
The synthesis and comparison of chondroitin sulfate-modified PDMAEMA with chondroitin sulfate-modified PEI as a potential gene delivery vector†
Yu-Lun Loa,
Hung-Wei Wanga,
Zi-Xian Liaob and
Li-Fang Wang*ab
aDepartment of Medicinal & Applied Chemistry, College of Life Science, Kaohsiung Medical University, Kaohsiung 807, Taiwan. E-mail: lfwang@kmu.edu.tw; Fax: +886-7-3125339; Tel: +886-7-3121101-2217
bInstitute of Medical Science and Technology, National Sun Yat-Sen University, Kaohsiung 804, Taiwan
First published on 31st March 2016
Abstract
Our previous research has confirmed a chondroitin sulfate-polyethylenimine copolymer (CS-PEI) as a potential gene delivery vector because of its recognition by CD44, which enhances the cellular uptake of CS-PEI/pDNA polyplexes. As poly(N,N-dimethylaminoethyl methacrylate) (PDMAEMA) is also a commonly used non-viral gene delivery vector, a CS-PDMAEMA copolymer was synthesized using a similar method with CS-PEI via Michael addition. The physicochemical properties of CS-PDMAEMA and CS-PEI copolymers were thoroughly characterized. The gel electrophoresis results demonstrate that the weight ratio of CS-PEI/pDNA required to completely encapsulate pDNA is ≥1 while that of CS-PDMAEMA/pDNA is ≥3. The CS-modified cationic polymers show lower cytotoxicity than compared with the unmodified ones. At the same weight ratio, CS-PEI/pDNA has a smaller particle size than CS-PDMAEMA/pDNA. The cellular uptake of CS-modified polyplexes is higher in U87 cells (high CD44 expression) than in 3T3 cells (low CD44 expression). However, the transfection efficiency of CS-modified polyplexes is higher in 3T3 cells than in U87 cells. The contrasting results may be attributed to the variation of cell types. In addition, the high level of asialoglycoprotein receptor (ASGP-R) expressed in 3T3 cells seems beneficial for triggering the lectin receptor-mediated endocytosis and results in high transfection efficiency when compared with U87 cells.
Introduction
Gene therapy shows great potential in the treatment of a wide range of diseases. However, success in clinical trials is limited because of numerous technical barriers.1 A fundamental engineering challenge of gene-based therapy is the development of safe and effective delivery vectors. Both viral vectors and non-viral vectors have been used for systemic delivery in clinical settings. Although viral vectors show excellent transfection efficiency, they cannot be largely applied in clinics because of limited DNA packing cargo size and safety issues.2 Non-viral vectors like cationic polymers have the potential capacity for transferring larger gene sequences and are more easily synthesized. In addition, non-viral vectors have the advantages of low immune response and no pathogenesis.3,4Polyethyleneimine (PEI) is the gold standard for non-viral gene delivery vectors. There are two commercially available PEIs, linear (LPEI, 22 kDa) and branched (BPEI, 25 kDa). Both have been extensively used in gene delivery for their high levels of gene expression.5 PEI has a high density of positively charged amino groups, forming complexes with negatively charged DNA, siRNA, and antisense oligodeoxynucleotides (ODN).6–9 Nevertheless, the positively charged surface characteristics of PEI also lead to its inherent cytotoxicity, nonspecific interactions with non-targeted tissue and blood cells, as well as its self-aggregation with serum proteins.10,11 To address the issue of efficacy and toxicity associated with PEI, numerous alternative polymers have been evaluated for gene delivery, including poly(N,N-dimethylaminoethyl methacrylate) (PDMAEMA), poly(β-amino ester)s, and carbohydrate-based cationic polymers and dendrimers.12 PDMAEMA exhibits a relatively low charge density when compared with PEI due to the presence of a tertiary amine that is approximately 50% protonated at physiological pH.13 However, PDMAEMA is more easily tailored than PEI because its monomer can be polymerized in differently controlled manners like atom transfer radical polymerization (ATRP)14–16 and reversible addition–fragmentation chain transfer (RAFT) polymerization.17,18 In addition, PDMAEMA can be synthesized to produce various macromolecular architectures, including linear, graft, block, star shapes with similar repetitive units or complex branched structures of higher homogeneity.19–21
The development of a gene delivery system on the basis of carbohydrate polymers has progressed in parallel to PEG-decorated ones owing to the prevention of recognition by the reticuloendothelial system (RES).22 The presence of derivable groups on sugar chains enriches the versatility for chemical reactions with different kinds of molecules for targeting specific cells and tissues.23 The diversity in carbohydrate structure also entails the formation of nucleic acid nanoparticles (NPs) with varying biophysical properties24 affecting the efficiency with which the various existing extra- and intracellular barriers in nucleic acid delivery can be overcome.25–28 Moreover, carbohydrate polymers are often described as biodegradable, biocompatible and non-immunogenic, which are attractive properties for pharmaceutical and biomedical applications. Thus, carbohydrate polymers are often employed in hybrid nanostructures to reduce the toxicity of synthetic materials such as PEI and PDMAEMA.29–34
Chondroitin sulfate (CS) is a natural anionic polysaccharide with good biocompatibility, biodegradability, bioactivity, low cytotoxicity, and non-allergenicity. CS is an important structural component in connective tissues and cartilage, belonging to the glycosaminoglycan (GAG) family. CS is the copolymer of sulfated D-glucuronic acid at C4 or C6, and N-acetyl-D-galactosamine. The ternary complex of pDNA/PEI and CS showed transgene efficiencies comparable with that of the pDNA/PEI complex, but with minimized cytotoxicity and agglutination of erythrocytes.25 The intracellular uptake of the ternary complex was hypothesized to have been the result of a CS-specific receptor-mediated energy-dependent processes.26 Intraperitoneally injecting PEI carrying a plasmid harboring the murine granulocyte macrophage-colony stimulating factor (mGM-CSF) gene coated with CS yielded 100% mouse survival.35
To solve the problem of strong electrostatic interactions between CS and PEI before being complexed with pDNA,25 CS-PEI was synthesized via Michael addition using the amino groups of PEI and the double bonds of methacrylated CS.36 Preparing CS-PEI via Michael addition also saved the carboxyl groups of CS to permit recognition by CD44.37 CD44 is a type I transmembrane glycoprotein participating in many cellular functions, such as cell orientation, adhesion, migration, and matrix-cell signaling processes.38 CD44 overexpresses in many solid tumors;39 thus, any NPs containing CS enhances the therapeutic outcome of drugs to cancer diseases because of the enhanced cellular uptake via CD44-mediated endocytosis.
Our previous study has utilized different degrees of methacylated CS as a crosslinker to obtain a sol-type PDMAEMA for gene delivery. The combination of CS and PDMAEMA showed lower cytotoxicity and better transfection efficiency than PDMAEMA.40 Nevertheless, the yield of the sol-type PDMAEMA was low. PDMAEMA having a similar molecular weight to that of PEI (10 kDa) with a thiol group was designed (HS-PDMAEMA), which was utilized to conduct the Michael addition with methacrylated CS to yield CS-PDMAEMA, following the same technique for preparing CS-PEI. The merits of this study not only solve the low yield of the sol-type PDMAEMA but also facilitate the same chemical structure of CS-PDMAEMA and CS-PEI for better comparison. The physicochemical properties of CS-PEI and CS-PDMAEMA copolymers were thoroughly characterized. The comparative performances of CS-PEI and CS-PDMAEMA as a gene delivery carrier, including DNA binding ability, cytotoxicity, transfection efficiency, and internalization pathways of their complexes with pDNA were studied.
Experimental
Materials
Sodium chondroitin sulfate (CS) was purchased from Tohoku Miyagi Pharmaceutical Co., Ltd. (Tokyo, Japan). Methacrylic anhydride and polyethylenimine (PEI, 10 kDa) were from Alfa Aesar (Ward Hill, MA) and used as received. Bis(2-hydroxyethyl) disulfide, 2,2′-bipyridyl, copper(I) bromide, 1,6-hexanedioldimethyl, fluorescein isothiocyanate (FITC), 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide (MTT), and deuterated chloroform (CDCl3) were purchased from Sigma (St. Louis, MO). Amberlite® IR120, 2-bromo-2-methylpropionyl bromide, 2-(dimethylamino)ethyl methacrylate (DMAEMA), and rhodamine 123, were purchased from Acros (Morris Plains, NJ). Aluminum oxide neutral (Al2O3) was from Seedchem Company PTY. LTD (Melbourne, Australia). Dithiothreitol (DTT) was purchased from J.T. Baker (Center Valley, PA). Ethidium bromide (EtBr) was purchased from MP Biomedicals (Verona, Italy). The reporter genes pEGFP-C1 and pGL3-Control were purchased from Clontech (Palo Alto, CA) and Promega (Madison, WI), respectively. Fetal bovine serum (FBS) was purchased from Biological Industries (Beit Haemek, Israel). Agarose, Minimum Essential Medium (MEM) and Dulbecco's Modified Eagle's Medium (DMEM) were purchased from Invitrogen (Carlsbad, CA). Potassium dihydrogen phosphate, disodium hydrogen phosphate, glycine, boric acid, and hydrochloric acid were purchased from Fluka (Buchs, Switzerland) and used for buffer preparation. All other unstated chemicals were purchased from Sigma-Aldrich and used without further purification.Synthesis of bis[2-(2′-bromoisobutyryloxy)ethyl]disulfide macroinitiator (iBuBr–SS–iBuBr)
An iBuBr–SS–iBuBr compound was prepared as previously published.41,42 In brief, bis(2-hydroxyethyl)disulfide (1 g, 6.48 mmol) was dissolved in 20 mL of tetrahydrofuran (THF) containing 2.7 mL of triethylamine (TEA) in a two-neck round-bottom flask. After six consecutive standard freeze–pump–thaw cycles to remove oxygen, 2-bromo-2-methylpropionyl bromide (2.4 mL, 19.42 mmol) was added dropwise into the above solution at 0 °C. The resulting solution was stirred overnight at room temperature (RT). Next, THF was removed by rotary evaporation and the product was purified by silica column chromatograph with a mixture of hexane/ethyl acetate (1/13 v/v) as the eluent. The product was isolated by rotary evaporation and further dried in vacuum at 50 °C. The yield of the oily product was 2.23 g (∼78%).Synthesis of PDMAEMA–SS–PDMAEMA and HS-PDMAEMA
The initiator iBuBr–SS–iBuBr (40.0 mg, 0.088 mmol) dissolved in 2-propanol (1.6 mL) and the monomer DMAEMA (4.4 mL, 26.1 mmol) in double-deionized (DD) water (0.4 mL) were added into a two-neck round-bottom flask, followed by six standard freeze–pump–thaw cycles. The catalyst CuBr (27.2 mg, 0.18 mmol) and 2,2′-bipyridyl (12.5 mg, 0.088 mmol) were added into the flask in the frozen condition, followed by two additional freeze–pump–thaw cycles to further remove oxygen. The reaction was carried out at 0 °C under argon for 6 h. The crude product was dissolved in DD water and purified by dialysis using a dialysis membrane (Spectra/Por MWCO 1000; Spectrum Laboratories, Rancho Dominguez, CA) against DD water to remove the unreacted monomer. The lyophilized product was dissolved in THF and passed through an aluminum oxide column and Amberlite® IR120 to remove the residual copper catalyst. Finally, the product was obtained from precipitation into excess n-hexane and dried under vacuum with a yield of 25.2 g (63%).The disulfide linkages of PDMAEMA–SS–PDMAEMA were reduced by DTT. PDMAEMA–SS– PDMAEMA (1 mg mL−1) and DTT (1 mg mL−1) at the weight ratio of 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 in DD water was stirred at RT for 24 h. The solution was dialyzed against DD water using a dialysis membrane (MWCO 1k). The final HS-PDMAEMA product was obtained by freeze-drying.
:1 in DD water was stirred at RT for 24 h. The solution was dialyzed against DD water using a dialysis membrane (MWCO 1k). The final HS-PDMAEMA product was obtained by freeze-drying.
Synthesis of CS-modified PDMAEMA (CS-PDMAEMA) and CS-modifies PEI (CS-PEI)
Methacrylated CS (CSMA) was synthesized as previously described.43 The degree of methacrylation on CS was controlled at ∼70%. HS-PDMAEMA/PEI-10k was grafted onto CSMA via Michael addition as performed in previous research.36 Briefly, CSMA (10 mg) and HS-PDMAEMA/PEI (300 mg) were respectively dissolved in DD water at a concentration of 1 mg mL−1. The CSMA solution was slowly added into the HS-PDMAEMA/PEI solution, followed by stirring at RT for 24 h. The product was purified by dialysis using a dialysis membrane (MWCO 25k) to remove the unreacted HS-PDMAEMA/PEI. The final product was obtained by freeze-drying with a yield of 224.87 mg (∼73%).Synthesis of rhodamine 123-conjugated CS-PDMAEMA and CS-PEI
CS-PDMAEMA/CS-PEI (10 mg) and N-(3-dimethylaminopropyl)-N′-ethylcarbodiimide hydrochloride (EDAC) (2 mg) were dissolved in 2 mL DD water. The solution was stirred at RT for 1 h. Rhodamine 123 (Rh123) (1 mg in 1 mL DD water) was added to the above solution and stirred at RT for 24 h. The solution was dialyzed against DD water using an MWCO 1k membrane. All steps were performed in the dark and the product was collected by freeze-drying.Characterization
1H-NMR spectra were recorded using a Varian Mercury plus-200 (200 MHz) NMR spectrometer in D2O or CDCl3 at a concentration of 10 mg mL−1. The molecular weight and molecular weight distribution of polymers were measured by gel permeation chromatograph (GPC) using an Agilent 1100 series equipped with a Shodex KF-803 column for the organic phase or with a Shodex Sugar KS-804 column for the aqueous phase. THF or DD water was used as an eluent at a flow rate of 1 mL min−1, respectively. Ten polystyrene or 5 dextran monodisperse standards were used to generate a calibration curve. The buffer capacity was analyzed by acid–base titration. Each 20 mg sample was added to 20 mL of 150 mM NaCl, and the pH was adjusted to 9.5 using 0.1 N NaOH. The titration was done at 25.0 ± 0.1 °C by adding 0.1 N HCl under an atmosphere of CO2-free N2.Preparation and characterization of polymer/pDNA polyplexes
PEGFP-C1 plasmids inserted with a cDNA sequence encoding enhanced green fluorescent protein (eGFP) driven by a cytomegalovirus (CMV) promoter and pGL3-control plasmid with a Hind III/Xba I firefly luciferase cDNA fragment, cloned into the pCDNA vector, were introduced into the E. coli strain DH5α and purified using a kit. CS-PEI/CS-PDMAEMA was dissolved in DD water to a final concentration of 1 mg mL−1. The pH value of solution was adjusted to 7. Plasmid DNA concentration was fixed at 4 μg/40 μL in DD water to measure DNA binding and at 8 μg/500 μL for other measurements. Equal volumes of polymers and pDNA solutions were then mixed and immediately vortexed at a high speed for 60 s. The DNA binding ability of polyplexes was evaluated at different weight ratios. The stability of polyplexes with 10% FBS was analyzed using gel electrophoresis with 0.8% agarose in tris–acetate–EDTA (TAE) with EtBr (1 μg mL−1). A current of 100 V was applied to gels for 40 min and DNA retention was visualized under UV illumination at 365 nm. To measure the resistance of polyplexes to DNase I, one unit of the enzyme was mixed with polyplexes and incubated for 1 h. The enzyme was inactivated by adding 150 mM EDTA and incubating at 65 °C for 30 min. Subsequently, a 25% (w/v) heparin solution was used to release DNA from polyplexes. The resistance capacity of polyplexes was evaluated using agarose gel electrophoresis.The hydrodynamic diameter and zeta potential of polyplexes were measured using a Zetasizer Nano ZS instrument (Malvern, Worcestershire, UK). Light scattering measurements were done with a laser at 633 nm and a 90° scattering angle. The particle size and zeta potential of each polyplex was measured three times. The size and morphology of polyplexes were also observed using a transmission electron microscope (TEM, Jeol TEM-1200; Tokyo, Japan). A carbon-coated 200-mesh copper specimen grid was glow-discharged for 1.5 min. Polyplexes (10 μL) were deposited on a TEM grid and allowed to dry for three days at RT before examining the samples by TEM. Images were observed under an acceleration voltage of 60 kV and captured with a CCD camera.
Cytotoxicity and transfection
U87 cells (a human glioblastoma cell line) and 3T3 cells (a mouse fibroblast cell line) were cultivated at 37 °C under humidified 5% CO2 in MEM and DMEM, respectively, supplemented with 10% FBS and 100 μg mL−1 penicillin–streptomycin. The cells were seeded in 96-well tissue culture plates at a density of 5 × 103 per well in the medium containing 10% FBS. The cytotoxicity of copolymers was evaluated by determining cell viability after 24 h of incubation with various concentrations (1–200 μg of polymer per mL). The number of viable cells was determined by estimating their mitochondrial reductase activity using the tetrazolium-based colorimetric method (MTT conversion test).44In vitro transgene expression was performed at a density of 1 × 105 per well in 12-well plates and incubated in the medium containing 10% FBS for 24 h before transfection. Polyplexes with a weight ratio (polymers/pDNA) of 1–20 were prepared using different amounts of polymers and a fixed pDNA amount of 8 μg to a final volume of 1000 μL. Following 2 or 4 h of incubation with 10% FBS, the medium was replaced with 1 mL of fresh complete medium, and the cells were incubated for 72 h post-transfection. The GFP expression was directly visualized using a fluorescence microscope.
For the luciferase assay, the procedures stated above were repeated to determine the transfection efficiency in U87 cells and 3T3 cells. To quantify the luciferase expression, the transfected cells were gently rinsed twice with 1 mL of 0.1 M PBS, added to a 200 μL per well of 1× Glo lysis buffer (Promega, Madison, WI), and allowed to stand overnight at −20 °C. The luciferase activity was monitored using a microplate scintillation and luminescence counter after mixing the contents of a 50 μL per well of supernatant with the contents of 50 μL per well of luciferase assay reagent. The total protein content of the cell lysate was examined using a BCA protein assay kit and measured according to the manufacturer's instructions. Transfection efficiency was expressed as the relative fluorescence unit per mg protein (RLU per mg protein). PEI-25k and Lipofectamine 2000 (Lipo-2000) were used as positive controls.
Transfection with or without proton pump inhibitor, bafilomycin A1
To evaluate the proton-sponge effect of the cationic polymers, a proton pump inhibitor, bafilomycin A1, on polycation-mediated transfection efficiency was tested.45 U87/3T3 cells were transfected by CS-PEI/pDNA or CS-PDMAEMA/pDNA polyplexes at a weight ratio of 7 with or without bafilomycin A1. The cells were seeded at a density of 1 × 105 per well in 12-well plates and incubated in MEM/DMEM medium containing 10% FBS for 24 h before transfection. When the cells were at 50–70% confluence, the culture medium was replaced with 1 mL of fresh medium containing 200 nM bafilomycin A1 and incubated for 20 min. Following 20 min of incubation, the medium containing the inhibitor was changed to fresh medium with 10% FBS, and the cells were further treated with CS-PEI/pDNA or CS-PDMAEMA/pDNA and incubated for another 4 h. PEI(25k)/pDNA at the N/P ratio of 10 was performed as a positive control. Luciferase expression was assayed after 48 h transfection.Cellular uptake and transfection in presence of endocytosis inhibitors
To understand the endocytosis pathways of the polyplexes, three inhibitors and one competitor were tested.46,47 U87/3T3 cells were seeded at a density of 1.5 × 105 per well in 12-well plates in MEM/DMEM supplemented with 10% FBS and incubated for 24 h. The cells were pre-incubated with various concentrations ranging 5–20 μg mL−1 for chlorpromazine, 100–400 nM for genistein, 25–100 nM for wortmannin, and 0.25–1 mg mL−1 for CS. Following 30 min of incubation, the medium containing an inhibitor was changed to fresh medium and the polymer/pDNA polyplex at a weight ratio of 7 was treated and incubated for another 4 h. Next, the cells were trypsinized, centrifuged and re-suspended in 1 mL of cold PBS, and the luciferase expression of polyplexes was analyzed as aforementioned.The same procedure as stated above was repeated using a single dose of inhibitors: 10 μg mL−1 chlorpromazine, 200 μM genistein, 50 nM wortmannin, and 100 μg mL−1 CS, respectively, to determine the inhibition of cellular uptake in the cells incubated using a flow cytometer.
Asialoglycoprotein and CD44 receptor expression on the surface of cells
To determine the expression level of the asialoglycoprotein receptor (ASGP-R), U87/3T3 cells were harvested and lysed by RIPA buffer (G-biosciences, St. Louis, MO). The protein content of the cell lysate was determined using a BCA protein assay kit (Thermo Fisher Scientific Inc., Rockford, IL). An equal amount of protein (40 μg) was separated using 10% SDS-PAGE gel and transferred to a PVDF membrane (Merck Millipore Life Science, Darmstadt, Germany). The PVDF membrane was blocked in blocking buffer [5% nonfat milk powder dissolved in tris-buffered saline buffer containing 0.1% Tween 20 (TBST)] at RT. Following 1 h of incubation, the blocked PVDF membrane was probed with a 1:1000 dilution of anti-asialoglycoprotein receptor antibody (Abcam, Cambridge, MA) in 1% nonfat milk–TBST buffer at 4 °C overnight. The next day, the PVDF membrane was washed three times with TBST and incubated with 1:10000 dilution of a HRP-conjugated secondary antibody (Leadgene biomedical, Tainan city, Taiwan) in TBST at RT for 1 h. The PVDF membrane was washed three times with TBST and developed using an enhanced chemiluminescence (ECL) detection system. To determine the expression level of CD44, an FITC conjugate anti-CD44 antibody against CD44 receptor assay was employed to measure the expression of CD44 receptor in U87/3T3 cells. The cells were trypsinized and then washed in PBS buffer. The cells (3 × 105) were then resuspended in PBS buffer and incubated with anti-human/mouse CD44 FITC antibody for 30 min on ice. The CD44 antibody concentration was prepared according to the manufacturer's instructions. After incubation, the cells were washed three times with PBS, re-suspended in 1 mL of PBS buffer, and analyzed using a flow cytometer.
Statistical methods
Means and standard deviations (SD) of data were calculated. Differences between groups were tested using Student's t-test and P < 0.05 was considered significant.Results and discussion
Preparation of CS-PEI and CS-PDMAEMA
CS-PEI was synthesized using Michael addition between the amino groups of PEI and the methacrylated groups of CSMA as previously reported.36 The 1H-NMR spectrum of CSMA (Fig. S1A, ESI†) shows two distinctive proton peaks of the double bond at δ 5.58 and 6.15 ppm (peaks a & b), the methyl groups adjacent to the double bond of MA at δ 1.93 ppm (peak c), and the methyl group of acetylamide of CS at δ 2.02 ppm (peak d). From the peak intensity ratio between peak (c) and peak (d), the degree of methacrylation of CSMA was estimated to be ∼70%. To synthesize CS-PDMAEMA using a similar method with CS-PEI via Michael addition, an iBuBr–SS–iBuBr macro-initiator was first prepared41,42 to yield a symmetrical PDMAEMA–SS–PDMAEMA diblock copolymer via ATRP (Scheme 1). The 1H-NMR spectrum of iBuBr–SS–iBuBr shows two distinctive proton peaks at δ 3.02 and 4.45 ppm (peaks e and f), attributable to the methylene units of 2-hydroxyethyl disulfide and the three protons at δ 1.98 ppm (peak g), attributable to the methyl group of α-bromoisobutyryl bromide (Fig. S1B†). The intensity ratio among the three peaks of e:f:g was exactly 1:1:3, implying the successful synthesis of iBuBr–SS–iBuBr.
 | ||
| Scheme 1 Synthesis flow charts of iBuBr–SS–iBuBr, PDMAEMA–SS–PDMAEMA, and HS-PDMAEMA. | ||
PDMAEMA–SS–PDMAEMA was obtained using iBuBr–SS–iBuBr as an initiator and DMAEMA as a monomer via ATRP. Fig. 1A shows the 1H-NMR assignment of PDMAEMA–SS–PDMAEMA, where the characteristic peaks of DMAEMA were observed at δ 2.28 (a, N–CH3), 2.57 (b, N–CH2), 4.08 (c, O–CH2), 1.82 (C–CH2), and 0.91 ppm (C–CH3), respectively. Next, the disulfide linkages of PDMAEMA–SS–PDMAEMA were reduced by DTT to yield HS-PDMAEMA (Scheme 1). The number average molecular weights measured by GPC were 14800 Da for PDMAEMA–SS–PDMAEMA and 8100 Da for HS-PDMAEMA with polydispersity indexes (PDI) of 1.55 and 1.33, respectively (Fig. S2†). This was the optimized molecular weight of HS-PDMAEMA and obtained to approximate that of PEI. CS-PDMAEMA was obtained by reacting the thiol groups of HS-PDMAEMA and the methacrylated groups of CSMA via Michael addition, similar to the preparation of CS-PEI. Fig. 1B is the 1H-NMR spectrum of CS-PDMAEMA where the peaks of PDMAEMA became wider and no new peaks appeared. Thus, GPC was explored to check the molecular weight. As clearly seen in Fig. 1C, the GPC profiles of CS-PEI and CS-PDMAEMA shifted to the left, as compared with that of CS, indicating an increase in molecular weight. The number average molecular weights were 53700 Da for CS, 89000 Da for CS-PEI, and 77000 Da for CS-PDMAEMA with a narrow PDI of ∼1.05. The lower molecular weight of CS-PDMAEMA compared with that of CS-PEI was reasonable because the parent molecular weight of PEI (10 kDa) was relatively higher and many amino groups of the branched PEI could attack the double bonds of CSMA.
 | ||
| Fig. 1 1H-NMR spectra of (A) PDMAEMA–SS–PDMAEMA in chloroform-d and (B) CS-PDMAEMA in deuterium water; (C) GPC profiles of CS, CS-PEI, and CS-PDMAEMA using DD water as an eluent. | ||
Acid/base titration profiles were generated to determine the proton buffering capacity of the polymers. Solutions were adjusted to ∼ pH 9.5 using 0.1 N NaOH and titrated using 0.1 N HCl (Fig. S3†). CS-PDMAEMA had a buffer capacity close to that of HS-PDMAEMA because a similar amount of HCl was utilized to change the pH from 5.1 to 7.4 (0.56 mL for CS-PDMAEMA and 0.60 mL for HS-PDMAEMA), which has been defined as a measurement of buffer capacity.48 In contrast, the buffer capacity of CS-PEI (0.93 mL) was higher than that of PEI-10k (0.69 mL) and PEI-25k (0.73 mL). Owing to the high density of amines, PEI leads itself to protonation, with the charge density proportional to the pH of the biological environment.12 This buffering capacity enables PEI polyplexes to avoid lysosomal trafficking and degradation once inside cells. It can be presumed from the same fact that CS-PEI should have a better proton sponge effect than CS-PDMAEMA.
Characterization of polymers/pDNA polyplexes
Independent of a w/w ratio of polymers/pDNA ranging 1–9, the hydrodynamic diameters were within 73–100 nm for CS-PDMAEMA/pDNA and within 59–76 nm for CS-PEI/pDNA, with a narrow size distribution (PDI < 0.3) (Fig. 2A). CS-PEI/pDNA had smaller sizes than CS-PDMAEMA/pDNA. Xu et al. had prepared PEI nanogels with mean diameters of 38, 75, 87, 121, 132, and 167 nm and showed the highest transfection efficiency for polyplexes with mean diameters of 75 and 87 nm,49,50 suggesting that the optimal size for gene transfer of non-targeting cationic vector-DNA polyplexes is between 70 and 90 nm.50 Particle size is a major factor for deciding the extravasation rate of nanoparticles from the bloodstream, as well as recognition by the RES; hence, nanoparticle sizes of 10–200 nm are preferred for in vivo delivery.51 The sizes of CS-PEI/pDNA and CS-PDMAEMA/pDNA were within 50–100 nm, which could be hypothesized to prevent recognition by the RES, thus benefitting gene transfection. | ||
| Fig. 2 (A) Hydrodynamic diameters and (B) zeta potentials of polymers/pDNA polyplexes in DD Water. The concentration of test samples was 0.1 mg mL−1 and the temperature was 25 °C. The data were means ± SD of an experiment done in triplicate. | ||
The zeta potentials of the polymers/pDNA were all positive and increased from 5 eV at w/w = 1 to 15 eV at w/w = 9 (Fig. 2B). The zeta potentials of CS-PEI/pDNA were larger than those of CS-PDMAEMA/pDNA when compared at the same w/w ratio. This difference is because PEI bears the primary, secondary, and ternary amino groups while PDMAEMA bears only the ternary amino groups.12 The morphologies of polyplexes were visualized using TEM (Fig. S4†). Both polyplexes had a spherical shape and their sizes were close to those measured by DLS. In agreement with the DLS result, CS-PEI/pDNA had smaller sizes than CS-PDMAEMA/pDNA.
The binding ability of polymers and pDNA was studied using an agarose gel electrophoresis retardation assay. The result illustrates that pDNA was well complexed by CS-PEI at a w/w ratio of >1 and by CS-PDMAEMA at a w/w ratio of >3 (Fig. 3A). To verify the protection of pDNA against DNase I digestion, the electrophoretic mobility analysis of each polyplex was tested after it had been left standing in 1U DNase I at RT for 1 h. A competition reagent of heparin was explored to release pDNA from polyplexes. As shown in Fig. 3B, the DNA remained intact in both polyplexes at a w/w ratio of ≥3 and degraded at the w/w ratio of 1 after 1 h of incubation with DNase I. Thus, DNA was well protected by CS-PEI as well as CS-PDMAEMA if a polyplex was prepared at a weight ratio of ≥3.
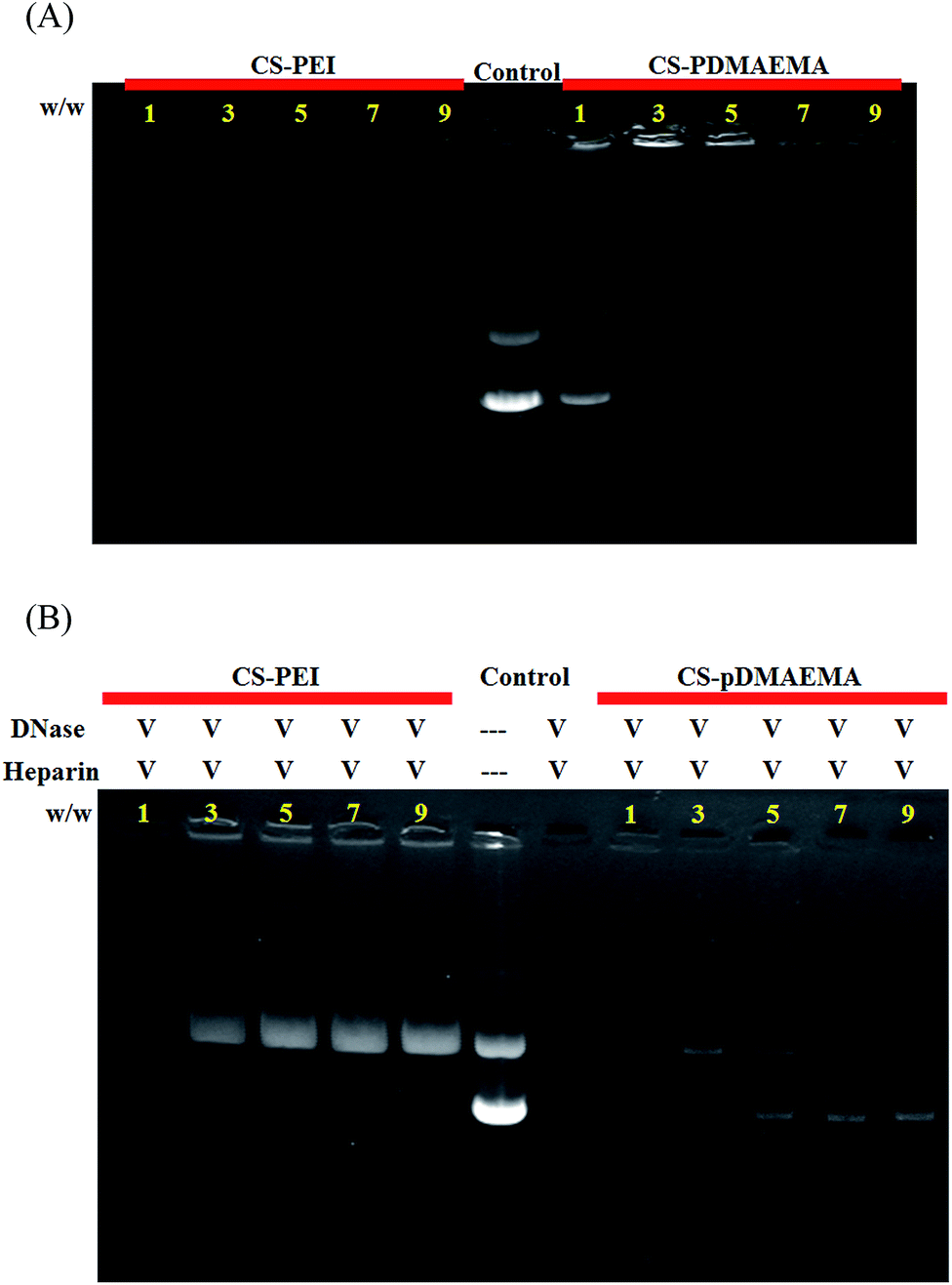 | ||
| Fig. 3 Agarose gel electrophoresis to test (A) pDNA retention and (B) DNase I digestion assay of polyplexes prepared at various w/w ratios. Polyplexes were incubated with 1U DNase I, followed by treatment with 25% heparin to release pDNA from polyplexes. Naked pDNA was used as a reference and the numeral of each graph indicates a weight ratio of polymers and pDNA. | ||
In vitro gene transfection
Cell viability decreased dramatically with PEI-10k and HS-PDMAEMA at a concentration of ≥20 μg mL−1 against 3T3 and U87 cells, but with CS-PEI and CS-PDMAEMA, cell viability remained ≥80% at a concentration of 20 μg mL−1 and was significantly higher than that with PEI-10k and HS-PDMAEMA (Fig. S5†).Plasmids pEGFP-C1 and pGL3-control were utilized to test the transfection efficiency of polymers as a non-viral gene vector. To evaluate the gene expression, 3T3 and U87 cells were exposed to polymers/pDNA at a w/w ratio of 1–20 with 10% FBS. A commercial Lipo-2000 and a gold standard of the non-viral gene vectors, PEI-25k, were used as positive controls. PEGFP-C1 was qualitatively used for efficient GFP expression (Fig. S6†) and the pGL3-control plasmid was quantitatively used for luciferase expression (Fig. 4).
 | ||
| Fig. 4 Luciferase activity of pGL3-control expression normalized with protein amounts containing 10% FBS in (A, B) 3T3 cells and (C, D) U87 cells as a function of polymers/pDNA weight ratios for (A, C) 2 h and (B, D) 4 h of incubation followed by 72 h of post-transfection. (n = 3, *P < 0.05, **P < 0.01). | ||
In 3T3 cells, the transfection efficiency of CS-PDMAEMA/pDNA consistently increased with an increasing w/w ratio up to 15 at both 2 and 4 h of incubation. Transfection efficiency at 4 h was higher than that at 2 h and comparable with that of Lipo-2000 and PEI-25k. The transfection efficiency of CS-PEI/pDNA was significantly higher than that of CS-PDMAEMA at 2 h of incubation. At w/w = 20, the decreased transfection efficiency was presumably due to the cytotoxicity caused by the high polymer concentration used (40 μg mL−1, Fig. S6A and B†). A similar result of transfection efficiency of CS-PEI/pDNA and CS-PDMAEMA/pDNA was observed in U87 cells (Fig. S6C and D†). However, the transfection efficiency was much lower in U87 cells than in 3T3 cells and CS-PEI/pDNA still had a higher transfection efficiency than CS-PDMAEMA/pDNA.
In parallel, the quantitative transfection efficiencies of polyplexes were measured and compared with those of PEI-25k/pGL3 and Lipo-2000/pGL3 with 10% FBS as positive controls. In 3T3 cells, the transfection efficiency of CS-PDMAEMA/pDNA increased consistently with increasing w/w ratio at 2 h of incubation and reached a plateau up to w/w = 5 at 4 h of incubation (Fig. 4A and B). The transfection efficiency at 4 h was higher than that at 2 h. Nevertheless, the transfection efficiency of CS-PEI/pDNA was close at both 2 h and 4 h of incubation and independent of the w/w ratio. Similar to the result of pEGFP-C1 expression, the transfection efficiency of CS-PEI/pDNA appeared significantly higher with increasing w/w ratio up to 11 at 2 h of incubation but only up to 5 at 4 h of incubation. This difference is presumably due to an increase in the transfection efficiency with incubation time for CS-PDMAEMA/pDNA but not for CS-PEI/pDNA. The transfection efficiency was relatively lower in U87 cells than in 3T3 cells while CS-PEI/pDNA still had significantly higher transfection efficiency than CS-PDMAEMA/pDNA. Though the tendency for transfection efficiency of CS-PEI/pDNA increase was not obvious with w/w ratio, the optimized transfection efficiency of CS-PDMAEMA/pDNA was seen at w/w = 7 (Fig. 4C and D). Thus, this w/w ratio was utilized for subsequent experiments.
According to the proton sponge hypothesis, the buffering capacity of PEI leads to osmotic swelling and rupture of endosomes, resulting in the release of a payload into the cytoplasm. The relatively high transfection efficiency of PEI as a gene vector was attributed to the ability to avoid lysosomal trafficking and degradation.45 Bafilomycin A1 is a specific inhibitor of the vacuolar ATPase proton pump found in early endosomes.45 Bafilomycin A1 inhibits proton transport into endosomes; hence, it should inhibit the protonation of proton sponge vectors inside endosomes and prevent endosomal escape, leading to reduced transfection efficiency. Therefore, 3T3 and U87 cells were transfected with polymers/pDNA at the weight ratio of 7 and with PEI-25k/pDNA at the N/P ratio of 10 with or without the pretreatment of bafilomycin A1 for testing the proton sponge effect. As seen in Fig. 5, the use of bafilomycin A1 significantly reduced the transfection efficiency of PEI and CS-PEI by approximately one order of magnitude in 3T3 cells and by approximately two orders of magnitude in U87 cells. In CS-PDMAEMA, the reduced transfection efficiency was relatively small in the presence of bafilomycin A1 as compared with CS-PEI. This result clearly evidenced the higher proton sponge effect of CS-PEI.
 | ||
| Fig. 5 Effect of proton pump inhibitors on luciferase activity of pGL3-control expression normalized with protein amounts containing 10% FBS in (A) 3T3 cells and (B) U87 cells at a polymers/pDNA weight ratio of 7 with or without bafilomycin A1 (a vacuolar proton pump inhibitor). PEI-25k/pDNA at and N/P ratio of 10 was performed as a positive control (n = 3, *P < 0.05, **P < 0.01). | ||
Cellular uptake and luciferase transfections with inhibitors
Prior to treating cells with Rh123–polymers/pDNA with 10% FBS for 4 h of incubation, all cells were treated with three chemical inhibitors or one competitor without 10% FBS for 30 min. Wortmannin was selected for macropinocytosis; chlorpromazine, for clathrin-mediated endocytosis (CME); genistein, for caveolae-mediated endocytosis; and CS, for CD44, respectively.52 The inhibition effects of the inhibitors and competitor on the cellular uptake of the polyplex at w/w = 7 were tested using the following concentrations: 10 μg mL−1 chlorpromazine, 50 nM wortmannin, 200 μM genistein, and 100 μg mL−1 CS. From the flow cytometric diagrams of Rh123–CS-PDMAEMA/pDNA, uptake inhibition was observed when 3T3 cells were pretreated with genistein and slightly with CS, but not with chlorpromazine and wortmannin as compared with the control group (Fig. 6A). In the Rh123–CS-PEI/pDNA polyplex, the flow cytometry diagram shifted slightly to the left when 3T3 cells were pretreated with wortmannin and CS (Fig. 6B). This left shift indicated the blocking uptake of the polyplex. Thus, the inhibition effect of the cellular uptake of CS-PDMAEMA/pDNA into 3T3 cells was presumed to involve caveolae-mediated endocytosis; however, in CS-PEI/pDNA, a macropinocytosis-mediated endocytosis pathway was involved. In regards to the pretreatment with CS, the left shift of the flow cytometry diagram of the polyplexes was too small to draw any inhibition conclusions. In contrast to the results for 3T3 cells, the flow cytometry diagrams of U87 cells pretreated with chlorpromazine and CS shifted remarkably to the left while those of cells pretreated with genistein and wortmannin remained intact (Fig. 6C and D). It can be concluded that clathrin- and CD44-mediated endocytosis are two possible pathways involved in the internalization of CS-PDMAEMA/pDNA as well as CS-PEI/pDNA into U87 cells.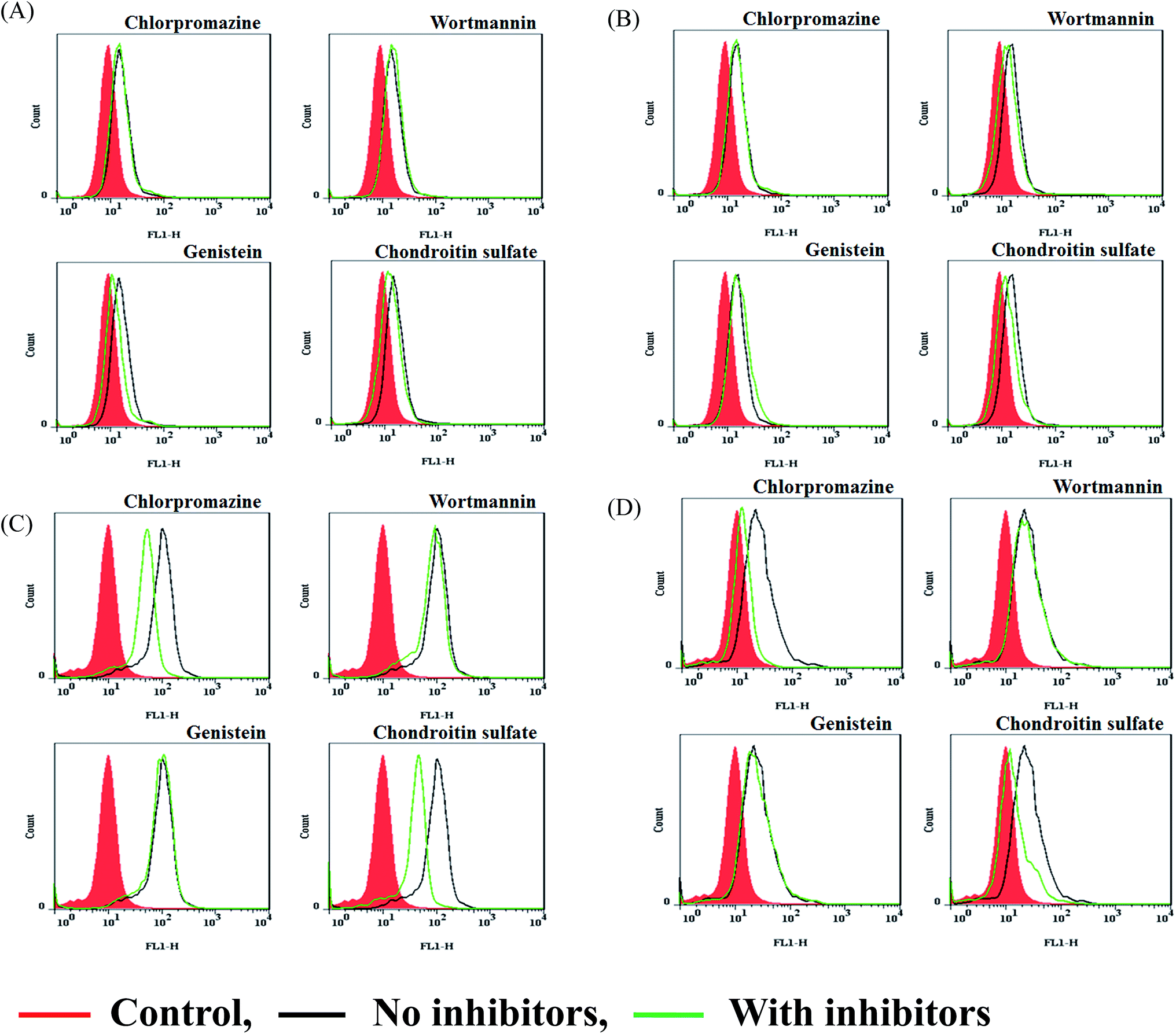 | ||
| Fig. 6 Flow cytometric diagrams of the inhibition of intracellular uptake of Rh123-linked polymers/pGL3 at a weight ratio of 7 containing 10% FBS. (A, B) 3T3 cells and (C, D) U87 cells had been pretreated with inhibitors (10 μg mL−1 chlorpromazine, 200 nM genistein, 50 nM wortmannin, and 100 μg mL−1 CS) before the cells were exposed to (A, C) CS-PDMAEMA/pDNA and (B, D) CS-PEI/pDNA. | ||
The dose-dependent inhibition in the use of the three chemical inhibitors or one competitor on gene expression was further tested. The cytotoxicities of the four chemical reagents were tested against 3T3 and U87 cells (Fig. S7†). The cell viabilities of wortmannin, genistein, and CS were ≥80% in any test concentration, but decreased dramatically with chlorpromazine at 10 μg mL−1 (∼50%). Comparing the cytotoxicities of various reagents revealed that chlorpromazine was very toxic against 3T3 and U87 cells. Subsequently, the percentage of gene expression in the cells pretreated with or without an inhibitor was measured. In 3T3 cells, the percentages of gene expression of the cells exposed to CS-PDMAEMA/pDNA decreased consistently with an increase in the concentration of chlorpromazine, wortmannin, and CS, but remarkably reduced to 40% when treated with 100 μM genistein (Fig. 7A). A similar trend of inhibition for CS-PEI/pDNA was observed in the cells pretreated with chlorpromazine, wortmannin, and CS. However, the percentage of gene expression in the cells pretreated with genistein decreased slightly with an increase in concentration (Fig. 7B). In U87 cells, the inhibition effect for both CS-PDMAEMA/pDNA and CS-PEI/pDNA was remarkable in the cells pretreated with chlorpromazine and CS (Fig. 7C and D). The inhibition effect might occur due to the cytotoxicity of inhibitors. Herein, the reduced percentages of gene expression in the cells exposed to inhibitors were plotted at the non-toxic concentration and the significance of inhibition was compared with the control group at the following concentrations: 5 μg mL−1 chlorpromazine, 100 nM wortmannin, 400 μM genistein, and 1 mg mL−1 CS. Significant inhibition of gene expression using CS-PDMAEMA and CS-PEI as a gene vector appeared in 3T3 cells pretreated with any of the four chemical reagents (Fig. S8A and B†). However, in U87 cells, the inhibition effect was observed only in cells pretreated with chlorpromazine, wortmannin, and CS (Fig. S8C and D†).
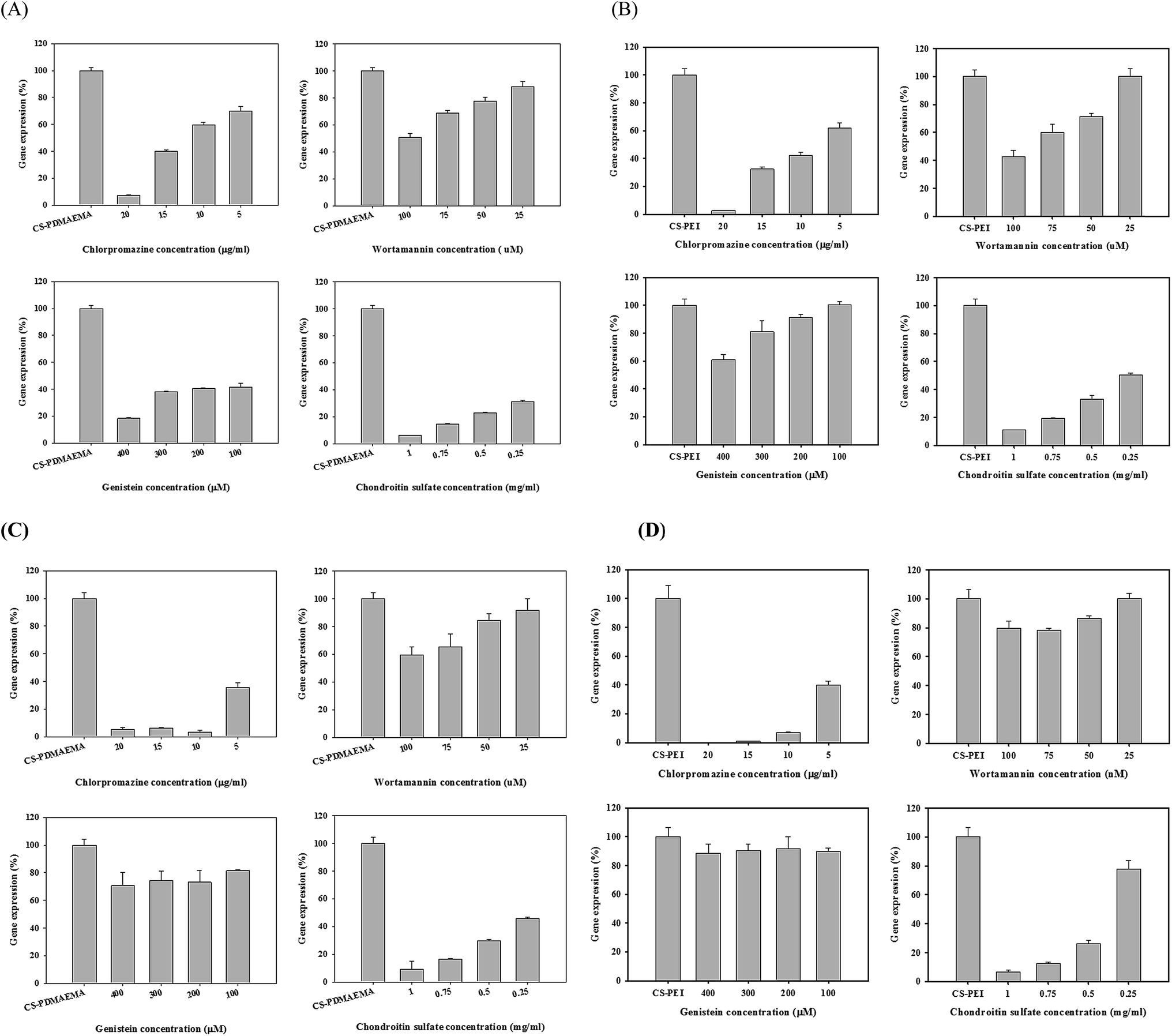 | ||
| Fig. 7 Dose dependence of four chemical inhibitors on the percentage of luciferase expression of pGL3-control containing 10% FBS in (A, B) 3T3 cells and (C, D) U87 cells at the weight ratio of 7 of (A, C) CS-PDMAEMA/pDNA and (B, D) CS-PEI/pDNA (n = 3). | ||
The results are summarized in Fig. 8. Reduced gene expression of more than 40% in 3T3 cells pretreated with any of the four chemical reagents is shown. In particular, cells pretreated with genistein had a more significant decrease in gene expression of CS-PDMAEMA/pDNA than of CS-PEI/pDNA, implying that CS-PDMAEMA/pDNA was more favorable for uptake via caveolae-mediated endocytosis (Fig. 8A). In U87 cells, the percentages of gene expression decreased more than 60% when the cells were pretreated with chlorpromazine and CS (Fig. 8B). This result was in agreement with the findings from the flow cytometry study. The internalization of CS-PDMAEMA/pDNA and CS-PEI/pDNA into U87 cells was mainly via clathrin- and CD44-mediated endocytosis because of a tremendously reduced percentage of gene expression in the use of inhibitors and competitor. Compared with CS-PEI/pDNA, CS-PDMAEMA/pDNA seems more sensitive to the pretreatment of inhibitors, showing highly reduced percentages of gene expression (Fig. 8B). Similarly, comparing the reduced percentages of gene expression (Fig. 8A and B), 3T3 cells are more responsive to chemical inhibitors and competitor than U87 cells.
 | ||
| Fig. 8 Reduced percentages of gene expression in the cells pretreated with inhibitors or a competitor in (A) 3T3 cells and (B) U87 cells (n = 3, *P < 0.05). Prior to treating cells with Rh123–polymers/pDNA with 10% FBS for 4 h of incubation, all cells were treated with inhibitors or a competitor without 10% FBS for 30 min. | ||
Our previous study had confirmed that U87 cells were CD44-overexpressing and 3T3 cells were low-expression cells (Fig. S9†).53 Comparing the flow cytometry diagram of each polyplex entering into these two cells shows that the cellular uptake of the polyplex into 3T3 cells (Fig. 6A and B) was less than that into U87 cells (Fig. 6C and D). To understand why lower CD44-expressing 3T3 cells when pretreated with CS still had an inhibition effect in transfection efficiency, the amount of ASGP-R expression in the two cells were examined using a western blotting assay. The hepatic ASGP-R, which recognizes glycoconjugates containing terminal galactose or N-acetylgalactosamine residues, was the first membrane lectin discovered and has been a classical system for studying receptor-mediated endocytosis before the hyaluronan/chondroitin sulfate receptor was studied as another endocytic membrane lectin.54 Western blotting results revealed higher expression of ASGP-R in 3T3 cells than in U87 cells (Fig. 9).
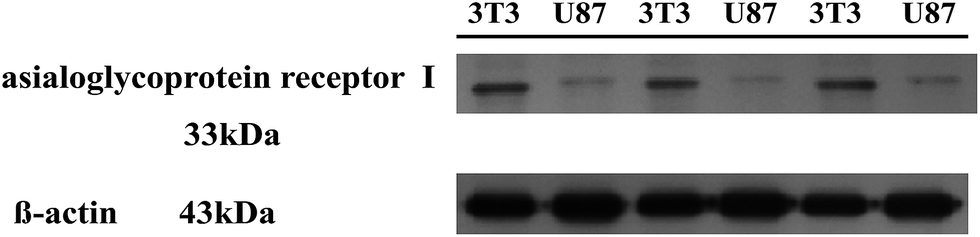 | ||
| Fig. 9 Western blotting analysis of asialoglycoprotein receptor I expression in the cell membrane. β-Actin was used as an internal control. | ||
The amount of CD44 expression in 3T3 cells has been reported to be dependent on serum concentration.55 3T3 cells were found to contain low levels of CD44 and did not bind hyaluronan when grown in medium containing 0.5 or 10% serum. In 5% serum, however, the cells had much higher levels of CD44 and were able to bind hyaluronan. CD44 levels also increased in 3T3 cells re-stimulated with either 5 or 10% serum after prior maintenance in low serum.55 In this study, all transfection studies were performed in the condition with 10% FBS, which may stimulate the expression of CD44 in 3T3 cells. In addition, the uptake of polyplexes varies significantly with the cationic carrier vector, cell type, and amount of cell-surface GAGs present. Transfection efficiency was shown to be inhibited by GAGs, suggesting that cell-surface GAGs probably direct polyplexes into intracellular compartments that do not promote transcription.56 Non-viral gene delivery systems need to overcome several cellular barriers for successful delivery of DNA into the nucleus; hence, the precise mechanism mediating DNA delivery still remains obscure. The internalization pathway depends not only on chemical and structural nature (size, charge, shape, and stability), but also on cell type and cell metabolism.47 The polyelectrolyte complexes of pDNA can undergo multiple structural transitions.57 It is hard to precisely control their characteristics, especially in the cellular environment; however, the high level of ASGP-R expressed in 3T3 cells seems beneficial for triggering the lectin receptor-mediated endocytosis and results in high transfection efficiency when compared with U87 cells (CD44-overexpressing cells).
Conclusions
This study successfully synthesized HS-PDMAEMA, which could be simply grafted onto CSMA to yield CS-PDMAEMA using a Michael addition similar to the preparation of CS-PEI. Higher buffering sponge effect and a smaller size of CS-PEI/pDNA resulted in better transfection efficiency in both 3T3 and U87 cells. Although PDMAEMA exhibits a relatively low buffering sponge effect, it is more easily tailored than PEI because its monomer can be polymerized in differently controlled manners to yield different architectures. Fine tuning the architecture of CS-PDMAEMA to improve transfection efficiency will be the focus of our future study. The cellular uptake of CS-modified polyplexes was higher in U87 cells (high CD44 expression) than in 3T3 cells (low CD44 expression). However, the transfection efficiency of CS-modified polyplexes is higher in 3T3 cells than in U87 cells. The contrasting results may be attributed to a higher expression of ASGP-R in 3T3 cells. Nevertheless, further studies are necessary to investigate other possible mechanisms involved in gene expression besides recognition of ASGP-R in 3T3 cells.Abbreviations
| ASGP-R | Asialoglycoprotein receptor |
| ATRP | Atom transfer radical polymerization |
| iBuBr–SS–iBuBr | Bis[2-(2′-bromoisobutyryloxy)ethyl]disulfide macroinitiator |
| CMV | Cytomegalovirus |
| CME | Clathrin-mediated endocytosis |
| CS | Chondroitin sulfate |
| CS-PEI | Chondroitin sulfate-polyethyleneimine |
| CS-PDMAEMA | Chondroitin sulfate-poly(N,N-dimethylaminoethyl methacrylate) |
| DTT | Dithiothreitol |
| MTT | 3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyltetrazolium bromide |
| DMEM | Dulbecco's modified Eagle's medium |
| DD | Double-deionized water |
| eGFP | Enhanced green fluorescent protein |
| EtBr | Ethidium bromide |
| FITC | Fluorescein isothiocyanate |
| GAG | Glycosaminoglycan |
| Lipo-2000 | Lipofectamine 2000 |
| CSMA | Methacrylated chondroitin sulfate |
| mGM-CSF | Murine granulocyte macrophage-colony stimulating factor |
| EDAC | N-(3-Dimethylaminopropyl)-N′-ethylcarbodiimide hydrochloride |
| NPs | Nanoparticles |
| ODN | Oligodeoxynucleotide |
| PDMAEMA | Poly(N,N-dimethylaminoethyl methacrylate) |
| PEI | Polyethyleneimine |
| RAFT | Reversible addition–fragmentation chain transfer |
| RES | Reticuloendothelial system |
| Rh123 | Rhodamine 123 |
| THF | Tetrahydrofuran |
| TEA | Triethylamine |
| TAE | Tris–acetate–EDTA |
| TBST | Tris-buffered saline buffer containing 0.1% Tween 20 |
Acknowledgements
We are grateful for the financial support from the Ministry of Science and Technology of Taiwan (MOST104-2314-B-037-006-MY3 and MOST103-2320-B-037-012-MY3). This study is also supported by “Aim for the Top Journals Grant (KMU-DT105009)” and by “NSYSU-KMU Joint Research Project, (NSYSUKMU 105-P018)” from Kaohsiung Medical University. We appreciate the experiment support of transmission electron microscope from Center for Research Resources and Development of KMU.References
- S. L. Ginn, I. E. Alexander, M. L. Edelstein, M. R. Abedi and J. Wixon, J. Gene Med., 2013, 15, 65–77 CrossRef CAS PubMed.
- M. Giacca and S. Zacchigna, J. Controlled Release, 2012, 161, 377–388 CrossRef CAS PubMed.
- F. Schlenk, S. Grund and D. Fischer, Ther. Delivery, 2013, 4, 95–113 CrossRef CAS PubMed.
- H. Yin, R. L. Kanasty, A. A. Eltoukhy, A. J. Vegas, J. R. Dorkin and D. G. Anderson, Nat. Rev. Genet., 2014, 15, 541–555 CrossRef CAS PubMed.
- X. Guo and L. Huang, Acc. Chem. Res., 2012, 45, 971–979 CrossRef CAS PubMed.
- M. S. Shim and Y. J. Kwon, Bioconjugate Chem., 2009, 20, 488–499 CrossRef CAS PubMed.
- M. S. Shim and Y. J. Kwon, J. Controlled Release, 2009, 133, 206–213 CrossRef CAS PubMed.
- O. M. Merkel, A. Beyerle, D. Librizzi, A. Pfestroff, T. M. Behr, B. Sproat, P. J. Barth and T. Kissel, Mol. Pharm., 2009, 6, 1246–1260 CrossRef CAS PubMed.
- J. H. Jeong, S. H. Kim, S. W. Kim and T. G. Park, J. Biomater. Sci., Polym. Ed., 2005, 16, 1409–1419 CrossRef CAS PubMed.
- T. Nomoto, Y. Matsumoto, K. Miyata, M. Oba, S. Fukushima, N. Nishiyama, T. Yamasoba and K. Kataoka, J. Controlled Release, 2011, 151, 104–109 CrossRef CAS PubMed.
- M. Ogris, S. Brunner, S. Schuller, R. Kircheis and E. Wagner, Gene Ther., 1999, 6, 595–605 CrossRef CAS PubMed.
- M. A. Mintzer and E. E. Simanek, Chem. Rev., 2009, 109, 259–302 CrossRef CAS PubMed.
- C.-W. Lo, W.-H. Liao, C.-H. Wu, J.-L. Lee, M.-K. Sun, H.-S. Yang, W.-B. Tsai, Y. Chang and W.-S. Chen, Langmuir, 2015, 31, 6130–6136 CrossRef CAS PubMed.
- A. P. Majewski, A. Schallon, V. Jerome, R. Freitag, A. H. Mueller and H. Schmalz, Biomacromolecules, 2012, 13, 857–866 CrossRef PubMed.
- F. Dai, P. Sun, Y. Liu and W. Liu, Biomaterials, 2010, 31, 559–569 CrossRef CAS PubMed.
- F. J. Xu, H. Li, J. Li, Z. Zhang, E. T. Kang and K. G. Neoh, Biomaterials, 2008, 29, 3023–3033 CrossRef CAS PubMed.
- S. Guo, Y. Huang, W. Zhang, W. Wang, T. Wei, D. Lin, J. Xing, L. Deng, Q. Du, Z. Liang, X. J. Liang and A. Dong, Biomaterials, 2011, 32, 4283–4292 CrossRef CAS PubMed.
- C. Zhu, S. Jung, S. Luo, F. Meng, X. Zhu, T. G. Park and Z. Zhong, Biomaterials, 2010, 31, 2408–2416 CrossRef CAS PubMed.
- A. M. Alhoranta, J. K. Lehtinen, A. O. Urtti, S. J. Butcher, V. O. Aseyev and H. J. Tenhu, Biomacromolecules, 2011, 12, 3213–3222 CrossRef CAS PubMed.
- K. M. Xiu, J. J. Yang, N. N. Zhao, J. S. Li and F. J. Xu, Acta Biomater., 2013, 9, 4726–4733 CrossRef CAS PubMed.
- Y.-L. Lo, G.-J. Chen, T.-H. Feng, M.-H. Li and L.-F. Wang, RSC Adv., 2014, 4, 11089 RSC.
- K. Raemdonck, T. F. Martens, K. Braeckmans, J. Demeester and S. C. De Smedt, Adv. Drug Delivery Rev., 2013, 65, 1123–1147 CrossRef CAS PubMed.
- Y. Liu, J. Wu, L. Meng, L. Zhang and X. Lu, J. Biomed. Mater. Res., 2008, 85, 435–443 CrossRef PubMed.
- W. Khan, H. Hosseinkhani, D. Ickowicz, P. D. Hong, D. S. Yu and A. J. Domb, Acta Biomater., 2012, 8, 4224–4232 CrossRef CAS PubMed.
- A. Pathak, P. Kumar, K. Chuttani, S. Jain, A. K. Mishra, S. P. Vyas and K. C. Gupta, ACS Nano, 2009, 3, 1493–1505 CrossRef CAS PubMed.
- T. Kurosaki, T. Kitahara, S. Fumoto, K. Nishida, J. Nakamura, T. Niidome, Y. Kodama, H. Nakagawa, H. To and H. Sasaki, Biomaterials, 2009, 30, 2846–2853 CrossRef CAS PubMed.
- G. Jiang, K. Park, J. Kim, K. S. Kim and S. K. Hahn, Mol. Pharm., 2009, 6, 727–737 CrossRef CAS PubMed.
- S. H. Yu, D. W. Tang, H. Y. Hsieh, W. S. Wu, B. X. Lin, E. Y. Chuang, H. W. Sung and F. L. Mi, Acta Biomater., 2013, 9, 7449–7459 CrossRef CAS PubMed.
- H. Huang, H. Yu, G. Tang, Q. Wang and J. Li, Biomaterials, 2010, 31, 1830–1838 CrossRef CAS PubMed.
- S. K. Tripathi, R. Goyal, P. Kumar and K. C. Gupta, Nanomedicine, 2012, 8, 337–345 CAS.
- F. J. Xu, Z. X. Zhang, Y. Ping, J. Li, E. T. Kang and K. G. Neoh, Biomacromolecules, 2009, 10, 285–293 CrossRef CAS PubMed.
- Z. H. Wang, W. B. Li, J. Ma, G. P. Tang, W. T. Yang and F. J. Xu, Macromolecules, 2011, 44, 230–239 CrossRef CAS.
- Y. Ping, C.-D. Liu, G.-P. Tang, J.-S. Li, J. Li, W.-T. Yang and F.-J. Xu, Adv. Funct. Mater., 2010, 20, 3106–3116 CrossRef CAS.
- J. Yao, Y. Fan, R. Du, J. Zhou, Y. Lu, W. Wang, J. Ren and X. Sun, Biomaterials, 2010, 31, 9357–9365 CrossRef CAS PubMed.
- K. Hamada, C. Yoshihara, T. Ito, K. Tani, M. Tagawa, N. Sakuragawa, H. Itoh and Y. Koyama, J. Gene Med., 2012, 14, 120–127 CrossRef CAS PubMed.
- Y.-L. Lo, H.-L. Chou, Z.-X. Liao, S.-J. Huang, J.-H. Ke, Y.-S. Liu, C.-C. Chiu and L.-F. Wang, Nanoscale, 2015, 7, 8554–8565 RSC.
- D. Hagenfeld, T. Schulz, P. Ehling, T. Budde, U. Schumacher and P. Prehm, J. Cell. Biochem., 2010, 111, 858–864 CrossRef CAS PubMed.
- H. Ponta, L. Sherman and P. A. Herrlich, Nat. Rev. Mol. Cell Biol., 2003, 4, 33–45 CrossRef CAS PubMed.
- D. Naor, R. V. Sionov and D. Ish-Shalom, Adv. Cancer Res., 1997, 71, 241–319 CrossRef CAS PubMed.
- Y. L. Lo, Y. S. Wang and L. F. Wang, Adv. Healthcare Mater., 2013, 2, 1458–1468 CrossRef CAS PubMed.
- T. X. Wang, R. Sridhar, A. Korotcov, A. H. Ting, K. Francis, J. Mitchell and P. C. Wang, Colloids Surf., A, 2011, 375, 147–155 CrossRef CAS PubMed.
- L. G. Bach, M. R. Islam, Y. T. Jeong, Y. S. Gal and K. T. Lim, Appl. Surf. Sci., 2012, 258, 2816–2822 CrossRef CAS.
- L. F. Wang, S. S. Shen and S. C. Lu, Carbohydr. Polym., 2003, 52, 389–396 CrossRef CAS.
- Y. L. Lou, Y. S. Peng, B. H. Chen, L. F. Wang and K. W. Leong, J. Biomed. Mater. Res., Part A, 2009, 88, 1058–1068 CrossRef PubMed.
- A. Akinc, M. Thomas, A. M. Klibanov and R. Langer, J. Gene Med., 2005, 7, 657–663 CrossRef CAS PubMed.
- A. F. Adler and K. W. Leong, Nano Today, 2010, 5, 553–569 CrossRef CAS PubMed.
- G. Sahay, D. Y. Alakhova and A. V. Kabanov, J. Controlled Release, 2010, 145, 182–195 CrossRef CAS PubMed.
- W. C. Tseng, C. H. Tang and T. Y. Fang, J. Gene Med., 2004, 6, 895–905 CrossRef CAS PubMed.
- D. Xu, J. Hong, K. Sheng, L. Dong and S. Yao, Radiat. Phys. Chem., 2007, 76, 1606–1611 CrossRef CAS.
- D.-M. Xu, S.-D. Yao, Y.-B. Liu, K.-L. Sheng, J. Hong, P.-J. Gong and L. Dong, Int. J. Pharm., 2007, 338, 291–296 CrossRef CAS PubMed.
- V. I. Shubayev, T. R. Pisanic II and S. Jin, Adv. Drug Delivery Rev., 2009, 61, 467–477 CrossRef CAS PubMed.
- Y.-S. Liu, C.-C. Chiu, H.-Y. Chen, S.-H. Chen and L.-F. Wang, Mol. Pharm., 2014, 11, 1164–1175 CrossRef CAS PubMed.
- Y.-L. Lo, K.-H. Sung, C.-C. Chiu and L.-F. Wang, Mol. Pharm., 2013, 10, 664–676 CrossRef CAS PubMed.
- P. H. Weigel and J. H. Yik, Biochim. Biophys. Acta, 2002, 1572, 341–363 CrossRef CAS.
- P. Kogerman, M. S. Sy and L. A. Culp, J. Cell. Physiol., 1996, 169, 341–349 CrossRef CAS PubMed.
- M. Ruponen, P. Honkakoski, M. Tammi and A. Urtti, J. Gene Med., 2004, 6, 405–414 CrossRef CAS PubMed.
- A. V. Kabanov and V. A. Kabanov, Bioconjugate Chem., 1995, 6, 7–20 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c6ra01957a |
| This journal is © The Royal Society of Chemistry 2016 |