DOI:
10.1039/C1FO10048C
(Paper)
Food Funct., 2011,
2, 395-399
Cardioprotective properties of raw and cooked eggplant (Solanum melongena L)
Received
24th March 2011
, Accepted 23rd May 2011
First published on 10th June 2011
Abstract
Although eggplants are known to be part of a healthy diet, the effects of this fruit on cardioprotection are not known. The present study examined the role of raw and grilled eggplants on cardioprotection using an isolated perfusion heart model. The animals were fed freeze-dried products of either raw or grilled eggplants for 30 days. After 30 days, isolated working hearts were subjected to 30 min ischemia followed by 2 h of reperfusion. Left ventricular function was monitored, and myocardial infarct size and cardiomyocyte apoptosis were assessed. To determine the antioxidant function of eggplants, their DPPH scavenging ability were determined, and polyphenolic components, especially nasunin content, were determined. The chemical composition of raw and grilled eggplants were determined in order to examine whether grilling was associated with major changes in their composition. The results of this study demonstrated eggplants as containing potent cardioprotective compounds judging by their ability to increase left ventricular function, and reduce myocardial infarct size and cardiomyocyte apoptosis. However, there was no difference in cardioprotective ability between the raw and grilled products. The antioxidant vitamins, including vitamin A, vitamin C and β-carotene, were lower and some of the polyphenolic components, especially nasunin content, were higher in grilled eggplants, but they were unable to demonstrate better cardioprotective properties compared to the raw fruit.
Introduction
Eggplants belong to the nightshade (Solanaceae) family of vegetables, which includes tomatoes, bell peppers and potatoes. Eggplants are considered as fruits and popular among people in Asian countries and some European countries like Italy. In the USA, they are rapidly gaining popularity among Italian food lovers. Eggplants are a rich source of vitamins and dietary fibers, as well as phytonutrients, including phenolic compounds such as caffeic and chlorogenic acids, and flavonoids.1 These chlorogenic acids possess anti-cancer, anti-microbial, anti-LDL and anti-viral activities. Eggplants are also rich in a unique anthocyanin phytonutrient called nasunin, present mainly in its skin. Nasunins are potent antioxidants, unique for chelating irons2 and can protect lipids in brain cell membranes.3 By its ability to remove excess iron, nasunins can reduce free radical formation.4
Eggplants are generally consumed after frying, boiling in water or grilling. A recent study demonstrated that the thermal treatment of eggplants increases their antioxidant content and their inhibitory effect on human neutrophil burst.5 The current study was undertaken to prove the hypothesis that eggplants possess cardioprotective effects, and if so, whether their cardioprotective ability is reduced after heat treatment. Two groups of rats were given the freeze-dried products of raw eggplants or grilled eggplants by gavaging, and their cardioprotective abilities were compared against a control group. Our results demonstrate the cardioprotective properties of eggplants without showing any difference between the raw and the cooked groups.
Materials and methods
Animal preparation
Sprague Dawley male rats of about 300 g body weight were used for our study. All animals used in this study received humane care in compliance with the principles of laboratory animal care formulated by the National Society for Medical Research, and the Guide for the Care and Use of Laboratory Animals prepared by the National Academy of Sciences and published by the National Institutes of Health (publication number NIH 85–23, revised 1985). The rats were fed ad libitum regular rat chow with free access to water. A group of rats were given (orally) for 30 days one of three preparations: (i) water only (control), (ii) freeze-dried extracts of eggplants or (iii) freeze-dried extracts of grilled eggplants. The raw eggplants were immediately frozen and lyophilized, as described previously.5 The grilling was done for 4–5 min using professional grilling apparatus followed by freeze-drying.5 The chemical composition of the freeze-dried products of raw and grilled eggplants was determined according to standard AOAC methods. After 30 days of feeding the eggplant products, the rats were sacrificed and the hearts were excised and perfused via working-mode.6
Isolated working rat heart preparation
The rats were properly anesthetized with pentobarbital (65 mg kg−1). After intravenous administration of heparin (500 IU kg−1), the chests were opened, and the hearts rapidly excised and mounted on a non-recirculating Langendorff perfusion apparatus.7,8 Retrograde perfusion was established at a pressure of 100 cm H2O with an oxygenated normothermic Krebs–Henseleit bicarbonate (KHB) buffer with the following ion concentrations (in mM): 118.0 NaCl, 24.0 NaHCO3, 4.7 KCl, 1.2 KH2PO4, 1.2 MgSO4, 1.7 CaCl2 and 10.0 glucose. The KHB buffer had been previously equilibrated with 95% O2/5% CO2, pH 7.4 at 37 °C. After perfusing the heart via the Langendorff mode for 10 min, the pulmonary vein was cannulated and the Langendorff perfusion discontinued for subsequent working heart perfusion, as described previously.6–8 It is essentially a left-heart preparation in which oxygenated KHB at 37 °C entered the cannulated pulmonary vein and left atrium at a filling pressure of 17 cm H2O. The perfusion fluid then passed to the left ventricle, from which it was spontaneously ejected through the aortic cannula against a pressure of 100 cm H2O. Aortic flow could be measured by a calibrated rotameter while coronary flow was measured by the timed collection of the coronary perfusate dripping from the heart. The aortic flow was recirculated while coronary effluent could be collected or recirculated. Heart rate, left ventricular developed pressure, and its first derivative (dp/dtmax) were acquired and recorded. Coronary flow was terminated for 15 min to induce global ischemia, which was followed by 2 h of reperfusion. Aortic pressure was measured using a Gould P23XL pressure transducer connected to a sidearm of the aortic cannula, and the signal was amplified using a Gould 6600 series signal conditioner and monitored on a real-time data acquisition and analysis system (CORDAT II). For the measurement of the left ventricular developed pressure (LVDP), a Millar catheter was inserted in the left ventricle via the tube and cannula of the left atrium and mitral valve, as described previously,6–8 and the exact LVDP was measured and recorded.
At the end of 10 min, after the attainment of steady-state cardiac function, baseline functional parameters were recorded and coronary effluent collected for biochemical assays. The circuit was then switched back to the retrograde mode and the heart perfused for 15 min with KHB buffer. The heart was then subjected to global ischemia for 30 min, followed by 2 h reperfusion.
Measurement of myocardial infarct size
At the end of reperfusion, a 10% (w/v) solution of triphenyltetrazolium in phosphate buffer was infused into the aortic cannula.9 The hearts were excised and stored at −70 °C. Sections (0.8 mm) of frozen heart were fixed in 2% paraformaldehyde, placed between two cover slips and digitally imaged using a Microtek ScanMaker 600z. To quantitate the areas of interest in pixels, NIH Image 5.1 (a public-domain software package) was used. The Infarct size (transmural) was quantified in pixels.
Assessment of cardiomyocyte apoptosis
Immunohistochemical detection of apoptotic cells was carried out using TUNEL, in which residues of digoxigenin-labeled dUP were catalytically incorporated into the DNA by terminal deoxynucleotiyl transferase II, an enzyme that catalyzes a template-independent addition of nucleotide triphosphate to the 3′-OH ends of double- or single-stranded DNA.9 The fluorescence staining was viewed with a confocal laser microscope. The number of apoptotic cells was counted and expressed as a percent of the total myocyte population.
MDA was assayed in the heart as described previously10 to monitor the development of oxidative stress. The MDA was derivatized using 2,4-dinitrophenylhydrazine (DNPH). Aliquots of 25 μL of the derivatized MDA in acetonitrile were injected onto a Beckman Ultrasphere C18 (3 mm) column in a Waters HPLC (Waters Corp., Milford, MA, USA). The products were eluted isocratically and detected at 307, 325 and 356 nm. The amount of MDA was quantitated using the Maxima software program (Waters Corp., Milford, MA, USA).
Scavenging activity of DPPH radicals
This method was based on the ability of eggplants to quench DPPH.11DPPH (1,1-diphenyl-2-picrylhydrazyl) is a synthetic and stable free radical product whose quenching by a scavenger substrate could be spectrophotometrically followed at 517 nm. An aliquot of 0.5 mL solution containing eggplant extracts was added to 1.5 mL of ethanol and a 0.5 mL ethanolic solution containing DPPH (0.5 mmol L−1). The absorbance was measured with a UV spectrophotometer at 517 nm at 20 °C.11 The ROS scavenging activity was measured against Trolox (6-hydroxy-2,5,7,8-tetramethilchroman-2-carboxylic acid), an α-tocopherol derivative.
The polyphenolic compounds were measured after separating by HPLC, followed by estimating at specific wavelengths, as described previously.11 Total phenolics were quantified by the Folin–Ciocalteau method.
Statistical analysis
For statistical analyses, a two-way analysis of variance (ANOVA) followed by Scheffe's test was first carried out to test for any differences between groups. If differences were established, the values were compared using Student's t-test for paired data. Values were expressed as mean ± SEM. The results were considered significant for p < 0.05.
Results
Chemical composition of raw and grilled eggplants
Table 1 shows the chemical composition of the raw and grilled eggplants. There was no significant difference in the composition between the raw and grilled eggplants, except for chlorine, vitamin A, β-carotene and vitamin C content, where these parameters were reduced.
Table 1 Chemical composition per 100 gm of raw and grilled eggplanta
| |
Raw |
Grilled |
|
Results are expressed as mean ± SEM of n = 2 per group. Since the determination is a very involved process, we only repeated the experiment twice, and hence no attempt was made to show a p value.
|
|
Carbohydrates (%) |
5.2 ± 0.8 |
4.9 ± 0.4 |
|
Protein/g |
1.9 ± 0.4 |
1.9 ± 0.2 |
| Fat/g |
0.4 ± 0.05 |
0.5 ± 0.04 |
| Fiber/g |
1.5 ± 0.02 |
1.6 ± 0.03 |
|
Oxalic acid/mg |
20 ± 4 |
24 ± 3 |
|
Calcium/mg |
15 ± 2 |
17 ± 2 |
|
Magnesium/mg |
14 ± 4 |
16 ± 3 |
| Phosphorus/mg |
42 ± 2 |
40 ± 1 |
|
Sodium/mg |
4 ± 0.02 |
5 ± 0.08 |
|
Iron/mg |
1.0 ± 0.05 |
1.1 ± 0.03 |
|
Copper/mg |
0.22 ± 0.01 |
0.25 ± 0.03 |
|
Potassium/mg |
2.4 ± 0.04 |
2.7 ± 0.07 |
|
Sulphur/mg |
39 ± 3.2 |
42 ± 2.0 |
|
Chlorine/mg |
48 ± 2 |
43 ± 1 |
|
Vitamin A/I. U. |
100 ± 3 |
85 ± 2 |
|
Thiamine/mg |
0.05 ± 0.01 |
0.04 ± 0.007 |
|
Riboflavin/mg |
0.15 ± 0.004 |
0.11 ± 0.01 |
|
β-Carotene/μg |
0.82 ± 0.1 |
0.68 ± 0.07 |
|
Vitamin C/mg |
15 ± 1.2 |
10 ± 1.0 |
| Energy/cal |
20 ± 2.2 |
17 ± 1.7 |
Effect of raw and grilled eggplants on myocardial function
The hearts of the rats given eggplant products orally for 30 days significantly improved post-ischemic contractile function as compared to those given the control diet. Surprisingly, both raw and grilled eggplants improved post-ischemic ventricular function in an identical manner. As shown in Fig. 1 (right), aortic flow was reduced in all groups during the post-ischemic reperfusion. Both raw and grilled eggplants significantly improved the aortic flow during the reperfusion as compared to the control group. There was no difference in the pattern of coronary flows between any of the groups (Fig. 1, left). Post-ischemic LVDP (Fig. 2, left) and LVmax![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) dp/dt (Fig. 2, right) also showed significant improvement for the hearts of rats given either of the eggplants.
dp/dt (Fig. 2, right) also showed significant improvement for the hearts of rats given either of the eggplants.
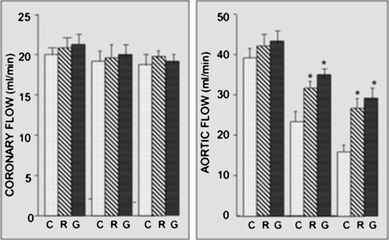 |
| | Fig. 1 Effect of raw and grilled eggplants on aortic flow (right) and coronary flow (left). Rats were given either raw or grilled eggplants for 30 days, while control experiments were performed by giving the rats water only. At the end of 30 days, isolated rat hearts were subjected to 30 min ischemia followed by 2 h of reperfusion. Aortic flow and coronary flow were determined at baseline and during the post-ischemic reperfusion. Results are expressed as mean ± SEM of six hearts per group. *p < 0.05 vs. control. | |
 |
| | Fig. 2 Effect of raw and grilled eggplants on the LVmaxdp/dt (right) and LVDP (left). Rats were given either raw or grilled eggplants for 30 days, while control experiments were performed by giving the rats water only. At the end of 30 days, isolated rat hearts were subjected to 30 min ischemia followed by 2 h of reperfusion. LVDP and LVmaxdp/dt were determined at baseline and during the post-ischemic reperfusion. Results are expressed as mean ± SEM of six hearts per group. *p < 0.05 vs. control. | |
Effect of raw and grilled eggplants on myocardial infarct size and cardiomyocyte apoptosis
Myocardial infarct size expressed as the percentage infarct of the entire risk area was only 34.5% for the control hearts subjected to 30 min ischemia followed by 2 h of reperfusion (Fig. 3 left). There was a significant reduction in the infarct size for the hearts of animals given either raw or grilled eggplants. However, there was no difference in infarct size between the two experimental groups. Cardiomyocyte apoptosis has an identical trend to that of infarct size (Fig. 3, right).
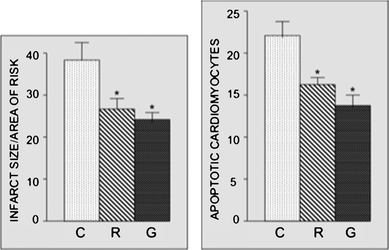 |
| | Fig. 3 Effect of raw and grilled eggplants on infarct size (left) and apoptosis (right). Rats were given either raw or grilled eggplants for 30 days, while control experiments were performed by giving the rats water only At the end of 30 days, isolated rat hearts were subjected to 30 min ischemia followed by 2 h of reperfusion. Myocardial infarct size and cardiomyocyte apoptosis were determined at the end of each experiment, as described in the Materials and methods section. Results are expressed as mean ± SEM of six hearts per group. *p < 0.05 vs. control. | |
Effect of raw and grilled eggplants on MDA formation in the heart
MDA is the presumptive marker for the development of oxidative stress and reactive oxygen species (ROS) activity in the heart. As shown in Fig. 4, the MDA content of the heart was significantly less in the hearts of animals fed either the skin or flesh of grapes. Interestingly, there were no differences between the raw and grilled eggplant-fed groups.
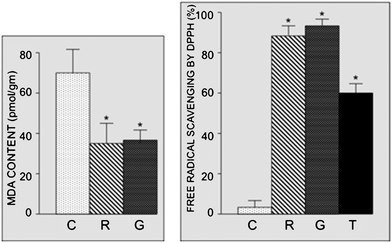 |
| | Fig. 4 Effect of raw and grilled eggplants on malonaldehyde content (left) and the DPPH scavenging of free radicals (right). Rats were given either raw or grilled eggplants for 30 days, while control experiments were performed by giving the rats water only At the end of 30 days, isolated rat hearts were subjected to 30 min ischemia followed by 2 h of reperfusion. Myocardial MDA content and DPPH scavenging activity against trolox (T) were determined at the end of each experiment, as described in the Materials and methods section. Results are expressed as mean ± SEM of six hearts per group. *p < 0.05 vs. control. | |
Phenolics content
Total phenolics were quantified by the Folin–Ciocalteau method. As was expected (Table 2), the phenolics content of the cooked (grilled) eggplants was higher compared to those present in the raw eggplants.
Table 2 Polyphenolic profiles of raw and grilled eggplantsa
| |
Raw |
Grilled |
|
Total polyphenolics were quantified by the Folin–Ciocalteau method. Results are expressed as mean ± SEM of n = 3 per determination. *p < 0.05 vs. raw.
|
|
Chlorogenic acid/ng mL−1 |
1118 ± 43 |
1958 ± 66* |
|
Caffeic acid/ng mL−1 |
14.5 ± 2.1 |
28.0 ± 2.7* |
| Nasunin/ng mL−1 |
85 ± 2.0 |
120 ± 3.6* |
| Total polyphenols/ng mL−1 |
1120 ± 56 |
1988 ± 47* |
Antioxidant activity determined by the DPPH˙ quenching method
The scavenging activity of the eggplants were determined by the DPPH˙ quenching method. The scavenging activity of 0.1 mmol L−1 Trolox is also shown as a reference for the DPPH˙ test. Each sample was analyzed in triplicate. p < 0.05 for the scavenging activity by the DPPH quenching method.
Discussion
The results of this study demonstrate that eggplants of both raw and grilled varieties can comparatively provide cardioprotection against ischemia reperfusion injury. Thus, in the eggplant groups, the aortic flow, developed pressure and maximum first derivative of the developed pressure were all higher after the hearts were subjected to 30 min ischemia and up to 2 h of reperfusion. The infarct size remained lower and the number of apoptotic cardiomyocytes were reduced. The chemical composition of the raw and grilled eggplants did not vary significantly, except for vitamin A, β-carotene and vitamin C, which were slightly higher. All three vitamins can function as intracellular free radical scavengers in the body. In addition, significant amounts of chlorogenic acid, caffeic acid and nasunin were present in the eggplants, all of which were higher in the grilled eggplants as compared to the raw eggplants.
Nasunin is the special anthocyanin that is present in the skins of eggplants (solanum melongena L). There are two isomers of nasunin, delphinidin 3-[4-(cis-p-coumaroyl)-L-rhamnosyl (1–6)glucopyranoside]-5-glucopyranoside (cis) and delphini-din-3[4-(trans-p-coumaroyl)-L-rhamnosyl-(1–6)glucopyranoside]-5 glucopyranoside (trans) [Fig. 5].12 Not only is Nasunin a potent antioxidant that is capable of scavenging free radicals, this anthocyanin is also a potent chelator of iron. Nasunin does not directly scavenge free radicals, rather it interferes with hydroxyl radical generation by chelating iron.12,13 Although, they are primarily present in their purple skin, they are equally present in our freeze-dried eggplants containing both flesh and skin. The amount of nasunin is likely to be very high in the peels of eggplants. In addition, nasunin is anti-angiogenic,13 and hence may not be very suitable for new blood vessel formation. The increase in aortic flow observed in our study could be due to the vasodilatory action of some other polyphenols.14
![Chemical structure of nasunin [delphinidin-3-(p-coumaroylrutinoside]-5-glucoside.](/image/article/2011/FO/c1fo10048c/c1fo10048c-f5.gif) |
| | Fig. 5 Chemical structure of nasunin [delphinidin-3-(p-coumaroylrutinoside]-5-glucoside. | |
In summary, our results demonstrate that both raw and grilled eggplants could reduce myocardial ischemia/reperfusion injury in ex vivo rat hearts. Interestingly enough, although the grilled eggplants had higher polyphenols, including chlorogenic acid, caffeic acid and nasunin, the cardioprotective ability of the grilled fruits were identical to those of raw eggplants. Consistent with these results, the MDA content and free radical scavenging activity of DPPH were also identical in both groups. Although, it appears that primarily the antioxidant activity of the eggplants is responsible for cardioprotection, perhaps some other parameters, such as intracellular signaling components, are also responsible for the cardioprotective effect of eggplants.
Acknowledgements
The authors are thankful to Dr Roberto Lo Scalzo of the Agricultural Research Council (CRA), Research Unit for Processing and Agrofood Industry (IAA, Milan, Italy) for supplying the eggplant extracts.
References
- A. P. Singh, D. Luthria, T. Wilson, N. Vorsa, V. Singh, G. S. Banuelos and S. Pasakdee, Polyphenols content and antioxidant capacity of eggplant pulp, Food Chem., 2009, 114, 955–961 Search PubMed.
- D. L. Luthria and S. Mukhopadhyay, Influence of sample preparation and assay of phenolic acids from eggplant, J. Agric. Food Chem., 2006, 54, 41–47 CrossRef CAS.
- Y. Noda, T. Kaeyuki, K. Igarashi, A. Mori and L. Packer, Antioxidant activity of nasunin, an anthocyanin in eggplant, Res. Commun. Mol. Pathol. Pharmacol., 1998, 102, 175–187 Search PubMed.
- Y. Noda, T. Kneyuki, K. Igarashi, A. Mori and L. Packer, Antioxidant activity of nasunin, an anthocyanin in eggplant peels, Toxicology, 2000, 148, 119–123 Search PubMed.
- R. L. Scalzo, M. Fibiani, G. Mennella, G. L. Rotino, M. D. Sasso, M. Culici, A. Spallino and P. C. Braga, Thermal treatment of eggplant (solanum melongena L) increases the antioxidant content and the inhibitory effect on human neutrophil burst, J. Agric. Food Chem., 2010, 58, 3371–3379 Search PubMed.
- D. T. Engelman, M. Watanabe, R. M. Engelman, J. A. Rousou, E. Kisin, V. E. Kagan, N. Maulik and D. K. Das, Hypoxic preconditioning preserves antioxidant reserve in the working rat heart, Cardiovasc. Res., 1995, 29, 133–140 CAS.
- R. Hattori, H. Otani, N. Maulik and D. K. Das, Pharmacological preconditioning with resveratrol: role of nitric oxide, Am. J. Physiol.: Heart Circ. Physiol., 2002, 282, H1988–H1995 Search PubMed.
- S. Das, N. Khan, S. Mukherjee, D. Bagchi, N. Gurusamy, H. Swartz and D. K. Das, Redox regulation of resveratrol-mediated switching of death signal into survival signal, Free Radical Biol. Med., 2008, 44, 82–90 Search PubMed.
- N. Maulik, S. Goswami, N. Galang and D. K. Das, Differential regulation of Bcl-2, AP-1 and NFkB on cardiomyocyte apoptosis during myocardial ischemic stress adaptation, FEBS Lett., 1999, 443, 331–336 Search PubMed.
- G. A. Cordis, N. Maulik and D. K. Das, Detection of oxidative stress in heart by estimating the dinitrophenylhydrazine derivative of malonaldehyde, J. Mol. Cell. Cardiol., 1995, 27, 1645–1653 Search PubMed.
- M. Falchi, A. Bertelli, R. L. Scalzo, M. Morassut, R. Morelli, S. Das, J. Cui and D. K. Das, Comparison of cardioprotective abilities between the flesh and skin of grapes, J. Agric. Food Chem., 2006, 54, 6613–6622 Search PubMed.
- T. Ichiyanagi, Y. Kashiwada, Y. Shida, Y. Ikeshiro, T. Kaneyuki and T. Konishi, Nasunin from eggplant consists of cis-trans isomers of delphinidin 3-[4-(p-coumaroyl)-L-rhamnosyl (1–6)glucopyranoside)-5-glucopyranoside, J. Agric. Food Chem., 2005, 53, 9472–9477 CrossRef CAS.
- K. Matsubara, T. Kaneyuki, T. Miyak and M. Mori, Antiangiogenic activity of nasunin, an antioxidan anthocyanin, in eggplant peels, J. Agric. Food Chem., 2005, 53, 6272–6275 Search PubMed.
- Y. Cao, R. Cao and E. Bråkenhielm, Antiangiogenic mechanisms of diet-derived polyphenols, J. Nutr. Biochem., 2002, 13, 380–390 Search PubMed.
|
| This journal is © The Royal Society of Chemistry 2011 |
Click here to see how this site uses Cookies. View our privacy policy here.