
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Sample-to-answer microfluidic device towards the point-of-need detection of Staphylococcus aureus enterotoxin genes in ruminant milk†
Maha
Shalaby
 *abc,
Valentina
Busin
a,
Xiaoxiang
Yan
b,
Seyda
Cengiz
d,
Mehmet Cemal
Adiguzel
e,
Jonathan M.
Cooper
b,
Taya
Forde
a and
Julien
Reboud
*b
*abc,
Valentina
Busin
a,
Xiaoxiang
Yan
b,
Seyda
Cengiz
d,
Mehmet Cemal
Adiguzel
e,
Jonathan M.
Cooper
b,
Taya
Forde
a and
Julien
Reboud
*b
aSchool of Biodiversity, One Health & Veterinary Medicine, University of Glasgow, Glasgow, UK. E-mail: 2427372S@student.gla.ac.uk
bJames Watt School of Engineering, University of Glasgow, Glasgow, UK. E-mail: Julien.reboud@glasgow.ac.uk
cFood Control Department, Faculty of Veterinary Medicine, Kafrelsheikh University, Kafr-El-Sheikh 33516, Egypt
dDepartment of Microbiology, Faculty of Veterinary Medicine, Sıtkı Koçman University, Muğla 48100, Turkey
eDepartment of Microbiology, Faculty of Veterinary Medicine, Ataturk University, Erzurum, Turkey
First published on 8th January 2025
Abstract
Milk is commonly screened both for indicators of animal disease and health, but also for foodborne hazards. Included in these analyses is the detection of Staphylococcus aureus, that can produce an enterotoxin, causing staphylococcal food poisoning (SFP), which often leads to sudden onset of significant gastrointestinal symptoms in humans. Epidemiological data on SFP are limited, particularly in low- and middle-income countries. Many conventional assays for the detection of staphylococcal enterotoxins rely on the detection of the genes coding for them, either directly in food samples or after bacterial culture. Currently, many of the nucleic acid-based methods used require specific expertise and equipment, whilst bacterial culture takes 24–48 hours; both are contributory factors that limit efforts either during food safety emergencies or routine screening. Here we present the development of a “sample-to-answer” isothermal nucleic acid loop-mediated amplification (LAMP) assay in a microfluidic device for the detection of Staphylococcus aureus enterotoxin genes in ruminant milk. A multiplex LAMP assay targeting two of the most prevalent S. aureus enterotoxin-encoding genes (A and B) was integrated into a microfluidic device combining simple 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :10 dilution for sample preparation and a lateral flow assay for easy readout. We achieved a limit of detection of 104 colony forming units per ml in spiked cow and goat milk samples, an order of magnitude more sensitive than the European recommendation for the maximum allowable presence of coagulase-positive staphylococci in raw milk. The assay showed no cross-reactivity in detecting other tested non-enterotoxigenic S. aureus strains or associated foodborne pathogens. The test integrated the simplicity of use of microfluidic devices with the sensitivity, specificity and rapidity of a nucleic acid-based assay, and a simple lateral flow readout to provide an appropriate device to ensure the safety of milk for human consumption. To illustrate its potential for point-of-need practical applications, the test was performed in agricultural settings in rural Turkey in a limited feasibility exercise.
:10 dilution for sample preparation and a lateral flow assay for easy readout. We achieved a limit of detection of 104 colony forming units per ml in spiked cow and goat milk samples, an order of magnitude more sensitive than the European recommendation for the maximum allowable presence of coagulase-positive staphylococci in raw milk. The assay showed no cross-reactivity in detecting other tested non-enterotoxigenic S. aureus strains or associated foodborne pathogens. The test integrated the simplicity of use of microfluidic devices with the sensitivity, specificity and rapidity of a nucleic acid-based assay, and a simple lateral flow readout to provide an appropriate device to ensure the safety of milk for human consumption. To illustrate its potential for point-of-need practical applications, the test was performed in agricultural settings in rural Turkey in a limited feasibility exercise.
1. Introduction
Staphylococcal food poisoning (SFP) occurs following the consumption of food containing sufficient amounts of one (or more) preformed staphylococcal enterotoxins (SEs).1,2 Symptoms can arise from as little as 0.5 ng of toxin per mL of milk.3 These enterotoxins are encoded by SE genes, which form part of the staphylococcal virulence gene repertoire,1,4 and are serologically classified into classical antigenic SEs (A-J) and the newly described group of SEs (K-U).5–7 Around 95% of reported SFP cases are caused by the classical staphylococcal enterotoxins,8,9 with SEA and SEB most commonly causing gastrointestinal symptoms.10 SFP symptoms comprise nausea, vomiting and abdominal cramping, with or without diarrhea, often with rapid onset (2–8 hours after exposure).11,12 It is usually a self-limiting disease, with requirement of hospitalization in some cases, particularly in children, the elderly and in people with co-morbidities.4,13S. aureus is an opportunistic pathogen that can be carried by or infect various hosts, including humans and domesticated ruminants (cows, buffalo, sheep and goats).14 It is commonly reported as a cause of clinical and sub-clinical mastitis (i.e., inflammation of the udder) in dairy animals.15–17 Humans are considered as the most common source of S. aureus contamination involved in SFP, however, animals with S. aureus carriage or active infections are also recognized as an important source of contamination with enterotoxigenic S. aureus, for instance in raw animal products (i.e., raw milk, milk products, and raw meat products).18,19
Staphylococcal growth and toxin production is commonly promoted by inadequate storage, cooking or handling, as well as cooling of food contaminated with enterotoxigenic S. aureus of human or animal origin.20 One of the common routes of SFP is consumption of milk and milk products, especially fresh dairy products.21 It is also noteworthy that although pasteurisation temperatures are sufficient to destroy the pathogen, such treatments do not inactivate the enterotoxins,22 which require prolonged boiling or autoclaving to gradually decrease their potency (Ortega et al. 2010). In the case of ultra heat-treated milk (UHT milk), the heating temperature should, theoretically, be sufficient, although the overall efficacy is affected by the toxin's initial concentration.23,24
Specific incidence data of SFP outbreaks are limited, with available reports from only some global subregions.25,26 The limitation of data availability is most problematic in low- and middle-income countries, most probably as many affected people do not seek medical attention. In addition to the lack of routine surveillance tools, the limited availability of clinical specimens and the complicated laboratory confirmation protocols all contribute to under-reporting.27 The number of SFP outbreaks reported in the European Union increased from 25 outbreaks between 2007 and 201128 to 117 outbreaks in 2019 and 202029,30 whilst in the United States, 42 outbreaks were recorded between 1993 and 199731 and 75 outbreaks between 2009 and 2015.32
According to data reported in our recent study, there has been a gradual change in the methods employed for the detection of S. aureus enterotoxins, where nucleic acid amplification tests (NAAT, identifying the genes) have been replacing the detection of the toxins themselves,26 opening up the potential for the development of new the point-of-need technologies.33,34 Within the context of the dairy value chain, detecting toxins or a risk of contamination either at the milking stage next to the animal, or within the dairy storage tank on the farm, would allow an early intervention to prevent the milk from entering the food chain. Such detection at the point-of-need is important for food safety.
Several assays have been developed for the detection of S. aureus enterotoxin genes, either directly from food samples using molecular tools or after an enrichment and/or culture step to increase bacterial concentrations.35 However, these assays are time-consuming,36 labor-intensive,35 and often require significant expertise and sophisticated equipment,37 limiting their application in food safety emergencies, such as SFP outbreaks.
Isothermal methods, such as loop-mediated isothermal amplification (LAMP), eliminate the need for thermal cycling, and can often be performed in a single step in a single tube.38 Laboratory-based LAMP assays have already been developed for the detection of enterotoxin genes of S. aureus, either as a single or multiplex assay, with a high analytical sensitivity (limit of detection (LOD) of 10 and 102 CFU ml−1, respectively) and specificity (100%).39,40 However, these assays still require access to a laboratory facility, which delays detection and subsequently the response to potential disease outbreaks.
To overcome the limitations of these laboratory-based assays, microfluidic technologies have emerged as portable, rapid and cost-efficient platforms that could be deployed at the point-of-need and thus provide near real-time responses to emergencies.41 Such methods have previously been applied to the detection of methicillin-resistant S. aureus; however these have still shown barriers for applicability in the field. For example, a study reporting a microfluidic-based PCR assay required pre-treatment of the sample to extract nucleic acids42 and a microfluidic LAMP-based assay required access to pneumatic and control systems to drive fluids through complex processes,43 preventing their wider application at the point-of-need.
With the goal of moving towards point-of-need applications, in this work, we have developed a prototype “sample-to-answer” LAMP-based microfluidic point-of-need device for the detection of S. aureus enterotoxin genes in ruminant milk (cow and goat). The test combines simple sample preparation steps (including direct dilution) integrated onto a microfluidic device, combined with incubation in a heat block and lateral flow strip detection (Fig. 1).
 | ||
| Fig. 1 Schematic diagram showing the process and structure of the prototype point-of-need test. (A) After adding the milk to the diluent chamber containing PCR-grade nuclease-free water, effecting a finger pump on the dilution chamber results in the sample being diluted and mixed with LAMP reagents present in the amplification chamber. (B) The amplification reaction occurs at 60 °C for 45 minutes. (C) The amplicons are labelled with biotin and fluorescein isothiocyanate (FITC) through the primers, leading to binding to the test line and streptavidin-coated beads for visualisation on a lateral flow strip. (D) The process was integrated into a simple microfluidic format, including dilution, amplification and read out, using finger pumps to move liquids through the chip. The lower three ovals with dark blue color represent the dilution chambers with the diluent, and those with pink color represent the dilution chambers after finger pumping the diluted sample to the amplification chamber. The upper three ovals with dark blue color represent amplification chambers with amplification reagents (master mix and LAMP primer mix) and with light blue color represent the mixture of diluted sample with amplification reagents. From left to right, the first strip is a negative control, showing only the control line, while the two strips for positive control and sample show both the control line and the test line, illustrating a positive test, i.e. indicating amplification of SEA and/or SEB (C and D). | ||
2. Materials and methods
2.1 Staphylococcus aureus reference strains: source and maintenance
Enterotoxigenic Staphylococcus aureus reference strains were purchased from the Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures (ESI† Table S1). To evaluate the specificity of the LAMP assay, reference strains of non-enterotoxigenic S. aureus (S. aureus 171.1), non-aureus staphylococcal strains (Staphylococcus epidermidis 49510 and Staphylococcus intermedius 63687) and non-staphylococcal species commonly causing food poisoning (Salmonella enteritidis 67199 and Bacillus cereus 48306) were sourced from Veterinary Laboratory Quality Assurance scheme provided by the Veterinary Laboratories Agency and identified by Scottish methicillin resistant S. aureus reference laboratory.Reference strains were grown on brain heart infusion (BHI) agar (Sigma Aldrich) at 37 °C overnight, then inoculated into BHI broth (Sigma Aldrich) (one colony for each tube) and incubated at 37 °C overnight. After adjusting concentrations (using optical density at 600 nm) to ∼1 OD600 using a spectrophotometer (Eppendorf Bio-photometer V1.35), they were ten-fold serially-diluted in sterile BHI broth, and colony counts were determined by conventional plate counting (using three 10 μl drops per dilution, and counting those drops with 5–50 colonies). Stocks from overnight BHI broth cultures of each reference strain were maintained in 15% glycerol from BHI broth cultures and stored at −80 °C.
2.2 Evaluation of specificity and sensitivity of LAMP assay for detection of Staphylococcus aureus enterotoxin A and B genes
The different steps taken for the evaluation, optimisation and transfer of LAMP assays for detection of S. aureus enterotoxin A and B genes (SEA and SEB) are summarized as a diagram in ESI† Fig. S1.The SEA gene LAMP primer set was based on published primers,40 with modifications to increase assay specificity. The new loop forward primer was designed using Primer Explorer V5web server (Eiken Chemical Co. LTD, Tokyo, Japan; http://primerexplorer.jp/e),44 based on the SEA sequence obtained from GenBank accession number M18970, and manual modification of the position of the F1 primer (ESI† Table S2 and Fig. S2). The SEB LAMP primer set was used as described40 (ESI† Table S2). The primers were BLASTed against the NCBI nucleotide collection and IDT OligoAnalyzer™ Tool was used to check for formation of hairpin, self-, and hetero-dimers. Artificial oligonucleotide dsDNA sequences (Integrated DNA technologies (IDT)) for each gene were designed manually for use as positive controls, mapping the target region of each primer set for both genes (ESI† Table S3).
Cross reactivity of SEA and SEB LAMP primers was evaluated against the artificial oligonucleotide dsDNA sequences of the other gene, using high (9.3 × 1010 and 1.78 × 1011 copies per μl) and low (9.3 and 1.78 copies per μl) concentrations (to ensure the accuracy of the assay, even when low concentrations of cross-reactants might be present) of SEA and SEB artificial dsDNA oligonucleotide sequences, respectively. Specificity of the LAMP assay for the detection of SEA and SEB genes was evaluated using DNA extracts of overnight cultures of reference strains from non-enterotoxigenic Staphylococcus aureus, non-aureus staphylococcal strains and other non-staphylococcal species commonly causing food poisoning (described in 3.1). The analytical sensitivity of the LAMP assay for the detection of SEA and SEB genes (LOD) was determined in two ways: (i) using DNA extracts of overnight cultures of the three S. aureus reference strains, which were subsequently diluted, and (ii) by preparing ten-fold serial dilutions of overnight cultures of the three S. aureus reference strains (S. aureus DSMZ 19041, S. aureus DSMZ 19044 and S. aureus DSMZ 20652), from which DNA was subsequently extracted. The LOD was identified as the lowest concentration detected for all the replicates (three biological and three technical replicates).
Genomic DNA for testing was prepared as follows: from each overnight BHI broth culture of each reference strain, cells were harvested from 1 ml by centrifugation at 9600 × g (10000 rpm) for 10 minutes and DNA was extracted using a MagaZorb® DNA Mini-Prep Kit (Promega) following the manufacturer's protocol. The quantity of DNA in the extract of each culture was determined using a Nanodrop 1000 Microvolume spectrophotometer (Thermofisher), and the quality of the DNA was confirmed by ensuring that it had an A260/A280 ratio >1.8.
2.3 Sample preparation optimisation
To evaluate the effect of dilution, instead of DNA extraction, as a simple sample preparation step, homogenized pasteurized whole cow and goat milk samples (purchased from commercial food outlets, on different occasions as biological replicates) were spiked with cells collected from overnight culture of S. aureus DSMZ 19041 reference strain (1 ml of the overnight culture was centrifuged at 1000 rpm for 10 minutes, the supernatant was discarded and the collected cells were resuspended in PBS and used to spike milk samples), then diluted (1:2, 1:4, 1:5 and 1:10); each dilution was tested with three biological and three technical replicates. Milk samples that had been spiked and diluted were used with and without DNA extraction to run LAMP assays.
In addition to diluting milk samples spiked with a single concentration of S. aureus, we also considered milk samples spiked with different concentrations of S. aureus. Homogenized pasteurized whole cow and goat milk samples were spiked with bacterial cells collected from ten-fold serially diluted overnight cultures of each S. aureus DSMZ 19041, 19044 and 20652 reference strains. Each overnight culture was serially diluted by adding 1 ml of the culture to 9 ml of sterile broth media. Bacterial cells from each dilution were collected by centrifugation at 1000 rpm for 10 minutes, the supernatant was discarded, and the cells were resuspended in PBS. These resuspended cells were then used to spike the milk samples. Following spiking, each milk sample with the corresponding bacterial concentration was further diluted 1:10 and directly used in the LAMP assay without any DNA extraction.
In a similar process as that described above for dilution, the following two approaches were assessed for their impact on analytical sensitivity: i) heating pre-treatment was added as another simple physical sample preparation step to explore the potential benefits for bacterial lysis, incubating at 95 °C for 10 minutes before running the assay; and ii) increasing the assay time (60 °C isothermal amplification) to 60 minutes instead of 30 minutes.
2.4 LAMP assay optimisation
The LAMP reaction for each gene (SEA and SEB) was carried out in a 20 μl reaction mixture containing 0.8 μl (10 pmol μl−1) of each inner primer (FIP and BIP), 0.2 μl of each outer primer (F3 primer and B3 primer), 1 μl (10 pmol μl−1) of each loop primer (LF and LB), 1 μl nuclease-free PCR-grade water (Invitrogen) and 15 μl of GspSSD2.0 Isothermal Master Mix (ISO_004) (OptiGene). The duplex LAMP assay was also carried out in a 20 μl reaction mixture containing half of the primers' volume from individual reactions. A volume of 5 μl of the sample under investigation (1:10 diluted spiked milk samples, or relevant controls) was added to each reaction (total volume 25 μl). The reaction mixture was incubated in a heat block (in tubes) for 30, 45 and 60 minutes, or a Rotor-Gene Q L2-0694-00 thermocycler (Qiagen), with an initial cycle of 60 °C for 20 seconds, followed by 30 cycles of 60 °C for 1 minute then termination at 95 °C for 10 minutes. Negative control reactions using nuclease-free PCR-grade water, PBS (used for reconstitution of collected cells from overnight cultures of reference strains), media (BHI broth) and cow/goat milk, as well as positive controls using artificial dsDNA oligonucleotide sequences, were prepared with each run to screen for the contamination of the reagents, and to confirm that the assay was working properly.
2.5 Statistical analysis
Statistical analysis was performed using SIGMAPLOT (v13.0), using a two-way ANOVA to compare the difference in amplification time associated with different factors including dilution, heating, assay time, enterotoxigenic S. aureus strains, single-plex and duplex assays, and milk sources (cow and goat). P-Value < 0.05 was considered significant based on the studentized range distribution (q value).2.6 Design, fabrication and manipulation of microfluidic device
Plastic microfluidic devices were designed using Inkscape (v 0.92.4) and FreeCAD software as previously described,34,45 and fabricated using CNC machining by Epigem Ltd, sealed with adhesive acetate film on both sides (Fig. 1D). The dimensions are provided in the ESI† (Fig. S3A). The device comprised six finger pump chambers, three for sample preparation (containing sample diluent, i.e., nuclease free water) and three for amplification (containing LAMP master mix and primer mix), as well as three chambers containing lateral flow strips (LFS) for visualisation of results (ESI,† Fig. S3B).Eighteen μl of sample diluent (nuclease free water) and 30 μl master mix (containing primers) were loaded separately and the chambers were sealed. The volumes were adjusted to meet the requirements for the lateral flow strip wetting (without the need for an extra step for adding running buffer), which needs ∼40 μl of liquid. This represents the final state of a manufactured device, as would be presented to the user before testing. For the test, 2 μl of sample was added into the diluent chambers, which were then sealed with adhesive acetate film. The finger pumps on the sample diluent chambers were actuated to both mix the sample and move all components into the LAMP reaction mix. The devices were inserted in the heat block to run the LAMP assay at 60 °C (ESI,† Fig. S4). The LAMP reaction products were then transferred to the lateral flow strips using the same finger pumps again, then the assay proceeded under capillary flow for 2 minutes before results were analysed by eye. The diagnostic device included a negative control strip (with just PCR grade nuclease free water as the sample, pre-loaded in the device) and a positive control consisting of SEA and SEB targets (1 ng μl−1), amplified from artificial dsDNA oligonucleotide sequences. This quality control step allows false positives arising from contamination of reagents to be ruled out, and false negatives related to the assay reagents, respectively (ESI,† Fig. S5).
2.7 Field testing
For preliminary evaluation of robustness and user friendliness under field conditions using raw milk samples, we collected 26 samples over two days in one of the production dairy farms of Ataturk University, Turkey with total capacity of 143 cows: 18 samples from individual cows (composite; i.e., the four udder quarters were sanitized with iodine, then the milk was collected from each quarter and combined) and eight bulk tank samples. Samples were tested on the farm using the microfluidic device for the detection of S. aureus enterotoxin A and B genes as per 2.6 above. For volume accuracy, a micropipette was used but could be replaced by droppers46,47 and Pasteur pipettes48 in future.As a comparator assay, performed blind to the field results by a different person, all 26 milk samples were streaked on sheep blood agar containing 7% sheep blood, MacConkey agar, and Mannitol Salt agar (all Oxoid) and incubated at 37 °C for 24 hours. Growing colonies of each morphotype were tested by Gram stain. Isolated Gram-positive cocci were identified as members of the genus Staphylococcus using routine biochemical tests (e.g., catalase, coagulase, DNase, haemolysis on blood agar, urease, mannitol fermentation on mannitol salt agar). They were also analysed for susceptibility to bacitracin and polymyxin B antibiotics and carbohydrate fermentation tests.49 For SEA and SEB gene confirmation, DNA was extracted from biochemically-confirmed staphylococcal isolates using the boiling method. Briefly, two overnight colonies were transferred to a 0.2 ml PCR tube and dissolved in 40 μL single-cell lysis buffer (including Tris–HCl, disodium EDTA, and TE Buffer). The tube was heated to 80 °C for 10 minutes and cooled down to 55 °C for 10 minutes in a thermocycler (Kyratec SC300). The resulting suspension was diluted with PCR-grade nuclease-free water and centrifuged at 4500 g for 30 s to eliminate debris. The supernatant (containing the DNA) was used to run a multiplex PCR assay. The primers used for the detection of SEA and SEB genes were as described39,50 (ESI† Table S4). The reaction mixture used was the same as the multiplex LAMP mixture mentioned before (section 3.4). A reaction mixture containing no added DNA and another with the DNA extract of non-enterotoxigenic S. aureus reference strain (S. aureus ATCC 25923) were used as a PCR negative controls. A reaction mixture containing the DNA extract of enterotoxigenic S. aureus reference strain carrying enterotoxin B gene was used as a positive control. The thermal running conditions were: 95 °C for 5 minutes; 30 cycles of 95 °C for 30 seconds, 58 °C for 30 seconds and 72 °C for 1 minute; 5 min at 72 °C. PCR products of each strain were analysed by gel electrophoresis, with expected band size of 102 and 164 bp for SEA and SEB genes, respectively.
3. Results and discussion
3.1 Evaluation of specificity and sensitivity of a LAMP assay to detect Staphylococcus aureus enterotoxin A and B genes
We first assessed the cross reactivity and specificity of the LAMP primer sets, and noted that there was no amplification with either SEA or SEB primer sets when testing high and low concentrations of the artificial dsDNA oligonucleotides sequences of the other gene, nor with any of the other non-enterotoxigenic reference strains evaluated (Fig. 2). | ||
| Fig. 2 Loop-mediated isothermal assay (LAMP) amplification (solid lines) and melt curves (dotted lines; insets) for Staphylococcus aureus enterotoxin A (SEA) and B (SEB) genes. Amplification curves were only observed for positive controls using the artificial oligonucleotide dsDNA sequence of each gene with its LAMP primers set. Specificity of the assay was confirmed, with no amplification observed when testing i) a non-enterotoxigenic Staphylococcus aureus strain; ii) non-aureus staphylococcal strains (Staphylococcus epidermidis and Staphylococcus intermedius), iii) other foodborne pathogens (Salmonella enteritidis and Bacillus cereus), and iv) when SEA or SEB primer sets were used to amplify the artificial oligonucleotide dsDNA sequence of the other gene (all in negative control test). Each sample was run in duplicate. The two orange (light and dark) curves represent the two replicates of positive control for SEA gene, while dark green and light blue curves represent the two replicates of positive control for SEB gene. | ||
The LOD of the LAMP assays for the detection of SEA and SEB genes from bacterial cultures was established through serial dilutions of DNA extracts, reaching 10−3 ng μl−1 in diluted DNA extracts and 104 CFU ml−1 in DNA extracts from diluted cultures for the SEA gene, whilst it was 10−5 ng μl−1 and 102 CFU ml−1 for the SEB gene (ESI† Fig. S6 and S7).
The LOD for the SEB gene when using extracted DNA (102 CFU ml−1) was comparable with those available from literature.39,40 Whilst the LOD for the SEA gene was higher than that previously reported, we speculate that this may be due to modifications made to the associated primer set (which was done to improve specificity), primer concentration and/or the fact that we used a different DNA extraction kit than the one described in the previous publication(s).39,40 Our modified assay also could detect the SEA gene in a culture with bacterial concentration lower than that needed for enterotoxins to be formed (105 CFU ml−1), and thus stipulated as the required LOD by European regulation.51 The LOD was calculated as the lowest concentration consistently detected for all the replicates. In this work, we use the LOD as a performance parameter to study the different implementations of the assay and its optimisation. The LOD for detection of the SEA and SEB genes in milk samples was determined as part and parcel of the optimisation of sample preparation.
3.2 Sample preparation optimisation
Milk sample dilution in water resulted in faster amplifications as dilution was increased. For 1:2 (0.5) diluted samples, amplification was detected in all three technical replicates among one of three biological replicates after 27–28 minutes of amplification, whereas 1:10 diluted samples (0.1) amplified within 17–18 minutes (Fig. 3). By contrast, amplification was not detected when using undiluted milk samples for LAMP. Extracted DNA from spiked milk samples could be detected at 7–8 minutes. Amplification curves of the SEB gene, using spiked diluted cow milk samples with and without DNA extraction, are shown in the ESI† Fig. S8.
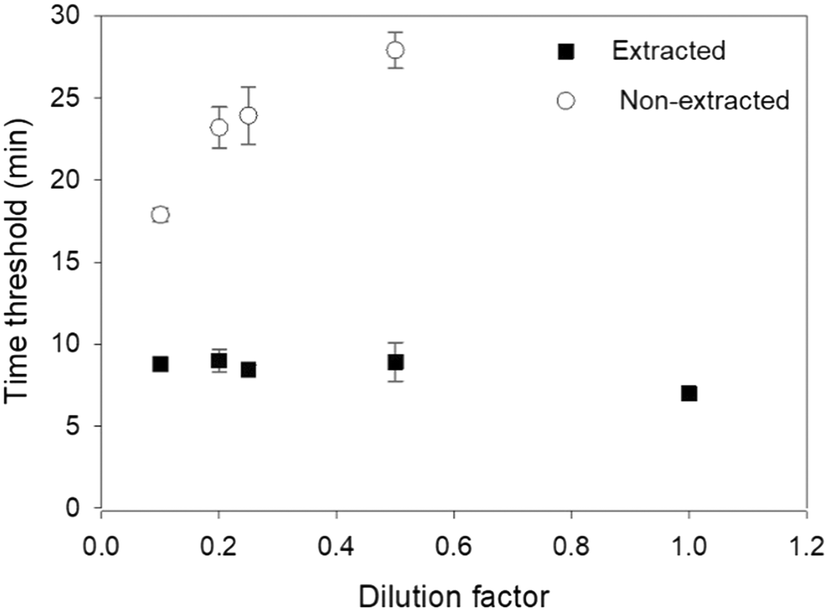 | ||
| Fig. 3 Evaluation of sample dilution as a preparation step for the detection of Staphylococcus aureus enterotoxin B (SEB) gene in spiked cow milk samples using loop-mediated isothermal amplification (LAMP) assay. Black squares: DNA was extracted from cow milk samples spiked with cells collected from overnight culture of Staphylococcus aureus DSMZ 19041 reference strain. The samples were then diluted in water (from no dilution to factor 1:10 (0.1)) and subjected to LAMP after (black squares) or without (open circles) DNA extraction. Each dilution was repeated three times (three biological replicates), each with three technical replicates. Error bar represents standard error of the mean. | ||
A key challenge in using milk samples for molecular assays is the inhibitory nature of milk components,52 such that sample preparation steps have always been required before molecular analysis of milk samples.53 These preparation steps often include chemical treatment, e.g., using DNA extraction kits and lysis buffers54 and are time consuming, costly and require trained personnel, which is not suitable for field application.55 Many approaches have been developed to simplify the sample preparation process,56 including, for example, physical preparation such as centrifugation,57 heating,58 filtration,59 and dilution,60 or often several such steps in combination.
In our approach, we used only a single dilution, one of the simplest modes of physical preparation, which has been shown to facilitate cell lysis through osmotic shock61 when combined with increased LAMP temperature (lysis and heating shock) and could also provide efficient dilution of the inhibitory components in milk samples, as also implemented in other assays.60,62,63 Our results showed that, in general, the dilution of spiked milk samples resulted in significant improvements to the analytical sensitivity of the assay and faster amplification times as dilution was increased. Our results thus indicate that dilution can be used as a simple field-applicable sample preparation step for milk, taking inspiration from other samples (e.g. blood64,65).
Heating decreased amplification time (i.e. time to reach a visible amplification signal) in 1:10 diluted spiked milk samples from both cows and goats (Fig. 4A–C; ESI† Table S5). Heating had no impact on the analytical sensitivity of detection of the SEA gene in cow milk samples spiked with S. aureus DSMZ 19041 (LOD was 104 CFU ml−1 with or without additional heat treatment), while it led to a one log reduction, i.e., an improvement, in the LOD for spiked goat milk samples (104 CFU ml−1 without heating vs. 103 CFU ml−1 using heating at 95 °C) (Fig. 4A). There was no amplification in any of the replicates or the experiments with cow or goat milk samples spiked with 102 and 101 CFU ml−1. LOD of multiplex LAMP assay for SEA and SEB genes in 1:10 diluted cow and goat milk samples spiked with S. aureus DSMZ 19041 was 104 and 105 CFU ml−1 with and without heating, respectively (Fig. 4B). Following demonstration that it was possible to detect DNA in milk spiked with bacterial cells following dilution and heat incubation at 60 °C for amplification, we explored if pre-heating at 95 °C for 10 minutes could enhance bacterial cell lysis and thus decrease LOD. Heating has been previously reported to be a simple, affordable, rapid and effective method for DNA extraction from bacteria.66,33
 | ||
| Fig. 4 Limit of detection (LOD) for Staphylococcus aureus enterotoxin (SE) genes using loop-mediated isothermal amplification (LAMP). All tests were performed using spiked milk samples diluted 1:10 with molecular grade water. Values for LOD are circled. (A) Amplification of SEA gene; (B) multiplex assay for SEA and SEB genes; and (C) amplification of SEB gene, comparing the effect of no heat treatment (closed symbols) vs. heating (open symbols) at 95 °C for 10 minutes prior to LAMP amplification at 60 °C for 30 minutes. For (A) and (B) both cow milk (discs) and goat milk (squares) samples were spiked with S. aureus DSMZ 19041 ten-fold serially diluted. For (C) goat milk samples were spiked with S. aureus DSMZ 19041, 19044 and 20652 reference strains ten-fold serially diluted (results for cow milk are shown in ESI† Fig. S9). (D) LAMP and multiplex LAMP assays for SEA and SEB genes in goat milk samples spiked with S. aureus DSMZ 19041 reference strain incubated at 60 °C for 60 minutes without 95 °C heat treatment. Assays were performed on three independent samples (three biological replicates) with three technical replicates at each concentration. Error bars are standard error. Data were artificially shifted (jittered) along the x-axis for ease of visualization. | ||
Combining sample dilution and heat treatment (i.e. lysis at 95 °C for 10 minutes), the LOD in spiked 1:10 diluted cow milk samples with S. aureus DSMZ 19041 reference strain with and without heating was 104 and 105 CFU ml−1, respectively (ESI† Fig. S9). The LOD in 1:10 diluted cow milk spiked with S. aureus DSMZ 19044 and 20652 reference strains were 103 and 104 CFU ml−1, respectively. LOD of SEB gene in 1:10 diluted goat milk samples spiked with each reference strain with and without heating was 103 and 104 CFU ml−1, respectively (Fig. 4C).
The LOD of LAMP assays for SEA and SEB genes in 1:10 diluted goat milk samples spiked with the cells collected from ten-fold serially diluted overnight culture of S. aureus DSMZ 19041 reference strain and incubated at 60 °C for 60 minutes without 95 °C heat treatment was 103 CFU ml−1 (Fig. 4D; ESI† Table S6), with different amplification times between 39 and 59 minutes. There was no amplification in milk samples spiked with 102 CFU ml−1 and LAMP assay for SEA gene with 101 CFU ml−1 concentration amplified in only one of the three replicates. The duration of heat treatment has been evaluated in combination with other DNA extraction protocols for bovine milk microorganisms.67 We found that combining two physical preparation steps (dilution and heating) improved the assay sensitivity by an order of magnitude and decreased amplification time; this effect was consistent across cow and goat milk and three strains of S. aureus. Of the factors described above, there were statistically significant differences between amplification times of DNA extracts from spiked, diluted milk samples versus non-extracted spiked diluted milk samples (P < 0.001) for both genes, and for detection of SEA in heated when compared with unheated spiked diluted milk of both cows and goats. P-Values associated with the difference between milk from heated versus unheated milk samples from different species (cow versus goat) are summarized in ESI† Table S7.
3.3 Optimization of developed LAMP and multiplex LAMP assays in a heat block
To translate the assay closer to a point-of-need context, the assay was transferred from a thermocycler to a heat block. Consistent with results observed on the thermocycler, on the heat block, the LOD decreased from 104 to 103 CFU ml−1 for goat milk samples spiked with S. aureus DSMZ 19041 reference strain when increasing the incubation time from 30 to 60 minutes (Table 1; ESI† Table S8). When spiked cow milk samples diluted 1:10 were tested using the optimized assay (i.e. incubation at 60 °C for 30 min on the heat block), the LOD was 105 CFU ml−1 for the SEA gene and multiplex LAMP assay and 104 CFU ml−1 for SEB gene (Fig. 5); this is consistent with the LODs observed on the thermocycler, except for SEA gene it was one log higher when incubated on the heat block. We also demonstrated that increasing the amplification time from 30 to 60 minutes led to the same improvement in performance as the inclusion of a thermal lysis step (Table 1). From an end-user point of view, achieving the same (clinically relevant) LOD without the need for an extra processing step that could complicate the protocol may be preferable. In summary, optimal LOD for S. aureus enterotoxin genes using this device can be achieved using either sample heating (lysis) or by increasing the incubation time, thus providing two alternative assay protocols for end-users that might be more relevant for different scenarios, depending on the specific testing context, and the speed of response needed. The LOD were determined under laboratory conditions and may differ when used in the field. However, the performance is better than the recommended EU guidelines,51 often by more than 10 times, providing room to improve.
:10 diluted cow milk samples at 60 °C on a heat block for 30, 45 and 60 minutes
| Target | LOD of LAMP and multiplex LAMP assays for detection of SEA and SEB in spiked 1:10 diluted goat milk samples (colony forming units per ml) |
|||
|---|---|---|---|---|
| Time (minutes) | ||||
| 30 | 45 | 60 | ||
| SEA gene (S. aureus reference strain DSMZ 19041) | 105 | 104 | 103 | |
| SEB gene (S. aureus reference strain DSMZ 19041) | 105 | 104 | 103 | |
| Multiplex LAMP assay (SEA and SEB genes) | S. aureus reference strain DSMZ 19041 | 105 | 104 | 103 |
| S. aureus reference strain DSMZ 19044 | 105 | 104 | 103 | |
| S. aureus reference strain DSMZ 20652 | 104 | 104 | 103 | |
 | ||
| Fig. 5 Limit of detection (LOD) of loop-mediated isothermal amplification (LAMP) assay for the detection of Staphylococcus aureus enterotoxin A (A) and B (B) genes, as well as for the multiplex LAMP assay for both genes (C) in 1:10 diluted cow milk spiked with the cells collected from ten-fold serially diluted overnight culture of S. aureus DSMZ 19041 reference strain, incubated at 60 °C for 30 minutes. +ve = positive control using artificial oligonucleotide dsDNA and −ve = negative control using PCR grade nuclease free water and unspiked milk, respectively. The photos shown were cropped and edited, while original photos are in the ESI† (Fig. S10–S12), where * in each panel represent the LOD. | ||
The device and the multiplexed assay provide a positive result when either or both SEA and SEB genes are present in a sample. From a public health perspective, the presence of one or both enterotoxin genes does not change the sample's interpretation, as detecting either would lead to the same public health response. This approach is also more affordable, as it only requires the immobilization of a single capture molecule on the lateral flow strip instead of two (or the use of multiple strips as per our previous work).45 The multiplex LAMP assay, therefore, was developed as a cumulative, non-differentiating multiplex assay for a single readout, additionally minimizing the risk of confusion for end users in field settings.
3.4 Field testing
Out of the 26 milk samples, only one individual cow sample was positive on our multiplex LAMP assay. All of the microfluidic device's negative and positive controls showed expected results (ESI† Fig. S13). After culturing all milk samples, two samples (including the one with the positive result on LAMP) showed suspect colonies with morphology consistent with Staphylococci. Biochemical testing revealed that these samples contained Gram positive, coagulase negative staphylococci (ESI† Table S9). Multiplex PCR did not detect the presence of enterotoxin A or B in these isolates (ESI† Fig. S14), suggesting the single positive outcome obtained on the device may have been a false positive. Our preliminary field validation suggests the potential ability of this test to be used in field settings; however, wider validation is required to establish robust performance metrics and usability at the point-of-need.4. Conclusion
This study presents the development of a prototype “sample-to-answer” device for the detection of SEA and SEB genes in ruminant milk, suitable for use in low-resource settings. The developed test was rapid (results obtained within one hour), analytically sensitive and specific for the detection of S. aureus enterotoxin genes when using spiked samples in a laboratory setting, and robust to using commercially homogenized and pasteurised milk from different host species (cow and goat), with different S. aureus strains. We successfully incorporated this assay into a microfluidic device with a simple fabrication technique and an easy-to-read diagnostic result in which we combined the multiplexed test for SEA and SEB genes into a single readout, facilitating easy interpretation. By doing this, we propose that the test could greatly simplify the diagnostic testing for the most commonly occurring enterotoxin genes, especially in low resource areas, including on farms and in the case of food-borne illness emergency situations, but further work with fresh milk samples, in field settings, and with potential end-users is needed to validate the scientific and convenience characteristics of the prototype assay.Data availability
Please note that all data associated with this publication referred to by http://dx.doi.org/10.5525/gla.researchdata.1670 is available through the University of Glasgow's Enlighten online data repository.Author contributions
Conceptualization, M. S., V. B. and J. R. methodology, validation and data analysis, M. S., S. C., and M. C. A. performed the experiments. All authors participated to data analysis. Designs and device fabrication, M. S. and X. Y. contributed to the fabrication. All authors contributed to design. Field testing, M. S., J. R., M. C. A. and S. C. writing – original draft, M. S. writing – review & editing, M. S. made modifications of the manuscript, V. B., T. F., J. C. and J. R. reviewed and edited, supervision, V. B., T. F. and J. R.Conflicts of interest
No conflict of interest.Acknowledgements
The authors are grateful for the discussions with Professor Ruth Zadoks on the challenges of designing molecular assays for food safety applications. This work was funded by scholarship funding from the Egyptian Ministry of Higher Education [MM28/19] and from Scottish Universities Life Sciences Alliance (SULSA) [309273]. The work was also part of an Institution Links project [623818418] under the Newton Fund, supported by the UK Department for Business, Energy and Industrial Strategy and Türkiye Bilimsel Araştırma Kurumu (TÜBİTAK) and delivered by the British Council. The funders were not involved in the design, analysis or interpretation of the study data, writing or decision to submit for publication. Diagrams in Fig. 1 and the Table of Contents figure were created in BioRender (https://app.biorender.com).References
- M. M. Dinges, P. M. Orwin and P. M. Schlievert, Exotoxins of Staphylococcus aureus, Clin. Microbiol. Rev., 2000, 13(1), 16–34 CrossRef.
- Y. Le Loir, F. Baron and M. Gautier, Staphylococcus aureus and food poisoning, Genet. Mol. Res., 2003, 2(1), 63–76 Search PubMed.
- M. L. Evenson, M. W. Hinds, R. S. Bernstein and M. S. Bergdoll, Estimation of human dose of staphylococcal enterotoxin A from a large outbreak of staphylococcal food poisoning involving chocolate milk, Int. J. Food Microbiol., 1988, 7(4), 311–316 CrossRef.
- M. Á. Argudín, M. C. Mendoza and M. R. Rodicio, Food poisoning and Staphylococcus aureus enterotoxins, Toxins, 2010, 2(7), 1751–1773 CrossRef.
- V. Rall, F. Vieira, R. Rall, R. Vieitis, A. Fernandes Jr. and J. Candeias, et al. PCR detection of staphylococcal enterotoxin genes in Staphylococcus aureus strains isolated from raw and pasteurized milk, Vet. Microbiol., 2008, 132(3–4), 408–413 CrossRef PubMed.
- M. S. Bergdoll, Enterotoxins, in Staphylococci and Staphy-lococcal Injections, ed. C. S. F. Easman and C. Adlam, Academic Press, New York, 1983, vol. II, pp. 559–598 Search PubMed.
- S. H. Munson, M. T. Tremaine, M. J. Betley and R. A. Welch, Identification and characterization of staphylococcal enterotoxin types G and I from Staphylococcus aureus, Infect. Immun., 1998, 66(7), 3337–3348 CrossRef PubMed.
- C. Letertre, S. Perelle, F. Dilasser and P. Fach, Identification of a new putative enterotoxin SEU encoded by the egc cluster of Staphylococcus aureus, J. Appl. Microbiol., 2003, 95(1), 38–43 CrossRef PubMed.
- M. R. Pelisser, C. S. Klein, K. R. Ascoli, T. R. Zotti and A. C. M. Arisi, Ocurrence of Staphylococcus aureus and multiplex PCR detection of classic enterotoxin genes in cheese and meat products, Braz. J. Microbiol., 2009, 40(1), 145–148 CrossRef PubMed.
- I. V. Pinchuk, E. J. Beswick and V. E. Reyes, Staphylococcal enterotoxins, Toxins, 2010, 2(8), 2177–2197 CrossRef PubMed.
- N. Balaban and A. Rasooly, Staphylococcal enterotoxins, Int. J. Food Microbiol., 2000, 61(1), 1–10 CrossRef PubMed.
- H. S. Tranter, Foodborne staphylococcal illness, Lancet, 1990, 336(8722), 1044–1046 CrossRef PubMed.
- R. Murray, Recognition and management of Staphylococcus aureus toxin-mediated disease, Intern. Med. J., 2005, 35, S106–S119 CrossRef PubMed.
- V. Peton and Y. Le Loir, Staphylococcus aureus in veterinary medicine, Infect., Genet. Evol., 2014, 21, 602–615 CrossRef PubMed.
- D. J. Wilson, R. N. Gonzalez and H. H. Das, Bovine mastitis pathogens in New York and Pennsylvania: prevalence and effects on somatic cell count and milk production, J. Dairy Sci., 1997, 80(10), 2592–2598 CrossRef.
- D. Bergonier, R. De Crémoux, R. Rupp, G. Lagriffoul and X. Berthelot, Mastitis of dairy small ruminants, Vet. Res., 2003, 34(5), 689–716 CrossRef PubMed.
- A. Fagiolo and O. Lai, Mastitis in buffalo, Ital. J. Anim. Sci., 2007, 6(sup2), 200–206 CrossRef.
- J. Kadariya, T. C. Smith and D. Thapaliya, Staphylococcus aureus and staphylococcal food-borne disease: an ongoing challenge in public health, BioMed Res. Int., 2014, 2014, 827965 Search PubMed.
- E. Ortega, H. Abriouel, R. Lucas and A. Gálvez, Multiple roles of Staphylococcus aureus enterotoxins: pathogenicity, superantigenic activity, and correlation to antibiotic resistance, Toxins, 2010, 2(8), 2117–2131 CrossRef PubMed.
- J.-A. Hennekinne, M.-L. De Buyser and S. Dragacci, Staphylococcus aureus and its food poisoning toxins: characterization and outbreak investigation, FEMS Microbiol. Rev., 2012, 36(4), 815–836 CrossRef.
- M. L. de Garnica, B. Linage, J. A. Carriedo, J. A. Santos and C. Gonzalo, Staphylococcus aureus and Escherichia coli prevalence in ovine bulk tank milk, Small Rumin. Res., 2013, 115(1–3), 108–112 CrossRef.
- T. Asao, Y. Kumeda, T. Kawai, T. Shibata, H. Oda and K. Haruki, et al. An extensive outbreak of staphylococcal food poisoning due to low-fat milk in Japan: estimation of enterotoxin A in the incriminated milk and powdered skim milk, Epidemiol. Infect., 2003, 130(1), 33 CrossRef PubMed.
- W. L. Claeys, S. Cardoen, G. Daube, J. De Block, K. Dewettinck and K. Dierick, et al. Raw or heated cow milk consumption: Review of risks and benefits, Food Control, 2013, 31(1), 251–262 CrossRef.
- L. Necidová, Š. Bursová, D. Haruštiaková, K. Bogdanovičová and I. Lačanin, Effect of heat treatment on activity of staphylococcal enterotoxins of type A, B, and C in milk, J. Dairy Sci., 2019, 102(5), 3924–3932 CrossRef.
- WHO, WHO estimates of the global burden of foodborne diseases: foodborne disease burden epidemiology reference group 2007–2015, World Health Organization, 2015 Search PubMed.
- M. Shalaby, J. Reboud, T. Forde, R. N. Zadoks and V. Busin, Distribution and prevalence of enterotoxigenic Staphylococcus aureus and staphylococcal enterotoxins in raw ruminants' milk: A systematic review, Food Microbiol., 2023, 104405 Search PubMed.
- E. Scallan, T. F. Jones, A. Cronquist, S. Thomas, P. Frenzen and D. Hoefer, et al. Factors associated with seeking medical care and submitting a stool sample in estimating the burden of foodborne illness, Foodborne Pathog. Dis., 2006, 3(4), 432–438 CrossRef PubMed.
- EFSA, The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2011, EFSA J., 2013, 11(4), 3129 Search PubMed.
- EFSA, The European Union one health 2019 zoonoses report, EFSA J., 2021, 19(2), e06406 Search PubMed.
- EFSA, The European Union one health 2020 zoonoses report, EFSA J., 2021, 19(12), e06971 Search PubMed.
- S. J. Olsen, L. C. MacKinnon, J. S. Goulding, N. H. Bean and L. Slutsker, Surveillance for foodborne-disease outbreaks—United States, 1993–1997, MMWR CDC Surveill. Summ., 2000, 49(1), 1–62 Search PubMed.
- D. Dewey-Mattia, K. Manikonda, A. J. Hall, M. E. Wise and S. J. Crowe, Surveillance for foodborne disease outbreaks—United States, 2009–2015, MMWR Surveill. Summ., 2018, 67(10), 1 CrossRef PubMed.
- M. Shehadul Islam, A. Aryasomayajula and P. R. Selvaganapathy, A review on macroscale and microscale cell lysis methods, Micromachines, 2017, 8(3), 83 CrossRef.
- J. Reboud, G. Xu, A. Garrett, M. Adriko, Z. Yang and E. M. Tukahebwa, et al. based microfluidics for DNA diagnostics of malaria in low resource underserved rural communities, Proc. Natl. Acad. Sci. U. S. A., 2019, 116(11), 4834–4842 CrossRef PubMed.
- Y. Geng, S. Liu, J. Wang, H. Nan, L. Liu and X. Sun, et al. Rapid Detection of Staphylococcus aureus in Food Using a Recombinase Polymerase Amplification-Based Assay, Food Anal. Methods, 2018, 11(10), 2847–2856 CrossRef.
- N. Sowmya, M. Thakur and H. Manonmani, Rapid and simple DNA extraction method for the detection of enterotoxigenic Staphylococcus aureus directly from food samples: comparison of PCR and LAMP methods, J. Appl. Microbiol., 2012, 113(1), 106–113 CrossRef.
- A. Van Belkum and H. Niesters, Nucleic acid amplification and related techniques in microbiological diagnostics and epidemiology, Cell. Mol. Biol., 1995, 41(5), 615–623 Search PubMed.
- M. Fakruddin, Loop mediated isothermal amplification (LAMP)–an alternative to polymerase chain reaction (PCR), Bangladesh Research Publications Journal, 2011, 5(4), 425–439 Search PubMed.
- H. Yin, T. Fang and H. Wen, Combined multiplex loop-mediated isothermal amplification with lateral flow assay to detect sea and seb genes of enterotoxic Staphylococcus aureus, Lett. Appl. Microbiol., 2016, 63(1), 16–24 CrossRef PubMed.
- M. Goto, H. Hayashidani, K. Takatori and Y. Hara-Kudo, Rapid detection of enterotoxigenic Staphylococcus aureus harbouring genes for four classical enterotoxins, SEA, SEB, SEC and SED, by loop-mediated isothermal amplification assay, Lett. Appl. Microbiol., 2007, 45(1), 100–107 CrossRef.
- A. De Mello, Focus: plastic fantastic?, Lab Chip, 2002, 2(2), 31N–36N RSC.
- D. L. House, C. H. Chon, C. B. Creech, E. P. Skaar and D. Li, Miniature on-chip detection of unpurified methicillin-resistant Staphylococcus aureus (MRSA) DNA using real-time PCR, J. Biotechnol., 2010, 146(3), 93–99 CrossRef CAS PubMed.
- C.-H. Wang, K.-Y. Lien, J.-J. Wu and G.-B. Lee, A magnetic bead-based assay for the rapid detection of methicillin-resistant Staphylococcus aureus by using a microfluidic system with integrated loop-mediated isothermal amplification, Lab Chip, 2011, 11(8), 1521–1531 RSC.
- N. Tomita, Y. Mori, H. Kanda and T. Notomi, Loop-mediated isothermal amplification (LAMP) of gene sequences and simple visual detection of products, Nat. Protoc., 2008, 3(5), 877–882 CrossRef CAS.
- X. Guo, M. A. Khalid, I. Domingos, A. L. Michala, M. Adriko and C. Rowel, et al. Smartphone-based DNA diagnostics for malaria detection using deep learning for local decision support and blockchain technology for security, Nat. Electron., 2021, 4(8), 615–624 CrossRef CAS.
- P. K. Bharti, N. Silawat, P. P. Singh, M. P. Singh, M. Shukla and G. Chand, et al. The usefulness of a new rapid diagnostic test, the First Response® Malaria Combo (pLDH/HRP2) card test, for malaria diagnosis in the forested belt of central India, Malar. J., 2008, 7, 1–6 CrossRef.
- P. Gillet, J. Maltha, V. Hermans, R. Ravinetto, C. Bruggeman and J. Jacobs, Malaria rapid diagnostic kits: quality of packaging, design and labelling of boxes and components and readability and accuracy of information inserts, Malar. J., 2011, 10, 1–15 CrossRef PubMed.
- K. C. Paap, A. M. van Loon, F. M. Koene, L. W. van Buul, S. Jurriaans and M. Smalbrugge, et al. Clinical evaluation of single-swab sampling for rapid COVID-19 detection in outbreak settings in Dutch nursing homes, Eur. Geriatr. Med., 2021, 1–8 Search PubMed.
- P. J. Quinn, B. K. Markey, F. C. Leonard, P. Hartigan, S. Fanning and E. Fitzpatrick, Veterinary microbiology and microbial disease, John Wiley & Sons, 2011 Search PubMed.
- M. Mehrotra, G. Wang and W. M. Johnson, Multiplex PCR for detection of genes for Staphylococcus aureus enterotoxins, exfoliative toxins, toxic shock syndrome toxin 1, and methicillin resistance, J. Clin. Microbiol., 2000, 38(3), 1032–1035 CrossRef CAS.
- Commission Regulation (EC) No 2073. 2005.
- Y.-J. Moon, S.-Y. Lee and S.-W. Oh, A review of isothermal amplification methods and food-origin inhibitors against detecting food-borne pathogens, Foods, 2022, 11(3), 322 CrossRef CAS PubMed.
- J. Vidic, P. Vizzini, M. Manzano, D. Kavanaugh, N. Ramarao and M. Zivkovic, et al. Point-of-need DNA testing for detection of foodborne pathogenic bacteria, Sensors, 2019, 19(5), 1100 CrossRef CAS.
- S. Merk, H. Meyer, I. Greiser-Wilke, L. Sprague and H. Neubauer, Detection of Burkholderia cepacia DNA from Artificially Infected EDTA-blood and Lung Tissue Comparing Different DNA Isolation Methods, J. Vet. Med., Ser. B, 2006, 53(6), 281–285 CrossRef CAS.
- B. P. Casavant, D. J. Guckenberger, D. J. Beebe and S. M. Berry, Efficient sample preparation from complex biological samples using a sliding lid for immobilized droplet extractions, Anal. Chem., 2014, 86(13), 6355–6362 CrossRef CAS.
- B. Pancza, M. Szathmáry, I. Gyurján, B. Bánkuti, Z. Tudós and S. Szathmary, et al. A rapid and efficient DNA isolation method for qPCR-based detection of pathogenic and spoilage bacteria in milk, Food Control, 2021, 130, 108236 CrossRef CAS.
- A. Ramesh, B. Padmapriya, A. Chrashekar and M. Varadaraj, Application of a convenient DNA extraction method and multiplex PCR for the direct detection of Staphylococcus aureus and Yersinia enterocolitica in milk samples, Mol. Cell. Probes, 2002, 16(4), 307–314 CrossRef CAS PubMed.
- J. Jose and K. Brahmadathan, Evaluation of simplified DNA extraction methods for emm typing of group A streptococci, Indian J. Med. Microbiol., 2006, 24(2), 127–130 CrossRef CAS.
- S. J. Wu and C. Kado, Preparation of milk samples for PCR analysis using a rapid filtration technique, J. Appl. Microbiol., 2004, 96(6), 1342–1346 CrossRef CAS PubMed.
- P. Cremonesi, B. Castiglioni, G. Malferrari, I. Biunno, C. Vimercati and P. Moroni, et al. Improved method for rapid DNA extraction of mastitis pathogens directly from milk, J. Dairy Sci., 2006, 89(1), 163–169 CrossRef CAS.
- K. R. Acharya, N. K. Dhand, R. J. Whittington and K. M. Plain, PCR inhibition of a quantitative PCR for detection of Mycobacterium avium subspecies paratuberculosis DNA in feces: diagnostic implications and potential solutions, Front. Microbiol., 2017, 8, 115 Search PubMed.
- E. M. Burd, Validation of laboratory-developed molecular assays for infectious diseases, Clin. Microbiol. Rev., 2010, 23(3), 550–576 CrossRef CAS PubMed.
- CLSI E. 14-A2; Evaluation of Matrix Effects: Approved Guideline, Clinical and Laboratory Standards Institute, Document EP, 2005 Search PubMed.
- A. J. Neri, J. Roy, J. Jarrett, Y. Pan, C. Dooyema and K. Caldwell, et al. Analysis of a novel field dilution method for testing samples that exceed the analytic range of point-of-care blood lead analyzers, Int. J. Environ. Health Res., 2014, 24(5), 418–428 CrossRef CAS PubMed.
- B. P. Sullivan, A. T. Bender, D. N. Ngyuen, J. Y. Zhang and J. D. Posner, Nucleic acid sample preparation from whole blood in a paper microfluidic device using isotachophoresis, J. Chromatogr., B, 2021, 1163, 122494 CrossRef CAS.
- A. A. Dashti, M. M. Jadaon, A. M. Abdulsamad and H. M. Dashti, Heat treatment of bacteria: a simple method of DNA extraction for molecular techniques, Kuwait Med. J., 2009, 41(2), 117–122 Search PubMed.
- J. A. Schwenker, M. Friedrichsen, S. Waschina, C. Bang, A. Franke and R. Mayer, et al. Bovine milk microbiota: Evaluation of different DNA extraction protocols for challenging samples, MicrobiologyOpen, 2022, 11(2), e1275 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4lc00907j |
| This journal is © The Royal Society of Chemistry 2025 |