
Multi-parametric imaging of the invasiveness-permissive acidic microenvironment in human glioma xenografts†
Cuiyun Huanga,
Huan Qinb,
Jun Qiana,
Jingye Zhanga,
Shengyuan Zhaoa,
Yinzhi Changyia,
Bingbing Lib,
Jianping Zhangc,
Jianhua Zhua,
Da Xingb,
Sihua Yang*b and
Cong Li*a
aKey Laboratory of Smart Drug Delivery, Ministry of Education, School of Pharmacy, Fudan University, Shanghai, 201203, China. E-mail: congli@fudan.edu.cn; Fax: +86 21 5198 0001
bKey Laboratory of Laser Life Science and Institute of Laser Life Science, Ministry of Education, College of Biophotonics, South China Normal University, Guangzhou, 510613, China. E-mail: yangsh@scnu.edu.cn; Fax: +86 20 8521 6052
cCenter for Biomedical Imaging, Department of Nuclear Medicine, Fudan University Shanghai Cancer Center, Fudan University, Shanghai, 200032, China
First published on 11th June 2015
Abstract
Gliomas, the intrinsic tumors of the brain, are currently incurable because they are extremely invasive. The infiltrative nature of gliomas makes them difficult to completely excise and leads to a high recurrence rate. Acidification of extracellular microenvironment plays an important role in the invasion, migration and metastasis of solid tumors. To elucidate the relationship between acidic microenvironment and the invasiveness of gliomas, multi-parametric imaging studies were conducted between a highly invasive orthotopic (ortho.) human U87MG glioma xenograft and a lowly invasive subcutaneous (s.c.) U87MG xenograft. In vivo optical imaging showed higher overall acidity of the ortho. tumor than that of the s.c. tumor after administration of a pH responsive near-infrared (NIR) fluorescence probe. Positron emission tomography/computed tomography (PET/CT) imaging demonstrated higher glucose uptake in the ortho. tumor after injection of [18F]-fluorodeoxyglucose (18F-FDG). Photoacoustic microscopy imaging (PAM) revealed a higher vascular density but a more aberrant vessel morphology in the ortho. tumor. Immunofluorescence microscopic imaging indicated significantly up-regulated acidification associated enzymes in the gliomas than in normal brain tissue. This work not only reveals the acidity correlated glioma invasiveness, but also shows the promise for curbing glioma invasiveness by neutralizing intratumoral acidity via down-regulation of glucose uptake, normalizing tumor vasculatures or blocking the acidification associated metabolic/physiological processes.
1. Introduction
Gliomas, the most common and formidable type of primary brain tumor, are currently not curable resulting in a disproportionate level of mortality worldwide.1 For example, despite of substantial treatments such as maximal surgical resection, conformal radiation and systemic chemotherapy, the median overall survival (OS) is only 14.6 months for patients with glioblastomas multiforme (GBM),2 the most aggressive glioma variant. High invasiveness is a dominant reason leading to the poor prognosis of glioma patients. Unlike the normal glial cells, whose proliferation, differentiation, and migration are tightly controlled,3 the transformed glial cells infiltrate into brain tissue so aggressively that “crab claw-like” or “firework-like” spreading patterns are formed along the white matter track, blood vessels and parenchyma.4 Due to the infiltrated and heterogeneous nature of glioma cells, it is extremely difficult to completely eliminate the neoplastic tissue, which leads to high recurrence. Therefore, attenuating the invasiveness of gliomas will be helpful to reduce their high invasiveness and mortality.Acidification of extracellular pH (pHe) is a universal characteristic of solid tumors regardless of genotype and phenotype.5 While the pHe in normal tissues is measured in a range of 7.3–7.4, the value in solid tumors is determined in a range of 6.2–6.9.6,7 Tumor acidification plays an important role in maintaining cancer cell invasiveness6,7 by up-regulating tumorigenic transformation,4 promoting angiogenesis,3 accelerating extracellular matrix (ECM) remodeling,8 resisting chemotherapy9 and suppressing the immune response.10 For example, cancer cells exposed to a low pH in vitro displayed an increased invasive tendency11 and regions of highest tumor invasion usually corresponded to areas with the lowest pH.12 The proton concentration gradient from the tumor to adjacent normal tissue promotes tissue remodeling at the tumor–stroma interface13,14 and increases the activity of proteases such as cathepsin,11 which is actively involved in local invasion. Furthermore, neutralization of intratumoral acidosis with systemic buffers such as bicarbonate inhibited tumor local invasion.15,16 Based on the above experimental results, we hypothesize that the acidic microenvironment plays an important role in maintaining the high invasiveness of gliomas. Evaluating acidity and acidity associated physiological/metabolic processes will help to understand the acidification correlated invasiveness and hence identify potential therapeutic targets for glioma treatment.
To support rapid growth and proliferation, cancer cells metabolize glucose through glycolysis14 instead of oxidative phosphorylation used in normal cells in the presence or absence of oxygen. This phenomenon was first described by Warburg about 80 years ago and named the “Warburg effect”.15 A large amount of lactic acid, as the end product of glycolysis, is believed to be the main reason leading to extracellular acidity. For example, an experimentally induced hyperglycemia acidified tumor pHe from 7.4 to 6.7 coincides with an increase in the glycolytic rate.17 Glycolysis relies on a series of enzymes to metabolize glucose to lactic acid (Fig. 1). They include the facultative glucose transporter 1 (GLUT1) that increases the intracellular glucose uptake rate,18,19 lactate dehydrogenase (LDH) that converts the pyruvate into lactate,20,21 and monocarboxylate transporter 4 (MCT-4) that exports lactate out of the cells.20 Besides glycolysis, tumor acidity is also maintained by the up-regulation of plasma membrane ion transporters such as the Na+/H+ exchanger (NHE)22 that effluxes protons to the extracellular fluid, and carbonic anhydrase 9 (CA9) that accelerates the hydration of extracellular CO2 to H+ and HCO3−.21 Besides the metabolic alternations, tumor acidity has also been reported to result from the poor diffusion rate that prevents the timely drainage of protons.12 Bullitt et al. quantitatively measured vessel attributes including vessel density, radius, tortuosity and terminal branch degree within five types of gliomas by high-resolution magnetic resonance imaging (MRI).23 Compared to normal brains, all glioma patients demonstrated the marked increase in vessel tortuosity and terminal branch count. Therefore, studying fermentative glycolysis, acidification associated enzymes/transporters and vascular morphology help to understand the acidosis associated invasiveness in gliomas.
 | ||
| Fig. 1 Key enzymes/membrane ion channel proteins involved in tumor acidification. Acidification is associated with fermentative glycolysis that produces excess lactic acid, increased activities of the membrane proton transporters such as Na+/H+ exchangers (NHEs) that accelerate the intracellular proton efflux and carbonic anhydrases (CAs) that catalyze hydration of extracellular CO2 to carbonic acid. Up-regulated glycolysis is characterized by enhanced glucose uptake via glucose transporters (GLUTs), conversion of glucose to lactate and H+ via lactate dehydrogenase (LDH) and increased efflux of lactic acid via monocarboxylate transporters (MCTs). | ||
Previous works demonstrated that while orthotopic glioma xenografts showed high invasiveness including widespread dissemination, infiltration along white matter tracts, seeding of the ventricular system and extension along the leptomeninges, subcutaneous xenografts possessing characteristic glioma receptors, such as epidermal growth factor receptor (EGFR), lost their invasiveness by forming a discrete mass with a well-circumscribed border that pushed aside rather than invaded adjacent normal tissue.24 In this work, mouse models inoculated with both ortho. human U87MG glioma xenografts and s.c. U87MG glioma xenografts were developed to investigate the relationship between the acidity and invasiveness of gliomas. Because molecular imaging shows promise in monitoring biological events at molecular and cellular levels with high sensitivity and spatial resolution, multiparametric imaging strategies were used to compare the acidity and acidity associated physiological processes between ortho. and s.c. glioma xenografts. Acidity in tumors was determined by in vivo optical imaging after intravenous (i.v.) administration of a home-made pH responsive near-infrared fluorescence probe DiIRB-S.25 Glucose uptake efficiency was evaluated by in vivo positron emission tomography/computed tomography after i.v. injection of the radioactive glucose derivative [18F]-fluorodeoxyglucose. The vascular density and morphology in tumor margin areas were imaged by in vivo photoacoustic microscopy imaging. Additionally, the expression levels of acidity associated enzymes/transporters were evaluated by ex vivo immunofluorescence microscopic imaging. Overall, this work not only elucidates the relationship between glioma acidity and its invasiveness, but also helps to identify glioma treatment by blocking tumor acidity or acidity associated physiological/metabolic pathways.
2. Results
2.1 pH responsive NIR fluorescence and photoacoustic probe
In a previous study, we developed a pH responsive NIR fluorescence probe DiIRB-S that showed the capability to visualize intratumoral acidosis.26 In this probe, two NIR fluorophores with high photochemical stability and extinction coefficient were conjugated via an acid liable hydrazone bond. Under a neutral pH, the fluorophores formed a face to face H-type aggregate through van der Waals forces and their fluorescence was suppressed efficiently (Scheme 1). The cleavage of acid liable bonds in acidic pHs with concomitant aggregate disruption resulted in substantial fluorescence enhancement. DiIRB-S exhibited two absorption bands at 686 and 756 nm in an aqueous solution, which were assigned to the H-type aggregate and fluorophore monomer27 (Fig. 2A and S1A–C†). Notably, while absorbance at 756 nm (A756) increased consistently at pH 5.5, absorbance at 686 nm (A686) decreased gradually, which indicated the conversion of the aggregate to a monomer in an acidic environment. The time course of the absorbance ratio A756/A686 was further measured at pH 7.4, 6.5 and 5.5 that mimics the environment of normal tissues, tumor extracellular fluid and lysosomal compartments, respectively. The hydrolytic velocities of the aggregates under pH 5.5 and 6.5 were about 52.6 and 13.2 times faster than that under pH 7.4, respectively (Fig. 2B). Fluorospectroscopy studies demonstrated that the fluorescence intensity of DiIRB-S centered at 778 nm increased as many as 22.6 times in the first 2 h after incubation at pH 5.5 (Fig. 2C and S1D–F†). The time dependent fluorescence intensity of DiIRB-S indicated that the signal enhancement velocities at pH 5.5 and 6.5 were 17.0 and 7.1 times higher than that under pH 7.4 in the first 2 h post-incubation, respectively (Fig. 2D). Similar to the absorption spectra, while the photoacoustic signal excited at 686 nm decreased consistently (Fig. 2E and S2A–C†), the signal excited at 756 nm increased gradually at pH 5.5 (Fig. S2D–F†). Fig. 2F shows the time dependent PA signal ratio (PA756/PA686) of DiIRB-S in the first 2 h after incubation under pH 7.4, 6.5 and 5.5. The slopes of the fitting straight lines between the incubation times and the PA756/PA686 ratios correlated to the acidity of the buffer solutions. The slopes measured at pH 5.5 and 6.5 were 9.5 and 2.6 times higher than the value measured at pH 7.4, respectively. | ||
| Scheme 1 pH responsive NIR fluorescence probe DiIRB-S evaluating tumor acidity. (A) Chemical structure of DiIRB-S. The acid liable hydrazone bond is indicated by a red color. (B) Cartoon representing the fluorescence enhancement mechanism of DiIRB-S under an acidic pH. | ||
 | ||
| Fig. 2 DiIRB-S determines pH values by acidity correlated absorption, emission and photoacoustic signal variation velocities. The time dependent absorption spectra (1.0 μM) at pH 5.5 (A) and the time courses of the A756/A686 absorption ratios (B) at pH 7.4, 6.5 and 5.5. The time dependent emission (1.0 μM) under pH 5.5 (C) and the time courses of fluorescence intensities (D) at pH 7.4, 6.5 and 5.5 (excited at 756 nm). The time dependent photoacoustic spectra (1.0 μM) under pH 5.5 (excited at 756 nm) (E) and the time courses of the PA756/PA686 photoacoustic signal ratio (F) at pH 7.4, 7.0, 6.5, 6.0 and 5.5. | ||
2.2 The ortho. tumors show higher gross acidity than the s.c. tumors
Besides the tumor extracellular fluid, acidosis also exists in the lysosomes of cancer cells. To verify the activation of DiIRB-S in the tumor extracellular matrix rather than in the lysosomes,28 the cultured U87MG cells were treated with DiIRB-S (25 μM) for 0.5, 2.0 or 12 h in a media (pH 6.5) mimicking the tumor extracellular environment (Fig. 3A). While the probe signals were not detected after incubation for 0.5 and 2.0 h, the signals of small vesicular structures were observed at 12 h post treatment and they colocalized well with the lysosomes stained by Lyso-tracker, which indicates that lysosomes are the intracellular delivering destination of the probe. Therefore, DiIRB-S actually senses acidosis in the tumor extracellular fluid but not in the lysosomes if the imaging studies are conducted within 2 h after treatment with the probe. DiIRB-S (40 nmol per mouse) successfully visualized both ortho. and s.c. glioma xenografts with significant NIR fluorescence enhancement at 2 h after intravenous injection of the probe (Fig. 3B). Ortho. and s.c. tumors demonstrated similar time courses of NIR fluorescence intensity. The fluorescence signal in both types of tumor increased substantially and reached their maximum as 8.3 ± 1.3 and 6.4 ± 0.4 × 109 p s−1 cm−2 per Sr (mean ± SD, n = 4) at 2 h post-injection (PI) (Fig. 3C). The signals of both tumors decreased after reaching the maximal values and the minimal values were observed at 24 h PI. However, the fluorescence intensities on the ortho. tumor were higher than those of the s.c. tumor during the whole imaging process, which implied the higher gross acidity of the extracellular fluid in the ortho. tumor. | ||
| Fig. 3 The ortho. glioma xenograft showed higher overall acidity than the s.c. glioma xenograft. (A) Fluorescence microscopic images of live U87MG cells treated with DiIRB-S (25 μM) in a media with pH 6.5 for selected incubation times. The fluorescence of Lyso-tracker is displayed in green and the NIR fluorescence is displayed in red. Bar = 50 μm. (B) Representative in vivo NIR fluorescence images of a mouse bearing the ortho. glioma xenograft at 2 h PI with DiIRB-S (40 nmol per mouse), unit: p s−1 cm−2 per Sr. Arrows point to the tumor position; (C) time dependent NIR fluorescence intensities on the tumors after i.v. injection of DiIRB-S. Data are presented as the mean ± SD (n = 4 mouse models bearing both ortho. and s.c. tumor xenografts). C.C. = color coded. | ||
2.3 The ortho. tumors show a more rapid glucose uptake than the s.c. tumors
Fig. 4A shows the representative 2D projection of in vivo microPET/CT images of a mouse model bearing both ortho. and s.c. glioma xenografts at 0.5 and 3.5 h PI with 18F-FDG (11 MBq per mouse). Prominent uptakes in both types of tumors, kidneys, intestinal tract, urinary bladder and brain were clearly observed at 0.5 h PI. Radioactivity accumulation was observed in normal brain tissues, especially in the cerebellum, which led to the compromised T/B ratio (1.19 ± 0.05) of the ortho. tumor (mean ± SD, n = 3). The radioactivity in normal brain tissues decreased with time after FGD injection, which resulted in the increased T/B ratio of the ortho. tumor (2.26 ± 0.10) at 3.5 h PI. Notably, in the s.c. tumor, the radioactivity initially located in the core area but extended to the whole tumor at 3.5 h PI. Fig. 4B presents the in vivo standardized uptake values (SUVs) of the ortho. and s.c. tumors at selected time-points PI with 18F-FDG. The minimal SUVs of both the ortho. (1.63 ± 0.07) and s.c. tumors (1.25 ± 0.14) were found at 3.0 min after 18F-FDG injection. The SUV of the ortho. tumor reached its maximal value, 2.03 ± 0.09, at 2 h PI. In contrast, the SUV of the s.c. tumor increased more rapidly and its maximum (2.10 ± 0.06) was recorded at 1 h PI. The SUVs of both the ortho. and s.c. tumors decreased after reaching their maximums and their values were determined as 1.76 ± 0.10 and 1.89 ± 0.08 respectively at 3.5 h PI. In vivo bio-distribution data demonstrated the highest SUVs in the heart at either 0.5 h (7.50 ± 2.4) or 3.5 h (7.13 ± 2.2) after FDG injection. While the SUV in the kidneys, brain and s.c. tumor decreased remarkably, the values in the heart and ortho. tumor were barely changed from 0.5 to 3.5 h PI (Fig. 4C). | ||
| Fig. 4 The ortho. glioma xenograft showed a more rapid uptake of radioactive glucose derivative 18F-FDG than the s.c. glioma xenograft. (A) Representative in vivo PET/CT images of a mouse model bearing both ortho. and s.c. glioma xenografts at 0.5 h and 3.5 h PI with i.v. 18F-FDG (11 MBq per mouse). The inserted pictures show the coronal positions of the ortho. and s.c. tumors, pointed out by the yellow and red arrows respectively. (B) The time course of the SUVs in the ortho. and s.c. tumors after administration of i.v. 18F-FDG. (C) In vivo bio-distribution of 18F-FDG in glioma xenografts and main organs at 0.5 and 3.5 h PI. Data are presented as the mean ± SD (n = 3 mouse models bearing both the ortho. and s.c. tumor xenografts). | ||
2.4 Ortho. gliomas show more aberrant vascular morphology than s.c. tumors
Fig. 5A shows the in vivo photoacoustic microscopy (PAM) images of the vasculature in the normal brain tissue, and the margin areas of the ortho. and s.c. glioma xenografts. The vasculature in normal brains formed a well-organized and regulated architecture, but the tumor vasculature lost the conventional hierarchy and were highly disorganized, tortuous, and characterized with inconsistent diameters and uneven shapes with abnormal bulges and blind ends. While the vessel diameters in normal brain tissue were determined in a range of 17.2–76.3 μm, broader diameter distribution were measured in the ortho. tumors (19.1–128.4 μm) and s.c. tumors (15.8–85.2 μm) (Fig. 5B). The average vessel diameter of the ortho. tumor was measured as 67.7 μm, which was higher than the s.c. tumor (51.1 μm) as well as normal brain tissue (47.4 μm). Fig. 5C shows the microscopic fluorescence images of the normal brain tissue and glioma sections, in which the vessels were immunohistochemically stained by CD31 antibody. Compared to the normal brain tissue, the nuclei in tumor margins were distributed with a higher density, which presents the fast proliferation rate of glioma cells. Similar to in vivo photoacoustic imaging, the vessels in both the ortho. and s.c. tumors showed aberrant morphology such as heterogeneous diameters, uneven shapes and blind ends. Additionally, the microvessel density (MVD) of the ortho. tumor was measured as 6.3 and 3.2 times higher than normal brain tissue and the s.c. tumors, respectively (Fig. 5D). | ||
| Fig. 5 The ortho. tumor showed a more variable vessel diameter and a higher vascular density than the s.c. tumor and normal brain tissue. (A) Representative in vivo PAM images of the vasculature in the normal brain tissue, and the margin areas of the ortho. and s.c. tumors. The yellow lines indicate vessels with maximal and minimal diameters in the field of view (excited at 532 nm). Bar: 200 μm. (B) Vascular diameters in the normal brain tissue, and the margin areas of the ortho. and s.c. tumors. (C) Microscopic fluorescence images of the normal brain, and the ortho. and s.c. tumor margins immuno-stained by CD31 antibody. The nuclear stain by DAPI is displayed in blue and the immunofluorescence, presenting vessels, is displayed in green. Bar: 50 μm. (D) Vascular densities in the normal brain, and the ortho. and s.c. tumor margins. Data are presented as the mean ± SD (n = 12 slides from 3 mouse models bearing both the ortho. and s.c. tumor xenografts). **P < 0.01. | ||
2.5 Both types of glioma shows higher expression levels of acidity associated enzymes
Ex vivo immunofluorescence microscopic imaging was conducted to evaluate the expression level of acidity associated enzymes in normal brain tissues and the invasive edges of the glioma xenografts. As shown in Fig. 6A, acidity associated enzymes including GLUT1, NHE1, MCT4, CA9 were clearly observed in the normal brain tissues and they located as tiny vesicular structures in the cytoplasm. Compared to normal brain tissues, all the enzymes mentioned above were over-expressed in both the ortho. and s.c. tumors. In the ortho. tumors, the enzymes distributed as star-like cluster structures extended from the peri-nuclei area to the extracellular space. In the s.c. tumors, these enzymes located as stripe-like structures, filling the whole intracellular and extracellular space. The enlarged MCT4 immunofluorescence images clearly show the different distribution patterns between the ortho. and s.c. tumors (Fig. 6B). Fig. 6C demonstrates the immunofluorescence intensities of these enzymes in normal brain tissue, and in the ortho. and s.c. tumors. Obviously, the s.c. tumor demonstrated the highest expression levels of GLUT1, NHE1, CA9 that were on average 10.4, 10.9 and 4.8 times higher than those of normal brain tissue, respectively. The highest expression level of MCT4 was detected in the ortho. tumor and it was 2.9 times higher than that of the normal brain tissue.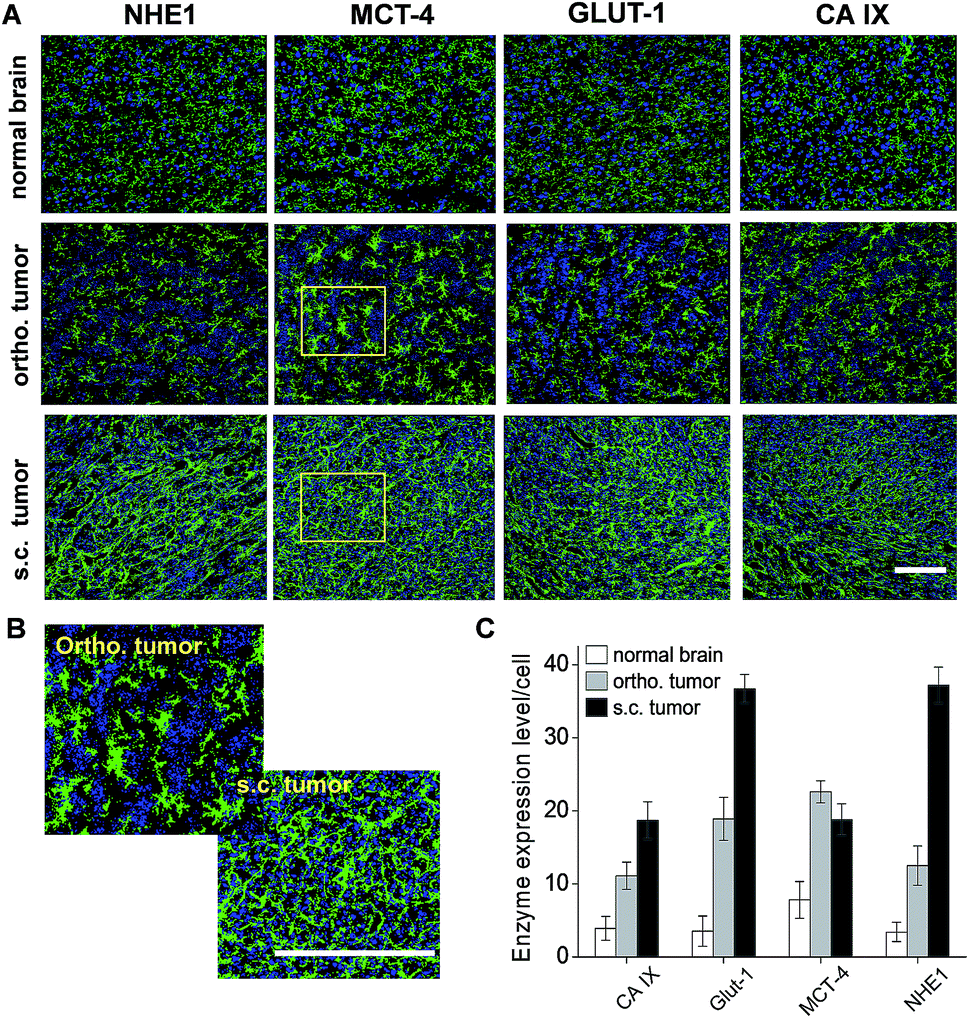 | ||
| Fig. 6 The expression level of acidity associated enzymes/ion channel proteins. (A) Representative microscopic immunofluorescence images of normal brain tissue and ortho. and s.c. tumor sections. The nuclear stain by DAPI is displayed in blue and the immunofluorescence is displayed in green. (B) Enlarged pictures of MCT4 immunofluorescence images highlighted in the yellow boxes of panel (A). (C) Average immunofluorescence intensity per cell in the normal brain tissues and the ortho. and s.c. tumors. Data are presented as the mean ± SD (n = 12 slides from 3 mouse models). Bar: 50 μm. | ||
3. Discussion
Magnetic resonance imaging (MRI) technologies such as MR spectroscopy imaging (MRSI)7 and chemical exchange saturation transfer (CEST) MRI29,30 have been used to determine the pH values in solid tumors. However, due to the inherently low sensitivity of MRI, a high dose of imaging agents (0.1–10 mM) and a long acquisition time (min–h) are inevitable. Compared to MRI, optical imaging demonstrates high sensitivity, a fast response rate and convenience for manipulation.31 NIR fluorescence imaging shows advantages for in vivo studies because NIR light, with wavelengths in the range of 650–900 nm, achieves low phototoxicity, and minimized absorption and autofluorescence from endogenous tissues, which allows it to visualize biological events in deep tissues (<1.0 cm).31 We previously developed a series of pH response NIR fluorescence probes to image tumors32,33 or evaluate tumor metastatic potentials25,34 by sensing acidic tumor microenvironment (TME). In this work, DiIRB-S was chosen to evaluate glioma invasiveness because of its predominant signal activation in tumor extracellular fluid. This capability of DiIRB-S could be explained by its remarkable pH responsive fluorescence enhancement and overall negative charge that prevents the efficient intracellular delivery into acidic lysosomes.PAM is a non-invasive imaging modality that detects optical absorption contrast by collecting the acoustic signal generated via a photoacoustic effect.35 As an emerging imaging technology, PAM combines the advantages of the high sensitivity of optical imaging and the spatial resolution of ultrasonic imaging.36 By using a NIR fluorophore as the imaging probe, PAM successfully detected objects embedded at depths of as much as 5.2 cm in chicken breast muscle, with a resolution of <780 μm and a sensitivity of <7 pmol probe in blood.37 Considering the pH correlated time dependent PA756/PA686 ratio of DiIRB-S, it is possible to determine the local pH values in gliomas after systemic injection of this probe.
Compared to oxidative phosphorylation (36 ATP per glucose), glycolysis is a less efficient glucose metabolic pathway to generate energy molecules (4 ATP per glucose).38 Due to the metabolic alternation from oxidative phosphorylation to aerobic glycolysis in cancer cells, tumors have to elevate glucose consumption to compensate for the low efficiency of glycolysis. Increased glucose uptake as a characteristic of glycolysis has been used to evaluate tumor malignancy. For example, Di Chiro et al. first showed that there was a positive correlation between glioma malignancy and glycolytic activity in clinical studies.39 High grade gliomas with high glycolytic levels demonstrated a much stronger radiotracer 18F-FDG (glucose analogue) uptake than low grade gliomas with low glycolytic levels. Therefore, it is possible to evaluate glycolytic level and invasiveness by quantifying the 18F-FDG signal intensity in the ortho. and s.c. tumors. The SUV, defined as the 18F-FDG retention normalized to the injected dose and tissue weight, is an established index for quantifying glucose metabolic activity in tissues. As shown in Fig. 4, a much higher SUV of ortho. tumors than s.c. tumors in the first 5 min PI with 18F-FDG implied the higher glycolytic level in the ortho. tumors. Lodge et al. reported that the high-grade gliomas took a longer time to reach the maximal FDG uptake value than benign lesions.40 Compared to s.c. tumors, ortho. tumors need one more hour to reach the maximum SUV, which indicates the higher malignancy/invasiveness of ortho. tumors compared to s.c. tumors.
The vasculatures in the high-grade gliomas had typical characteristics such as hyperpermeability, high vessel density, inconsistent diameters, unidentified arterioles and venules, tortuous shapes with abnormal bulges and blind ends.41,42 Imaging the micro-vasculature in glioma xenografts also helps to elucidate the invasiveness of gliomas. Even though MRI offers high spatial resolution to visualize tumor vasculature, it is still difficult to detect the vasculature with diameters less than 100 μm.43 Because the PA signal intensity usually correlates to its optical absorption, the much higher extinction coefficient of hemoglobin at 532 nm (200 cm−1) than other endogenous molecules such as epidermis (20 cm−1) or dermis (0.4 cm−1), provides its feasibility to non-invasively monitor micro-vasculature that are hard to detect by MRI.44 For example, PAM visualized vasculature in mouse ears with a lateral resolution as high as 0.5 μm under in vivo conditions.45 As shown in Fig. 5A, PAM demonstrated the highest vascular density and the most aberrant vascular morphology in the ortho. glioma xenograft. The heterogeneous vasculatures may accelerate the tumor acidity and invasiveness by the following explanations: (1) the poor oxygenation increases glucose metabolism via the glycolytic pathway and produces more lactic acid;41 (2) the limited perfusion rate hampers the drainage of the metabolized protons, which inevitably results in acidification of the local micro-environment; (3) the structural abnormalities of the tumor vasculature leads to spatial and temporal heterogeneity in the tumor blood flow, which elevates interstitial fluid pressure and pushes fluid flow from the tumor margin into the peri-tumor area and finally facilitates peri-tumor invasion by acidifying the local extracellular fluid.24,46 Therefore, ‘normalization’ of aberrant structures and functions of tumor vasculature will make them more efficient for oxygen delivery and then decrease the acidity and invasiveness of gliomas.
Compared to normal brain tissues, all four acidity associated enzymes tested in this work were up-regulated in both types of glioma xenograft, which partially explained the higher extracellular acidity in the tumors. Interestingly, in these enzymes, only MCT4 showed its highest expression level in the ortho. tumors. Previous work showed that MCTs induce intracellular acidification by the efflux of excess lactic acid out of the cytoplasm. Baltazar et al. reported that MCT1 and MCT4 were over-expressed in the plasma membrane of glioblastomas and MCT1 inhibitors exhibited anti-tumoral and antiangiogenic activity.47 As a potential therapeutic target, the down-regulation of MCTs may attenuate the acidic microenvironment and limit the invasiveness of gliomas.
4. Experimental
4.1 Materials
Rabbit anti-mouse CD31 primary antibody was purchased from Abcam (Cambridge, UK). Rabbit anti-mouse CA9, GLUT1, MCT4 and NHE1 primary antibody were purchased from Santa Cruze Biotechnology (California, USA). Alexa Fluo488-labeled goat anti-rabbit secondary antibody was purchased from Cell Signaling Technology (Danvers, USA). DAPI was purchased from Sigma (St. Louis., USA). 18F-FDG was produced automatically by a cyclotron (RDS Eclips ST; Siemens) and an Explora FDG4 module (Siemens) in Fudan University Shanghai Cancer Center. Human glioblastoma U87MG cells were purchased from the Chinese Academy of Science Cell Bank. Lysosome Tracker was purchased from Life Technology (Eugene, USA).4.2 Methods
All absorption spectra were recorded on a Shimadzu UV-2550 spectrophotometer. All fluorescence spectra were recorded on a Shimadzu RF-5301PC spectrophotometer. All pH measurements were performed with a Mettler Toledo MP220 pH meter equipped with a Mettler Toledo InLab® glass electrode. In vivo and ex vivo NIR fluorescence images were acquired in an IVIS Spectrum In Vivo Imaging System (Cailper Perkin Elmer, USA). In vivo photoacoustic microscopic images were collected from a home-made system equipped with an Nd:YAG laser, operating at a wavelength of 532 nm with a full-width at high magnitude (FWHM) of 4.0 ns and a repetition of 20 Hz. Meanwhile, a tunable pulsed laser source (Surelite OPO Plus, USA) was applied with wavelengths in a range of 675–100 nm, a pulse width of 4–6 ns, and a pulse repetition rate of 20 Hz. In vivo PET/CT imaging was performed on an Inveon system (Siemens, Germany) after intravenous injection of 18F-FDG. Live cell fluorescence microscopic images were collected on a Leica DMF4000B laser-scanning inverted microscope (Leica Inc., Germany) with epifluorescence by using 20× objective lenses. The Lyso-tracker was excited by a N3 filter (546 ± 6 nm) and its emission was collected with a 600 ± 20 nm band-pass filter. Meanwhile, DiIRB-S was excited by a Y7 filter (710 ± 37.5 nm) and its emission was detected by a 810 ± 45 nm band-pass filter. The immunofluorescence images were collected on a Zeiss LSM 710 META confocal laser scanning microscope (Carl Zeiss, Germany) by using a 20× lens. DAPI was excited with a 405 nm laser and the emission was detected with a photomultiplier by a 420–480 nm band-pass filter. Alexa-Flour488 was excited with a 488 nm laser and the emission was detected by a second photomultiplier using a 505–550 nm band-pass filter.4.3 pH dependent spectroscopic studies
The stock solution was prepared by dissolving DiIRB-S (9.1 mg, 5 × 10−6 mol) in anhydride dimethyl sulfoxide (DMSO) with a concentration of 10 mM. A working solution of DiIRB-S was prepared by diluting the stock solution with buffers to a final concentration of 1.0 μM. Absorption profiles of the DiIRB-S working solution were measured under pH 7.4, 6.5 and 5.5 at selected time points after incubation. The scanning speed was 0.5 nm s−1 and the slit width was set to 2 nm. Emission spectra of the DiIRB-S working solution with pH 7.4, 6.5 and 5.5 were excited at 745 nm and collected at selected time points after incubation. For photoacoustic spectroscopic studies, the laser beam (excited at 686 nm and 756 nm) was focused to illuminate the working solutions of DiIRB-S in a 0.5 mm diameter glass tube and the PA signals were received by an ultrasonic transducer. The PA signal profiles of the working solutions with pH 7.4, 7.0, 6.5, 6.0 and 5.5 were obtained at different time points after incubation. Each PA signal was generated by averaging data from scanning 128 times.4.4 Live cell fluorescence microscope imaging
Human glioblastoma U87MG cells (1.0 × 104) cultured in a 24 well-plate with approximately 50% confluence were treated with 25 μM DiIRB-S for 0.5, 2.0 or 12 h in 1.0 mL DMEM media (pH 6.5), supplemented with 10% FBS and 1% penicillin/streptomycin at 37 °C. After lysosome staining with Lyso-tracker (500 nM), the cells were washed 3× with PBS prior to microscope imaging.4.5 Tumor model development
U87MG cells grown as mono-layers in 75 cm2 flasks were cultured at 37 °C and 5% CO2 in high-glucose DMEM (Hyclone) containing 10% FBS and 1% penicillin/streptomycin. Male nude mice were purchased from Hua Bukang BioTek (Beijing, China) at 4–5 weeks of age and maintained under standard housing conditions. All animal experiments were carried out in accordance with guidelines approved by the Ethics Committee of Fudan University. U87MG cells (5.0 × 106 cells suspended in 150 μL PBS) were inoculated into the right thigh of the nude mice. U87MG cells (1 × 106 suspended in 3.0 μL PBS) were injected into the corpus striatum at 4–6 days after subcutaneous inoculation. After brain inoculation for 20 days, the mice bearing both the ortho. and s.c. glioma xenografts were ready for the imaging experiments.4.6 In vivo micro PET/CT imaging studies
After fasting for 8 h, the tumor bearing mice were anesthetized by isofluorine and then underwent dynamic PET/CT imaging after i.v. injection of 18F-FDG (11 MBq per mouse). Whole body PET was first performed with a 350–650 keV energy window and a 3.432 timing window. CT scanning covering the identical transverse field of view was obtained and the PET imaging datasets were reconstructed iteratively by application of the CT data for attenuation correction and co-registration. The PET/CT images were analyzed on a Host IAW workstation with Inveon Research Workplace software. An elliptic volume of interest (VOI) was drawn manually around areas with abnormal 18F-FDG uptake to calculate the SUV peak (the hottest spot in the tumor foci).4.7 In vivo optical imaging
NIR fluorescence images were acquired using an Integrin 750 filter set (excitation filter: 745 nm; emission filter: 800–840 nm) after systemic injection of DiIRB-S (40 nmol per mouse). The mice were anesthetized by isoflurane and placed on an imaging plate in a position to visualize both the ortho. and s.c. glioma xenografts at the same time. At the end of imaging, the mice were scarified and perfused with saline followed by 4% paraformaldehyde (PFA) via the heart to douche blood and pre-fix the mouse. The brain, subcutaneous tumor and main organs were isolated, imaged and the fluorescence intensities of isolated organs were quantified respectively.4.8 In vivo photoacoustic microscopic imaging
In vivo PAM imaging of blood vessels in the normal brain tissue and invasive margins of the tumors were conducted in mouse models after anesthesia (10% chloral hydrate, 85 μL per mouse). A field flattening len with a magnification of 4× was used to scan a region with a diameter of 1.0 mm. An Nd:YAG laser (Surelite II-20, Continuum, USA), operating 4–6 ns pulses at a 532 nm wavelength with a repetition rate of 20 Hz, was used as the light source. A tunable pulsed laser with a repetition rate of 20 Hz and a pulse width of 4 ns (Nd:YAG Surelight-II-20 connected to Surelite OPO Plus, spectral tuning range 675–1000 nm, Continuum) was used as the light source. The light was scanned by a 2D scanning galvanometer (6231 H, Cambridge Technology) and focused by the objective lens to irradiate the tested tissues. An unfocused ultrasound transducer with a center frequency of 15 MHz and a −6 dB bandwidth was used to receive the PA signals generated by the tested sample. A silicon photodiode (ET 2000, Electro-Optics Technology, Inc., USA) was used to monitor and calibrate the intensity and stability of the laser beam. The PA signals were recorded through the signal amplifier and a dual-channel data acquisition card. The blood vessels of the tumor xenografts were imaged by PAM at 532 nm.4.9 Immunofluorescence microscopic imaging
Tumor bearing mice were scarified and perfused with saline followed by 4% PFA via the heart to douche blood. The isolated brains and s.c. tumors were fixed in formalin for 12 h, 30% sucrose solution for 24 h and then sectioned coronally with a thickness of 10 μm. The brain and s.c. tumor sections were stained by a primary antibody including MCT4, GLUT1, NHE1, CA9 respectively with a 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :100 dilution. After washing in PBS, the slides were further stained by Alexa fluor 488-conjugated goat anti-rabbit secondary antibody (1:500 dilutions). Confocal fluorescence microscopic images were captured in a laser confocal living cell imaging system (Carl Zeiss LSM710) equipped with a 20× objective lens. The fluorescence intensities per cell were quantified by ImageJ (NIH, Bethesda, MD) software (n = 8).
:100 dilution. After washing in PBS, the slides were further stained by Alexa fluor 488-conjugated goat anti-rabbit secondary antibody (1:500 dilutions). Confocal fluorescence microscopic images were captured in a laser confocal living cell imaging system (Carl Zeiss LSM710) equipped with a 20× objective lens. The fluorescence intensities per cell were quantified by ImageJ (NIH, Bethesda, MD) software (n = 8).
4.10 Statistics
Values are presented as the mean ± SD when the sample number was above 4 (n > 4).5. Conclusion
Overall, by using multi-parametric imaging technologies, we verified the positive correlation between intratumoral acidity and glioma invasiveness. It is promising for curbing glioma invasiveness by directly neutralizing intratumoral acidity or blocking the acidification associated physiological incidents such as aerobic glycolysis, vascular heterogeneity and the over-expression of acidity associated enzymes/transporters.Acknowledgements
This work was financially supported by the National Basic Research Program of China (973 Program, 2011CB910404, 2013CB932500), the National Natural Science Foundation of China (nos 81171384, 81371624), the New Century Excellent Talents in University Award and the Special Nano-Projects of Shanghai Science and Technology Committee (13NM1400400).Notes and references
- P. Y. Wen and S. Kesari, N. Engl. J. Med., 2008, 359, 492 CrossRef CAS PubMed.
- R. Stupp, W. P. Mason, M. J. van den Bent, M. Weller, B. Fisher, M. J. Taphoorn, K. Belanger, A. A. Brandes, C. Marosi, U. Bogdahn, J. Curschmann, R. C. Janzer, S. K. Ludwin, T. Gorlia, A. Allgeier, D. Lacombe, J. G. Cairncross, E. Eisenhauer and R. O. Mirimanoff, European Organisation for Research and Treatment of Cancer Brain Tumor and Radiotherapy Groups and National Cancer Institute of Canada Clinical Trials Group, N. Engl. J. Med., 2005, 352, 987 CrossRef CAS PubMed.
- N. J. Abbott, L. Ronnback and E. Hansson, Nat. Rev. Neurosci., 2006, 7, 41 CrossRef CAS PubMed.
- A. Giese, Acta Neurochirurgica Supplement, 2003, 88, 153 CAS.
- P. Friedl and K. Wolf, Nat. Rev. Cancer, 2003, 3, 362 CrossRef CAS PubMed.
- B. A. Webb, M. Chimenti, M. P. Jacobson and D. L. Barber, Nat. Rev. Cancer, 2011, 11, 671 CrossRef CAS PubMed.
- R. van Sluis, Z. M. Bhujwalla, N. Raghunand, P. Ballesteros, J. Alvarez, S. Cerdan, J. P. Galons and R. J. Gillies, Magn. Reson. Med., 1999, 41, 743 CrossRef CAS.
- A. F. Chambers and L. M. Matrisian, J. Natl. Cancer Inst., 1997, 89, 1260 CrossRef CAS PubMed.
- Z. M. Bhujwalla, C. L. McCoy, J. D. Glickson, R. J. Gillies and M. Stubbs, Br. J. Cancer, 1998, 78, 606 CrossRef CAS PubMed.
- K. Fischer, P. Hoffmann, S. Voelkl, N. Meidenbauer, J. Ammer, M. Edinger, E. Gottfried, S. Schwarz, G. Rothe, S. Hoves, K. Renner, B. Timischl, A. Mackensen, L. Kunz-Schughart, R. Andreesen, S. W. Krause and M. Kreutz, Blood, 2007, 109, 3812 CrossRef CAS PubMed.
- E. K. Rofstad, B. Mathiesen, K. Kindem and K. Galappathi, Cancer Res., 2006, 66, 6699 CrossRef CAS PubMed.
- V. Estrella, T. Chen, M. Lloyd, J. Wojtkowiak, H. H. Cornnell, A. Ibrahim-Hashim, K. Bailey, Y. Balagurunathan, J. M. Rothberg, B. F. Sloane, J. Johnson, R. A. Gatenby and R. J. Gillies, Cancer Res., 2013, 73, 1524 CrossRef CAS PubMed.
- R. A. Gatenby and E. T. Gawlinski, Cancer Res., 1996, 56, 5745 CAS.
- R. A. Gatenby and R. J. Gillies, Nat. Rev. Cancer, 2004, 4, 891 CrossRef CAS PubMed.
- O. Warburg, Science, 1956, 123, 309 CAS.
- I. F. Robey, B. K. Baggett, N. D. Kirkpatrick, D. J. Roe, J. Dosescu, B. F. Sloane, A. I. Hashim, D. L. Morse, N. Raghunand, R. A. Gatenby and R. J. Gillies, Cancer Res., 2009, 69, 2260 CrossRef CAS PubMed.
- E. A. Mazzio, B. Smith and K. F. Soliman, Cell Biol. Toxicol., 2010, 26, 177 CrossRef CAS PubMed.
- J. Yun, C. Rago, I. Cheong, R. Pagliarini, P. Angenendt, H. Rajagopalan, K. Schmidt, J. K. Willson, S. Markowitz, S. Zhou, L. A. Diaz Jr, V. E. Velculescu, C. Lengauer, K. W. Kinzler, B. Vogelstein and N. Papadopoulos, Science, 2009, 325, 1555 CrossRef CAS PubMed.
- T. Amann and C. Hellerbrand, Expert Opin. Ther. Targets, 2009, 13, 1411 CrossRef CAS PubMed.
- R. Le Floch, J. Chiche, I. Marchiq, T. Naiken, K. Ilc, C. M. Murray, S. E. Critchlow, D. Roux, M. P. Simon and J. Pouyssegur, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 16663 CrossRef CAS PubMed.
- J. Chiche, K. Ilc, J. Laferriere, E. Trottier, F. Dayan, N. M. Mazure, M. C. Brahimi-Horn and J. Pouyssegur, Cancer Res., 2009, 69, 358 CrossRef CAS PubMed.
- R. A. Cardone, V. Casavola and S. J. Reshkin, Nat. Rev. Cancer, 2005, 5, 786 CrossRef CAS PubMed.
- E. Bullitt, G. Gerig, S. Aylward, S. Joshi, K. Smith, M. Ewend and W. L. Lin, in Medical Image Computing and Computer-Assisted Intervention – Miccai, ed. R. Ellis and T. Peters, Springer, Berlin, Heidelberg, 2003, vol. 2878, Pt 1, p. 671 Search PubMed.
- D. Fukumura and R. K. Jain, Microvasc. Res., 2007, 74, 72 CrossRef CAS PubMed.
- L. Wang, X. Zhu, C. Xie, N. Ding, X. F. Weng, W. Y. Lu, X. B. Wei and C. Li, Chem. Commun., 2012, 48, 11677 RSC.
- L. Wang, Z. Fan, J. Zhang, Y. Changyi, C. Huang, Y. Gu, Z. Xu, Z. Tang, W. Lu, X. Wei and C. Li, Int. J. Cancer, 2015, 136, E107 CrossRef CAS PubMed.
- R. Khairutdinov and N. Serpone, J. Phys. Chem. B, 1997, 101, 2602 CrossRef CAS.
- Z. Li, Y. Song, Y. Yang, L. Yang, X. Huang, J. Han and S. Han, Chem. Sci., 2012, 3, 2941 RSC.
- V. R. Sheth, Y. G. Li, L. Q. Chen, C. M. Howison, C. A. Flask and M. D. Pagel, Magn. Reson. Med., 2012, 67, 760 CrossRef PubMed.
- D. D. Castelli, G. Ferrauto, J. C. Cutrin, E. Terreno and S. Aime, Magn. Reson. Med., 2014, 71, 326 CrossRef PubMed.
- S. Luo, E. Zhang, Y. Su, T. Cheng and C. Shi, Biomaterials, 2011, 32, 7127 CrossRef CAS PubMed.
- C. Li, J. Xia, X. Wei, H. Yan, Z. Si and S. Ju, Adv. Funct. Mater., 2010, 20, 2222 CrossRef CAS PubMed.
- C. Li, K. Li, H. Yan, G. Li, J. Xia and X. Wei, Chem. Commun., 2010, 46, 1326 RSC.
- Z. Si, C. Huang, X. Gao and C. Li, RSC Adv., 2014, 4, 55548 RSC.
- H. F. Zhang, K. Maslov, G. Stoica and L. V. Wang, Nat. Biotechnol., 2006, 24, 848 CrossRef CAS PubMed.
- X. D. Wang, Y. J. Pang, G. Ku, X. Y. Xie, G. Stoica and L. H. V. Wang, Nat. Biotechnol., 2003, 21, 803 CrossRef CAS PubMed.
- G. Ku and L. V. Wang, Opt. Lett., 2005, 30, 507 CrossRef.
- M. G. Vander Heiden, L. C. Cantley and C. B. Thompson, Science, 2009, 324, 1029 CrossRef CAS PubMed.
- G. Di Chiro, R. L. DeLaPaz, R. A. Brooks, L. Sokoloff, P. L. Kornblith, B. H. Smith, N. J. Patronas, C. V. Kufta, R. M. Kessler, G. S. Johnston, R. G. Manning and A. P. Wolf, Neurology, 1982, 32, 1323 CrossRef CAS.
- M. A. Lodge, J. D. Lucas, P. K. Marsden, B. F. Cronin, M. J. O’Doherty and M. A. Smith, Eur. J. Nucl. Med., 1999, 26, 22 CrossRef CAS.
- D. W. Siemann, Cancer Treat. Rev., 2011, 37, 63 CrossRef CAS PubMed.
- E. Bullitt, D. L. Zeng, G. Gerig, S. Aylward, S. Joshi, J. K. Smith, W. L. Lin and M. G. Ewend, Acad. Radiol., 2005, 12, 1232 CrossRef PubMed.
- A. P. Pathak, E. Kim, J. Zhang and M. V. Jones, PLoS One, 2011, 6, e22643 CAS.
- G. Ku, X. D. Wang, X. Y. Xie, G. Stoica and L. H. V. Wang, Appl. Opt., 2005, 44, 770 CrossRef.
- Y. Yuan, S. H. Yang and D. Xing, Appl. Phys. Lett., 2012, 100, 023702 CrossRef PubMed.
- R. T. Tong, Y. Boucher, S. V. Kozin, F. Winkler, D. J. Hicklin and R. K. Jain, Cancer Res., 2004, 64, 3731 CrossRef CAS PubMed.
- V. Miranda-Goncalves, M. Honavar, C. Pinheiro, O. Martinho, M. M. Pires, C. Pinheiro, M. Cordeiro, G. Bebiano, P. Costa, I. Palmeirim, R. M. Reis and F. Baltazar, Neuro-Oncology, 2013, 15, 172 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c5ra07685d |
| This journal is © The Royal Society of Chemistry 2015 |