
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceSilicon alleviates cadmium toxicity in muskmelon (Cucumis melo): integrative insights from photosynthesis to antioxidant activity to gene expression
Ram
Krishna†
a,
Mohammad
Shahid
 b,
Waquar Akhter
Ansari†
*c,
Khalid Mashay
Al-Anazi
d,
Mohammad Abul
Farah
d,
Durgesh Kumar
Jaiswal
e,
Akhilesh
Yadav
f,
Sudhakar
Pandey
g and
Md Azizul
Haque
*h
b,
Waquar Akhter
Ansari†
*c,
Khalid Mashay
Al-Anazi
d,
Mohammad Abul
Farah
d,
Durgesh Kumar
Jaiswal
e,
Akhilesh
Yadav
f,
Sudhakar
Pandey
g and
Md Azizul
Haque
*h
aInstitute of Environment and Sustainable Development, Banaras Hindu University, Varanasi 221005, Uttar Pradesh, India
bDepartment of Agricultural Microbiology, Faculty of Agricultural Science, Aligarh Muslim University, Aligarh 202002, Uttar Pradesh, India
cMarwadi University Research Centre, Department of Agriculture, Faculty of Science, Marwadi University, Morbi Road, Rajkot 360003, Gujarat, India. E-mail: waquar.ansari@gmail.com; waquar.ansari@marwadieducation.edu.in
dDepartment of Zoology, College of Science, King Saud University, P. O. Box 2455, Riyadh 11451, Saudi Arabia
eDepartment of Biotechnology, Graphic Era (Deemed to be University), Dehradun 248002, Uttarakhand, India
fDepartment of Plant Science, University of California, Davis 95616, California, USA
gIndian Council of Agricultural Research, Pusa Campus, New Delhi-110012, India
hDepartment of Biotechnology, Yeungnam University, Gyeongsan 38541, Republic of Korea. E-mail: azizul@ynu.ac.kr
First published on 1st May 2025
Abstract
Cadmium (Cd) toxicity is a potential abiotic stress that significantly affects plant growth and development, especially in vegetable crops like muskmelon. Silicon (Si), a vital element, plays an essential role in alleviating Cd-induced stress by various mechanisms. The present study explored the impact of Cd and Si treatments at 25, 50, 100, and 200 mg per kg soil concentrations on the physio-biochemical and molecular changes in muskmelon. Si treatment stabilized key physiological factors like increasing photosynthesis by 16% to 98%, decreasing electrolyte leakage by 28% to 69%, and increasing the relative water content by 7% to 48%, respectively, under increasing concentrations of Cd treatments supplemented with Si, improving the overall plant health. The results indicated that Cd exposure led to increased oxidative damage, as evidenced by the levels of hydrogen peroxide, malondialdehyde, and proline, under a maximum concentration of 200 mg kg−1 with respective fold increases of 3.44, 2.84 and 5.03, while Si supplementation significantly mitigated these effects, with respective fold changes of 0.83, 0.92 and 0.87, under similar concentrations, when compared with the control plants. Si enhanced the activity of the antioxidant enzyme and improved the gene expression associated with oxidative stress, providing a protective effect against Cd-induced damage. Si also promoted nutrient uptake and alleviated the unfavourable impacts of Cd on soil physio-biochemical properties, like pH, electrical conductivity (EC), and organic carbon (OC) content. The combined application of Cd and Si showed a synergistic effect, enhancing plant resilience and reducing Cd accumulation. These findings suggest that Si is an effective strategy for mitigating Cd toxicity, improving plant health, and promoting crop productivity, particularly in muskmelon and other vegetable crops.
Environmental significanceCadmium (Cd) is a non-biodegradable, hazardous microelement prevalent in the atmosphere. The transfer of cadmium through ecosystems presents significant threats to the ecology. Human activity, including metal extraction, smelting, and application of chemical pesticides and fertilizers, has increased the levels of cadmium in the ecosystem. Cadmium exposure to crops induces the formation of reactive oxygen species (ROS), which impair the functioning of cells and numerous biological molecules in plants. The detrimental impact of cadmium results in a disruption of physiological systems, development, and overall growth in plants. To date, the effect of cadmium has not been explored in detail. This study shows that silicon reduces the severity of cadmium in muskmelon by improving the plant’s overall performance. |
Introduction
Cadmium (Cd), a nonessential heavy metal, is considered a severely toxic contaminant due to its potential to accumulate in organisms and remain for a long duration, even at negligible concentrations.1 It causes serious health issues in humans and animals when it enters the food chain.2 The soil contaminated with Cd has a negative impact on plant growth attributes, leading to symptoms such as yellowing of leaves, reduced growth and yields, decreased activity of enzymes, accumulation of reactive oxygen species (ROS), and protein damage.3,4 Cd is easily taken up by plants because of its high movement in soil and plant systems. Excessive build-up of cadmium within the body of an individual can lead to calcification of the kidneys and bones, problems with metabolism, ostealgia, hypertension, diabetes, emphysema, and various other disorders. It can result in oxidative damage to deoxyribonucleic acid (DNA), impede the repair mechanisms, and promote the formation of cancer cells.1Muskmelon variety Kashi Madhu has been developed by the Indian Council of Agriculture Research-Indian Institute of Vegetable Research (ICAR-IIVR). It is known for its high yield, good fruit quality, and resistance to various diseases. Kashi Madhu has a round, slightly flattened shape with a smooth, golden-yellow skin and sweet, orange-colored flesh. The variety is preferred for its taste, attractive appearance, and relatively high market value. It is well-suited for cultivation in regions with a warm climate and is known for its ability to thrive under both rainfed and irrigated conditions. Kashi Madhu also has a good shelf life, making it suitable for commercial farming and marketing.5 Interestingly, China, Turkey, and India produced 14.07, 1.64, and 1.48 million metric tons of muskmelons, respectively, during 2023–24 (Foreign Agriculture Service, USDA, assessed on 24th January, 2025).
Silicon (Si) constitutes approximately 27.7% of the Earth’s crust, ranking as the second largest component after oxygen, and is not necessary for the survival of higher plants on the Earth. Nonetheless, it plays a significant role in maintaining plant growth attributes and product qualities under abiotic and biotic stresses.6 It has been determined that silicon dioxide nanoparticles are an attractive kind of nanomaterial that has the potential to address a wide range of stressors, including the toxicity of heavy metals in polluted soil. In addition, when compared to traditional silicon fertilizers, SiO2 NPs have the ability to successfully minimize the effects of cadmium stress. Increases in Cd toxicity, plant growth, photosynthetic parameters, and chlorophyll contents can be achieved through the use of silicon dioxide nanoparticles. Additionally, the concentration of Cd within plants can be decreased through the utilization of these nanoparticles.7 Si enhances soil qualities by boosting soil microbiome, regulating nutrient cycling, increasing retention of soil water, and creating a suitable environment for plant growth.8 Although, in nature, Si is present in plentiful amounts, it is generally found in complex forms to which plants cannot have access. Solubilized, bio-available forms such as monosilicic/protosilicic acid [Si(OH)4/H4SiO4] generally occur at low concentrations (0.1–0.6 mM) in soil.9 Si is absorbed by plants solely as H4SiO4, the only accessible form in the soil. Once absorbed, H4SiO4 moves radially through the cortex of the root via the plasmodesmata or cytoplasm.10,11 After reaching the xylem, it is fastly transported to the aerial parts of the plants such as stems, leaves, flowers, and fruits with the help of a transpiration stream.12 In plant organs like leaves and stems, H4SiO4 is moved via the xylem to the extracellular apoplastic space.
The phytotoxic effects of Cd in the soil become evident only after plants absorb it and translocate it to different tissues and organs. There are two pathways by which Cd can enter plant roots: (1) apoplast and (2) symplast pathways.13,14 In the apoplast pathway, Cd is absorbed by roots through positive diffusion and moves through the spaces between the rhizodermis cell walls and cortex, eventually entering the xylem and phloem without entering the cells. The symplast pathway, on the other hand, involves the movement of Cd through cellular plasmodesmata facilitated by membrane transporters of the cells.15,16
Si can interfere with both pathways of Cd uptake, reducing Cd accumulation in plants and thus mitigating its toxicity.15,17 However, Si improves plant growth attributes under Cd toxicity.18 The photosynthesis disruption is a key impact of Cd stress that can be minimized by Si application.19 Furthermore, Si constitutes barriers in physical form within the cell wall and also alters the expression genes that participated in Cd absorption and transportation. Under Cd stress, high ROS amounts such as hydrogen peroxide (H2O2), hydroxyl radical (OH−), and superoxide ion (O2−), accumulate in the cells of the plant.20,21 Antioxidant enzymes like ascorbate peroxidase (APX), catalase (CAT), guaiacol peroxidase (GPX), peroxidase (POD) and superoxide dismutase (SOD) are synthesized by plants to counteract the negative impact of ROS at high concentrations under Cd toxicity.22 Si can increase the plant's antioxidative capability, and the application of SiO2 (silicon dioxide) nanoparticles activates the antioxidative defense machinery.23
Although there is extensive research on the role of Si in alleviating environmental stresses caused by cadmium in different crops, to the best of our knowledge, the role of silicon in mitigating cadmium stress in muskmelon has not been studied. Therefore, based on the aforementioned considerations, the current experiment was planned to investigate whether silicon can enhance cadmium tolerance in muskmelon by modulating different physio-biochemical and molecular parameters. Therefore, implementing effective strategies to mitigate Cd stress and reduce its concentration in plants via Si treatment is a future aspect for enhancing muskmelon plant growth and ensuring better productivity.
Materials and methods
Plant materials and treatments
Muskmelon seeds of the variety Kashi Madhu were used in the present study. Experiments were carried out at the Dept. of Agriculture, Marwadi University Research Center, Marwadi University, Rajkot, Gujarat, India, with 22.36° N latitude and 70.79° E longitude. Seeds were disinfected using 3% sodium hypochlorite (NaOCl) for 5 min followed by washing thrice using distilled water. Seeds were germinated in plastic trays filled with a coco peat and vermiculite mixture. Plant at the true leaf stage was used for transplanting in pots with diameter and height of 21 and 23 cm, respectively. Pot soil pH and bulk density were 6.8 and 129 g cm−3, respectively. CdSO4 (cadmium sulfate), in the concentration range of 25 mg, 50 mg, 100 mg, and 200 mg Si per kg soil, were mixed with the soil in pots before two days of seed sowing; similarly, Si (SiO2) in the concentration range of 25 mg, 50 mg, 100 mg, and 200 mg Cd per kg were mixed with the soil. While the soil moisture content was maintained up to 65%. The setup of treatments was as follows: T1 = control (untreated), T2 = 25 mg Cd per kg soil, T3 = 50 mg Cd per kg soil, T4 = 100 mg Cd per kg soil, T5 = 200 mg Cd per kg soil, T6 = 25 mg Si per kg soil, T7 = 50 mg Si per kg soil, T8 = 100 mg Si per kg soil, T9 = 200 mg Si per kg soil, T10 = 25 mg Cd per kg soil + 25 mg Si per kg soil, T11 = 50 mg Cd per kg soil + 50 mg Si per kg soil, T12 = 100 mg Cd per kg soil + 100 mg Si per kg soil, and T13 = 200 mg Cd per kg soil + 200 mg Si per kg soil. Each of the experiments were carried out in triplicate (n = 3). Pots were irrigated with 1.5 liters of water at three days intervals, and data were recorded after 50 days of germination when the fruit setting starts.Root and shoot length measurement
Three representative plants from each treatment were carefully and meticulously uprooted from their pots, ensuring minimal disturbance to the roots. This careful approach was taken to maintain the accuracy of the data and to provide a reliable basis for our research. Root and shoot measurements were carried out using meter tape.Root and shoot dry and fresh weight measurements
Plants extracted intact from the roots were separated into shoot and root parts, and their fresh mass (FM) was recorded. Furthermore, these parts were dried for 48 h at 80 °C in a hot air oven to obtain the dry mass (DM). The dry-to-fresh mass ratio of each portion was calculated as a percentage using formulae, which were adapted from Kausar24 with slight modifications.25Net photosynthetic rate (Pn), stomatal conductance (gs), chlorophyll fluorescence (Fv/Fm) and chlorophyll color index (CCI) measurement
The Pn and gs were meticulously recorded using a portable photosynthetic system (LI-6200; LICOR, USA). These measurements were conducted on 03 leaves per replication between 11:00 AM and 1:00 PM. Fv/Fm was assessed by recording the photosystem II maximum quantum efficiency, by employing a hand usable Plant Efficiency Analyzer. For this measurement, 30 min dark adaption of leaves was carried out by employing clips on the adaxial surface. Excitation irradiance was maintained at 3000 μmol m−2 s−1. Higher (F0) and lower (Fm) fluorescence were recorded to calculate Fv/Fm.For CCI measurements, the top three leaves were selected during daytime hours (10:00–11:00 AM) using a handy chlorophyll meter at wavelengths of 655/940 nm, which are known to be optimal for chlorophyll content determination.
Electrolyte leakage (EL) and relative water content (RWC)
For EL assessment, 1 cm diameter leaf discs were soaked in 20 ml of water at room temperature for 4 h to measure the initial conductivity (EC1). The discs were further autoclaved for 30 min at 121 °C to measure the final conductivity (EC2). EL was calculated with the help of the formula given below:| EL% = (EC1/EC2) × 100.25 |
The RWC was measured following meticulous protocols.26 The fresh weight (FW) of leaf samples was recorded first followed by soaking in water for full turgidity for 6 h and measurement of the turgid weight (TW). Subsequently, the leaf samples were dried at 80 °C using an oven to measure the dry weight (DW). The calculation of RWC% was performed using the formula: RWC% = (FW − DW/TW − DW) × 100.
Chlorophyll (Chl) and carotenoid (Car) measurement
For Chl and carotenoid estimation, approximately 250 mg of fresh leaf tissue was crushed in 80% of chilled acetone by utilizing a mortar and pestle. The supernatant absorbance was measured at 663 nm, 645 nm, and 470 nm wavelengths for Chl a, Chl b, and Car, respectively, on a UV-vis spectrophotometer (UV-1900, Shimadzu, Japan). The pigment concentrations were calculated according to Ansari.26Leaf proline content
The free proline content was measured using the ninhydrin technique reported by Bates.27 After filtering the homogenate solution, 2 ml of the filtrate was transferred to fresh test tubes, and 2 ml of each of acid ninhydrin and glacial acetic acid was mixed. The tubes were incubated at 80 °C for 1 h followed by rapid cooling on ice to stop the reaction. The optical densities were noted at 520 nm following the separation of the toluene-based chromophore from the aqueous phase.Estimation of hydrogen peroxide (H2O2)
The KI reagent was used to quantify the H2O2 concentration in accordance with Jana and Choudhuri.28 Four ml of 5% trichloroacetic acid (TCA) was utilized for crushing 0.2 g of frozen roots from each pot. Following centrifugation at 10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 rpm for 15 min at 4 °C, the supernatant (0.5 ml) was added with 0.5 ml of phosphate buffer (0.1 M, pH = 7.0) and 1 ml of KI (1 M), and then dark incubation was performed at room temperature for 1 h.
000 rpm for 15 min at 4 °C, the supernatant (0.5 ml) was added with 0.5 ml of phosphate buffer (0.1 M, pH = 7.0) and 1 ml of KI (1 M), and then dark incubation was performed at room temperature for 1 h.
Leaf malondialdehyde (MDA)
Malondialdehyde (MDA), which is produced by the peroxidation of lipids, was estimated in fresh leaf tissues by following the methodology stated by Weisany.29 A total of 0.2 g of freshly collected leaf tissue was crushed in 5 ml of trichloroacetic acid (TCA; 0.1% w/v), followed by centrifugation at 12000 g for 5 min at 4 °C. Finally, the extract was cooled on ice, and the optical density was recorded at 532 nm.
Assay of antioxidant enzyme activity
To extract antioxidant enzymes, 1 g of leaf tissues was crushed in 50 mM phosphate buffer. After centrifuging at 15000 g for 20 min at 4 °C, the supernatant was collected and utilized as a source of enzymes. Using the technique described earlier,30 the amount of protein in the supernatant was determined. Estimation of superoxide dismutase (SOD, EC 1.15.1.1) was carried out as stated by Beyer.31 The optical density was measured at 560 nm, U per mg protein was used to express the activity of SOD. The activity of catalase (CAT, EC1.11.1.6) was measured by tracking the change in optical density at 240 nm for 2 min.32 Ascorbate peroxidase (APX, EC 1.11.1.11) activity was recorded using the procedure described in Nakano and Asada,33 and absorbance was measured for 3 min at 290 nm. The activity technique of Foyer and Fletcher34 was used for glutathione reductase (GR; EC 1.6.4.2), and the change in the optical density at 340 nm was monitored for 2 min.
RNA isolation, cDNA preparation, and real-time PCR
Total RNA was extracted from the leaf tissue using a Chromous RNA Isolation Kit. One μg of total RNA with random primers was used to synthesize 1st strand cDNA using a synthesis kit. The cDNA was further diluted and used for real-time PCR. 1st strand cDNA (2 μl) in 20 μl reaction was mixed with 10 μl iQ-SYBR Green Supermix (Bio-Rad, United States), 0.8 μl of 100 ng μl−1 of each gene-specific primer pair, and 6.4 μl of Milli-Q water. Changes in the expression pattern of antioxidant enzyme-related genes, SOD, CAT, PO, PAL, PR2 and PR3, were determined by qRT-PCR. The qRT-PCR was performed at 94 °C (5 min) for 1 cycle, 35 cycles at 95 °C (20 s), and annealing as per the Tm of individual primer (1 min) and extension at 72 °C (30 s). The output from the real-time software was analyzed and was used for the cycle threshold (CT) values. Calculations of relative expression were performed by implementing the ΔΔCT.35Element quantification
The concentration of Na, Mg, K, Ca, and Fe was measured using a 7700x ICP-MS system (Agilent Technologies, USA). Dried leaf tissues were finely powdered and subject to microwave digestion with HNO3 (SupraPur™, USA). The analysis procedure followed the guidelines outlined by Bhati.36 Metal analysis was conducted on three independent replicates. For data processing, we utilized Syngistix for ICP-MS software (PerkinElmer, Waltham, Massachusetts, USA).Determination of total and available Cd in muskmelon and soil
The nitrate method was utilized to determine Cd in muskmelon as described elsewhere.37 The plant and soil samples were dried and powdered by grinding. A total of 3 ml nitric acid (concentrated) was added and mixed followed by 1 h of boiling in a hot water bath. The mixture was allowed to cool, then 0.5 ml of 30% H2O2 solution was added to the reaction and warmed for 30 min in a hot water bath. The reaction mixture was cooled and diluted by adding deionized water to an optimum concentration. Diethylenetriaminepentaacetic acid (DTPA) was utilized to extract the soil samples followed by filtration for 2 h on a shaking table, the resultant solution was diluted by adding deionized water. The concentration of Cd in different muskmelon parts and soil was estimated with the help of an Avanta M atomic absorption spectrophotometer,38 while HMs were calculated using the equation given by Lu.39Statistical interpretation
All the statistical analysis for the present study was performed with the help of SPSS software (version 16.0, SPSS Inc.). Three independent replicates (biological) were taken for each experiment, and each was conducted in triplicate.Results
Soil parameters
The impact of different treatments of Cd and Si, individually and in combination, on soil properties was assessed, including electrical conductivity, pH, and organic carbon content. For the control, the EC was 209, pH was 7.1, and OC was 0.98. Cadmium (Cd) treatments increased the EC from 212 (25 mg Cd per kg) to 219 (200 mg Cd per kg), with pH remaining around 7.1–7.2 and OC slightly decreasing to 0.96, although these changes were non-significant (Fig. 1A and B). Silicon (Si) treatments increased the EC from 220 (25 mg Si per kg) to 228 (200 mg Si per kg), with pH 7.2 in all treatments and OC mostly stable around 0.99. Combined Cd and Si treatments further increased EC up to 235, with pH increasing to 7.3 and OC remaining fairly consistent around 0.97–0.99, indicating the combined impact on soil chemistry (Fig. 1C).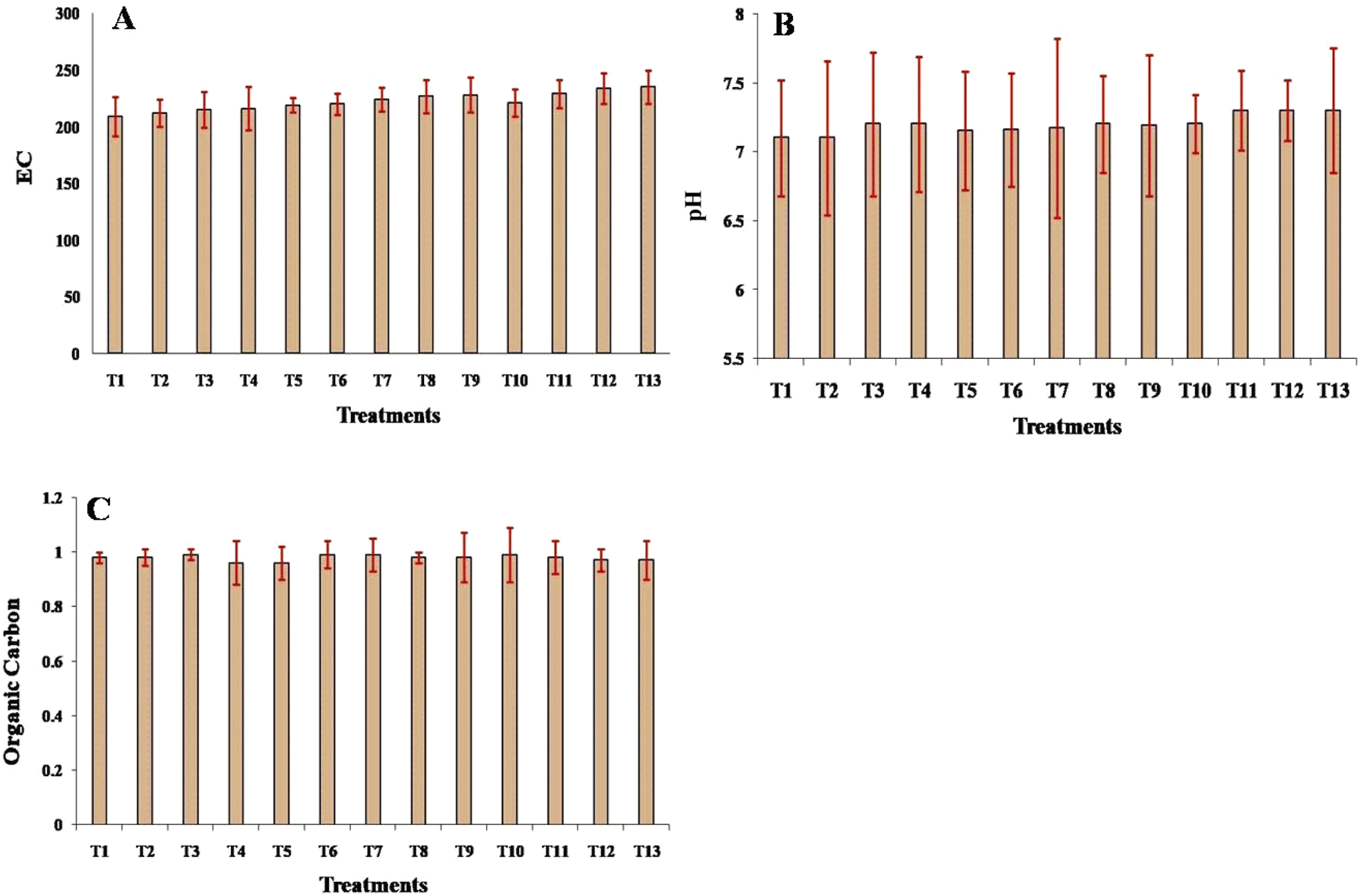 | ||
| Fig. 1 Effect of different treatment on (A) EC, (B) Ph, and (C) organic content of the soil. Means of three replicates ± SEs. *Bar symbol indicates T1 = control (untreated), T2 = 25 mg Cd per kg soil, T3 = 50 mg Cd per kg soil, T4 = 100 mg Cd per kg soil, T5 = 200 mg Cd per kg soil, T6 = 25 mg Si per kg soil, T7 = 50 mg Si per kg soil, T8 = 100 mg Si per kg soil, T9 = 200 mg Si per kg soil, T10 = 25 mg Cd per kg soil + 25 mg Si per kg soil, T11 = 50 mg Cd per kg soil + 50 mg Si per kg soil, T12 = 100 mg Cd per kg soil + 100 mg Si per kg soil, and T13 = 200 mg Cd per kg soil + 200 mg Si per kg soil. | ||
Root and shoot lengths
Under Cd stress, an increase in the length of the root was recorded although it is significant (p ≤ 0.05) at 100 and 200 mg Cd, respectively. A non-significant change in the root length was observed at 25, 50, and 100 mg Si per kg soil, although the increase in the root length was significant at 200 mg Si per kg soil. The data indicate that the root length increases with higher concentrations of Cd and Si, the highest root length being observed at 200 mg Cd + Si per kg soil. Under Cd stress, a significant (p ≤ 0.05) reduction in plant growth in terms of shoot length was noted, with the maximum reduction at 200 mg Cd concentration. Plants under Cd + Si treatment showed reduction, although it was lower when treated separately with Cd only; however, the reduction was non-significant up to 100 mg concentration, although a significant increase was recorded at 200 mg Cd + Si concentration (Fig. 2A). | ||
| Fig. 2 Effect of different treatments on (A) root and shoot length, (B) root and shoot dry and fresh mass ratio, and (C) leaf dry and fresh mass ratio of muskmelon plants. Means of three replicates ± SEs. *Treatment details as mentioned in Fig. 1. | ||
Root, shoot, and leaf dry and fresh mass ratio
It has been observed that root, shoot, and leaf dry and fresh mass ratio significantly (p ≤ 0.05) increases at all concentrations of Cd, compared to control plants. However, a non-significant change in root, shoot, and leaf dry-to-fresh mass ratio was recorded when the plants were treated with different concentrations of Si. However, a non-significant change in root dry-to-fresh mass ratio was noted under all concentrations of Cd and Si joint treatment. Although in the case of shoot and leaf dry and fresh mass ratio, a non-significant (p ≤ 0.05) increase was noted up to 50 mg Cd + Si concentration, but at 100 and 200 mg Cd + Si concentration, the increase was significant (Fig. 2B and C).Relative water content (RWC) and electrolyte leakage (EL)
The RWC decreased as the cadmium concentration increased, and the reduction was non-significant (p ≤ 0.05) at 25 mg Cd concentration. However, a significant (p ≤ 0.05) reduction compared to the control was noted up to 200 mg Cd concentration. Although, under Si treatment, a positive change in the RWC was recorded, which increased with respect to the control, these changes were non-significant. When treated with Cd + Si together, compared to the control, the change was non-significant up to 50 mg Cd + Si concentration; however, it was significant (p ≤ 0.05) at 100 and 200 mg concentration of Cd + Si. Electrolyte leakage (EL) significantly (p ≤ 0.05) increased with higher cadmium (Cd) concentrations, from 23.5 at 25 mg Cd per kg soil to 46.9 at 200 mg Cd per kg soil. Silicon (Si) treatments reduced EL, while combined Cd and Si treatments showed mitigated EL, with 19.8 at 25 mg Cd + Si per kg soil and 32.3 at 200 mg Cd + Si per kg soil (Fig. 3A).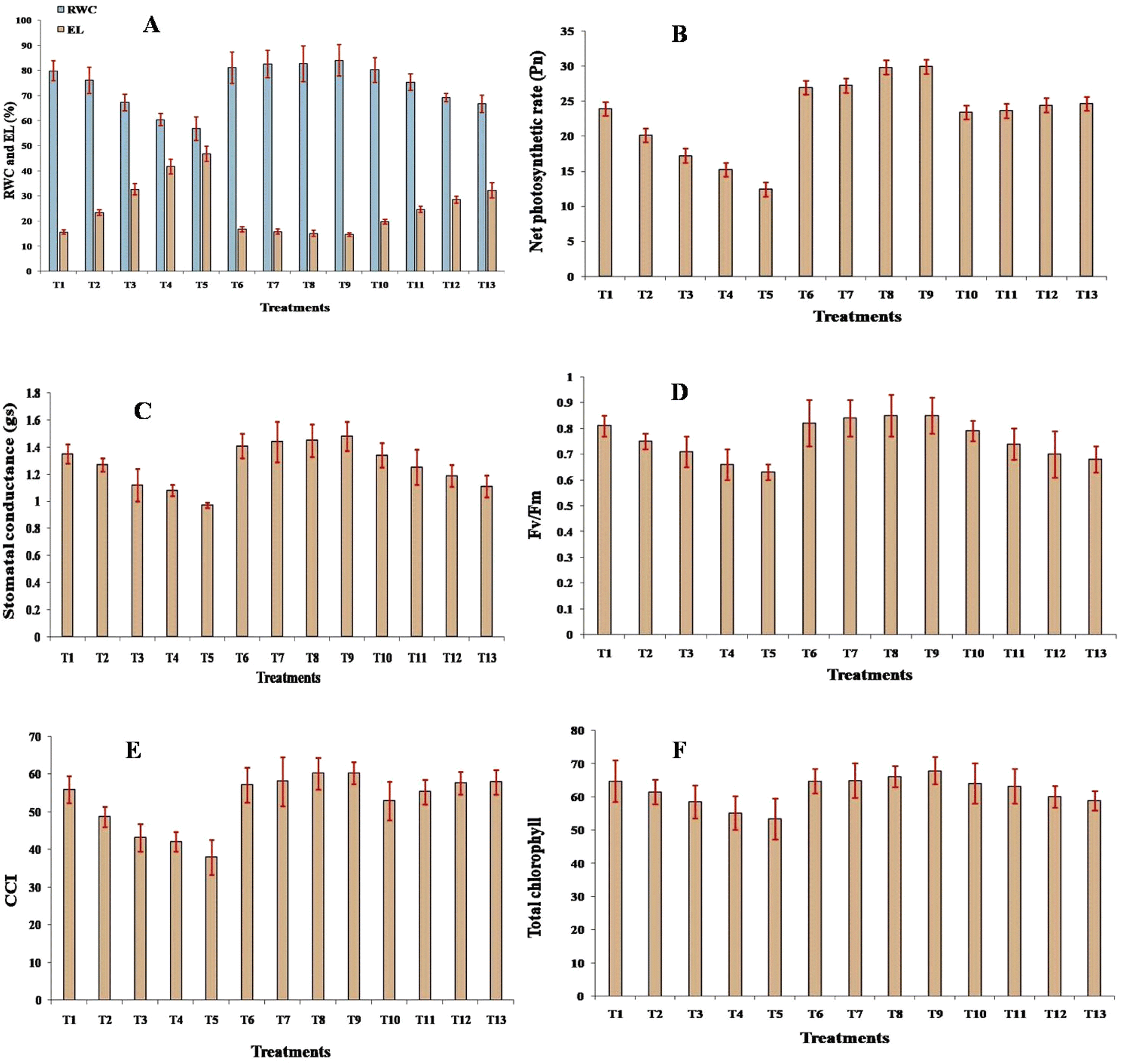 | ||
| Fig. 3 Effect of different treatments on (A) relative water content (RWC) and electrolyte leakage (EL), (B) net photosynthetic rate (Pn), (C) stomatal conductance (gs), (D) Fv/Fm, (E) chlorophyll color index (CCI), and (F) total chlorophyll content (mg g−1) in leaves of muskmelon plants. Means of three replicates ± SEs. *Treatment details as mentioned in Fig. 1. | ||
Photosynthesis parameters
The net photosynthetic rate (Fig. 3B) significantly (p ≤ 0.05) decreased with increasing cadmium (Cd) concentrations, compared to the control. Conversely, silicon (Si) treatments increased photosynthetic rates, while combined Cd and Si treatment resulted in a significant (p ≤ 0.05) increase when compared with Cd treatment alone. Stomatal conductance decreased significantly (p ≤ 0.05) with increasing cadmium (Cd) concentrations, from 1.27 to 0.97, compared to the control (1.35). Conversely, silicon (Si) treatments significantly improved stomatal conductance (Fig. 3C). Combined Cd and Si treatments show mitigated reductions in stomatal conductance when compared with individual treatments with Cd. Fv/Fm values decreased with increasing Cd concentrations, from 0.75 to 0.63; the reduction was non-significant at 25 mg concentration, beyond which it was significant (p ≤ 0.05). Conversely, silicon (Si) treatments increased Fv/Fm values, although the increase was non-significant when compared with control, while it was significant (p ≤ 0.05) when compared to respective individual CD treatments. Combined Cd and Si treatments showed mitigated reductions in Fv/Fm values, 0.79 to 0.68, although the reduction was significantly lower when compared with Cd treatment alone (Fig. 3D).The Chlorophyll Content Index (CCI) decreased significantly (p ≤ 0.05) with increasing cadmium (Cd) concentrations, compared to the control. Conversely, silicon (Si) treatments significantly (p ≤ 0.05) increased the CCI (Fig. 3E), while combined Cd and Si treatments showed improved CCI values compared with individual Cd treatments. The total chlorophyll content (Fig. 3F) and carotenoid levels decreased significantly (p ≤ 0.05) with increasing cadmium (Cd) concentrations. The total chlorophyll content dropped from 61.5 (25 mg Cd per kg soil) to 53.4 (200 mg Cd per kg soil), while carotenoid levels declined from 1.82 to 1.55, compared to the control (64.8 for chlorophyll and 1.87 for carotenoids). Silicon (Si) treatments enhanced both the chlorophyll and carotenoid content, while combined Cd and Si treatments showed mitigated reductions.
Hydrogen peroxide, proline, and MDA contents
The result showed that the H2O2, malondialdehyde (MDA), and proline levels increased significantly (p ≤ 0.05) with the increase in cadmium (Cd) concentrations (Fig. 4A–C). For instance, the H2O2 level increased from 22.4 to 45.4 compared to the control (13.2). MDA level increased from 1.32 to 3.24 and proline content from 53.2 to 144.5, indicating stress-induced oxidative damage and proline accumulation under Cd stress. In contrast, silicon (Si) treatments decreased H2O2, MDA, and proline levels, although these changes were non-significant when compared with control and significant (p ≤ 0.05) when compared with respective treatments. Combined Cd and Si treatments mitigated these increases, indicating Si involvement in alleviating Cd-induced oxidative damage. | ||
| Fig. 4 Effect of different treatments on (A) hydrogen peroxide in μmol per g (H2O2), (B) MDA content in μmol g−1, and (C) proline content in μg g−1 in leaves of muskmelon plants. Means of three replicates ± SEs. *Treatment details as mentioned in Fig. 1. | ||
Antioxidant enzyme activity
Activities of all enzymes, including SOD, CAT, APX, GR, and POD, increased significantly (p ≤ 0.05) with the increase of Cd concentration from 25 mg to 200 mg, with the maximum value achieved at 200 mg concentration. Under Si treatment, SOD and CAT activities changed non-significantly when compared with control, although the change was significant when compared with the respective treatment of Cd. Similarly, APX, GR, and POD activities changed non-significantly up to 50 mg of Si treatment, compared with control, although the change was significant with respect to the same concentration of Cd. Moreover, significant change was recorded for APX, GR, and POD activities under 100 and 200 mg Si treatment, with respect to control and similar treatment of Cd. Under combined treatment of Cd + Si, SOD, CAT, APX, GR, and POD activities increased significantly with an increase in the concentration of Cd + Si, and it was significantly (p ≤ 0.05) higher in the case of all the enzymes at 200 mg concentration. However, the increase in the enzyme activity was much lower compared with the activity of all the enzymes when treated with Cd alone and higher with respect to Si only treatment (Fig. 5A–E).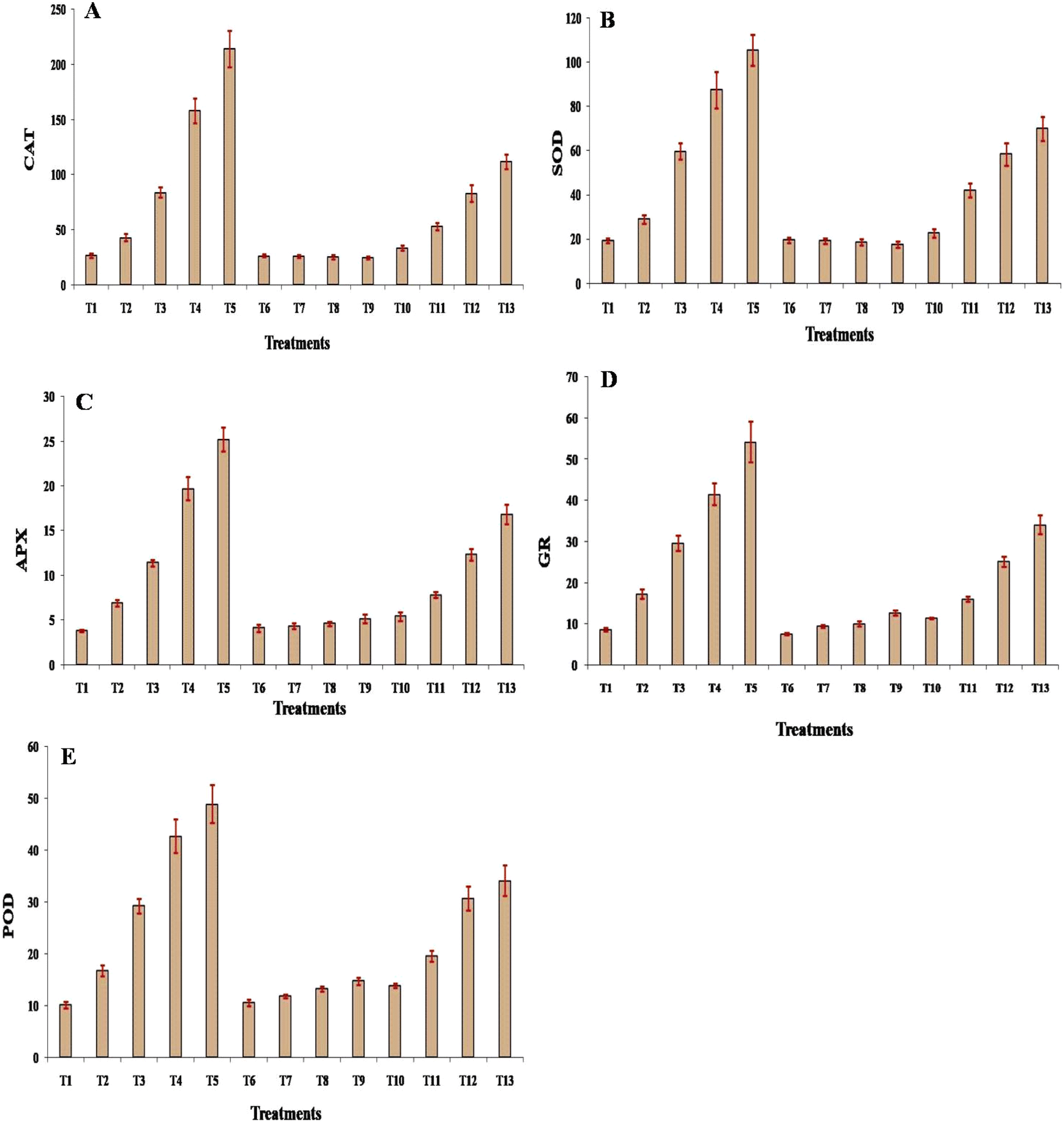 | ||
| Fig. 5 Effect of different treatments on activities of key antioxidant enzymes: (A) catalase [μmol (H2O2 oxidized) per mg (protein) per min] (CAT), (B) superoxide dismutase [units per mg (protein)] (SOD), (C) ascorbate peroxidase [μmol (ascorbate) per mg (protein) per min] (APX), (D) glutathione reductase [μmol (NADPH) per mg (protein) per min] (GR) and, (E) guaiacol peroxidase [μmol (H2O2 reduced) per mg (protein) per min] (POD) in leaves of muskmelon plants. Means of three replicates ± SEs. *Treatment details as mentioned in Fig. 1. | ||
Element concentration
Fig. 6A–E present the concentrations of various elements (Na, K, Mg, Fe, and Ca) in leaf tissues treated with different concentrations of cadmium (Cd) and silicon (Si), individually and in combination. With an increase in Cd concentration, Na, K, Fe, and Ca, concentration rises significantly (p ≤ 0.05), with maximum rise noted under 200 mg Cd concentration, with respective values, 2876, 62, 563, and 532, the increase in the concentration of elements under all concentration of Cd treatment was significant (p ≤ 0.05). However, a significant (p ≤ 0.05) reduction was recorded under all concentrations of Cd treatment in the case of the Mg element. Under 25 mg of Si treatments, the concentration of Na, K, Fe, and Ca increased non-significantly, while the Mg concentration decreased non-significantly, with respect to the control. At 50, 100, and 200 mg concentration of Si, the concentration of Na, K, Mg, Fe, and Ca increased significantly (p ≤ 0.05) with respect to the control, although the reduction in the concentration of K and Mg was recorded under 200 mg Si treatment. Under the combined treatments of Cd + Si, with concentrations of 25 mg, 50 mg, 100 mg, and 200 mg, a significant (p ≤ 0.05) increase in the concentration of Na, K, Mg, Fe, and Ca was recorded, and it was significantly (p ≤ 0.05) higher with respect to the values under the Si treatment of same concentration, while their values were significantly lower when compared with the concentration under Cd treatment of similar concentration.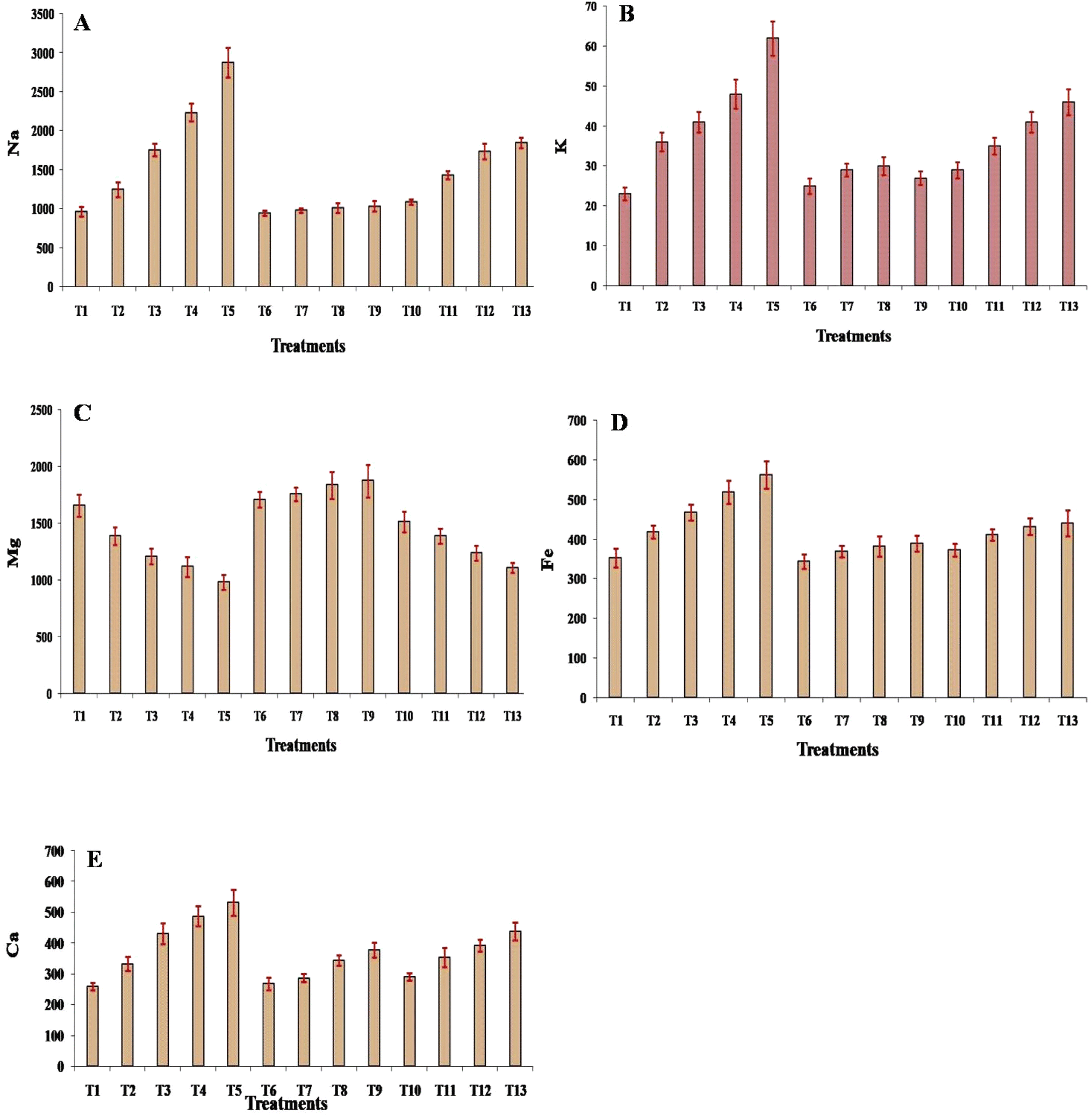 | ||
| Fig. 6 Effect of different treatments on the concentration of solutes: (A) Na, (B) K, (C) Mg, (D) Fe, and (E) Ca in the leaves of muskmelon plants. Means of three replicates ± SEs. *Treatment details as mentioned in Fig. 1. | ||
Gene expression changes
Expression of antioxidative enzyme related genes CAT, SOD, APX, GR, and POD was analyzed using RT-PCR. Under different concentrations of Cd stress, the gene expression of all the genes was significantly (p ≤ 0.05) increased with respect to the values under the control condition. Under 25 mg, 50 mg, 100 mg, and 200 mg Cd treatment, the gene expression of CAT, SOD, APX, GR, and POD was in the range 1.88–7.88, 1.23–4.43, 2.25–5.69, 2.25–5.67 and 1.98–5.33, respectively, with the maximum value under 200 mg Cd treatment in the case of all the enzymes. It has been noted that under all the treatments of Si, viz., 25 mg, 50 mg, 100 mg, and 200 mg, the expression of CAT, SOD, APX, GR, and POD did not change significantly, only slight changes were recorded, and the values were very close to the values under the control conditions. When the plants were treated with Cd + Si together at concentrations of 25 mg, 50 mg, 100 mg, and 200 mg, a significant (p ≤ 0.05) increase in the expression of CAT, SOD, APX, GR, and POD genes was recorded; however the increase in the gene expression level was comparatively lower in the case of individual Cd treatment (Fig. 7A–E).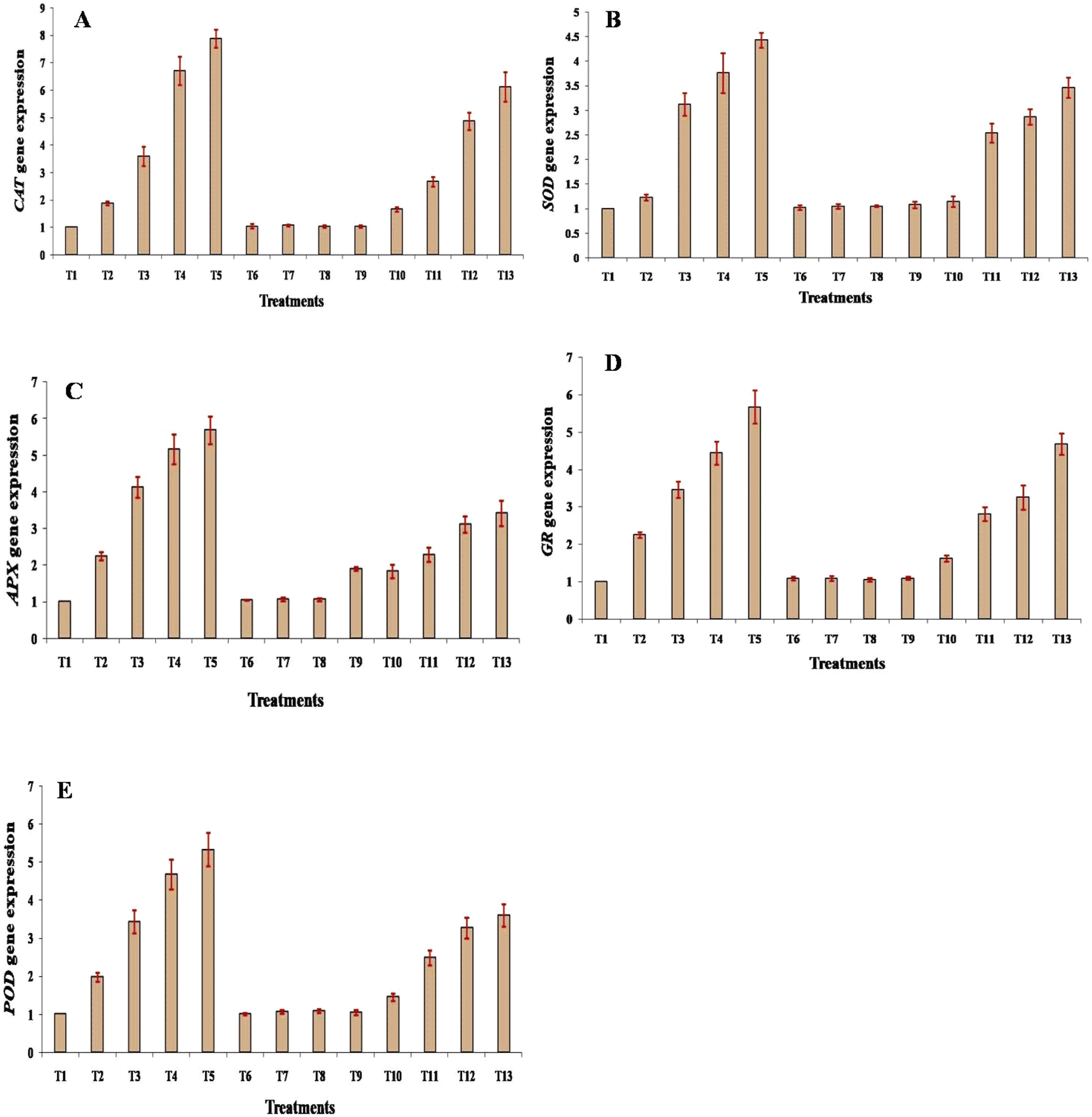 | ||
| Fig. 7 Effect of different treatments on the relative expression of antioxidative enzyme genes: (A) catalase (CAT), (B) superoxide dismutase (SOD), (C) ascorbate peroxidase (APX), (D) glutathione reductase (GR), and (E) guaiacol peroxidase (POD) in leaves of muskmelon plants. Means of three replicates ± SEs. *Treatment details as mentioned in Fig. 1. | ||
Multivariate analysis
The correlation matrix presented in the figure shows relationships between various physio-biochemical and molecular parameters in muskmelon (Fig. 8) under Cd, Si, and Cd + Si treatments. Most of the parameters recorded were significantly (p ≤ 0.05) correlated either negatively or positively. However, soil parameters EC and pH were non-significantly (p ≤ 0.05) correlated with all the parameters except for the root length and total carotenoid content. The RWC and Pn showed strong positive correlations with silicon (Si) treatment. Enzyme activities like SOD, CAT, APX, GR, and POD were positively correlated with H2O2 and MDA. Significant negative correlations between Cd treatments and parameters like RWC and photosynthetic parameters were recorded. Mg concentration showed a negative correlation with Cd treatments. The Principal Component Analysis (PCA) biplot (Fig. 9) visualizes the distribution of different treatments and variables, showcasing the variance explained by the first two principal components (PCA 1 and PCA 2). PCA 1 accounts for 85.06% of the variance, while PCA 2 accounts for 9.53% of the variance. Most of the parameters observed particularly all the enzymes, and their gene expression was found on the right side of the plot, which grouped with high concentration of Cd or Cd + Si, showing a positive correlation.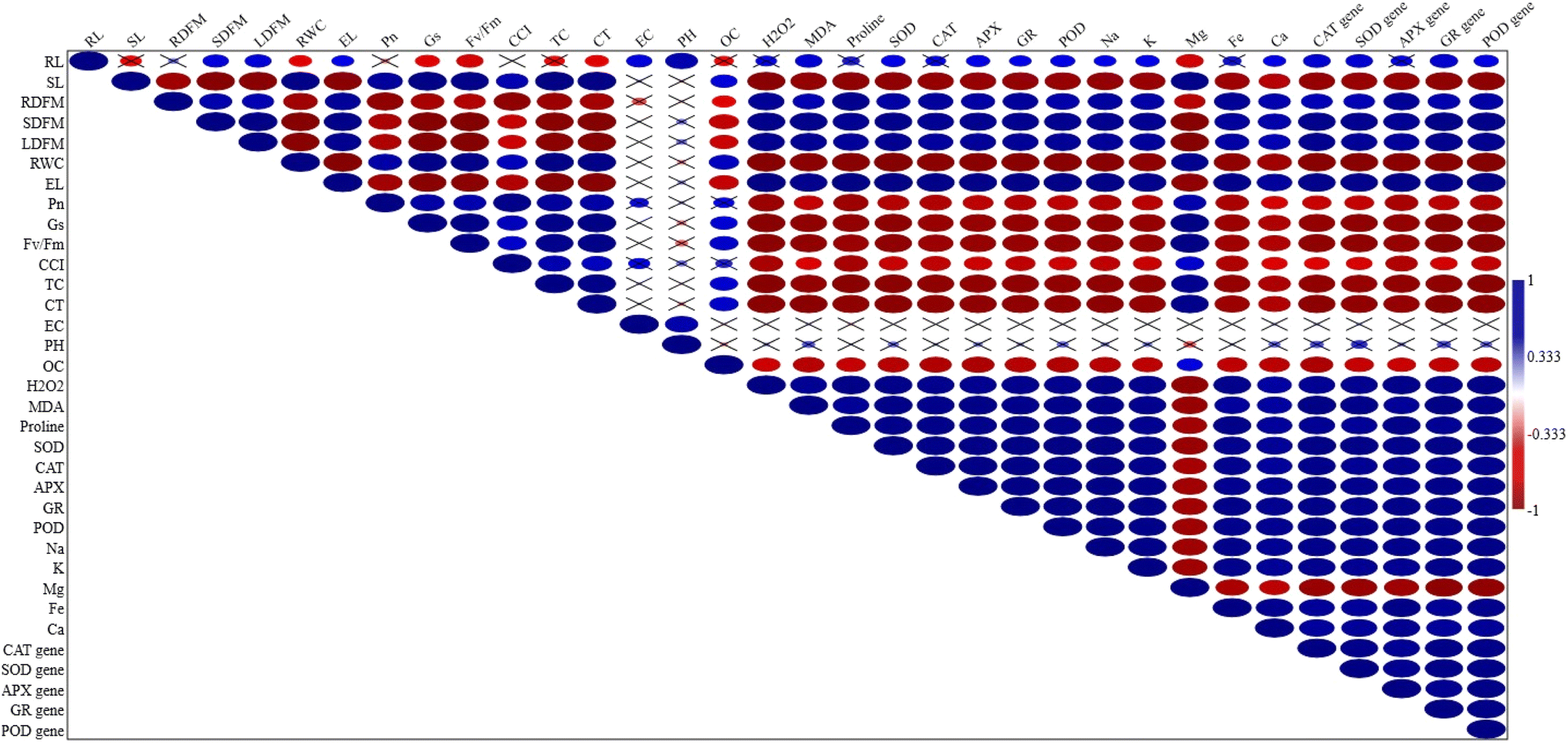 | ||
| Fig. 8 Pearson correlation analysis among the parameters analysed under different treatments in muskmelon plants. | ||
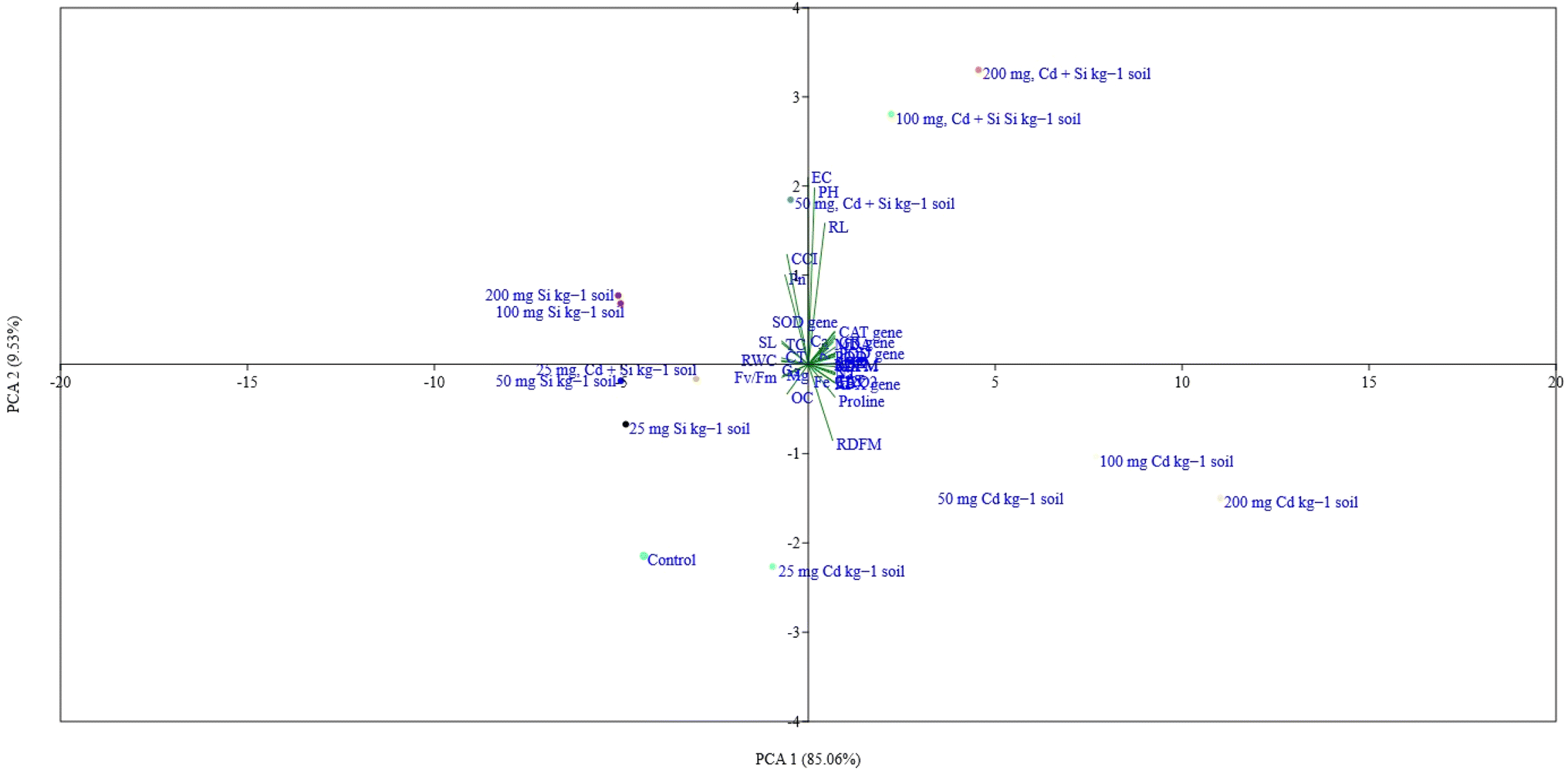 | ||
| Fig. 9 Principal component analysis among the parameters and under different treatments in muskmelon plants. | ||
Discussion
Silicon (Si) has an important role in alleviating cadmium (Cd) toxicity in plants by different modes of action; it decreases the absorption of Cd by plants, thereby restricting Cd accumulation in different plant tissues. The mechanisms are mediated by the regulation of gene/s associated with the lignin biosynthetic pathway and hormone signal transduction in plants. Si helps in mitigating Cd-induced oxidative stress by enhancing antioxidative enzyme activities; in addition to this, Si also enhances the plants' mineral nutrient uptake, which is helpful in retaining plant growth attributes under Cd toxicity. Furthermore, it helps in the regulation of water balance in plants, which can improve the plant resistance to Cd toxicity.40,41 It has been reported that the individual and combined Cd and Si utilization affected the EC, pH, and OC of soil to different degrees, although the changes were non-significant. The soil EC is one of the most important parameters that indicate the salinity and ionic concentration of the soil. In the present study, Cd treatments slightly increased the soil EC with no significant (p ≤ 0.05) difference over the control, similar to other studies in which Cd contamination resulted in enhanced soil salinity.42,43 In this study, the Si application mostly at higher concentrations, was reported to slightly increase the soil ionic concentration, although the changes were found to be non-significant statistically. The result is similar to previous findings, which suggests that Si amendments are helpful in plant growth and in slightly modifying the ionic balance of soil.44 The mixed Cd and Si application resulted in a collective effect on EC. The Cd and Si combination might be increasing the ionic exchange capacity of the soil, further enhancing the EC, as reported by Wang,45 where the Si application in contaminated soils influenced the absorption and mobility of various ions, including heavy metals. In the present study, we found that the pH of the soil largely remained unchanged across all the Cd treatments; it also suggests that Cd may have little to no direct impact on soil pH. Our findings are in support of the finding of Tong,46 in which they found that Cd application at lower concentrations has no significant impact on soil pH. On the other hand, the treatment of Si slightly increased the pH of the soil. The pH increment by Si application is well documented as it affects the soil buffering capacity, generally inducing a slightly alkaline condition.47 Importantly, the mixed Si and Cd application displayed a more pronounced pH increment, which was up to 7.3. The results revealed that Si might mitigate the Cd acidifying effects, or probably enhance the soil neutralization capacity. Previous studies also suggested this synergistic effect in which Si is supposed to modify other elements' impacts by affecting the base cations such as magnesium and calcium.48However, in this study, we found a marginal reduction in soil OC, suggesting that applied concentrations of Cd did not significantly affect the soil OC. The Si treatments, on the other hand, sustained the soil OC up to 0.99, revealing that Si has no negative impact on soil OC. Li49 concluded that Si can increase soil organic matter stability by altering the activity of microbes and promoting the OC concentration in soils, which could state the OC level stability in the present study. The Cd and Si mixed application maintained the OC level. This steadiness indicates that Si might play an OC stabilizing role in the presence of Cd, significantly countering the Cd adverse impact of soil organic matter. Ma22 also reported similar results, where Si minimized the heavy metal's negative impacts on soil organic matter. The results of this study provide significant insights into the effect of Cd stress and Si application on plant growth attributes, especially with respect to shoot and root growth. Cd is very toxic and is known to check plant growth attributes, mainly by affecting the physiology at the cellular level and the uptake of nutrients. Anyhow, the Si application is identified to play a role in abiotic stress alleviation and appears to mitigate Cd negative effects. The results of the present study are similar to the findings of Li,50 in which they found elongated roots under metal stress to improve the capacity of plants to cope with stressors. The Si application, on the other hand, displayed a non-significant increment in the height of the plant up to 100 mg Si per kg soil, but at 200 mg Si per kg soil a significant height increment was reported. This reveals that the application of Si may increase the growth of shoots, especially at higher Si concentrations. Si has been proven to improve plant's cellular mechanical properties, enhance resistance against oxidative stress, and balance different metabolic pathways, which can participate in better growth of shoots under abiotic stresses.51
The results reveal that while Cd application individually enhances EL and decreases the RWC, Si application helps to sustain these properties. The mixed Cd and Si application displays a synergistic effect, further sustaining EL and RWC, pointing out that Si can cope with Cd-induced stress in plants. These results support the potentiality of Si as an important tool in Cd stress contamination management in wheat, ensuring sustainable plant health and productivity.52 The results demonstrate that Cd stress notably damages the physiological functions of plants, including Pn, gs, CCI, total chlorophyll and carotenoid levels, and Fv/Fm values. The Cd alone treatment significantly (p ≤ 0.05) declines these parameters, pointing to the negative effect of Cd on plant health and efficiency of photosynthesis. However, the Si application resulted in a significant (p ≤ 0.05) improvement in the above parameters. Si not only improved the stomatal conductance and net photosynthetic rate but also maintained Fv/Fm values, chlorophyll content, CCI, and carotenoid concentration.53 These positive improvements reveal that Si has an important role in coping Cd stress by elevating the function of chloroplast, protecting photosynthetic pigments, and maintaining membrane integrity in muskmelon. Furthermore, the mixed Cd and Si treatment proved the protective effect of Si, displaying that Si mitigated the negative impacts of Cd on plant physiology to a significant (p ≤ 0.05) extent. The combined treatments showed intermediary values between individual treatments of Cd and Si, pointing out that Si can efficiently counter the toxic impact of Cd, thereby improving the plant's overall health and yield performance.41 For example, in wheat, Cd-induced stress reduced the stomatal conductance, and photosynthetic rates were maintained by Si treatment, which increased the stability of chlorophyll and led to an increment in Fv/Fm values, resulting in overall better plant performance.54 In the same way, in cucumber, Si application helped maintain higher photosynthetic rates and caused lower electrolyte leakage, suggesting the importance of Si in cell membrane protection and improvement of water use efficiency.55 Spinach is very sensitive to heavy metal stress, and Si application is beneficial by causing significant (p ≤ 0.05) increment in chlorophyll and carotenoid quantities, showing enhanced growth and tolerance to stress.
This study displayed that Cd exposure causes potential oxidative stress in muskmelon crops, as pointed out by the enhanced levels of H2O2, MDA, and proline. The results are similar to the previous findings on various vegetable crops, like lettuce and spinach.56 Si treatment potentially reduces Cd induced oxidative stress by reducing the H2O2, MDA, and proline levels in plants. In addition to this, the protective role of Si has been observed in different vegetables like cucumber, cabbage, and radish, in which antioxidant defense responses were improved by Si application which reduces oxidative damage.57–59 The mixed Cd and Si treatment was able to cope with oxidative stress, although not to baseline levels, stating that Si can partially alleviate the Cd adverse impact, improving muskmelon resilience against Cd toxicity. Cd toxicity induces potential oxidative stress in muskmelon,60 as proved by enhanced key antioxidative enzyme enhanced in the present study. Our findings are similar to the findings on different vegetable crops like tomato,61 cabbage,62 and radish,63 where an increment in antioxidative enzyme activity was also reported under Cd exposure. While Si alone did not potentially change the activities of the enzyme over the control, it adjusted the plant's response against Cd stress. In particular, Si significantly (p ≤ 0.05) enhanced APX, GR, and POD activities at higher doses (100 and 200 mg Si) under Cd treatment alone, pointing out that Si plays a protective role in minimizing oxidative damage. However, the increment in the activity of enzymes under the mixed application was lower than that reported over Cd alone and higher than that over Si alone treatment. This points out that Si mitigates oxidative stress induced by Cd. Similar reports have also been found in tomato and cucumber, in which mixed Si and Cd application resulted in improved activities of antioxidant enzymes over Cd alone, but the increment was not similar to the case of Si alone.64,65
The present study suggests that Cd exposure potentially affected the concentrations of key elements (Na, K, Mg, Fe, and Ca) in plants, with a notable increment in Na, K, Fe, and Ca and a reduction in Mg under Cd stress. The application of Si, especially at higher doses (50–200 mg Si), significantly increases the concentrations of the elements, revealing the importance of Si in modulating the uptake of the nutrient and mitigating Cd adverse impact on plant nutrition. The mixed Cd and Si application results in a significant (p ≤ 0.05) increment in the concentration of elements, with higher values over Si alone treatment but lower over Cd alone treatment. These results point out the significance of Si in the improvement of nutrient status and reducing the Cd detrimental impacts on vegetable crops. These results are similar to the findings on vegetables, such as lettuce, cucumber, cabbage, and tomato, where Si alleviates the heavy metal stress by improving the uptake and transport of nutrients.66 In crops such as cucumber and tomato, similar experiments also suggest that Si decreases the accumulation of Cd in the plant and improves the plant's overall health by sustaining the function of the membrane and absorption of nutrients.19 These findings point out that Si may decrease the uptake of Cd and toxicity by competition for the root binding site with Cd or by nutrient transport mechanism improvement, but it does not completely overcome the Cd induced interference.19
The Cd stress potentially upregulates the antioxidative enzyme gene expression in muskmelon, pointing to the plant's adaptation responses against oxidative damage caused by metal toxicity. The elevated CAT, SOD, APX, GR, and POD expression recorded under Cd exposure activates the antioxidative defense mechanisms of plants.67 This is supported by the studies where Cd exposure initiated a strong antioxidative response by upregulated gene expression and activity of enzymes.68 Though Si did not significantly activate the antioxidative enzyme gene expression under normal conditions and plays a protective role under Cd stress. Si has been proven to improve the antioxidative defense machinery, lowering the oxidative stress and improving plant sustainability against metal toxicity.67 Silicon additionally impacts gene expression associated with Cd absorption and transportation. The primary Cd carrier genes encode proteins that are members of the natural resistance associated macrophage protein, low affinity cation transporter, yellow stripe-like transporter, ZRT-like protein, IRT-like protein, P-type adenosine triphosphatase (ATPase, HMA), and cation diffusion facilitator families. Silicon attenuates the expression of Nramp5 in rice cells subjected to Cd contamination.69 The translocation of Cd to vesicles could work as an essential process for Cd detoxification. Si enhances the expression of OsHMA3 in the root system.70 Cd toxification elevates the expression of four genes (Nramp5, Nramp1, HMA2, and HMA3) responsible for encoding Cd transporter proteins in rice. The external application of Si reduces the expression of these four genes in response to Cd stress.71 Consequently, the application of Si can mitigate Cd-induced harm to plants by decreasing the absorption and translocation of Cd.
In this study, Si application, when mixed with Cd, displayed a moderate up-regulation in CAT, SOD, APX, GR, and POD gene expression over Cd alone. This points out that Si application can help in oxidative damage mitigation induced by Cd by up-regulating antioxidative gene expression, although to a lesser extent than the Cd alone treatments. These results are in support of previous findings in vegetable crops like cucumber, lettuce, and tomato where Si application reduced the oxidative stress induced by Cd by antioxidative enzyme activity improvement.72,73 The mixed Cd and Si application in the present study not only moderated the gene expression levels over Cd alone treatment but also pointed out that Si application might help in reducing Cd uptake in plant tissues, hence minimizing oxidative damage. This is in support of studies that show Si can minimize the uptake of heavy metals and mitigate their toxic impacts by improving the physio-biochemical properties of plants. The correlation matrix data highlight the intricate interactions between Cd and Si treatments and their impacts on physiological, biochemical, and molecular responses in the muskmelon plant. These findings highlight the protective role of Si in mitigating Cd-induced stress, improving antioxidant defense mechanisms, maintaining balanced nutrition and ultimately the overall plant health. This PCA biplot effectively demonstrates the complex interactions between different soil treatments and their effects on various physio-biochemical and molecular parameters in muskmelon. The significant variance explained by the first two principal components highlights the dominant effects of Cd and Si treatments on plant responses.
Conclusion
In conclusion, this study highlights the significant role of silicon (Si) in mitigating the toxic effects of cadmium (Cd) stress in muskmelon, a vegetable crop. Cd exposure leads to oxidative stress, nutrient imbalances, and reduced plant growth, which are alleviated by Si supplementation. Si enhances the antioxidative defense system by boosting the activity of key antioxidant enzymes and their respective gene expression and reducing oxidative damage. Si treatment with 200 mg Si per kg soil, showed maximum reduction in dry and fresh mass ratio, and it was 35%, 41% and 40% in case of root dry and fresh mass ratio, shoot dry and fresh mass ratio and leaf dry and fresh mass ratio, respectively. Additionally, Si helps maintain nutrient uptake, stabilizes key physiological parameters such as photosynthesis, and improves the overall plant growth under Cd stress. The combined application of Cd and Si shows a synergistic effect, further improving plant resilience. These findings suggest that Si could be a promising strategy to reduce Cd toxicity in muskmelon, which might be consequential in other cucurbits and vegetables, thereby improving crop productivity. Hence, Si fertilizers can be used in sustainable agriculture to mitigate HM toxicity and improve vegetable crop productivity. Further research is needed to better understand the molecular mechanisms underlying Si's protective role in muskmelon crops. In addition, studies should be carried out to validate the significance of Si in alleviating the negative impacts of Cd on other vegetable crops and under different environmental conditions.Data availability
All the data recorded during the experiments are presented in the manuscript.Author contributions
Conceptualization: Md Azizul Haque, Durgesh Kumar Jaiswal, and Waquar Akhter Ansari. Data curation: Mohammad Shahid and Waquar Akhter Ansari. Formal analysis: Mohammad Abul Farah, Md Azizul Haque and Waquar Akhter Ansari. Funding acquisition: Mohammad Abul Farah and Khalid Mashay Al-Anazi. Investigation: Ram Krishna, Waquar Akhter Ansari, Mohammad Shahid. Methodology: Ram Krishna. Project administration: Waquar Akhter Ansari and Durgesh Kumar Jaiswal. Resources: Waquar Akhter Ansari and Durgesh Kumar Jaiswal. Software: Mohammad Abul Farah, Md Azizul Haque, and Khalid Mashay Al-Anazi. Supervision: Waquar Akhter Ansari and Durgesh Kumar Jaiswal. Validation: Waquar Akhter Ansari. Visualization: Waquar Akhter Ansari. Writing – original draft: Ram Krishna, Akhilesh Yadav and Waquar Akhter Ansari. Writing – review & editing: Ram Krishna, Mohammad Abul Farah, Mohammad Shahid, Md Azizul Haque, Sudhakar Pandey, Durgesh Kumar Jaiswal, and Waquar Akhter Ansari.Conflicts of interest
All the authors declared that there is no competing interest including financial and personal which can influence the present research work.Acknowledgements
The authors would like to extend their sincere appreciation to the Researchers Supporting Project number (RSP2025R154), King Saud University, Riyadh, Saudi Arabia.References
- H. Chen, X. Huang, H. Chen, S. Zhang, C. Fan, T. Fu, T. He and Z. Gao, Effect of silicon spraying on rice photosynthesis and antioxidant defense system on cadmium accumulation, Sci. Rep., 2024, 14(1), 15265 CrossRef CAS.
- P. Mukherjee, J. Dutta, M. Roy, T. K. Thakur and A. Mitra, Plant growth-promoting rhizobacterial secondary metabolites in augmenting heavy metal (loid) phytoremediation: An integrated green in situ ecorestorative technology, Environ. Sci. Pollut. Res., 2024, 31(44), 55851–55894 CrossRef CAS.
- M. Rizwan, S. Ali, M. Z. ur Rehman, J. Rinklebe, D. C. Tsang, A. Bashir, A. Maqbool, F. M. Tack and Y. S. Ok, Cadmium phytoremediation potential of Brassica crop species: a review, Sci. Total Environ., 2018, 631, 1175–1191 CrossRef PubMed.
- T. El Rasafi, A. Oukarroum, A. Haddioui, H. Song, E. E. Kwon, N. Bolan, F. M. Tack, A. Sebastian, M. N. V. Prasad and J. Rinklebe, Cadmium stress in plants: A critical review of the effects, mechanisms, and tolerance strategies, Crit. Rev. Environ. Sci. Technol., 2022, 52(5), 675–726 CrossRef CAS.
- S. Pandey, M. Rai, H. C. Prasanna and G. Kalloo, ‘Kashi Madhu’: a new muskmelon cultivar with high total soluble solids, HortScience, 2008, 43(1), 245–246 Search PubMed.
- I. Martos-García, R. Fernández-Escobar and M. Benlloch-González, Silicon is a non-essential element but promotes growth in olive plants, Sci. Hortic., 2024, 323, 112541 CrossRef.
- H. Ashraf, F. Ghouri, J. Liang, W. Xia, Z. Zheng, M. Q. Shahid and X. Fu, Silicon Dioxide Nanoparticles-based amelioration of Cd toxicity by regulating antioxidant activity and photosynthetic parameters in a line developed from wild rice, Plants, 2024, 13(12), 1715 CrossRef CAS PubMed.
- S. Bhardwaj, D. Sharma, S. Singh, P. C. Ramamurthy, T. Verma, M. Pujari, J. Singh, D. Kapoor and R. Prasad, Physiological and molecular insights into the role of silicon in improving plant performance under abiotic stresses, Plant Soil, 2023, 486(1), 25–43 CrossRef CAS.
- A. Frew, L. A. Weston, O. L. Reynolds and G. M. Gurr, The role of silicon in plant biology: a paradigm shift in research approach, Ann. Bot., 2018, 121(7), 1265–1273 CrossRef CAS PubMed.
- I. Khan, S. A. Awan, M. Rizwan, S. Ali, M. J. Hassan, M. Brestic, X. Zhang and L. Huang, Effects of silicon on heavy metal uptake at the soil-plant interphase: A review, Ecotoxicol. Environ. Saf., 2021, 222, 112510 CrossRef CAS PubMed.
- S. Clemens and J. F. Ma, Toxic heavy metal and metalloid accumulation in crop plants and foods, Annu. Rev. Plant Biol., 2016, 67(1), 489–512 CrossRef CAS PubMed.
- A. Lux, Z. Lukačová, M. Vaculık, R. Švubová, J. Kohanová and M. Soukup, Silicification of root tissues, Plants, 2020, 9, 111 CrossRef CAS PubMed.
- F. U. Haider, C. Liqun, J. A. Coulter, S. A. Cheema, J. Wu, R. Zhang, M. Wenjun and M. Farooq, Cadmium toxicity in plants: Impacts and remediation strategies, Ecotoxicol. Environ. Saf., 2021, 211, 111887 CrossRef CAS PubMed.
- U. Zulfiqar, W. Jiang, W. Xiukang, S. Hussain, M. Ahmad, M. F. Maqsood, N. Ali, M. Ishfaq, M. Kaleem, F. U. Haider and N. Farooq, Cadmium phytotoxicity, tolerance, and advanced remediation approaches in agricultural soils; a comprehensive review, Front. Plant Sci., 2022, 13, 773815 CrossRef PubMed.
- N. Rhimi, M. Hajji, A. Elkhouni, M. Ksiaa, M. Rabhi, E. Lefi, A. Smaoui, K. Hessini, A. H. Hamzaoui, C. Cabassa-Hourton and A. Savouré, Silicon reduces cadmium accumulation and improves growth and stomatal traits in sea barley (Hordeum marinum Huds.) exposed to cadmium stress, J. Soil Sci. Plant Nutr., 2024, 1–17 Search PubMed.
- Z. Yang, F. Yang, J. L. Liu, H. T. Wu, H. Yang, Y. Shi, J. Liu, Y. F. Zhang, Y. R. Luo and K. M. Chen, Heavy metal transporters: Functional mechanisms, regulation, and application in phytoremediation, Sci. Total Environ., 2022, 809, 151099 CrossRef CAS PubMed.
- A. Shafiq, M. E. Munawar, M. Nadeem, A. Khan, G. H. Abbasi, M. A. U. Haq, M. A. Ayub, I. Iftikhar and M. Awais, Health risk assessment of bread wheat grown under cadmium and nickel stress and impact of silicic acid application on its growth, physiology, and metal uptake, Environ. Sci. Pollut. Res., 2024, 31(43), 55535–55548 CrossRef CAS PubMed.
- S. Pereira, A. Monteiro, J. Moutinho Pereira and L. T. Dinis, Silicon, an emergent strategy to lighten the effects of (a) biotic stresses on crops: A review, J. Agron. Crop Sci., 2024, 210(6), e12762 CrossRef CAS.
- L. Hou, S. Ji, Y. Zhang, X. Wu, L. Zhang and P. Liu, The mechanism of silicon on alleviating cadmium toxicity in plants: A review, Front. Plant Sci., 2023, 14, 1141138 CrossRef.
- A. S. Pereira, A. O. S. Dorneles, K. Bernardy, V. M. Sasso, D. Bernardy, G. Possebom, L. V. Rossato, V. L. Dressler and L. A. Tabaldi, Selenium and silicon reduce cadmium uptake and mitigate cadmium toxicity in Pfaffia glomerata (Spreng.) Pedersen plants by activation antioxidant enzyme system, Environ. Sci. Pollut. Res., 2018, 25, 18548–18558 CrossRef CAS.
- H. Hassan, S. H. Elaksher, S. Shabala and B. Ouyang, Cadmium uptake and detoxification in tomato plants: Revealing promising targets for genetic improvement, Plant Physiol. Biochem., 2024, 108968 CrossRef CAS.
- C. Ma, K. Ci, J. Zhu, Z. Sun, Z. Liu, X. Li, Y. Zhu, C. Tang, P. Wang and Z. Liu, Impacts of exogenous mineral silicon on cadmium migration and transformation in the soil-rice system and on soil health, Sci. Total Environ., 2021, 759, 143501 CrossRef CAS PubMed.
- L. Jia, Y. Song, F. You, S. Wang, X. Liu, L. Huang and L. Wang, Ameliorating the detrimental effects of chromium in wheat by silicon nanoparticles and its enriched biochar, Sci. Total Environ., 2024, 950, 175270 CrossRef CAS.
- A. Kausar, M. Y. Ashraf and I. Ali, et al., Evaluation of sorghum varieties/lines for salt tolerance using physiological indices as screening tool, Pakistan J. Bot., 2012, 44, 47–52 CAS.
- W. A. Ansari, N. Atri, B. Singh, P. Kumar and S. Pandey, Morpho-physiological and biochemical responses of muskmelon genotypes to different degree of water deficit, Photosynthetica, 2018, 56(4), 1019–1030 CrossRef CAS.
- W. A. Ansari, R. Krishna, S. P. Kashyap, K. Alanzi, M. Abul Farah, D. K. Jaiswal, A. Yadav, M. T. Zeyad and J. P. Verma, Relevance of Plant growth-promoting bacteria (PGPB) in reducing the severity of tomato wilt caused by Fusarium oxysporum f. sp. lycopersici by altering metabolites and related genes, Front. Microbiol., 2025, 15, 1534761 CrossRef.
- L. S. Bates, R. P. A. Waldren and I. D. Teare, Rapid determination of free proline for water-stress studies, Plant Soil, 1973, 39, 205–207 CrossRef CAS.
- S. Jana and M. A. Choudhuri, Glycolate metabolism of three submersed aquatic angiosperms during ageing, Aquat. Bot., 1982, 12, 345–354 CrossRef CAS.
- W. Weisany, Y. Sohrabi, G. Heidari, A. Siosemardeh and K. Ghassemi-Golezani, Changes in antioxidant enzymes activity and plant performance by salinity stress and zinc application in soybean (Glycine max L.), Plant Omics, 2012, 5(2), 60–67 CAS.
- O. H. Lowry, N. J. Rosebrough, A. L. Farr and R. J. Randall, Protein measurement with the Folin phenol reagent, J. Biol. Chem., 1951, 193, 265–275 CrossRef CAS PubMed.
- W. F. Beyer and I. Fridovich, Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions, Anal. Biochem., 1987, 161, 559–566 CrossRef CAS PubMed.
- H. Lück, Catalase, in Methods of Enzymatic Analysis, Academic press, 1965, pp. 885–894 Search PubMed.
- Y. Nakano and K. Asada, Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts, Plant Cell Physiol., 1981, 22(5), 867–880 CAS.
- C. H. Foyer and J. M. Fletcher, Plant antioxidants: colour me healthy, Biologist, 2001, 48(3), 115–120 CAS.
- K. J. Livak and T. D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method, Methods, 2001, 25(4), 402–408 CrossRef CAS.
- K. Bhati, S. Aggarwal and S. Sharma, et al., Differential expression of structural genes for the late phase of phytic acid biosynthesis in developing seeds of wheat (Triticum aestivum L.), Plant Sci., 2014, 224, 74–85 CrossRef CAS PubMed.
- M. M. Antonijević and M. Marić, Determination of the content of heavy metals in pyrite contaminated soil and plants, Sensors, 2008, 8(9), 5857–5865 CrossRef PubMed.
- A. U. Haq, T. E. Bates and Y. K. Soon, Comparison of extractants for plant-available zinc, cadmium, nickel, and copper in contaminated soils, Soil Sci. Soc. Am. J., 1980, 44(4), 772–777 CrossRef CAS.
- M. Lu, Y. Liang, P. Lakshmanan, X. Guan, D. Liu and X. Chen, Magnesium application reduced heavy metal-associated health risks and improved nutritional quality of field-grown Chinese cabbage, Environ. Pollut., 2021, 289, 117881 CrossRef CAS PubMed.
- M. Hasanuzzaman, K. Nahar, T. I. Anee and M. Fujita, Exogenous silicon attenuates cadmium-induced oxidative stress in brassica napus l. by modulating AsAGSH pathway and glyoxalase system, Front. Plant Sci., 2017, 8, 1061–1070 CrossRef PubMed.
- T. Shah, Z. Khan, S. R. Khan, A. Imran, M. Asad, A. Ahmad and P. Ahmad, Silicon inhibits cadmium uptake by regulating the genes associated with the lignin biosynthetic pathway and plant hormone signal transduction in maize plants, Environ. Sci. Pollut. Res., 2023, 30(59), 123996–124009 CrossRef CAS.
- L. Wang, L. Qin, X. Sun, S. Zhao, L. Yu, M. Wang and S. Chen, Linking Bacterial Growth Responses to Soil Salinity with Cd Availability, Bull. Environ. Contam. Toxicol., 2022, 109(2), 286–297 CrossRef CAS.
- J. K. Sharma, N. Kumar, N. P. Singh and A. R. Santal, Phytoremediation technologies and their mechanism for removal of heavy metal from contaminated soil: An approach for a sustainable environment, Front. Plant Sci., 2023, 14, 1076876 CrossRef PubMed.
- K. Alharbi, E. M. Hafez, N. Elhawat, A. E. D. Omara, E. Rashwan, H. H. Mohamed and S. I. Gadow, Revitalizing Soybean Plants in Saline, Cd-Polluted Soil Using Si-NPs, Biochar, and PGPR, Plants, 2024, 13(24), 3550 CrossRef CAS PubMed.
- Y. Wang, W. Xing, X. Liang, Y. Xu, Y. Wang, Q. Huang and L. Li, Effects of exogenous additives on wheat Cd accumulation, soil Cd availability and physicochemical properties in Cd-contaminated agricultural soils: A meta-analysis, Sci. Total Environ., 2022, 808, 152090 CrossRef CAS PubMed.
- W. Tong, Y. Hamid, B. Hussain, M. Usman, A. Sher, L. Liu and X. Yang, The effect of single and composite additives on Cd and Pb mobility, speciation and accumulation in late rice (Oryza sativa L.) grown on contaminated soil, Appl. Ecol. Environ. Res., 2020, 18(3), 4207–4221 CrossRef.
- Silicon and Nano-Silicon in Environmental Stress Management and Crop Quality Improvement: Progress and Prospects, ed. H. Etesami, A. H. Al Saeedi, H. El-Ramady, M. Fujita, M. Pessarakli and M. A. Hossain, 2022 Search PubMed.
- Q. Dong, J. Fang, F. Huang and K. Cai, Silicon amendment reduces soil Cd availability and Cd uptake of two Pennisetum species, Int. J. Environ. Res. Public Health, 2019, 16(9), 1624 CrossRef CAS.
- N. Li, A. Feng, N. Liu, Z. Jiang and S. Wei, Silicon application improved the yield and nutritional quality while reduced cadmium concentration in rice, Environ. Sci. Pollut. Res., 2020, 27(16), 20370–20379 CrossRef CAS PubMed.
- T. Li, X. Yang, L. Lu, E. Islam and Z. He, Effects of zinc and cadmium interactions on root morphology and metal translocation in a hyperaccumulating species under hydroponic conditions, J. Hazard. Mater., 2009, 169(1–3), 734–741 CrossRef CAS.
- A. Alam, B. Hariyanto, H. Ullah, K. Salin and A. Datta, Effects of silicon on growth, yield and fruit quality of cantaloupe under drought stress, Silicon, 2021, 13, 3153–3162 CrossRef CAS.
- M. Rizwan, J. D. Meunier, J. C. Davidian, O. S. Pokrovsky, N. Bovet and C. Keller, Silicon alleviates Cd stress of wheat seedlings (Triticum turgidum L. cv. Claudio) grown in hydroponics, Environ. Sci. Pollut. Res., 2016, 23, 1414–1427 CrossRef CAS.
- R. Tobiasz-Salach, M. Mazurek and B. Jacek, Physiological, biochemical, and epigenetic reaction of maize (Zea mays L.) to cultivation in conditions of varying soil salinity and foliar application of silicon, Int. J. Mol. Sci., 2023, 24(2), 1141 CrossRef CAS PubMed.
- S. M. Howladar, S. A. Al-Robai, F. S. Al-Zahrani, M. M. Howladar and A. Y. Aldhebiani, Silicon and its application method effects on modulation of cadmium stress responses in Triticum aestivum (L.) through improving the antioxidative defense system and polyamine gene expression, Ecotoxicol. Environ. Saf., 2018, 159, 143–152 CrossRef CAS PubMed.
- T. Gou, L. Yang, W. Hu, X. Chen, Y. Zhu, J. Guo and H. Gong, Silicon improves the growth of cucumber under excess nitrate stress by enhancing nitrogen assimilation and chlorophyll synthesis, Plant Physiol. Biochem., 2020, 152, 53–61 CrossRef CAS.
- Y. Meng, L. Zhang, L. Wang, C. Zhou, Y. Shangguan and Y. Yang, Antioxidative enzymes activity and thiol metabolism in three leafy vegetables under Cd stress, Ecotoxicol. Environ. Saf., 2019, 173, 214–224 CrossRef CAS.
- A. Harızanova, Z. Zlatev and L. K. A. Universityplovdiv, Effect of silicon on activity of antioxidant enzymes and photosynthesis in leaves of cucumber plants (Cucumis sativus L.), Turkish Journal of Agricultural and Natural Sciences, 2014, 1(Özel Sayı-2), 1812–1817 Search PubMed.
- N. Li, K. Wang, Y. Lv, Z. Zhang, B. Cao, Z. Chen and K. Xu, Silicon enhanced the resistance of Chinese cabbage (Brassica rapa L. ssp. pekinensis) to ofloxacin on the growth, photosynthetic characteristics and antioxidant system, Plant Physiol. Biochem., 2022, 175, 44–57 CrossRef CAS.
- S. Bhardwaj, T. Verma, A. Raza and D. Kapoor, Silicon and Nitric Oxide-Mediated Regulation of Growth Attributes, Metabolites and Antioxidant Defense System of Radish (Raphanus sativus L.) under Arsenic Stress, Phyton, 2023, 92(3), 0031–9457 Search PubMed.
- Y. Cheng, L. Qiu, P. Shen, Y. Wang, J. Li, Z. Dai and Z. Zou, Transcriptome studies on cadmium tolerance and biochar mitigating cadmium stress in muskmelon, Plant Physiol. Biochem., 2023, 197, 107661 CrossRef CAS PubMed.
- C. Song, M. A. Manzoor, D. Mao, X. Ren, W. Zhang and Y. Zhang, Photosynthetic machinery and antioxidant enzymes system regulation confers cadmium stress tolerance to tomato seedlings pretreated with melatonin, Sci. Hortic., 2024, 323, 112550 CrossRef CAS.
- N. Jinadasa, D. Collins, P. Holford, P. J. Milham and J. P. Conroy, Reactions to cadmium stress in a cadmium-tolerant variety of cabbage (Brassica oleracea L.): is cadmium tolerance necessarily desirable in food crops?, Environ. Sci. Pollut. Res., 2016, 23, 5296–5306 CrossRef CAS.
- J. Xin, X. H. Zhao, Q. L. Tan, X. C. Sun, Y. Y. Zhao and C. X. Hu, Effects of cadmium exposure on the growth, photosynthesis, and antioxidant defense system in two radish (Raphanus sativus L.) cultivars, Photosynthetica, 2019, 57(4), 967–973 CrossRef CAS.
- Z. Liu, X. Wu, J. Yan, W. Fan, T. Li, S. Wang and P. Liu, Silicon reduces cadmium accumulation and toxicity by regulating transcriptional and physiological pathways, and promotes the early growth of tomato seedlings, Ind. Crops Prod., 2023, 206, 117720 CrossRef CAS.
- J. Wu, J. Guo, Y. Hu and H. Gong, Distinct physiological responses of tomato and cucumber plants in silicon-mediated alleviation of cadmium stress, Front. Plant Sci., 2015, 6, 453 Search PubMed.
- J. Pavlovic, L. Kostic, P. Bosnic, E. A. Kirkby and M. Nikolic, Interactions of silicon with essential and beneficial elements in plants, Front. Plant Sci., 2021, 12, 697592 CrossRef.
- H. Zhang and L. Lu, Transcription factors involved in plant responses to cadmium-induced oxidative stress, Front. Plant Sci., 2024, 15, 1397289 CrossRef.
- D. Moravčíková and J. Žiarovská, The effect of cadmium on plants in terms of the response of gene expression level and activity, Plants, 2023, 12(9), 1848 CrossRef.
- J. Ma, H. Cai, C. He, W. Zhang and L. Wang, A hemicellulose-bound form of silicon inhibits cadmium ion uptake in rice (Oryza sativa) cells, New Phytol., 2015, 206, 1063–1074 CrossRef CAS.
- Y. Cai, B. Pan, B. Liu, K. Cai, J. Tian and W. Wang, The cd sequestration effects of rice roots affected by different Si management in Cd-contaminated paddy soil, Sci. Total Environ., 2022, 849, 157718 CrossRef CAS.
- D. Chen, D. Chen, R. Xue, J. Long, X. Lin and Y. Lin, et al., Effects of boron, silicon and their interactions on cadmium accumulation and toxicity in rice plants, J. Hazard. Mater., 2019, 367, 447–455 CrossRef CAS.
- E. Pilon-Smits, Phytoremediation, Annu. Rev. Plant Biol., 2005, 56(1), 15–39 CrossRef CAS PubMed.
- F. Zhang, X. Wan, Y. Zheng, L. Sun, Q. Chen, X. Zhu and M. Liu, Effects of nitrogen on the activity of antioxidant enzymes and gene expression in leaves of Populus plants subjected to cadmium stress, J. Plant Interact., 2014, 9(1), 599–609 CrossRef.
Footnote |
| † These authors contributed equally and share joint first authorship. |
| This journal is © The Royal Society of Chemistry 2025 |