Phytotoxins produced by microbial plant pathogens
Richard N.
Strange
*ab
aDepartment of Biology, University College London, Gower Street, London, WC1E 6BT, UK
bDepartment of Biological and Chemical Sciences, Birkbeck College, University of London, Malet Street, London, WC1E 7HX
First published on 28th November 2006
Abstract
Covering: up to the end of 2005
Phytotoxic compounds produced by plant pathogens are often crucial determinants of plant disease. Knowledge of them provides insights into disease syndromes and may be exploited by conventional breeding and biotechnology to obtain resistant crops.
 Richard Strange | Richard Strange is a Plant Pathologist and currently Honorary Professor of Biology, University College London and Honorary Research Fellow in the School of Biology and Chemistry, Birkbeck College, University of London. He has published over 90 scientific papers and two books: Plant Disease Control, Towards Environmentally Acceptable Methods (1992) and Introduction to Plant Pathology (2003). His current interests are production of toxin-minus mutants of Ascochyta rabiei and promoting food security (see paper written with Peter Scott, ‘Plant disease: A threat to global food security’, Annu. Rev. Phytopathol., 2005, 43, 83–116). He is married to Lilian, a professional pianist, and they have two children and three grandchildren. His main hobby is playing the ’cello. |
1 Introduction
Interest in compounds synthesised by plant pathogens, which adversely affect their hosts, has usually been fired by the possibility that they may provide an explanation, at least in part, for disease symptoms. Such investigations normally start with growing the pathogen in culture and bioassaying the filtrate or, where the organism has been grown on solid media, extraction and bioassaying the extracted material. The bioassay is then used to monitor purification of phytotoxic compounds, which are normally identified by physical techniques such as mass spectrometry , ultra-violet (UV), infra-red (IR) and nuclear magnetic resonance (NMR) spectroscopy, to which may be added X-ray crystallography if crystals are available. Such studies have shown that the compounds isolated are of diverse chemical structure. Usually, a given pathogen produces a family of related compounds rather than a single toxin , and members of the family often differ in biological activity. Moreover, different isolates of the pathogen or related organisms may produce mixtures of the toxins which differ in concentration. However, in some cases, unrelated organisms also produce similar compounds, suggesting that the genes necessary for their biosynthesis have been transferred horizontally.Determination of the role of phytotoxic compounds in pathogenesis (i.e. the ability to cause disease) or virulence (i.e. the severity of disease) is critical. Some evidence is provided by demonstrating that the toxin reproduces at least some of the symptoms of the disease and that all virulent isolates of the pathogen produce the toxin . Correlation of the amount of toxin produced by different isolates of the pathogen with their virulence, and correlation of toxin sensitivity of different genotypes of the host with susceptibility comprise further evidence of toxin involvement in disease. However, the most compelling evidence is that non-toxigenic mutants are non-pathogenic or reduced in virulence, and host plants that are able to detoxify the compound or bypass the biochemical lesion that it causes are resistant or less susceptible.
The aim of this review is not to be an encyclopaedia of phytotoxic compounds synthesised by plant pathogens but rather to pick out the main lines of research, to illustrate them with appropriate examples and to point out the practical applications of such research.
2 Toxin production
Phytotoxins are usually produced in liquid media, although some produce good titres on solid media. The composition of the medium is crucial. For example, Fusarium oxysporum f. sp. lilii produced the highest titres of toxin on Czapek Dox medium, and at least part of the toxicity was attributed to fusaric acid1.1 In contrast, the solanapyrone toxins 2, 3, and 4 were only produced by the fungal pathogen of chickpea, Ascochyta rabiei, on this medium when it was supplemented with an extract of chickpea.2,3 Rather than fractionate the chickpea extract in order to determine the compound(s) responsible for triggering synthesis, Czapek-Dox nutrients were supplemented with a rich mixture of cations, amino acids and vitamins. The supplement allowed the production of the solanapyrone toxins and this finding led, by process of elimination, to the demonstration that cations were the essential components necessary for toxin production.3 Production of the partially characterised albicidins is very sensitive to medium composition: these compounds are not only phytotoxic and responsible for the systemic pathogenesis of sugar cane by Xanthomonas albilineans, but are also of interest as antibiotics , as they selectively block prokaryote DNA replication.4 Optimisation of a chemically defined medium yielded 30 times as much albicidin relative to sucrose peptone medium from half the biomass of the organism.4
Thaxtomins 5 and 6 are phytotoxins produced by Streptomyces scabies and S. acidiscabies, the causal agents of potato scab.5 Oatmeal broth was the best medium tested for their synthesis, although some production did occur in potato broth, fresh potato starch and some commercial starch preparations, but not in Luria broth or tryptic soy broth.5Toxin production was repressed 130-fold when 0.5% glucose was included in the medium,6 and tryptophan was a potent inhibitor of thaxtomin production by all five strains of S. scabies that were tested, whereas tyrosine and phenylalanine were less inhibitory.7 These results might explain the lack of thaxtomin production in Luria broth and tryptic soy broth.5 Despite the inhibitory effect of tyrosine and phenylalanine, these aromatic amino acids were shown by radiolabelling to be precursors of thaxtomin A.7

In a comparison of the production of mycotoxins and phytotoxins by isolates of the fungi Alternaria alternata and A. radicina on rice and carrot discs, most of the isolates of A. alternata produced tenuazonic acid7, alternariol8, alternariol methyl ether9 and altertoxin-I10 on rice, but only alternariol and alternariol methyl ether on carrot discs.8Radicinin11, epi-radicinol12 and radicinol13 were synthesised by A. radicina on carrot discs, but only radicinin and radicinol on rice.8

Temperature is also an important factor in toxin production. Production of thaxtomins 5 and 6 was optimal at 28 °C, whereas no coronatine14, a polyketide toxin from Pseudomonas syringae pv. glycinea, was synthesised at this temperature.6 High concentrations of coronatine, however, were produced at an optimum temperature of 18 °C.9,10

Recently, a novel environment for the production of a toxin has been discovered. A number of fungi belonging to the genus Rhizopus are responsible for seedling blight of rice. Swelling of roots is a prominent symptom and is caused by a macrocyclic polyketide , rhizoxin15. However, a search for genes encoding fungal polyketide synthases using degenerate PCR primers was unsuccessful.11 In contrast, primers designed for bacterial polyketide synthase type 1 gave a product of the expected size of 0.7 kb. When four rhizoxin-producing strains of the fungus and two non-toxigenic strains were examined by PCR , using universal primers for 16S bacterial ribosomal RNA , only the toxigenic strains gave an amplicon. Sequencing of the product showed that the bacterium belonged to the genus Burkholderia. Microscopy studies demonstrated the presence of the bacterium in the fungus, and the bacterium in pure culture synthesised the toxin and several derivatives that had previously been isolated from the fungus. In this instance the natural environment for production of the toxin by the bacterium appears to be an endosymbiotic association with a fungus!

3 Bioassays and detection
Many bioassays have been used to detect and quantify phytotoxins. Some of these involve seeds, seedlings and more mature, whole plants but many use parts of plants, isolated cells, suspension cultures or protoplasts. Once the biochemical lesion that results from application of the toxin has been discovered, this can form the basis of an assay. Also, once the chemistry of the phytotoxin has been established, it may be detected and quantified by physico-chemical techniques such as high performance liquid chromatography (HPLC ). There are a number of recent examples of the use of bioassay -guided identification of active compounds that are representative. For example, fungi from Egyptian soils have been assayed for the production of germination inhibitors of the parasitic weed Orobanche crenata.12 The assay was based on inhibition of seed germination of O. crenata seed promoted by the synthetic germination stimulant GR24. Use of this assay enabled the selection of a fungus which produced a filtrate that was highly inhibitory to the germination of O. crenata. The fungus was identified as Myrothecium verrucaria and the inhibitor as verrucarin A16.12
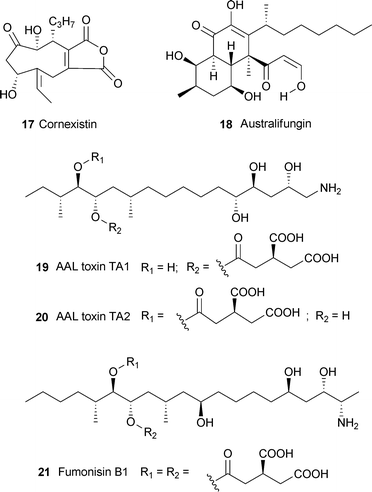
Duckweed (Lemna pausicostata) is a widely used bioassay plant that has been used to assay several toxins . For example, cornexistin17 caused inhibition of growth at 333 nM within 72 h.13 At the higher concentration of 5 µM, australifungin18, and the sphingolipid precursors phytosphingosine and sphinganine, accumulated owing to inhibition of sphingolipid synthesis, but this toxin was about eight times less potent than AAL toxins TA119 and TA220 or fumonisin B121,14 which act similarly (see Section 9). Duckweed has also been used to evaluate the toxicity of macrocyclic trichothecene toxins produced by Myrothecium verrucaria. Roridins E22 and H23, epi-isororidin E24 and verrucarins A16 and J25 were all toxic, with ED50 values ranging from 0.1 to 9.7 µM.15

Soaking rice grains in a toxin preparation from Pseudomonas fuscovaginae, which presumably consisted of a mixture of syringotoxin 26, fuscopeptin A 27 and fuscopeptin B 28,16 enhanced germination, had no effect on the number of roots, but caused drastic inhibition of seedling elongation, the extent of which correlated with susceptibility of the variety to disease in the field.17 Cuttings of seedlings were used to evaluate the toxicity of culture filtrates of Fusarium solani f. sp. glycines, the causal agent of sudden death syndrome of soybean.18 Foliar symptoms were caused on all legumes tested except mung bean (Vigna radiata) but these differed from symptoms of sudden death syndrome in soybean foliage.18

Various plant parts have been used by other authors as assay material for toxins . For example, leaf discs of kudzu (Pueraria lobata) were used to assay roridins E22 and H23, epi-isororidin E24, and verrucarins A16 and J25, which gave ED50 values ranging between 1.5 and >80 µM.15 Measurement of electrolyte leakage from plant tissues is a popular method of determining toxicity of compounds. For example, phomalactone29 caused rapid electrolyte leakage from cucumber cotyledons at concentrations of >50 ppm.19 Some authors have used tissue cultures: apple leaves cultured from meristem cells were more sensitive to AM-toxin30, the host-selective toxin of the apple pathotype of Alternaria alternata, and gave more reproducible results than an assay using leaves from trees.20 The toxicity of this host-selective toxin and some of its analogues was also evaluated using inhibition of the photosynthetic evolution of oxygen, measured with an oxygen electrode: the results obtained correlated with the ability of the compounds to induce necrosis .21 Fluorescence imaging is a method which allows comparison of toxin -damaged and healthy tissue in the same leaf,22 and has been used to examine the response of canola (Brassica napus) and white mustard (Sinapis alba) to destruxins A31 and B32: the limits of detection were 50 ng ml–1 and 10 µg ml–1, respectively.23

A drawback to the use of assays involving whole plants, plant parts and tissue cultures is the variation within the assay material owing to diffusion gradients of the toxin . These may by avoided by the use of isolated cells or protoplasts. For example, isolated cells from leaflets of chickpea (Cicer arietinum) were subjected to culture filtrates of Ascochyta rabiei, and cell viability, determined by the vital dye fluorescein diacetate, provided a score of phytotoxicity. The assay was used to monitor the isolation of the toxins , which were identified as solanapyrones A–C2–4.2,3
Another way to avoid the diffusion problems inherent in multicellular plant material is to use suspension cultures. With suspension cultures of tobacco, the LD50 value and lethal dose of the acidic protein cactorein from Phytophthora cactorum were 38 nM and >100 nM, respectively.24 Similarly, suspension cultures of Brassica species were used to evaluate the toxicity of phomalide33, destruxin B32, homodestruxin B34 and the metabolites of the latter two compounds, hydroxydestruxin B35 and hydroxyhomodestruxin B36.25,26
Where the objective is simply to quantify a known toxin , albeit in a complex mixture of compounds such as a culture filtrate, HPLC is often suitable.27 Immunoassays have been used by a number of authors to detect toxins . For example, an indirect ELISA method was developed to detect coronatine14 in host plants of Pseudomonas syringae using a specific monoclonal antibody 11B8, which reliably quantified the toxin between the limits of 5 and 40 ng ml–1.28 Moreover, use of a second antibody and immunofluorescence microscopy and immunogold labelling demonstrated that coronatine was associated with chloroplasts and particles of proteinase inhibitor in tomato cells, and that the toxin was able to migrate to healthy tissue adjacent to bacterial lesions.28
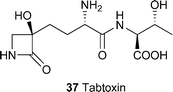
Organisms which possess the genes necessary for toxin elaboration may be detected by PCR if the appropriate sequences are known. For example, two sets of PCR primers have been developed which gave amplification products of 829 bp and 1020 bp corresponding to sequences in the tblA and tabA genes, which are required for tabtoxin37 production.29 When tested on 32 strains of Pseudomonas syringae, amplicons were only obtained with tabtoxin-producing pathovars, making this a useful test for toxigenic strains of the bacterium. Similarly, amplification of the cfl gene from the gene cluster encoding coronatine14 has been used to detect strains of Xanthomonas campestris pv. campestris, X. campestris pv. armoraciae and Pseudomonas syringae pv. maulicola capable of producing the toxin .30
Toxins are often difficult to detect in the diseased plant and, in some instances, this may because they are bound to their site of action or are converted to another compound which may be the ultimate toxin . For example, phaseolotoxin38, produced by Pseudomonas syringae pv. phaseolicola, is attacked in the plant by peptidases to release N6-(N1-sulfo-diaminophospinyl)-L-ornithine (Psorn) 39. However, destruxin B32 and homodestruxin B34, toxins from Alternaria brassicae, were detected in leaves of Brassica napus infected with the fungus.31

4 Isolation
Phytotoxins are usually isolated from in vitro cultures of the pathogen grown on either solid or liquid media. For example, AAL toxin TA119 was extracted from rice cultures with chloroform followed by methanol–water. Purification of the toxin from the aqueous phase was accomplished by chromatographic techniques.32 More frequently, toxins are isolated from cultures grown on liquid media, where they usually occur in dilute solution together with the nutrients of the medium and other products of the organism. After removal of the toxigenic organism, toxins may be concentrated and purified from the aqueous solution by partitioning into an organic solvent or by solid-phase extraction. Further purification is usually achieved by chromatography .The solanapyrone toxins of Ascochyta rabiei were isolated by flash chromatography of ethyl extracts of culture filtrates on silica gel using solvents of increasing polarity to elute the three toxins 2–4.33 However, for routine preparation of small amounts of the toxins , they are adsorbed onto C18 cartridges before HPLC on a semi-preparative C18 column using an isocratic mobile phase determined by solvent optimisation.27 A C18 solid-phase cartridge was also used as an initial purification procedure for destruxin B32 and homodestruxin B34 from culture filtrates of Alternaria brassicae. Isolation of the compounds was then achieved by HPLC .31
Klebsiella oxytoca encodes a protein, AlbA, which binds albicidins, the partially characterised toxins of Xanthomonas albilineans that cause the symptoms of scald of sugar cane.34 The AlbA–albicidin complex was stable from 4 to 40 °C but the toxin could be dissociated from the complex by heat, suggesting that the AlbA protein could be used as an affinity matrix for the purification of albicidin.34
5 Chemical synthesis
Phytotoxins produced by plant pathogenic organisms have several uses (see Section 13) but because, as mentioned in Section 4, their preparation and purification from cultures of toxigenic organisms is often laborious, there has been interest in their chemical synthesis.A racemic mixture of thaxtomins 5 and 6 was synthesised by the reaction of the lithiated anion of sarcosine anhydride with gramine methosulfate in the presence of a catalytic amount of dilithium tetrachlorocuprate in tetrahydrofuran. The remainder of the sequence of reactions involved protection of the indole amine group, aldol condensation with 3-benzyloxybenzaldehyde, bromohydrin formation and catalytic hydrogenation.35
The activity of coronatine14 in triggering volatile biosynthesis in Lima bean at 10 µM and coiling of the touch-sensitive tendrils of Bryonia dioica at 20 µM was mimicked by 6-substituted indanoyl isoleucine.36 Synthesis of this compound was achieved in four steps from 1,2,3,4-tetrahydronaphthalene and included acylation, oxidative cleavage and intramolecular Friedel–Crafts acylation reactions.36 Methods for the rational design of 4-oxoindanoyl amino acid conjugates based on coronatine with interesting properties (see Section 10) have been reported.37
6 Genetics of the biosynthesis and production of phytotoxins
Many studies have been devoted to the elucidation of the genetics of the biosynthesis and production of phytotoxins. Therefore only examples that illustrate the main lines of enquiry will be discussed.6.1 Coronatine
Coronatine 14 is a non-host-specific toxin that is synthesised by several pathovars of Pseudomonas syringae including glycinea, morsprunorum, atropurpurea, and the closely related tomato and maculicola.38 The toxin consists of two components, coronafacic acid40 and coronamic acid41. Coronafacic acid is synthesised via the polyketide pathway, and coronamic acid (1-amino-2-ethylcyclopropane carboxylic acid) is a cyclised derivative of isoleucine.39 The two components are coupled together via an amide bond to form the toxin .40 Genes involved in synthesis of the toxin (COR genes) are generally located on a plasmid but are chromosomally borne in pvs. maculicola41 and tomato.42 A study of three plasmids containing COR genes showed that they were self-transmissible and shared an oriV/par region. Several IS elements were found on a plasmid designated p4180, which mapped to regions adjacent to the COR gene cluster, and it is thought that these may have contributed to horizontal transfer of the cluster and evolution of the coronatine biosynthetic pathway.41
Coronafacic acid 40 is derived from three acetate units, one butyrate unit and one pyruvate unit,43 and the polyketide synthase responsible for its synthesis consists of two open reading frames (ORFs) that encode type I multifunctional proteins and several ORFs that encode monofunctional proteins.44 The nucleotide sequence of a 3.92 kb DNA fragment involved in coronafacic acidbiosynthesis consists of four complete ORFs and one that is incomplete.45 The predicted translation products of three of the ORFs showed similarity to acyl carrier proteins, fatty acid dehydrases and β-ketoacylsynthases, all of which are required for polyketide synthesis.45 Three further ORFs have been identified in a 4 kb region located at the 3′-end of the coronafacic acid biosynthetic gene cluster, which showed homology with oxidoreductases, thioesterases and transposases.46Coronamic acid41 is derived from L-isoleucineviaL-alloisoleucine,43 and the enzyme responsible for its synthesis contains an adenylation domain and a thiolation domain.47
In P. syringae pv. glycinea, the genes for synthesis of coronatine are located within a 32 kb region of a 90 kb plasmid designated p4180A.48 The cfl gene in this organism encodes CFA ligase, an enzyme that is responsible for activating coronafacic acid and coupling it to coronamic acid. This gene and the CFA biosynthetic gene cluster (responsible for the synthesis of coronafacic acid) give rise to a single transcript, and transcription of all of the genes in the operon are directed by the cfl promoter, which is localised to a 0.37 kb region upstream of the transcriptional start site.48Transcription of the cfl/CFA operon was temperature-sensitive, with maximal activity at 18 °C, explaining the optimum production of the toxin at this temperature9,10 (see Section 2). The sequence of cfl showed homologies with a variety of adenylate-forming enzymes that bind and hydrolyze ATP in order to activate and effect the ligation of their substrates.
Further work on the regulation of coronatine synthesis initially showed that both coronafacic acid and coronamic acid structural genes are controlled by CorR, a transcriptional activator required for the synthesis of coronatine.49 Later work showed that toxin synthesis was controlled by a two-component regulatory system consisting of two response regulators, CorR and CorP, and the histidine protein kinase CorS. The latter had six transmembrane domains and was important as a temperature sensor.50,51 Further information on the effect of host and temperature on the expression of the cma gene responsible for the synthesis of coronamic acid was obtained by fusing the cma promoter to a promoterless efgp (green fluorescent protein) gene. In P. syringae pv. glycinea, expression was temperature-dependent in minimal medium and in the host plant, but in P. syringae pv. tomato it was not significantly affected by temperature, although expression in cells recovered from infected plants appeared to be greater than that in minimal medium.52
6.2 Helminthosporium carbonum toxin (HC-toxin)
HC-toxin 42 is a host-selective toxin consisting of a family of four related compounds of which the most abundant is a cyclic peptide, consisting of D-proline, L-alanine, D-alanine and L-Aeo, where Aeo represents 2-amino-9,10-epoxy-8-oxodecanoic acid.53,54Toxin production is controlled by several genes at a complex locus termed Tox2, which contains two copies of a gene, HTS1, that encodes a large peptide synthetase of about 574 kDa. Non-toxigenic mutants which are avirulent for maize lines susceptible to the wild type are only obtained if both copies are disrupted.55Tox2 is present on either a 4 Mb chromosome or a 2.3 Mb chromosome but, in crosses of the two types, the chromosomes in the progeny did not segregate independently. Thus, instead of a 1 : 2 : 1 ratio of progeny inheriting both chromosomes, one or other, and neither, they behaved as homologues during meiosis, all progeny inheriting only one copy of the Tox2 locus. This suggests that the difference in size of the chromosomes in which the Tox2 locus occurs arose by translocation.56
TOXC is another locus required for HC-toxin production, and is present in most toxigenic strains as three copies on the same chromosome as the Tox2 locus. Targeted gene disruption of all three copies gave a phenotype which grew and sporulated normally in vitro but was not pathogenic and did not produce HC-toxin. The locus is highly homologous to the β-subunit of fatty acid synthase from several eukaryotes, and is therefore thought to be involved in the synthesis of the decanoic backbone of Aeo.57,58TOXE is a further locus in the TOX2 complex which appears to be concerned with the regulation of toxin biosynthesis and pathogenicity.59
HC-toxin is not stored in dormant spores of the fungus but is synthesised concomitantly with spore germination, and this is matched by the transcription of HTS1.60 Moreover, toxin production and appressorial formation appear to be coordinately regulated, since spores which were incubated under conditions which did not induce appressoria also failed to synthesise toxin .61
Degenerate primers designed on the basis of the sequences of cyclic peptide synthase genes gave rise to amplicons in PCR reactions with DNA from Diheterospora chlantydosporia, which synthesises the HC-toxin analogue chlamydocin43, Cylindrocladium macrosporum, which makes Cyl-244 and Cochliobolus victoriae, which synthesises the unrelated cyclic pentapeptide, victorin45.62 The amplicons were cloned and their sequences were found to be more closely related to HTS1 than to other cloned peptide synthetases. However, the identity of their amino acids was not consistent with very recent horizontal movement of these genes.

6.3 Host-selective toxins from pathotypes of Alternaria alternata
Pathotypes of Alternaria alternata produce related compounds, e.g.31 and 46–50, that are selectively toxic for their host plants. For example, the pathotype that causes brown spot of tangerines produces ACT-toxin, and the pathotype that causes leaf spot of rough lemon produces ACR-toxin. Each pathotype possesses genomic regions which are specific, that of the tangerine pathotype being a 1.05 Mb chromosome and that of the rough lemon type being another small chromosome of 2.0 Mb. Recently, a strain of A. alternata that was pathogenic to both tangerine and rough lemon was isolated and found to contain both small chromosomes.63 The authors suggest that a dispensable chromosome carrying the gene cluster that controls biosynthesis of one of the host-selective toxins was transferred horizontally and rearranged by duplication or translocation in an isolate of the fungus carrying genes for biosynthesis of the other host-selective toxin .
6.4 The solanapyrone toxins
Solanapyrones A2, B3 and C4 are produced both by Alternaria solani, the causal agent of early blight of potatoes,64 and Ascochyta rabiei, the causal agent of a devastating disease of chickpea.2,3 Both fungi belong to the order Pleosporales but to different families. It would therefore be interesting to ascertain if other fungi belonging to this order also synthesise the toxins and whether they have a role to play in any plant disease that they may cause.7 Signalling of phytotoxin synthesis
In some plant pathogens, toxin biosynthesisin vitro appears to require chemical factors found in the host. For example, Helminthosporium saccharitoxin 51 was only synthesised when an extract of sugar cane was included in the medium. The active factor was identified as serinol52 (2-amino-1,3-propanediol).65Expression of SyrB is necessary for the production of syringomycin 53 and for full virulence of Pseudomonas syringae pv. syringae. When a strain of the bacterium containing a syrB–lacZ fusion was used to monitor the activation of SyrB, syringomycin production was found to be modulated by two classes of plant signal molecules, phenolic glucosides and sugars.66 Active phenolic glucosides included arbutin 54, phenyl-β-D-glucopyranoside 55 and salicin56.66 Sugars such as D-fructose, D-mannose and sucrose enhanced the activity of the phenolic glucosides.66 Most toxigenic strains of P. syringae pv. syringae produced larger quantities of toxin in the presence of arbutin or D-glucose, 13 out of 42 produced a least 10-fold more and some strains produced toxin only in the presence of these compounds.67 Using the syrB–lacZ fusion, leaf extracts of sweet cherry were found to activate the syrB gene.68 Two flavonol glycosides , quercetin-3-rutinosyl-4′-glucoside 57 and kaempferol-3-rutinosyl-4′-glucoside 58, and a flavanone glucoside, dihydrowogonin-7-glucoside 59, were identified: the flavonoid glycosides were similar in activity to arbutin54 and their activity was enhanced by D-fructose, which itself had low activity.68 In similar work with P. syringae pv. tomato, malic and citric acids were found to be the main components in tomato leaf extracts and intercellular fluids that induced coronatine14 production.69


Host factors may be important for the production of other toxins but the compounds concerned have not been identified. For example, oat bran was the best medium for production of thaxtomin A5, and suberin was the only plant polymer that allowed the production of the toxin in minimal medium.70 Consequently, the authors suggested that extracellularesterases from the bacterium might have allowed the release of toxin -inducing molecules from lipid precursors.70
8 Role in symptom expression and disease
Several lines of evidence point to the importance of phytotoxins in a disease syndrome. In the first instance, a cell-free preparation of the toxin should reproduce at least some of the symptoms of the disease. Virulent isolates of the pathogen produce the toxin whereas avirulent ones do not, and there may be a positive correlation between toxin production by the pathogen and its virulence. There may also be a positive correlation between toxin sensitivity of the host and its susceptibility to the pathogen. Some of the best evidence for the role of toxins in plant disease comes from experiments in which mutants have been produced that lack toxin production and are reduced in virulence or have become non-pathogenic.8.1 Coronatine
When Tn5 mutants of Pseudomonas syringae pv. tomato DC3000 were screened for reduced virulence, six were found that caused reduced disease symptoms and were unable to multiply to high titres in planta. All six contained insertions in genes required for the biosynthesis of coronatine14.42 As previously mentioned, coronatine consists of two components, coronafacic acid40 and coronamic acid41, and the two components are coupled together via an amide bond to form the toxin .40 Reduced virulence of mutants with impairment of genes necessary for the synthesis of both coronafacic acid and coronamic acid was similar to that of mutants impaired in the synthesis of one or other of these two components of the toxin , leading to the conclusion that the intact toxin was required for full virulence.42In an exciting new development, coronatine14 has been found to induce systemic susceptibility in Arabidopsis thaliana, the converse of the widely reported systemic acquired resistance, which is induced by necrotrophic pathogens and mediated by salicylic acid.71 The authors hypothesise that this may be attributed to the fact that coronatine is a structural mimic of the phytohormone jasmonic acid, and that the jasmonate and salicylate signalling pathways are mutually antagonistic. However, coronatineinduced systemic resistance to herbivory by an insect, the cabbage looper Trichoplusia ni. A jasmonate-insensitive mutant (jai1) of tomato was not affected by coronatine14 and was highly resistant to P. syringae pv. tomato DC3000, which produces the toxin , but host genotypes that were defective in jasmonate biosynthesis were as susceptible to the pathogen as wild-type plants.72 On the other hand, application of methyl jasmonate to wild-type plants complemented the virulence defect of a bacterial mutant deficient in production of coronatine, further emphasising that the toxin may act as a structural mimic of jasmonate.72
8.2 Thaxtomins
Scab of potatoes is usually attributed to Streptomyces scabies and S. acidiscabies. These pathogens cause areas of necrosis surrounded by raised borders resulting from hypertrophy. Similar symptoms are caused by two piperazine toxins produced by the pathogens, which were given the trivial names thaxtomin A5 and thaxtomin B6.73 There is strong correlative data linking thaxtomin production with pathogenicity of bacteria causing scab. In one study there was a positive correlation between the pathogenicity of 23 strains of Streptomyces scabies and their ability to produce thaxtomin A5.74 In another study, 37 isolates of S. scabies, which were mainly from eastern and central Canada but also included five from the American Type Culture Collection (ATCC), showed that pathogenicity for potato was perfectly correlated with the production of thaxtomins.75 A further study showed that thaxtomin A production in culture was significantly and positively correlated with the percentage of tuber surface infected by 78 streptomycete isolates.76 When mutants of the pathogen were obtained by treatment with N-methyl-N′-nitro-N-nitrosoguanidine, one produced 300-fold less thaxtomin than the wild type and was non-pathogenic, but was also a glutamic acid auxotroph. Another mutant that was prototrophic but did not produce detectable concentrations of thaxtomins in culture supernatants was also non-pathogenic.778.3 Deoxynivalenol
When deoxynivalenol (DON) 60 was applied to the ears of 96 double haploid lines of wheat, symptoms typical of Fusarium head blight occurred in sensitive lines, consisting initially of water-soaking followed by bleaching that spread both acropetally and basipetally. A quantitative trait locus (QTL) for DON insensitivity was closely associated with a QTL for resistance to Fusarium head blight that governed symptom spread in the ear. Resistant lines detoxified DON by converting the compound to the 3-O-glucoside 61, leading the authors to suggest that the QTL for resistance might encode a DON-glucosyl transferase or regulator of the expression of such an enzyme.78
9 Mode of action
The primary sites of action of toxins fall into three categories: enzyme inhibition, interference with the properties of membranes and interference with defence responses.9.1 Enzyme inhibition
Four toxins have been described as inhibitors of ceramide synthase, an enzyme involved in sphingolipid synthesis: these are AAL toxins TA119 and TA220, fumonisin B121 and australifungin18. AAL toxins and fumonisin B1 are structurally related but australifungin is completely different.79,80 Inhibition of ceramide synthesis leads to the accumulation of sphingoid bases with consequent toxicity. In sensitive tomatoes with the asc/ascgenotype , apoptosis is induced but the co-dominant Ascallele confers toxin insensitivity and resistance to the pathogen.81Maize varieties that are sensitive to HC-toxin42 of Helminthosporium carbonum have the gentoype hm1/hm1 and are affected by doses of 100–1000 ng ml–1. Both the ketone and epoxide function of the 2-amino-9,10-epoxy-8-oxodecanoic acid moiety (Aeo) are essential for activity.82 Originally, inhibition of chlorophyll synthesis was thought to be the primary site of action of the toxin , and it was thought that this was achieved by the prevention of the formation of δ-aminolevulinic acid (ALA): reversal of the effects of the toxin by the addition of ALA supported this contention.83 Later evidence, however, showed that histone deacetylase, which reversibly deacetylates the core histones H3 and H4 in chromatin, is the site of action. Alteration of the acetylation of these histones in other systems affects gene expression and possibly, in maize, such genes might be involved in defence such as those encoding pathogenesis-related proteins or proteins that strengthen cell walls.84,85
Several pathovars and isolates of Pseudomonas syringae produce the toxin precursor tabtoxin37, a dipeptide of either threonine or serine linked to tabtoxinine-β-lactam, which is the active moiety. The compound is rather unstable and rapidly converts to the inactive δ-lactam 62. Because tabtoxin causes chlorosis, it was originally thought that the site of action was ribulose-1,5-bisphosphate carboxylase. However, this was incorrect and the mode of action of the toxin has been established as the irreversible inhibition of glutamine synthetase.86 In consequence, ammonia accumulates in sufficient concentrations to cause chlorosis.

Phaseolotoxin 38 is a tripeptide composed of homoarginine, alanine and N6-(N1-sulpho-diaminophosphinyl)-L-ornithine (PSorn) 39 produced by Pseudomonas syringae pv. phaseolicola, the causal agent of halo blight of bean. The toxin is also produced by P. syringae pv. actinidiae, which causes bacterial canker of kiwifruit.87 Besides causing chlorosis in bean, the toxin is necessary for systemic invasion of the plant by the pathogen.88 It is a potent and reversible inhibitor of ornithine transcarbamoylase (OCTase), an enzyme that catalyses the synthesis of citrulline from ornithine and carbamoyl phosphate. In planta, the toxin is attacked by peptidases to release 39, which causes the inhibition of OCTase. This results in the blocking of arginine synthesis, as production of its precursor, citrulline, is inhibited. Consequently, protein accumulation is inhibited and chlorosis is thought to be the result of lower protein levels. Significantly, toxin -treated tissue that had become chlorotic regreened on the addition of arginine.89
9.2 Interference with membranes
Water-soaking is the first visible symptom of the action of many phytotoxins, implying a perturbation of the membranes of the host plant. For example, the initial effects of two naphthazarin toxins , diyhdrofusarubin63 and isomarticin64, on the cytology of leaves of rough lemon (Citrus jambhiri) seedlings were on the chloroplast membranes, the plasmalemma and the tonoplast.90
Investigations of several phytotoxins have shown that they cause pore formation in lipid bilayers, and may also have surfactant properties. For example, several lipodepsipeptides such as the syringopeptins 65 and 66 and syringomycin 53 form pores in lipid bilayers, but often these have been studied in model membranes or red blood cells.91,92 A decrease in the rate of photosynthesis in leaves of Xanthium strumarium treated with syringopeptins was also attributed to the toxins ' effects on biological membranes.93
In tobacco protoplast assays, syringopeptin forms SP22A 65 and SP22B 66 were of similar activity in causing lysis of protoplasts and measurable 45Ca2+ influx with a threshold concentration of 50 ng ml–1.94 The toxins were also potent biosurfactants, lowering the interfacial tension of HPLC -grade water to 36 and 34.5 nm m–1, respectively; the critical micellar concentration was 0.8 mg ml–1 for both forms of the toxin . A model that integrates known effects of the toxins on ion flux in plasma membranes with the formation of ion channels and the consequential cascade of effects associated with cellular signalling has been proposed.95

A partially purified toxin from Phytophthora cactorum with a molecular weight of about 15 kDa specifically affected the trans-membrane electrical potential of apple cells of cultivars that were susceptible to the pathogen.96
9.3 Suppression of host defence responses
A recent review asserts that suppression of host defences is a key step in the pathogenesis of plants by Pseudomonas syringae.97 One agent of suppression is the toxin coronatine14. Infection of Arabidopsis thaliana by a coronatine-producing (Cor+) P. syringae pv. tomato strain DC3000 was compared with that caused by a coronatine-defective (Cor–) strain DC3661. Plants were inoculated either by infiltration or by dipping them into bacterial suspension. The Cor+ strain caused severe disease symptoms, and multiplied by 4–6 logs after inoculation by either method. In contrast, the Cor– strain failed to produce any disease symptoms and multiplied by only 1–1.5 logs in dipped plants, whereas it caused mild symptoms and multiplied by 6 logs over the 4 day experimental period in plants inoculated by infiltration. Similar results were obtained in experiments with tomato, the natural host of the bacterium. Higher levels of mRNAs of several distinct defence-related genes were consistently found in Arabidopsis leaves infiltrated with the Cor– strain than in those infiltrated with the Cor+ strain.98More recently, expression of NHO1, a gene required for limiting the growth in planta of non-host Pseudomonas bacteria, but completely ineffective against the virulent bacterium Pseudomonas syringae pv. tomato DC3000, has been investigated.99Transcription was activated by flagellin, and a strain of P. syringae pv. tabaci that lacked flagellin was unable to induce NHO1 in Arabidopsis, multiplied much better in the plant than the wild-type bacterium and caused disease symptoms. Although P. syringae pv. tomato DC3000 possesses flagellin, induction of NHO1 was rapidly suppressed by the bacterium in a type III secretion system-dependent manner. When bacterial molecules, known as effectors, which induce resistance or susceptibility according to the genotype of plant in which they are introduced, were tested in protoplasts for suppression of NHO1induction, nine were found: HopS1, HopAI1, HopAF1, HopT1-1, HopT1-2, HopAA1-1, HopF2, HopC1 and AvrPto. In addition, coronatine14 also suppressed flagellin-induced NHO1transcription.99
Some fungal toxins have been found to suppress defence responses by plants. For example, helminthosporol67, a toxin from Cochliobolus sativus, strongly inhibited β-1,3-glucan synthase.100 This enzyme is responsible for the synthesis of callose, an early response to pathogen attack that has been implicated as a resistance mechanism. Other fungal toxins suppress phenylalanine ammonia-lyase activity, a key enzyme in plant defence. Cytochalasin A68, zearalenone69, pinolidoxin70, putaminoxin71 and fusarenone X72 all suppressed phenylalanine ammonia-lyaseinduction in suspension cultures of hybrid poplar at 10–4 M or less.101

10 Analogues, metabolites and structure–activity relationships
Plant pathogens usually do not synthesise a single compound with phytotoxic properties but rather a family of related compounds that vary in their toxicity. Thaxtomin A5 is the principal toxin of Streptomyces scabies but the bacterium also produces a number of related compounds in vitro. Four of these were characterised spectroscopically as p-hydroxyphenyl- 73, 3,4-dihydroxyphenyl- 74, 15-de-N-methyl- 75, and 12-de-N-methyl- 76 analogues of thaxtomin A.102 Some analogues, as in the case of those of the toxin of Helminthosporium sacchari51, which is specific for sugar cane, may even compete with the principal toxin .103
Pathotypes of Alternaria alternata are of particular interest as they produce a number of related compounds (30 and 46–50) that are host-selective. As pointed out in Section 6, in some cases the genes necessary for their biosynthesis are borne on small dispensable chromosomes which may be horizontally transferable. In other instances, organisms that differ widely phylogenetically produce the same or similar compounds. For example, coronatine14 and its analogues were not known outside the genus Pseudomonas until it was found that Xanthomonas campestris pv. phormiicola produced N-coronafacoyl-L-valine77 and N-coronafacoyl-L-isoleucine78.104

Eutypine
79 has been implicated as the principal toxin of Eutypa lata,105 but later work showed that an isolate of the fungus from California which was pathogenic to grapevines did not produce the compound. Instead, the related compound eulatochromene80 was found and chemically synthesised.106 In a bioassay measuring chlorophyll loss of grapevine leaves, the compound showed greater toxicity than eutypine. The cyclised product 5-formyl-2-methylvinyl[1]benzofuran81 was also active, whereas the reduced product eutypinol82 and the quinol 83 were inactive.106

The structure–activity relationships of six sphaeropsidin toxins 84–89 produced by Sphaeropsis sapinea f. sp. cupressi, as well as eight derivatives from three of the compounds 90–97 were investigated. The C8–C14 double bond, the tertiary hydroxyl group at C9, the vinyl group at C13, and the carboxylic group at C10, as well as the integrity of the A-ring were necessary for the compounds to have non-selective phytotoxic and antimycotic activity.107


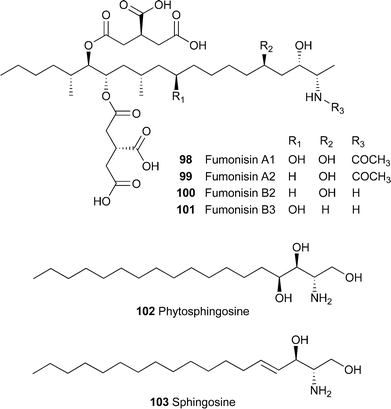
With the fumonisins A198, A299, B121, B2100 and B3101 and AAL toxins 19 and 20, the propanetricarboxylic acid groups were necessary for herbicidal activity, but this was markedly reduced by acetylation of the terminal amine group.108 The structurally related sphingolipids phytosphingosine102 and sphingosine103 were also phytotoxic, giving rise to similar symptoms as those caused by fumonisin B121, but their activity was about two orders of magnitude less than that of the toxin .108
In a rather different vein, indanoyl amino acid conjugates based on the structure of coronatine were found to induce defensive responses in plants that surpassed those of octodecanoid hormones and also caused the efficient abscission of fruit.37
11 Self-protection
How toxigenic organisms protect themselves from their own toxins has always been an intriguing question. In the case of plant pathogens, several mechanisms are used. Trichothecenes 104–108 are toxins produced predominantly by species of Fusarium which have multiple inhibitory effects on eukaryote cells: these include inhibition of protein, DNA and RNA synthesis, inhibition of mitochondrial function, and malfunctioning of membranes and cell division.109 A gene, Tril2, which appears to encode a trichothecene pump that protects Fusarium sporotrichioides from the effects of these toxins , has been isolated. When the gene was disrupted, incorporation of trichothecene in the medium inhibited growth, and growth on complex media and levels of trichothecene were reduced.110
A gene with similar properties to Tril2 has been found in Helminthosporium carbonum, which synthesises HC-toxin42.111 The gene, named TOXA, is present as two linked copies and flanks the HTS1 peptide synthetase gene. Sequence data suggested that the gene was very similar to several members of a family of genes which confer antibiotic resistance and that it codes for a 58 kDa hydrophobic protein with 10–13 membrane-spanning regions. It was possible to mutate one of the genes in targeted gene disruption experiments but not both. Putting all the data together, the authors proposed that TOXA codes for an HC-toxin efflux pump which protects the fungus from the toxin by secreting it.111
Pathovars of Pseudomonas syringae that produce tabtoxin37 may protect themselves from the toxin by adenylation of glutamine synthetase, which renders the enzyme less sensitive to inactivation by the active moiety of the toxin , tabtoxinine-β-lactam109.112 The production of β-lactamases that hydrolyse the β-lactam ring to the non-toxic tabtoxinine110 is a second mechanism by which the bacterium may protect itself from its own toxin .

The active moiety of phaseolotoxin 38 is N6-(N1-sulfo-diaminophospinyl)-L-ornithine (PSorn) 39. It is a powerful inhibitor of ornithine transcarbamoylase (OTCase), an enzyme required for the synthesis of citrulline. However, the pathogen does not inhibit the synthesis of its own citrulline as it has two OTCase genes, one of which, argK, encodes an enzyme that is resistant to phaseolotoxin. Expression of argK is co-ordinately regulated with phaseolotoxin production, both being produced at 18–20 °C but not at 28–30 °C, which is the optimum for growth of the bacterium.113
Cercosporin
111, which is produced by several species of Cercospora, is a singlet oxygen-generating photosensitiser and is toxic to plants, mice and many fungi, but not the fungi that produce it. When the redox potential of seven fungal species that differed in sensitivity to cercosporin was investigated, using 20 tetrazolium dyes differing in redox potential , cercosporin-insensitive fungi were found to be able to reduce more dyes than sensitive fungi, and that addition of reducing agents to the media such as ascorbate, cysteine, and reduced glutathione (GSH) reduced the toxicity of cercosporin to sensitive fungi.114 The authors proposed a model in which cercosporin insensitivity was attributed to reducing power production at cell surfaces, causing transient reduction and detoxification of the cercosporin molecule.114

12 Toxin insensitivity and toxin degradation
Plants have at least three mechanisms for rendering toxins produced by pathogens ineffective. They include failure to take up the compound, bypassing the biochemical lesion caused by the toxin and enzymic conversion to less toxic compounds. Plants that are able to mitigate the effects of toxins by such means are generally more resistant to the toxigenic organism than those that lack them. In addition, there are other organisms that degrade toxins produced by plant pathogens, and some of them have found utility as biocontrol agents for toxigenic pathogens.A mutant of Arabidopsis was identified which had a lower rate of uptake of the thaxtomins 5 and 6 than the wild type and was consequently more resistant to the toxins . The wild-type gene has homologues in all fully sequenced eukaryotic genomes, and encodes a small protein which the authors suggest is a regulator of a transport mechanism.115
Investigation of the differential sensitivity of tomatoes with the Asc/Asc, Asc/asc and asc/asc genotypes to the AAL toxins 19 and 20 and fumonisin B121 showed that complementation of the asc/ascgenotype with the Asc gene conferred toxin insensitivity to plants.116 When the two alleles were sequenced and the sequences compared, the ascallele was found to have several deletions, the most important of which was a 2-bp deletion in one of the exons, giving rise to a stop codon. Asc-1 is homologous to LAG1 of Saccharomyces cerevisiae, a gene that has been associated with longevity and facilitates the transport of glycosylphosphatdylinositol (GPI)-anchored proteins from the endoplasmic reticulum to the Golgi apparatus. Expression sequence tags showed that homologues of the gene are widely transcribed, as indicated by records of full-length cDNAs from fish, mammals, insects, higher fungi and monocotyledonous and dicotyledonous plants.116 The authors propose that Asc-1 is involved in the production of sphingolipids by a pathway that is not sensitive to 19 and 20 or other inhibitors of sphingolipid synthesis, effectively bypassing the biochemical lesion caused by these compounds and thus negating the catastrophic effect of inhibiting production of these important components of membranes. Plants over-expressing the tomato Asc-1 gene are highly insensitive to 19, 20 and 21.117
Conversion of phytotoxins both by plants and other organisms to less toxic compounds is a subject of much interest: plants that are able to render phytotoxins less toxic are of value as breeding material for resistance to toxigenic organisms, and micro-organisms that do so have potential as biocontrol agents and as a means for the biotechnological production of resistant crops (see Section 13).
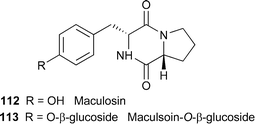
Glucosylation is a mechanism by which compounds can be rendered less toxic, and has been demonstrated in several systems. For example, maculosin112 was converted to at least three polar compounds by spotted knapweed (Centaurea maculosa). One of these was identified as maculosin β-O-glucoside113, which was not phytotoxic towards spotted knapweed or any of several other plants tested.118
Streptomyces scabies glucosylated thaxtomin A5in vitro, and the glycoside was six times less toxic to potato tuber slices than the native toxin .119 When potato cultivars differing in susceptibility to scab caused by S. scabies were treated with radiolabelled thaxtomin A, the scab-resistant cultivars produced greater amounts of a compound which, on chromatography , had an Rf value similar to the glycoside . Moreover, there was almost twice as much glucose transferase on a specific activity basis in resistant than in susceptible potato cultivars and selections. These results suggest that glucosylation is a mechanism of detoxification of thaxtomin A and is related to resistance to S. scabies.119 More recently, several hundred microorganisms that were randomly selected from the environment have been tested for their ability to transform thaxtomin A5 to much less phytotoxic metabolites. Only one was found, which was identified as Aspergillus niger. The toxin was converted by the fungus to two major and five minor metabolites. All the compounds, with the exception of two of the minor metabolites, were fully characterised by a combination of MS and NMR techniques as 114–118. The two major metabolites, 116 and 117, were far less toxic than thaxtomin A.120

Glucosylation is also a mechanism by which the toxin fommanoxin119, a product of the forest pathogen Heterobasidion annosum, is metabolised by cells of Pinus sylvestris.121 However, this is a late detoxification product, the non-toxic alcohol being produced first. A rather similar situation was found with destruxin B32, the major phytotoxin produced by Alternaria brassicae, which is the causal agent of one of the most damaging diseases of brassica crops. Sinapis alba, which is resistant to the disease, metabolised destruxin B faster than susceptible species. As in the case of fommanoxin, the first metabolite was the alcohol hydroxydestruxin B35, which was converted to the β-D-glucosyl derivative 120 more slowly. Remarkably, hydroxydestruxin B35 acted as an elicitor of the production of phytoalexins (low molecular weight, antimicrobial defence compounds) in resistant species but not in susceptible ones.122

A single, dominant Mendelian locus, Hm1, is responsible for resistance of maize to H. carbonum race 1 and insensitivity to HC-toxin42. Plants with the genotypes Hm1/Hm1 or Hm1/hm1 inactivate the toxin by reducing the carbonyl group on the side-chain of the Aeo moiety of the toxin 42.123,124 A duplicate gene, Hm2, has also been cloned but this gives only partial, adult resistance. Resistance is the wild-type condition in maize, and susceptibility arose from a transposon insertion in Hm1 and deletion of Hm2.125 Other monocots, which are not susceptible to H. carbonum race 1, possess homologues of these genes. In rice, the homologues may be related to the maize genes by vertical descent, as they are found in chromosomal regions that are syntenic with the maize loci, suggesting that their function is in defence of plants against HC-toxin or related molecules. The authors speculate that HC-toxin or related compounds may have been important in plant disease before the evolution of the HC-toxin reductase function encoded by the Hm homologues.125
Bacteria from sugar cane infected with Xanthomonas albilineans were screened for resistance to albicidins, the partially characterised toxins produced by the leaf scald pathogen. An isolate of Pantoea dispersa was highly effective in detoxifying albicidins and gave almost complete control of the disease, even when co-inoculated with a 10-fold excess of the pathogen. The authors speculate that the ability to detoxify albicidins not only allowed P. dispersa to compete with X. albilineans at wounds, but also protected the plant from the activity of the toxins .126
13 Exploitation
Where a phytotoxin produced by a plant pathogen is a determinant of pathogenicity (the ability to cause disease) or virulence (a measure of pathogenicity) the phytotoxin may be exploited in the production of plants that are resistant to the disease. One way in which this may be done is to use the toxic compound as a selective agent for genotypes that are insensitive, as such plants will be completely resistant to the toxigenic pathogen if the toxin is a determinant of pathogenicity, or more resistant than sensitive plants if the toxin is a component of virulence. In some instances the reasons for toxin insensitivity are known, and more direct approaches may be taken to produce toxin -insensitive plants that are resistant to disease. For example, plants may be selected in which an enzyme that converts the toxin to non-toxic compounds is well expressed, or plants may be genetically modified with a gene encoding such an enzyme. Conversely, toxin sensitivity might be caused by inhibition of a key enzyme in an important biochemical pathway. Here, selection could be for plants that have a variant of the enzyme that is not sensitive, or the plant could be genetically modified in order to express such an enzyme. Another way in which phytotoxins may be exploited, if they are sufficiently selective, is in weed control.13.1 Selection
A number of workers have used crude culture filtrates of pathogens or ill-defined toxins as a means of selecting genotypes of plants with increased disease resistance. This approach may lead to contradictory results.127 For example, calli from two citrus cultivars with known tolerance to mal secco disease, caused by Phoma tracheiphila, was in reverse order to the response of the two cultivars to natural and artificial inoculations.128 However, in contrast, when the toxicity of Fusarium subglutinans to pineapples was assessed by placing droplets of culture filtrates on wounded leaf segments of tissue-cultured plantlets, the results correlated with reactions to the pathogen, those genotypes that were most sensitive to the culture filtrate being the most susceptible to the pathogen.129Fusaric acid1, a product of F. subglutinans, was also tested in the assay but there was no correlation of sensitivity with susceptibility: consequently, other components of the culture were suggested to be responsible for the differential effect.129 Some scientists have been less circumspect in suggesting that culture filtrates or partially purified toxins may be used in selection. While this may be true in individual cases, the variation usually present in cultures of plant pathogens, which is difficult to control, may lead to variable results. The use of pure compounds would seem preferable.13.2 Genetic modification of plants for toxin insensitivity
In some instances it is clear why a plant is resistant to a toxigenic pathogen. For example, maize with the Hm1allele is able to detoxify the HC-toxin produced by Helminthosporium carbonum by reducing the carbonyl group on the side-chain of the Aeo moiety of the toxin 42. Here there is no difficulty in finding genotypes with this dominant allele that can be used in breeding programmes. In other systems, such breeding material may not be so readily available. For example, the tolerance of some cultivars of grapevine to Eutypa lata has been attributed to conversion of an aldehydic toxin , eutypine79, to the corresponding alcholol, eutypinol 82. In a search for plants that were a good source of such an enzyme, mung bean was identified, and a weak homology was found between the gene encoding this enzyme and the Hm1 gene.130 When calli of grapevine (Vitis vinifera) were transformed with the gene, they were far less sensitive to eutypine than the wild type.130Regeneration of such calli into plants may be of benefit to the viticulture industry where E. lata is a problem, if the introduced gene is stably maintained.Toxigenic pathogens are another source of genetic material with the potential for conferring toxin insensitivity, as the pathogens have to avoid poisoning themselves. As mentioned in Section 10, Pseudomonas phaseolicola secretes phaseolotoxin 38, a potent inhibitor of ornithine carbamoyl transferase (OCTase) which catalyses the synthesis of citrulline. However, the bacterium is not affected, as it has two OCT genes, one of which encodes an enzyme, argK, that is resistant to the toxin . When tobacco was transformed with the gene, some of the transformants were insensitive to the toxin .131
As mentioned previously (Section 12), an isolate of Pantoea dispersa, detoxified albicidins, which are produced by Xanthomonas albilineans, the causal agent of sugar cane leaf scald, and was an effective biocontrol agent.126 The gene responsible was cloned, and transgenic sugarcane plants, in which it was expressed, were generated. When expression was at levels of 1–10 ng of enzyme per mg of leaf protein, plants infected with pathogen did not develop the chlorotic symptoms typical of the disease, whereas all untransformed control plants were severely affected. Furthermore, transgenic plants with high activity of the enzyme in young stems were also protected against systemic multiplication of the pathogen.132 Therefore it seems that the expression of this gene confers resistance not only to disease symptoms, but also to multiplication of the toxigenic pathogen within its host.
13.3 Weed control
In 1993 it was pointed out that although natural products had made a large impact on the development of insecticides, there have been few successes with these products as herbicides, a situation that still appears to be the same today.133 The reasons suggested for this situation include the molecular complexity of the compounds, their limited stability and their low herbicidal activity. However, conventional herbicide discovery efforts based on “synthesise and screen” strategies have also had limited success, but advances in chemistry and biotechnology have increased the speed and ease with which natural products can be discovered and developed as herbicides.133Phosphinothricin121 is one success.134 This compound, which is known as glufosinate when synthetic, is a product of Streptomyces viridochromogenes, and is environmentally and toxicologically benign. It is also a degradation product in target plants of Bialaphos. Bialaphos is a product of S. hygroscopicus, and in 1995 was the only commercial herbicide produced by biosynthesis.134 One application of Bialaphos controls red raspberry (Rubus idaeus) in spruce plantations.
Although AAL toxins 19 and 20 are specific for tomato plants with the asc/ascgenotype , they are also toxic to jimsonweed (aka thorn-apple, Datura stramonium) and black nightshade (Solanum nigrum) at 1.56 µg ml–1 and 0.01 µg ml–1, respectively.135 The related compound fumonisin B121 was also effective, all three compounds probably exerting their effect by causing the accumulation of sphingolipid intermediates.136 However, the fumonisins were originally isolated on the basis of their carcinogenicity. In a search for analogues with high phytotoxicity and low mammalian toxicity, the best compound was analogue 9 122, which had ICs of 50–100 µM for mouse fibroblasts, but caused cellular leakage and loss of chlorophyll from tomato with the asc/ascgenotype and black nightshade leaf discs at 10 µM.137

A toxin produced by an isolate of Alternaria alternata was suggested to play an important role in a disease of water hyacinth (Eichhornia crassipes), one of the world's most important weeds.138 Unfortunately, the compound does not appear to have been chemically defined. In contrast, maculosin112 was isolated from cultures of Alternaria alternata, and was demonstrated to be specific for spotted knapweed, Centaurea maculosa.139
Septoria cirsii is specific to Canada thistle (aka creeping thistle, Cirsium arvense), causing chlorosis and necrosis of the leaves. In culture the fungus produces β-nitropropionic acid123, a known phytotoxin which causes the same symptoms in leaves as the pathogen and, in addition, is inhibitory to seed germination and root elongation.140 The fungus and its toxin would appear to be promising candidates for the biocontrol of Canada thistle.

Rather similar work has been done with a pathotype of Drechslera avenae, which appeared to be specific for wild oats (Avena sterilis). The fungus produced pyrenophorol124 in culture, and the compound was toxic to A. sterilis at a concentration of 320 µM but considerably less toxic at this concentration to A. fatua.Pyrenophorol at 640 M did not cause symptoms in any of the other monocotyledons or dicotyledons tested, with the exception of tomato (Lycopersicon esculentum).141 Rather similar results were obtained with pyrenophorin125 from the same fungus.142 The rather low activity of these compounds and the comparative lack of specificity of pyrenophorin would seem to preclude their use as herbicides, although it may prove possible to synthesise analogues with higher activity and the desired specificity.

Ascochyta caulina is another fungus with apparent specificity for a weed host, Chenopodium album. The fungus produces toxins in culture, and the main one has been identified as ascaulitoxin126. Results from field experiments with the fungus did not justify its development as a bioherbicide, probably owing to its low virulence.143 However, addition of the toxins to spores of the fungus did improve control in greenhouse experiments.144

Thaxtomins 5 and 6 have been considered as possible herbicides, but were considered to have insufficient systemic toxicity to ensure reliable weed control at low doses.145
Witchweed (species of Striga) and Orobanche are parasitic plants which tap into the roots of their hosts and sequester their nutrients and water. Striga is a hemiparasite in that it has chlorophyll and can therefore make some contribution to its own nutrition by photosynthesis. Orobanche is achlorophyllous and therefore a holoparasite, deriving all its nutrients from its host. Both genera produce copious numbers of small seeds which may remain dormant in the soil for >10 years, but germinate on the approach of roots of a susceptible plant, wreaking untold havoc among cereal and legume crops, particularly in Africa and Asia. There is therefore an urgent need to find control measures that are effective and can be applied in these areas. Although a number of pathogens of Striga and Orobanche have been described, there is little information on whether they cause disease through the production of toxins . Rather, attention has been focused on the inhibition of germination of the parasites. In an evaluation of 14 fungal toxins as inhibitors of germination of Striga hermonthica, T-2 toxin105 was the most active, causing 100% inhibition of seed germination at 10–5 M and 19% inhibition at 10–7 M. Deoxynivalenol60 was also active, causing 100% and 69% reduction in germination when assayed at 10–4 and 10–5 M, respectively.146
In tests of 188 isolates of fungi from Egyptian soil, five of one fungus were found which were potent inhibitors of the germination of seeds of Orobanche crenata in dual culture.12 Moreover, filtrates from liquid culture of the fungus inhibited germination even when diluted 1000-fold. As previously mentioned (Section 3), the fungus was identified as Myrothecium verrucaria and the active component in culture filtrates as the macrocylic trichothecene verrucarin A16.12M. verrucaria has been isolated from O. ramosa plants in Southern Italy and may therefore be a pathogen.147 Clearly, if M. verrucaria is to be of value in the control of Orobanche in crop plants, it is imperative that neither verrucarin A nor any other mycotoxin is present in the plant. In fact, a caution against the use of the fungus in biocontrol has been issued for this reason.147 Nevertheless, in experiments with kudzu (Pueraria montana) treated with an isolate of M. verrucaria that produced high concentrations of trichothecenes, including verrucarin A, none was found, suggesting that this may not prove as problematic as anticipated.148
14 References
- H. J. M. Loffler and J. R. Mouris, Neth. J. Plant Pathol., 1992, 98, 107–115 Search PubMed.
- S. S. Alam, J. N. Bilton, A. M. Z. Slawin, D. J. Williams, R. N. Sheppard and R. N. Strange, Phytochemistry, 1989, 28, 2627–2630 CrossRef CAS.
- Y. M. Chen and R. N. Strange, Plant Pathol., 1991, 40, 401–407 CAS.
- L. Zhang, J. Xu and R. G. Birch, J. Appl. Microbiol., 1998, 85, 1023–1028 CAS.
- R. Loria, R. A. Bukhalid, R. A. Creath, R. H. Leiner, M. Olivier and J. C. Steffens, Phytopathology, 1995, 85, 537–541 Search PubMed.
- M. J. Babcock, E. C. Eckwall and J. L. Schottel, J. Gen. Microbiol., 1993, 139, 1579–1586 CAS.
- A. Lauzier, C. Goyer, L. Ruest, R. Brzezinski, D. L. Crawford and C. Beaulieu, Can. J. Microbiol., 2002, 48, 359–364 CrossRef CAS.
- M. Solfrizzo, A. De Girolamo, C. Vitti, K. Tylkowska, J. Grabarkiewicz-Szczesna, D. Szopinska and H. Dorna, Food Addit. Contam., 2005, 22, 302–308 CrossRef CAS.
- I. P. Budde, B. H. Rohde, C. L. Bender and M. S. Ullrich, J. Bacteriol., 1998, 180, 1360–1367 CAS.
- I. P. Budde and M. S. Ullrich, Mol. Plant–Microbe Interact., 2000, 13, 951–961 Search PubMed.
- L. P. Partida-Martinez and C. Hertweck, Nature, 2005, 437, 884–888 CrossRef CAS.
- R. El Kassas, Z. K. El Din, M. H. Beale, J. L. Ward and R. N. Strange, Weed Res., 2005, 45, 212–219 CrossRef CAS.
- T. Amagasa, R. N. Paul, J. J. Heitholt and S. O. Duke, Pestic. Biochem. Physiol., 1994, 49, 37–52 CrossRef CAS.
- H. K. Abbas, S. O. Duke, A. J. Merrill, E. Wang and W. T. Shier, Phytochemistry, 1998, 47, 1509–1514 CrossRef CAS.
- H. K. Abbas, B. B. Johnson, W. T. Shier, H. Tak, B. B. Jarvis and C. D. Boyette, Phytochemistry, 2002, 59, 309–313 CrossRef CAS.
- H. Batoko, J. Bouharmont and H. Maraite, Euphytica, 1994, 76, 139–143 Search PubMed.
- H. Batoko, A. D. d'Exaerde, J. M. Kinet, J. Bouharmont, R. A. Gage, H. Maraite and M. Boutry, Biochim. Biophys. Acta, 1998, 1372, 216–226 CAS.
- G. L. Hartman, Y. H. Huang and S. Li, Australas. Plant Pathol., 2004, 33, 9–15 Search PubMed.
- T. Fukushima, M. Tanaka, M. Gohbara and T. Fujimori, Phytochemistry, 1998, 48, 625–630 CrossRef CAS.
- M. Miyashita, T. Nakamori, T. Murai, T. Yonemoto, H. Miyagawa, M. Akamatsu and T. Ueno, Z. Naturforsch., C: Biosci., 2001, 56, 1029–1037 CAS.
- M. Miyashita, T. Nakamori, H. Miyagawa, M. Akamatsu and T. Ueno, Biosci., Biotechnol., Biochem., 2003, 67, 635–638 CrossRef CAS.
- W. J. Bowyer, L. Ning, L. S. Daley, G. A. Strobel, G. E. Edwards and J. B. Callis, Spectroscopy, 1998, 13, 36–+ Search PubMed.
- J. Soukupova, S. Smatanova, L. Nedbal and A. Jegorov, Physiol. Plant., 2003, 118, 399–405 CrossRef CAS.
- I. A. Dubery, D. Meyer and C. Bothma, Phytochemistry, 1994, 35, 307–312 CrossRef CAS.
- M. S. C. Pedras and C. J. Biesenthal, Plant Cell Rep., 2000, 19, 1135–1138 CrossRef CAS.
- M. S. C. Pedras, C. J. Biesenthal and I. L. Zaharia, Plant Sci., 2000, 156, 185–192 CrossRef CAS.
- Y. M. Chen, E. K. Peh and R. N. Strange, Bioseparation, 1991, 2, 107–113 CAS.
- Y. F. Zhao, W. T. Jones, P. Sutherland, D. A. Palmer, R. E. Mitchell, P. H. S. Reynolds, J. P. Damicone and C. L. Bender, Physiol. Mol. Plant Pathol., 2001, 58, 247–258 Search PubMed.
- J. Lydon and C. D. Patterson, Lett. Appl. Microbiol., 2001, 32, 166–170 CrossRef CAS.
- Y. F. Zhao, J. P. Damicone and C. L. Bender, Plant Dis., 2002, 86, 883–888 Search PubMed.
- L. Buchwaldt and J. S. Jensen, Phytochemistry, 1991, 30, 2311–2316 CrossRef CAS.
- H. K. Abbas and R. F. Vesonder, Toxicon, 1993, 31, 355–358 CrossRef CAS.
- K. Hamid and R. N. Strange, Int. Chickpea Newsl., 1997, 4, 20–22 Search PubMed.
- L. H. Zhang, J. L. Xu and R. G. Birch, Microbiology (Reading, U. K.), 1998, 144, 555–559 CrossRef CAS.
- J. Moyroud, J. Gelin, A. Chene and J. Mortier, Tetrahedron, 1996, 52, 8525–8534 CrossRef CAS.
- G. Schuler, H. Gorls and W. Boland, Eur. J. Org. Chem., 2001, 1663–1668 CrossRef CAS.
- R. Lauchli and W. Boland, Chem. Rec., 2003, 3, 12–21 CrossRef CAS.
- D. A. Cuppels and T. Ainsworth, Appl. Environ. Microbiol., 1995, 61, 3530–3536 CAS.
- C. Bender, D. Palmer, A. Penaloza-Vazquez, V. Rangaswamy and M. Ullrich, Arch. Microbiol., 1996, 166, 71–75 CrossRef CAS.
- C. L. Bender, H. Liyanage, D. Palmer, M. Ullrich, S. Young and R. Mitchell, Gene, 1993, 133, 31–38 CrossRef CAS.
- F. J. Alarcon-Chaidez, A. Penaloza-Vazquez, M. Ullrich and C. L. Bender, Plasmid, 1999, 42, 210–220 CrossRef CAS.
- D. M. Brooks, G. Hernandez-Guzman, A. P. Kloek, F. Alarcon-Chaidez, A. Sreedharan, V. Rangaswamy, A. Penaloza-Vazquez, C. L. Bender and B. N. Kunkel, Mol. Plant–Microbe Interact., 2004, 17, 162–174 Search PubMed.
- R. J. Parry, S. V. Mhaskar, M. T. Lin, A. E. Walker and R. Mafoti, Can. J. Chem., 1994, 72, 86–99 CAS.
- S. Jiralerspong, V. Rangaswamy, C. L. Bender and R. J. Parry, Gene, 2001, 270, 191–200 CrossRef CAS.
- C. N. Penfold, C. L. Bender and J. G. Turner, Gene, 1996, 183, 167–173 CrossRef CAS.
- V. Rangaswamy, R. Mitchell, M. Ullrich and C. Bender, J. Bacteriol., 1998, 180, 3330–3338 CAS.
- R. Couch, S. E. O'Connor, H. Seidle, C. T. Walsh and R. Parry, J. Bacteriol., 2004, 186, 35–42 CrossRef CAS.
- H. Liyanage, C. Penfold, J. Turner and C. L. Bender, Gene, 1995, 153, 17–23 CrossRef CAS.
- A. Penaloza-Vazquez and C. L. Bender, J. Bacteriol., 1998, 180, 6252–6259 CAS.
- A. V. Smirnova, L. Wang, B. Rohde, I. Budde, H. Weingart and M. S. Ullrich, J. Mol. Microbiol. Biotechnology, 2002, 4, 191–196 Search PubMed.
- A. V. Smirnova and M. S. Ullrich, Microbiology (Reading, U. K.), 2004, 150, 2715–2726 CrossRef CAS.
- H. Weingart, S. Stubner, A. Schenk and M. S. Ullrich, Mol. Plant–Microbe Interact., 2004, 17, 1095–1102 Search PubMed.
- J. D. Walton, E. D. Earle and B. W. Gibson, Biochem. Biophys. Res. Commun., 1982, 107, 785–794 CrossRef CAS.
- J. B. Rasmussen and R. P. Scheffer, Plant Physiol., 1988, 86, 187–191 CrossRef CAS.
- D. G. Panaccione, J. S. Scott-Craig, J. A. Pocard and J. D. Walton, Proc. Natl. Acad. Sci. U. S. A., 1992, 89, 6590–6594 CrossRef CAS.
- S. R. Canada and L. D. Dunkle, Appl. Environ. Microbiol., 1997, 63, 996–1001 CAS.
- J. H. Ahn and J. D. Walton, Mol. Plant–Microbe Interact., 1997, 10, 207–214 Search PubMed.
- Y. Q. Cheng, L. D. Le, J. D. Walton and K. D. Bishop, J. Nat. Prod., 1999, 62, 143–145 CrossRef CAS.
- J. H. Ahn and J. D. Walton, Mol. Gen. Genet., 1998, 260, 462–469 CAS.
- M. J. Jones and L. D. Dunkle, Mol. Plant–Microbe Interact., 1995, 8, 476–479 Search PubMed.
- I. Weiergang, L. D. Dunkle, K. V. Wood and R. L. Nicholson, Fungal Genet. Biol., 1996, 20, 74–78 CrossRef CAS.
- A. N. Nikolskaya, D. G. Panaccione and J. D. Walton, Gene, 1995, 165, 207–211 CrossRef CAS.
- A. Masunaka, K. Ohtani, T. L. Peever, L. W. Timmer, T. Tsuge, M. Yamamoto, H. Yamamoto and K. Akimitsu, Phytopathology, 2005, 95, 241–247 Search PubMed.
- A. Ichihara, H. Tazaki and S. Sakamura, Tetrahedron Lett., 1983, 24, 5373–5376 CrossRef CAS.
- F. Pinkerton and G. Strobel, Proc. Natl. Acad. Sci. U. S. A., 1976, 73, 4007–4011 CrossRef CAS.
- Y. Y. Mo and D. C. Gross, J. Bacteriol., 1991, 173, 5784–5792 CAS.
- N. B. Quigley and D. C. Gross, Mol. Plant–Microbe Interact., 1994, 7, 78–90 Search PubMed.
- Y. Y. Mo, M. Geibel, R. F. Bonsall and D. C. Gross, Plant Physiol., 1995, 107, 603–612 CAS.
- X. Z. Li, A. N. Starratt and D. A. Cuppels, Phytopathology, 1998, 88, 1094–1100 Search PubMed.
- J. Beausejour, C. Goyer, J. Vachon and C. Beaulieu, Can. J. Microbiol., 1999, 45, 764–768 CrossRef CAS.
- J. Cui, A. K. Bahrami, E. G. Pringle, G. Hernandez-Guzman, C. L. Bender, N. E. Pierce and F. M. Ausubel, Proc. Natl. Acad. Sci. U. S. A., 2005, 102, 1791–1796 CrossRef CAS.
- Y. F. Zhao, R. Thilmony, C. L. Bender, A. Schaller, S. Y. He and G. A. Howe, Plant J., 2003, 36, 485–499 CrossRef CAS.
- C. H. Lawrence, M. C. Clark and R. R. King, Phytopathology, 1990, 80, 606–608 Search PubMed.
- R. R. King, C. H. Lawrence and M. C. Clark, Am. Potato J., 1991, 68, 675–680 Search PubMed.
- R. Loria, R. A. Bukhalid, B. A. Fry and R. R. King, Plant Dis., 1997, 81, 836–846 Search PubMed.
- L. L. Kinkel, J. H. Bowers, K. Shimizu, E. C. Neeno-Eckwall and J. L. Schottel, Can. J. Microbiol., 1998, 44, 768–776 CrossRef CAS.
- C. Goyer, J. Vachon and C. Beaulieu, Phytopathology, 1998, 88, 442–445 Search PubMed.
- M. Lemmens, U. Scholz, F. Berthiller, C. Dall'Asta, A. Koutnik, R. Schuhmacher, G. Adam, H. Buerstmayr, A. Mesterhazy, R. Krska and P. Ruckenbauer, Mol. Plant–Microbe Interact., 2005, 18, 1318–1324 Search PubMed.
- O. D. Hensens, G. L. Helms, E. T. T. Jones and G. H. Harris, J. Org. Chem., 1995, 60, 1772–1776 CrossRef CAS.
- S. M. Mandala, R. A. Thornton, B. R. Frommer, J. E. Curotto, W. Rozdilsky, M. B. Kurtz, R. A. Giacobbe, G. F. Bills, M. A. Cabello, I. Martin, F. Pelaez and G. H. Harris, J. Antibiot., 1995, 48, 349–356 CAS.
- H. Wang, J. Li, R. M. Bostock and D. G. Gilchrist, Plant Cell, 1996, 8, 375–391 CrossRef CAS.
- J. B. Rasmussen and R. P. Scheffer, Physiol. Mol. Plant Pathol., 1988, 32, 283–291 Search PubMed.
- S. D. Kim, H. W. Knoche and L. D. Dunkle, Physiol. Mol. Plant Pathol., 1987, 30, 433–440 Search PubMed.
- G. Brosch, R. Ramsom, T. Lechner, J. D. Walton and P. Loidl, Plant Cell, 1995, 7, 1941–1950 CrossRef CAS.
- J. D. Walton, Plant Cell, 1996, 8, 1723–1733 CrossRef CAS.
- J. G. Turner, Physiol. Mol. Plant Pathol., 1986, 29, 59–68 Search PubMed.
- H. Sawada, T. Takeuchi and I. Matsuda, Appl. Environ. Microbiol., 1997, 63, 282–288 CAS.
- S. S. Patil, A. C. Hayward and R. Emmons, Phytopathology, 1974, 64, 590–595 Search PubMed.
- J. G. Turner and R. E. Mitchell, Plant Physiol., 1985, 79, 468–473 CrossRef CAS.
- D. S. Achor, S. Nemec and R. A. Baker, Mycopathologia, 1993, 123, 117–126 CrossRef CAS.
- G. Agner, Y. A. Kaulin, P. A. Gurnev, Z. Szabo, L. V. Schagina, J. Y. Takemoto and K. Blasko, Bioelectrochemistry, 2000, 52, 161–167 CrossRef CAS.
- K. Blasko, L. V. Schagina, G. Agner, Y. A. Kaulin and J. Y. Takemoto, Biochim. Biophys. Acta, 1998, 1373, 163–169 CAS.
- D. DiGiorgio, L. Camoni, K. A. Mott, J. Y. Takemoto and A. Ballio, Plant Pathol., 1996, 45, 564–571 CrossRef CAS.
- M. L. Hutchison, M. A. Tester and D. C. Gross, Mol. Plant–Microbe Interact., 1995, 8, 610–620 Search PubMed.
- M. L. Hutchison and D. C. Gross, Mol. Plant–Microbe Interact., 1997, 10, 347–354 Search PubMed.
- B. Mezzetti, R. Capasso, A. Evidente, F. A. Hammerschlag, R. H. Zimmerman, G. Cristinzio and P. Rosati, J. Phytopathol., 1994, 142, 219–226 CrossRef CAS.
- K. Nomura, M. Melotto and S. Y. He, Curr. Opin. Plant Biol., 2005, 8, 361–368 CrossRef CAS.
- S. Mittal and K. R. Davis, Mol. Plant–Microbe Interact., 1995, 8, 165–171 Search PubMed.
- X. Li, H. Lin, W. Zhang, Y. Zou, J. Zhang, X. Tang and J. M. Zhou, Proc. Natl. Acad. Sci. U. S. A., 2005, 102, 12990–12995 CrossRef CAS.
- M. Briquet, D. Vilret, P. Goblet, M. Mesa and M. C. Eloy, J. Bioenerg. Biomembr., 1998, 30, 285–295 CrossRef CAS.
- M. Vurro and B. E. Ellis, Plant Sci., 1997, 126, 29–38 CrossRef CAS.
- R. R. King and C. H. Lawrence, J. Agric. Food Chem., 1996, 44, 1108–1110 CrossRef CAS.
- H. Nakajima and R. P. Scheffer, Phytochemistry, 1987, 26, 1607–1611 CrossRef CAS.
- R. E. Mitchell, Phytochemistry, 1991, 30, 3917–3920 CrossRef CAS.
- P. Teyrulh, I. Philippe, J. M. Renaud, G. Tsoupras, P. Deangelis, J. Fallot and R. Tabacchi, Phytochemistry, 1991, 30, 471–473 CrossRef CAS.
- L. R. Smith, N. Mahoney and R. J. Molyneux, J. Nat. Prod., 2003, 66, 169–176 CrossRef CAS.
- L. Sparapano, G. Bruno, O. Fierro and A. Evidente, Phytochemistry, 2004, 65, 189–198 CrossRef CAS.
- T. Tanaka, H. K. Abbas and S. O. Duke, Phytochemistry, 1993, 33, 779–785 CrossRef CAS.
- O. Rocha, K. Ansari and F. M. Doohan, Food Addit. Contam., 2005, 22, 369–378 CrossRef CAS.
- N. J. Alexander, S. P. Mccormick and T. M. Hohn, Mol. Gen. Genet., 1999, 261, 977–984 CrossRef CAS.
- J. W. Pitkin, D. G. Panaccione and J. D. Walton, Microbiology (Reading, U. K.), 1996, 142, 1557–1565 CrossRef CAS.
- T. J. Knight, A. Chatterton and P. J. Langstonunkefer, Fed. Proc., 1985, 44, 1803 Search PubMed.
- K. Lopez-Lopez, J. L. Hernandez-Flores, M. Cruz-Aguilar and A. Alvarez-Morales, J. Bacteriol., 2004, 186, 146–153 CrossRef CAS.
- C. C. Sollod, A. E. Jenns and M. E. Daub, Appl. Environ. Microbiol., 1992, 58, 444–449 CAS.
- W. R. Scheible, B. Fry, A. Kochevenko, D. Schindelasch, L. Zimmerli, S. Somerville, R. Loria and C. R. Somerville, Plant Cell, 2003, 15, 1781–1794 CrossRef CAS.
- B. F. Brandwagt, L. A. Mesbah, F. L. W. Takken, P. L. Laurent, T. J. A. Kneppers, J. Hille and H. J. J. Nijkamp, Proc. Natl. Acad. Sci. U. S. A., 2000, 97, 4961–4966 CrossRef CAS.
- B. F. Brandwagt, T. J. A. Kneppers, H. J. J. Nijkamp and J. Hille, Mol. Plant–Microbe Interact., 2002, 15, 35–42 Search PubMed.
- S. H. Park, A. Stierle and G. A. Strobel, Phytochemistry, 1994, 35, 101–106 CAS.
- I. A. Acuna, G. A. Strobel, B. J. Jacobsen and D. L. Corsini, Plant Sci., 2001, 161, 77–88 CrossRef CAS.
- G. Lazarovits, J. Hill, R. R. King and L. A. Calhoun, Can. J. Microbiol., 2004, 50, 121–126 CrossRef CAS.
- M. Zweimuller, S. Antus, T. Kovacs and J. Sonnenbichler, Biol. Chem., 1997, 378, 915–921 CrossRef CAS.
- M. S. C. Pedras, I. L. Zaharia, Y. Gai, Y. Zhou and D. E. Ward, Proc. Natl. Acad. Sci. U. S. A., 2001, 98, 747–752 CrossRef CAS.
- R. B. Meeley and J. D. Walton, Plant Physiol., 1991, 97, 1080–1086 CrossRef CAS.
- R. B. Meeley, G. S. Johal, S. P. Briggs and J. D. Walton, Plant Cell, 1992, 4, 71–77 CrossRef CAS.
- D. S. Multani, R. B. Meeley, A. H. Paterson, J. Gray, S. P. Briggs and G. S. Johal, Proc. Natl. Acad. Sci. U. S. A., 1998, 95, 1686–1691 CrossRef CAS.
- L. Zhang and R. G. Birch, Lett. Appl. Microbiol., 1996, 22, 132–136 CrossRef CAS.
- M. Buiatti and D. S. Ingram, Experientia, 1991, 47, 811–819 CrossRef CAS.
- A. Gentile, E. Tribulato, G. Continella and A. Vardi, Theor. Appl. Genet., 1992, 83, 759–764 CAS.
- O. Borras, R. Santos, A. P. Matos, R. S. Cabral and M. Arzola, Plant Breed., 2001, 120, 435–438 CrossRef.
- P. Guillen, M. Guis, G. Martinez-Reina, S. Colrat, S. Dalmayrac, C. Deswarte, M. Bouzayen, J. P. Roustan, J. Fallot, J. C. Pech and A. Latche, Plant J., 1998, 16, 335–343 CrossRef CAS.
- E. Hatziloukas and N. J. Panopoulos, J. Bacteriol., 1992, 174, 5895–5909 CAS.
- L. H. Zhang, J. L. Xu and R. G. Birch, Nat. Biotechnol., 1999, 17, 1021–1024 CrossRef CAS.
- S. O. Duke and J. Lydon, ACS Symp. Ser., 1993, 524, 110–124 CAS.
- S. O. Duke and H. K. Abbas, Allelopathy, 1995, 582, 348–362 Search PubMed.
- H. K. Abbas, C. D. Boyette, R. E. Hoagland and R. F. Vesonder, Weed Sci., 1991, 39, 673–677 Search PubMed.
- H. K. Abbas, S. O. Duke and T. Tanaka, J. Toxicol., Toxin Rev., 1993, 12, 225–251 Search PubMed.
- H. K. Abbas, T. Tanaka and W. T. Shier, Phytochemistry, 1995, 40, 1681–1689 CrossRef CAS.
- R. M. Babu, A. Sajeena and K. Seetharaman, Crop Prot., 2003, 22, 1005–1013 CrossRef.
- A. C. Stierle, J. H. Cardellina and G. A. Strobel, Proc. Natl. Acad. Sci. U. S. A., 1988, 85, 8008–8011 CAS.
- J. Hershenhorn, M. Vurro, M. C. Zonno, A. Stierle and G. Strobel, Plant Sci., 1993, 94, 227–234 CrossRef CAS.
- M. A. Kastanias and M. Chrysayi-Tokousbalides, Pest Manage. Sci., 2000, 56, 227–232 Search PubMed.
- M. A. Kastanias and M. Chrysayi-Tokousbalides, J. Agric. Food Chem., 2005, 53, 5943–5947 CrossRef CAS.
- J. Netland, L. C. Dutton, M. P. Greaves, M. Baldwin, M. Vurro, A. Evidente, G. Einhorn, P. C. Scheepens and L. W. French, BioControl, 2001, 46, 175–196 Search PubMed.
- M. Vurro, M. C. Zonno, A. Evidente, A. Andolfi and P. Montemurro, Biol. Control, 2001, 21, 182–190 CrossRef CAS.
- R. R. King, C. H. Lawrence and J. A. Gray, J. Agric. Food Chem., 2001, 49, 2298–2301 CrossRef CAS.
- M. C. Zonno and M. Vurro, Weed Res., 1999, 39, 15–20 CrossRef CAS.
- K. I. Anderson and S. G. Hallett, Weed Sci., 2004, 52, 623–627 Search PubMed.
- H. K. Abbas, H. Tak, C. D. Boyette, W. T. Shier and B. B. Jarvis, Phytochemistry, 2001, 58, 269–276 CrossRef CAS.
| This journal is © The Royal Society of Chemistry 2007 |