberstein, P. M. Ness, K. C. Anderson and J. D. Roback, in Blood Banking and Transfusion Medicine* Basic Principles & Practice, Churchill Livingstone/Elsevier, Philadelphia, 2nd edn, 2007, ch. 12, pp. 186–188 Search PubMed.
- M. M. Engelgau, K. M. Narayan and W. H. Herman, Screening for type 2 diabetes, Diabetes Care, 2000, 23, 1563–1580 CrossRef CAS.
- A. Faris and D. M. Spence, Measuring the simultaneous effects of hypoxia and deformation on ATP release from erythrocytes, Analyst, 2008, 133, 678–682 RSC.
- S. T. Halpin and D. M. Spence, Direct plate-reader measurement of nitric oxide released from hypoxic erythrocytes flowing through a microfluidic device, Anal. Chem., 2010, 82, 7492–7497 CrossRef CAS PubMed.
- R. S. Sprague, M. L. Ellsworth, A. H. Stephenson, M. E. Kleinhenz and A. J. Lonigro, Deformation-induced ATP release from red blood cells requires CFTR activity, Am. J. Physiol.: Heart Circ. Physiol., 1998, 275, H1726–H1732 CAS.
- G. Lambrecht, T. Friebe, U. Grimm, U. Windscheif, E. Bungardt, C. Hildebrandt, H. G. Baumert, G. Spatz-Kumbel and E. Mutschler, PPADS, a novel functionally selective antagonist of P2 purinoceptor-mediated responses, Eur. J. Pharmacol., 1992, 217, 217–219 CrossRef CAS.
- E. Beutler, Biochemistry of the erythrocyte, Experientia, 1969, 25, 470–473 CrossRef CAS.
- R. S. Sprague, A. H. Stephenson, M. L. Ellsworth, C. Keller and A. J. Lonigro, Impaired release of ATP from red blood cells of humans with primary pulmonary hypertension, Bull. Exp. Biol. Med., 2001, 226, 434–439 CAS.
- R. B. Sawant, S. K. Jathar, S. B. Rajadhyaksha and P. T. Kadam, Red cell hemolysis during processing and storage, Asian J. Transfus. Sci., 2007, 1, 47–51 CrossRef CAS PubMed.
- C. G. Koch, L. Li, D. I. Sessler, P. Figueroa, G. A. Hoeltge, T. Mihaljevic and E. H. Blackstone, Duration of red-cell storage and complications after cardiac surgery, N. Engl. J. Med., 2008, 358, 1229–1239 CrossRef CAS PubMed.
- P. C. Hebert, G. Wells, M. A. Blajchman, J. Marshall, C. Martin, G. Pagliarello, M. Tweeddale, I. Schweitzer and E. Yetisir, A multicenter, randomized, controlled clinical trial of transfusion requirements in critical care. Transfusion Requirements in Critical Care Investigators, Canadian Critical Care Trials Group, N. Engl. J. Med., 1999, 340, 409–417 CrossRef CAS PubMed.
- M. N. Gong, B. T. Thompson, P. Williams, L. Pothier, P. D. Boyce and D. C. Christiani, Clinical predictors of and mortality in acute respiratory distress syndrome: potential role of red cell transfusion, Crit. Care Med., 2005, 33, 1191–1198 CrossRef.
- M. C. Kneyber, M. I. Hersi, J. W. Twisk, D. G. Markhorst and F. B. Plotz, Red blood cell transfusion in critically ill children is independently associated with increased mortality, Intensive Care Med., 2007, 33, 1414–1422 CrossRef PubMed.
- A. Tinmouth, D. Fergusson, I. C. Yee and P. C. Hebert, Clinical consequences of red cell storage in the critically ill, Transfusion, 2006, 46, 2014–2027 CrossRef PubMed.
- E. Patten, M. Robbins, J. Vincent, J. Richardson and J. Hokanson, Use of red blood cells older than five days for neonatal transfusion, J. Perinatol., 1991, 11, 37–40 CAS.
- J. R. Hess, Red cell storage: when is better not good enough?, J. Blood Transfus., 2009, 7, 172–173 Search PubMed.
- J. T. Alexander, A. M. El-Ali, J. L. Newman, S. Karatela, B. L. Predmore, D. J. Lefer, R. L. Sutliff and J. D. Roback, Red blood cells stored for increasing periods produce progressive impairments in nitric oxide-mediated vasodilation, Transfusion, 2013, 53, 2619–2628 CrossRef CAS PubMed.
- M. L. Ellsworth, C. G. Ellis, D. Goldman, A. H. Stephenson, H. H. Dietrich and R. S. Sprague, Erythrocytes: oxygen sensors and modulators of vascular tone, Physiology, 2009, 24, 107–116 CrossRef CAS PubMed.
- H. Zhu, R. Zennadi, B. X. Xu, J. P. Eu, J. A. Torok, M. J. Telen and T. J. McMahon, Impaired adenosine-5′-triphosphate release from red blood cells promotes their adhesion to endothelial cells: a mechanism of hypoxemia after transfusion, Crit. Care Med., 2011, 39, 2478–2486 CrossRef CAS PubMed.
- R. S. Sprague, M. L. Ellsworth, A. H. Stephenson, M. E. Kleinhenz and A. J. Lonigro, Deformation-induced ATP release from red blood cells requires cystic fibrosis transmembrane conductance regulator activity, Am. J. Physiol., 1998, 275, H1726–H1732 CAS.
- R. S. Sprague, A. H. Stephenson, E. A. Bowles, M. S. Stumpf and A. J. Lonigro, Reduced expression of G(i) in erythrocytes of humans with type 2 diabetes is associated with impairment of both cAMP generation and ATP release, Diabetes, 2006, 55, 3588–3593 CrossRef CAS PubMed.
- R. Sprague, A. Stephenson, E. Bowles, M. Stumpf, G. Ricketts and A. Lonigro, Expression of the heterotrimeric G protein Gi and ATP release are impaired in erythrocytes of humans with diabetes mellitus, Adv. Exp. Med. Biol., 2006, 588, 207–216 CrossRef.
- N. S. Mangalmurti, S. Chatterjee, G. Cheng, E. Andersen, A. Mohammed, D. L. Siegel, A. M. Schmidt, S. M. Albelda and J. S. Lee, Advanced glycation end products on stored red blood cells increase endothelial reactive oxygen species generation through interaction with receptor for advanced glycation end products, Transfusion, 2010, 50, 2353–2361 CrossRef CAS PubMed.
- W. A. Heaton, S. Holme, K. Smith, M. E. Brecher, A. Pineda, J. P. AuBuchon and E. Nelson, Effects of 3-5 log10 pre-storage leucocyte depletion on red cell storage and metabolism, Br. J. Haematol., 1994, 87, 363–368 CrossRef CAS.
- C. C. Silliman, M. Dzieciatkowska, E. E. Moore, M. R. Kelher, A. Banerjee, X. Liang, K. J. Land and K. C. Hansen, Proteomic analyses of human plasma: Venus versus Mars, Transfusion, 2012, 52, 417–424 CrossRef CAS PubMed.
- C. C. Silliman, E. E. Moore, M. R. Kelher, S. Y. Khan, L. Gellar and D. J. Elzi, Identification of lipids that accumulate during the routine storage of prestorage leukoreduced red blood cells and cause acute lung injury, Transfusion, 2011, 51, 2549–2554 CrossRef CAS PubMed.
- C. D. Hillyer, L. E. Silberstein, P. M. Ness, K. C. Anderson and J. D. Roback, in Blood Banking and Transfusion Medicine: Basic Principles & Practice, Churchill Livingstone/Elsevier, Philadelphia, 2nd edn, 2007, ch. 12, pp. 188–200 Search PubMed.
- R. J. Sprung, R. S. Sprague and D. M. Spence, Determination of ATP release from erythrocytes using microbore tubing as a model of resistance vessels in vivo, Anal. Chem., 2002, 74, 2274–2278 CrossRef CAS.
- J. C. McDonald, D. C. Duffy, J. R. Anderson, D. T. Chiu, H. K. Wu, O. J. A. Schueller and G. M. Whitesides, Fabrication of microfluidic systems in poly(dimethylsiloxane), Electrophoresis, 2000, 21, 27–40 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2014 |