Generating cognitive dissonance in student interviews through multiple representations
Kimberly J.
Linenberger
and
Stacey Lowery
Bretz
*
Miami University, Department of Chemistry & Biochemistry, Oxford, OH 45056, USA. E-mail: bretzsl@muohio.edu
First published on 16th January 2012
Abstract
This study explores what students understand about enzyme–substrate interactions, using multiple representations of the phenomenon. In this paper we describe our use of the 3 Phase-Single Interview Technique with multiple representations to generate cognitive dissonance within students in order to uncover misconceptions of enzyme–substrate interactions. Findings from 25 student interviews are interpreted through the lens of multiple theoretical frameworks, including personal constructivism and coherence formation. The importance of classroom teachers engaging students in dialogue about representations is discussed.
Introduction
Biochemistry is inundated with representations of phenomena that students try to understand and faculty use to teach. For the purposes of this research, a representation is a physical depiction of a phenomenon and/or object at any level of abstraction (e.g. gestures, textbook images, and student generated diagrams). An important question for chemistry education researchers focuses on the extent to which students' interpretations of biochemistry representations correspond to those meanings intended by the instructor. According to constructivism, students will try to incorporate these representations, along with the details provided from their instructor, into their own understanding of the concept (Bodner, 1986). If the constructed knowledge includes a misinterpretation of the representation, misconceptions may arise.In this article, we describe our use of the 3 Phase-Single Interview Technique (3P-SIT) (Schönborn and Anderson, 2009) with multiple representations (in this specific instance, enzyme–substrate interactions) to generate cognitive dissonance within a student. Through the lens of several theoretical frameworks, we were able to elucidate the extent to which students make connections between the representations and integrate information from the representations into their cognitive framework.
Biochemistry visual literacy & role of representations on understanding
The discipline of biochemistry regularly uses multiple representations to describe just one construct. A single protein, for example, can be represented by a color coded ball-and-stick model, ribbon backbone, or space-filling model. These representations are often further differentiated as representations of domains, subunits, and/or active sites. Students are expected to assimilate these multiple representations into a coherent whole—all while still differentiating among them. Furthermore, chemists know that what professors teach and what students learn are rarely isomorphic with one another. We need to identify what salient features of external representations students attend to and to investigate how these representations impact their conceptual framework.Numerous studies have discussed the importance of representations for promoting conceptual, cognitive and visualization ability (Ainsworth, 2006; Harrison and Treagust, 2000; Kozma, 2003; Kozma and Russell, 1997; Lowe, 2003; Schnotz and Kürschner, 2008; Seufert, 2003; Tasker, 2005). Kozma (2003), along with Lowe (2003), have shown that upon introduction of a new concept, students look only at the surface-level features of the representation and are often not able to understand what these features represent. Tasker (2005) has argued that the type and clarity of representation is important to students' understanding. Furthermore, representations can also induce conceptual difficulties as reported by Hull (2003), Schönborn, et al. (2002), and Crossley et al. (1996) regarding representations of various biochemical processes.
An investigation of students' understanding of representations of Immunoglobulin G and antibody-antigen interaction revealed a range of difficulties (Schönborn et al., 2002). Students thought the simple non-covalent interaction between antibody and antigen represented complex processes such as secondary immune response or DNA processing. The structural features of the representation also created difficulties for the students: the red spherical shapes representing the variable and constant regions of the antibody were referred to by students as red blood cells because of their shape and color. Schönborn and Anderson (2002) concluded that
“…visualization of biochemical ERs [external representations] can be extremely challenging for students and can lead to a range of conceptual, visualization, and reasoning difficulties that can impact negatively on their understanding of molecular and cellular phenomenon.” (Schönborn et al., 2002, p. 97)
Current methods to determine role of representations on understanding
Schönborn et al. (2007) describe three methods for probing student understanding of representations: free response surveys, student generated diagrams, and clinical interviews. Free response questions allow a student to write what comes to mind without being forced into a specific line of thought. However, this technique does not permit in depth questioning to fully understand a student's cognitive construct. Asking a student to generate a diagram does allow easier access to the student's thinking, however, the drawing gives only a pictorial representation without full explanation of what objects represent. Therefore, Schönborn and Anderson (2009) developed a clinical interview they refer to as the 3P-SIT. This technique is a post-Piagetian, semi-structured interview technique (Bretz, 2007) that allows the student to speak freely, while still permitting the interviewer to probe more deeply into the student's understanding. The 3P-SIT consists of three phases: (1) probing a student's conceptual knowledge prior to the introduction of any representations; (2) measuring a student's ability to reason with the representation and their own knowledge; and (3) measuring the effect of the mode of representation on the student's understanding (Schönborn and Anderson, 2009). The 3P-SIT constrained students' interpretation to one ER per interview, but did not explore the influence of multiple representations on student understanding within a single interview.Methodology
Goals and research questions
The findings reported herein emerged in the context of a larger research study to investigate students' understandings of enzyme–substrate interactions and what their interpretations of multiple representations of the phenomenon revealed about those understandings. Student misconceptions as revealed by their discussions of the multiple representations were used to develop a concept inventory: the Enzyme-Substrate Interactions Concept Inventory (ESICI). Findings from a national administration of the ESICI are described elsewhere (Bretz and Linenberger, 2011). The specific research questions guiding this manuscript were did the use of a second representation induce cognitive dissonance within students, and if so, how did the students respond to the cognitive dissonance?Research design
In order to investigate the cognitive dissonance that might develop from student misinterpretation of multiple representations, we analyzed student data through the lens of three theories: personal constructivism (Geelan, 1997; Phillips, 1995), Ainsworth's theory of multiple representations (Ainsworth, 2006), and coherence formation (Seufert, 2003). The theory of personal constructivism (Geelan, 1997; Phillips, 1995) describes how students begin to construct their own understandings, and Baviskar et al. (2009) describe the four specific criteria for knowledge construction. Knowledge is connected via a comprehensive construct of facts, concepts, experiences, emotions, values and their relationships to each other. Therefore, prior knowledge must be elicited (criterion 1) before new information can be advantageously acquired into the construct. When new information does not fit the existing construct, cognitive dissonance emerges within the learner (criterion 2). Cognitive dissonance describes a psychological state where the learner's attitudes, beliefs, or behaviors are at odds with one another. This experience creates motivation for the learner to resolve the inconsistencies. The learner must choose either to reject the new information or to adapt the existing construct to fit the new information (Festinger, 1957). Application of the new knowledge (criterion 3) and reflection upon what was just learned (criterion 4) ensure that the learner incorporates the new information into the construct for long-term storage.We used the 3P-SIT interview technique as follows. Phase 1 probed students' understanding of enzyme–substrate interactions prior to the introduction of any representations. During Phase 2, the student was asked to describe and analyze the physical characteristics of the first representation for enzyme–substrate interactions, Fig. 1A. Also in phase 2, we subsequently introduced a second representation, Fig. 1B, that was similar to, but different from, the first representation. (For this particular research study, we used two pairs of representations, both conveying differing aspects of enzyme–substrate interactions. Students were given one of the two pairs of representations, either Fig. 1A/B or 2A/B) The purpose of introducing the second representation was to [potentially] generate cognitive dissonance regarding the student's efforts to simultaneously understand both representations while still describing enzyme–substrate interactions. The features of the representations and how they influenced students' understanding of enzyme–substrate interactions were explored in Phase 3. Finally, we added a 4th phase to allow the student to reflect upon any cognitive dissonance that arose during the interview. The exploration of the student's cognitive dissonance provided a metaphorical space in which we could detect and investigate misconceptions as the student tried to make meaning and connections both within and between the multiple representations.
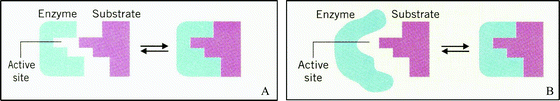 | ||
| Fig. 1 Representation set of a general enzyme–substrate interaction. The enzyme and substrate are represented along with the enzyme–substrate complex. The active site of the enzyme is also represented. (A) Representation of the lock & key theory of enzyme–substrate interaction. (B) Representation of the induced fit theory of enzyme–substrate interaction. Adapted from Chemistry and the Living Organism (p. 461), by M. M. Bloomfield and L. J. Stephens, 1996, United States: John Wiley & Sons, Inc. Copyright 1996 John Wiley & Sons Inc. Reprinted with permission. | ||
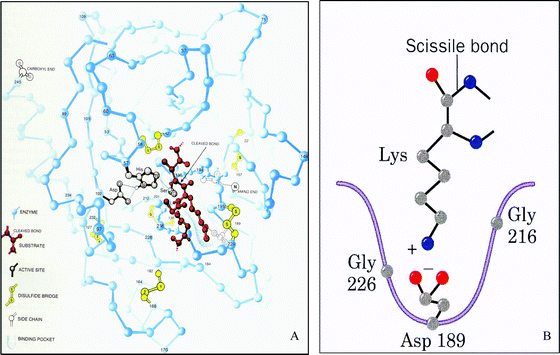 | ||
| Fig. 2 Representation set of the serine protease trypsin interacting with a peptide. (A) Representation of the backbone of trypsin interacting with the peptide substrate. The active site, binding pocket, and disulfide bonds are additionally labeled. From Chemistry: The Molecular Nature of Matter and Change, 4th Ed. (p.709), by M. S. Silberberg, 2006, New York, NY: McGraw Hill. Illustration by Irving Geis. Rights owned by Howard Hughes Medical Institute. Not to be used without permission. (B) Representation of the specificity pocket of trypsin interacting with the substrate. From Fundamentals of Biochemistry: Life at the Molecular Level, 3rd Ed (p. 351), by, D. Voet, J. G. Voet, C. W. Pratt, 2008, Hoboken, NJ: John Wiley & Sons, Inc. Copyright 2008 by D. Voet, J. G. Voet, C. W. Pratt. Reprinted with permission of John Wiley & Sons, Inc. | ||
Our second theoretical lens for analyzing student data comes from Ainsworth's (2006) general theory of learning from multiple representations. According to Ainsworth, representations serve three main functions: complementing each other in terms of both content and efficiency and also constraining one another. In combination, the effect of both complementing and constraining supports the construction of deeper understanding when the student abstracts relevant information from the representations. Seufert (2003) continues this idea of connected meaning construction from corresponding features of representations by the application of the theory of coherence formation, i.e., the third theoretical lens for analyzing our data. According to Seufert, learners are only able to acquire a deeper understanding and construct a coherent knowledge structure if they are able to make connections between features both within and between the representations of interest. Therefore, if the second representation did, in fact, induce cognitive dissonance within the student, then the connections reported by the students both within and between representations would illuminate what information the students used to build their construct of enzyme–substrate interactions. Collectively, these three frameworks enabled us to ascertain whether the connections the students made between the representations alleviated any dissonance within the student.
Representations
The interaction between enzymes and substrates is a fundamental concept in biochemistry. From the introduction of the concept during the discussion of kinetics and protein structure and function, to metabolism, and onto molecular biology, a sound understanding of enzyme–substrate interactions is essential. Due to the lack of literature on enzyme–substrate misconceptions and the breadth of representations used to teach the concept, two sets of representations typically found in textbooks were chosen (Fig. 1 and 2) to investigate what understandings each might elicit. The representations in Fig. 1 were chosen because these portray the historical models of enzyme–substrate interaction typically taught in high school and introductory chemistry and biology courses. These representations also portray a model system and not an actual enzyme and substrate as depicted in Fig. 2. The representations in Fig. 2 were chosen because serine proteases are a prototype class of enzyme typically taught along with enzyme catalysis, exemplifying acid–base and covalent catalysis. Trypsin was chosen as the specific enzyme system to ensure that the students were not just using recall of similar images of Chymotrypsin, another type of serine protease enzyme typically taught in lecture. For the representations in Fig. 2 the idea of whether or not students could connect the piece (Fig. 2B) to the whole (Fig. 2A) was also intriguing. Each representation was printed in color on a single sheet of paper, scaled to fill as much of the page as possible so a student could see the full detail of the representation. Each student was interviewed using either Fig. 1A/B or 2A/B. The representations in Fig. 1 were tested with 12 students and those in Fig. 2 with 13 students.We omitted the figure captions from the textbooks because Lowe (1993) concluded that when studying a representation that combines both text and graphics, it is difficult to discern between the contributions of each form regarding the students' mental construct of the representation. Rather, we presented each representation to the student in general terms as an enzyme–substrate interaction. This ensured that student interpretations of each representation were not influenced by what additional meanings the author conveyed through the captions.
Sample
The study was conducted at a predominately undergraduate institution in the midwestern United States. Students were purposefully sampled (see Table 1) from volunteers based upon age, sex, ethnicity and year in school from among three biochemistry courses, each serving a different population of students. Course 1 was a one semester, introductory survey course consisting of undergraduate dietetics, exercise science, zoology, and microbiology majors, with a fair majority of students intent on medical school. Course 1 followed a one semester course providing an overview of organic chemistry. Course 2 was also a one semester overview course for undergraduate chemistry majors that covered material in more depth than Course 1, but still focused on breadth of topics rather than depth. Course 3 was the first semester of an in-depth, year long biochemistry course for undergraduate biochemistry majors who completed a full year of organic chemistry. Both courses 2 and 3 included a small graduate student population. These courses were selected because enzyme–substrate interactions were taught in each of these courses for approximately two weeks. The breadth and depth of the concepts in these sections were equivalent across all three courses due to the importance of the material. Each course was taught by a different instructor.| Course 1 | Course 2 | Course 3 | Total | |||
|---|---|---|---|---|---|---|
| Undergraduate (n = 250) | Undergraduate (n = 70) | Graduate (n = 10) | Undergraduate (n = 50) | Graduate (n = 10) | ||
| Pilot | 7 | 0 | 0 | 0 | 3 | 10 |
| Full | 5 | 2 | 1 | 6 | 1 | 15 |
| Total | 12 | 2 | 1 | 6 | 4 | 25 |
Data collection
Prior to interviewing students, the three courses were observed by the first author in order to determine (1) the types of representations used by the instructors and (2) the information the students were provided regarding enzyme–substrate interactions. Students from all three courses were presented Fig. 2B in great detail during lecture. Instructors in all three courses put great emphasis on the specificity pocket, how the pocket determines which serine protease is present, and where the peptide substrate will be cleaved. Fig. 1A, B and 2A were not specifically introduced to the students prior to the interview.All participants signed an informed consent form permitting both audio and video recording of the interview. Pseudonyms are used to refer to the students in the discussion that follows. Students were interviewed for approximately 60 min at least two weeks after their course examination that tested the topics of enzyme–substrate interactions, specifically enzyme catalysis and enzyme kinetics. Digital pen and paper technology was used to capture any drawings by the students or markings upon the representations (Linenberger and Bretz, 2012). Audio recording was used to document student descriptions; video recording provided a record of student interactions with the representations (e.g., pointing at specific parts).
A pilot study was conducted to establish question efficacy and to identify any clarifications or additions to be made to the interview guide. We employed an emergent design approach to the pilot study where the interview guide was considered dynamic and, as such, adapted from interview to interview as needed to maximize the quality of data collected. The full study was conducted with a semi-structured methodology to allow for additional probes if further exploration of ideas was needed. The data presented herein includes students in both the pilot and full studies to show the range of cognitive dissonance experienced by a larger sample of students.
Data analysis
Data was managed using the software package NVivo 8 (QSR International Pty. Ltd., 2007). Both the interview transcripts and the documents collected were analyzed using the constant comparative method (Lincoln and Guba, 1985) to classify, describe, and connect data to look for categories and common themes. In order to answer the research questions, deductive analysis was used to determine if the second representation induced cognitive dissonance and, if so, how the students responded to the cognitive dissonance, using Table 2 as a rubric. A detailed description of the findings with regard to students' misconceptions about enzyme–substrate interactions will be discussed in future manuscripts. This manuscript focuses upon evidence from the interviews and data analysis for generating cognitive dissonance using multiple representations and the connections made between the representations.| Existence of cognitive dissonance | Student response upon introduction of 2nd representation | n (%) |
|---|---|---|
| Yes | sought new information from representations to support or refute their explanations | 8 (32) |
| Yes | student waited until reflection at end of interview to alleviate dissonance | 5 (20) |
| Yes | dismissed the 2nd representation as incorrect | 2 (8) |
| No | described correct understanding | 10 (40) |
| No | failed to connect the two representations | 0 (0) |
Findings
Evidence for cognitive dissonance
Recall that the purpose of introducing a second representation was to generate cognitive dissonance within the student in order to reveal any misconceptions regarding enzyme–substrate interactions. Analysis of the 25 interviews yielded multiple examples of cognitive dissonance as well as several different responses from those students who experienced cognitive dissonance.Aimeur et al. (1997) identifies several possible responses to cognitive dissonance when introduced with discrepant information: students can choose to dismiss the information as unimportant, dismiss the researcher as unimportant, or attempt to alleviate the dissonance by either changing opinion by being convinced otherwise or initiating a debate to attempt to change the researcher's opinion. We analyzed the interview data to identify (1) whether or not the students actually experienced cognitive dissonance and (2) for those students who did so, how they reacted to the cognitive dissonance per Aimeur's list of possible responses. Based on the interview guide, we chose to look for cognitive dissonance at the introduction of the second representation during Phase 2 and at the reintroduction of the first representation at the beginning of Phase 3.
The introduction of the second representation generated cognitive dissonance in the majority (n = 15, 60%) of the students. However, the ways in which these students dealt with the dissonance varied. Two of the fifteen students (one for each Figure) dismissed the second representation as incorrect. Melissa's explanation of Fig. 1A included her belief that the enzyme and substrate “just fit together perfectly. Like the enzyme and the substrate are specifically made for each other,” After Fig. 1B was introduced, however, Melissa said, “My first impression is that [the reaction] does not work, because the enzyme does not match up to the substrate. I don't see how it would bind to that funny looking thing?” As evidence of cognitive dissonance, she questioned herself upon introduction of the second representation. Instead of working to alleviate her dissonance, she chose to just dismiss the second representation. When asked to provide a caption to describe Fig. 1B, she suggested: “The incorrect reaction of an enzyme substrate intermediate.”
Eight students actively sought new information from the representations to support or refute their opinions in an effort to try to alleviate their cognitive dissonance. Todd held the same idea as Melissa in regards to Fig. 1B. His first impression was, “that with the enzyme and substrate and their configurations that they cannot come together to form this product, the enzyme substrate complex.” Unlike Melissa, however, Todd did not stop there. He went on to explain how the reaction could only occur if an additional molecule were present to change the enzyme's conformation. In fact, he changed the representation to match his understanding. Todd began to draw brackets around enzyme as shown in Fig. 3, saying
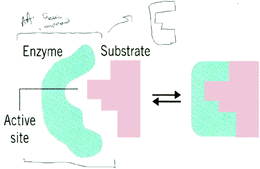 | ||
| Fig. 3 Todd's adaptation to the induced fit representation. An addition of an additional molecule was needed by Todd to understand the conformation change. | ||
“I guess these brackets represent the area around the enzyme and if you have another amino acid or protein or some other compound that can surround the enzyme and for this it would need to push [the top of the enzyme] in and push [the bottom of the enzyme] in and where I boxed it then these little grooves should fall into place to form or to match up with the substrate.”
He used information, though incorrectly, to support his opinion. Even though Todd's explanation and his adaptation to the representation reveal his lack of conceptual understanding, the cognitive dissonance Todd experienced was alleviated when he adapted the representation.
Pam, a first year biochemistry graduate student, used information from both representations to resolve her difficulties with the numbering of the amino acids and what part of the enzyme was actually represented. While explaining Fig. 2A she indicated that the numbering identified the location in the sequence of the enzyme, but when inspecting representation 2B, she became confused by the fact that the numbers were not in sequential order due to the orientation of the enzyme's backbone. Once both representations were side-by-side in Phase 3, she gathered her thoughts and resolved several issues she was having regarding the color of the backbone, the numbering of the amino acids, and exactly what was represented in Fig. 2B: “It's in um, oh no! Wait a minute, well I'm at 189. I'm right here. It's back here. Ok. So it's coming from the back, that's why it's that color.”
Megan also had issues similar to those of Pam, namely, connecting the two representations in Fig. 2. She disregarded the significance of the numbers associated with each amino acid and assumed (erroneously) that aspartate (Asp) 189 in Fig. 2b was the same as Asp 102 in Fig. 2A. She went on to explain that this was because a reaction occurs in Fig. 2B between the positive lysine (Lys) and the negative Asp 189. (Asp 102 is the amino acid found in the active site of the serine protease trypsin, the location of the reaction between enzyme and substrate, and is labeled as such in Fig. 2A.) During Phase 3 of the interview, while more closely examining the multiple representations, she realized her mistake and determined that Fig. 2B was actually the specificity pocket of trypsin—not the active site. She subsequently found the same specificity pocket in Fig. 2A.
Five students (20%), however, did not address the apparent contradictions in their explanations of the multiple representations until the end of the interview when explicitly asked to reflect by the researcher in Phase 4 of the interview. Although Megan was able to resolve some of her conflict, the multiple representations left her with unresolved conflict which she addressed during Phase 4 of the interview, asking “Why are these two there [Points to dotted lines for Asp-His and His-Ser interactions]?” She was still unclear as to the actual reasons for the other amino acids in the active site. After the interview, the first author described the roles of all three amino acids (Asp, histidine, and serine) in the active site, and Megan seemed relieved to finally figure out the piece she had been missing, namely what the active was and where it was located.
Although the majority of students experienced cognitive dissonance, ten students did not experience any form of cognitive dissonance upon the addition of the second representation. Of these ten students, six were administered the representations in Fig. 1 and four were administered the representations in Fig. 2. On first view of Fig. 1B, Abby went directly into a discussion of the induced fit model of enzyme–substrate interaction. Upon the reintroduction of Fig. 1A, she proceeded to compare and contrast the representations using the different models of interaction represented. Because she was able to correctly explain the various pieces of the representations both within the individual representations and between them, Abby was classified as not experiencing any cognitive dissonance upon addition of the second representation. The introduction of Fig. 2B to Jonathon triggered his recall of what a serine protease was, because as he stated he was more familiar with Fig. 2B (from lectures) than Fig. 2A. He accurately described the various pieces of the Fig. 2B and described the binding pockets in both representations as being similar, but with different substrate amino acid residues binding in each of the pockets.
The students' experiences with cognitive dissonance are summarized in Table 2.
Discussion and conclusion
Using the 3P-SIT interview with multiple representations created an opportunity to observe coherence formation. Phase 1 elicited the students' prior knowledge (Geelan's criterion 1). Upon the introduction of the second representation in Phase 2, some students experienced cognitive dissonance (criterion 2), discussing what connections, if any, they understood to connect the two representations. Phase 3 asked students to apply their knowledge by discussing the adequacy of the images (criterion 3). Finally, to conclude the interview, we asked the students if they had any further questions. This provided time for students to reflect on the interview and resolve any issues in their understanding that still remained (criterion 4).The addition of the second representation, which both complemented and constrained the first as described by Ainsworth (2006), generated cognitive dissonance in the majority of the students interviewed. The combination of each representation's functions forced the students to try to make connections both within and between the representations, per coherence formation (Seufert, 2003). The act of making (or, in some cases, not making) connections brought about cognitive dissonance for the majority of students. Students dealt with the dissonance, as seen in Table 2, in a variety of ways, similar to those discussed by Aimeur et al. (1997).
Implications for future research and practitioners
The cognitive dissonance generated in the interviews provided important insights into students' understanding of enzyme substrate interactions. Enzyme-substrate interactions are a vital concept in biochemistry. We need to know what students actually understand about the topic as a precursor to designing pedagogy to correct misconceptions. As the knowledge that defines the field of biochemistry continues to grow, representations to model processes will become increasingly important. Both chemistry education researchers and practitioners need methodologies to determine exactly what meanings students extract from representations and subsequently incorporate into their understanding. The use of the 3P-SIT with multiple representations not only reveals students' understandings of enzyme–substrate interactions by inducing cognitive dissonance, but also provides an additional method of examining visual literacy in biochemistry (Linenberger et al. (2012)).In the classroom, this methodology could be used by the practitioner to start an open discussion with students about what they find relevant in a representation compared to what the instructor finds relevant in a representation. Instructors could then provide students with a second representation, perhaps from another textbook or a research paper, and engage students in a discussion of what information is directly conveyed, as opposed to implied, by each of the representations. How do the multiple representations correspond? In what ways do they differ from one another? When would one representation be more appropriate to use rather than the other? Such discourse will serve both teachers and students in important ways. Teachers will have a direct window into their students' thinking. Students will not only be able to make coherent connections between representations and incorporate these into their knowledge framework, but will also be challenged to increase their visual literacy in biochemistry.
Acknowledgements
This work was supported by grant No. 0733642 from the National Science Foundation. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the author(s) and do not necessarily reflect the views of the National Science Foundation.References
- Aimeur E., Dufort H., Leibu D. and Frasson C., (1997), Some justifications about the learning by disturbing strategy, AI-ED 97 World Conference on Artificial Intelligence in Education, Kobe, Japan. 119–126.
- Ainsworth S., (2006), DeFT: A conceptual framework for considering learning with multiple representations, Learn. Instr., 16(3), 183–198.
- Baviskar S. N., Hartle R. T. and Whitney T., (2009), Essential criteria to characterize constructivist teaching: Derived from a review of the literature and applied to five constructivist-teaching method articles, Int. J. Sci. Educ., 31, 541–550.
- Bodner G. M., (1986), Constructivism—a theory of knowledge, J. Chem. Educ., 63(10), 873–878.
- Bretz S. L., (2007), Qualitative research designs in chemistry education research. In D. M. Bunce and R. S. Cole (Ed.), Nuts and bolts of chemical education research (pp. 79–99). Washington, D.C.: American Chemical Society.
- Linenberger K. J., (2011), “Biochemistry Students' Understandings of Enzyme–Substrate Interactions as Investigated through Multiple Representations and the Enzyme–Substrate Interactions Concept Inventory,” Doctoral Dissertation, Miami University, Oxford, OH 45056 USA.
- Crossley L. G., Anderson T. R. and Grayson D. J., (1996), 14th International Conference on Chemical Education, Queensland, Australia. 329.
- Festinger L., (1957), A theory of cognitive dissonance. Stanford, CA: Stanford University Press.
- Geelan D., (1997), Epistemological anarchy and the many forms of constructivism, Sci. Educ., 6, 15–28.
- Harrison A. G. and Treagust D. F., (2000), A typology of school science models, Int. J. Sci. Educ., 22(9), 1011–1026.
- Hull T. L., (2003), Students' use of diagrams for the visualization of biochemical processes. Unpublished MSc, University of KwaZulu-Natal, South Africa.
- Kozma R., (2003), Material and social affordances of multiple representations for science understanding, Learn. Instr., 13(2), 205–226.
- Kozma R. B. and Russell J., (1997), Multimedia and understanding: Expert and novice responses to different representations of chemical phenomena, J. Res. Sci. Teach., 34(9), 949–968.
- Lincoln Y. S. and Guba E. G., (1985), Naturalistic inquiry. Newbury Park, CA: Sage.
- Linenberger K. J. and Bretz S. L., (2012), A novel technology to investigate students' understandings of enzyme representations, J. Coll. Sci. Teach., in press.
- Linenberger K. J., Harle M., Orgill M. K., Towns M. H., Bretz S. L., (2012), Theoretical model in practice: Challenges to using the levels of representational competence to analyze students' thinking in biochemistry, Int. J. Sci. Educ., submitted.
- Lowe R. K., (1993), Diagrammatic information: Techniques for exploring its mental representation and processing, Inform. Design J., 7, 3–17.
- Lowe R. K., (2003), Animation and learning: Selective processing of information in dynamic graphics, Learn. Instr., 13(2), 157–176.
- Phillips D., (1995), The good, the bad, and the ugly: The many faces of constructivism, Educ. Researcher., 24(7), 5–12.
- QSR International Pty. Ltd., (2007), QSR NVivo.
- Schnotz W. and Kürschner C., (2008), External and internal representations in the acquisition and use of knowledge: Visualization effects on mental model construction, Instr. Sci., 36(3), 175–190.
- Schönborn K. J., Anderson T. R. and Mnguni L. E., (2007), Methods to determine the role of external representations in developing understanding in biochemistry. In D. e. a. Lemmermohle (Ed.), Professionell lehren—erfolgreich lernen (pp. 291–301). Munster, Germany: Waxmann.
- Schönborn K. J., Anderson T. R. and Grayson D. J., (2002), Student difficulties with the interpretation of a textbook diagram of immunoglobulin G (IgG), Biochem. Molec. Bio. Educ., 30(2), 93–97.
- Schönborn K. J. and Anderson T. R., (2009), A model of factors determining students' ability to interpret external representations in biochemistry, Int. J. Sci. Educ., 31(2), 193–232.
- Seufert T., (2003), Supporting coherence formation in learning from multiple representations, Learn. Instr., 13(2), 227–237.
- Tasker R., (2005), Using multimedia to visualize the molecular world: Educational theory in practice. In N. J. Pienta, M. M. Cooper and T. J. Greenbowe (Ed.), Chemists' Guide to Effective Teaching (pp. 195–211). Upper Saddle Ridge, NJ: Pearson.
| This journal is © The Royal Society of Chemistry 2012 |