
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Developing and improving enzyme-driven technologies to synthesise emerging prebiotics
Noelia Losada-Garcia b,
Milica Simovićc,
Marija Ćorovićc,
Ana Milivojevićc,
Nikola Nikačevićc,
Cesar Mateo*a,
Dejan Bezbradica*c and
Jose M. Palomo*a
b,
Milica Simovićc,
Marija Ćorovićc,
Ana Milivojevićc,
Nikola Nikačevićc,
Cesar Mateo*a,
Dejan Bezbradica*c and
Jose M. Palomo*a
aInstituto de Catálisis y Petroleoquímica (ICP), CSIC, C/Marie Curie 2, 28049 Madrid, Spain. E-mail: josempalomo@icp.csic.es; ce.mateo@icp.csic.es
bBioISI—Biosystems and Integrative Sciences Institute, Faculty of Sciences, University of Lisboa, Campo Grande, C8, 1749-016, Lisboa, Portugal
cFaculty of Technology and Metallurgy, University of Belgrade, Karnegijeva 4, 11000 Belgrade, Serbia. E-mail: dbez@tmf.bg.ac.rs
First published on 14th May 2025
Abstract
Emerging prebiotics, mainly oligosaccharides and phenolic compounds, are gaining increasing attention in the scientific community owing to their health benefits and broad industrial potential. Prebiotics are widely used in foods, cosmetic formulations and dietary supplements. Emerging prebiotics offer additional advantages as they can be derived from low-cost, renewable materials and produced sustainably, in line with the principles of a circular economy. Green technological approaches, integrating expertise from different scientific disciplines, will be essential to develop efficient and environmentally friendly methods for the production of emerging prebiotic-enriched products. This review provides a comprehensive overview of the advances in this field, highlighting the advantages and optimisation of enzyme-based catalysis. Insights into how enzymes enhance the control of oligosaccharide production by enabling the selective synthesis of regioisomers with desired chain lengths and modification of phenolic prebiotics are provided. Furthermore, different technologies to improve biocatalysts to contribute to the novel bioprocess intensification strategies applicable to emerging prebiotic processing are elucidated.
Green foundation1. We discussed greener methods to replace complex and tedious chemical processes in the synthesis of emerging oligosaccharides with prebiotic activities via catalytic conversion using enzymes. This approach allows their synthesis in one-step in aqueous media and at low or moderate temperatures.2. Prebiotics have a critical effect as health-promoting agents, with a global market estimated to be $13 billion by 2030. Their role is based on their beneficial effects on the host, stimulating the growth of “good” bacteria and inhibiting “bad” bacteria. They have been evaluated as nondigestible food ingredients and, more recently, for the treatment of skin problems. 3. Although some promising catalysts have been developed for the synthesis of prebiotics, the use of enzymes as catalysts offers significant advantages owing to their selectivity and specificity, with more sustainable and efficient synthesis approaches. This review will discuss methodologies to improve the final biocatalytic process. |
1. Introduction
Prebiotics are stable chemical molecules, mainly carbohydrates, which have critical effects as health promoting agents.1 They beneficially affect the host by selectively stimulating the growth and/or activity of one or a limited number of beneficial bacteria in their colon, thus improving the host health and, in the ideal case, inhibiting pathogenic bacteria.2 Current targets for prebiotics have now expanded beyond the typical probiotic genera Lactobacillus and Bifidobacterium to a wider range of microbial responders.3 Similar to probiotics, these include candidate health-promoting genera such as Roseburia spp., Eubacterium spp., Akkermansia spp., Christensensella spp., Propionibacterium spp., and Faecalibacterium spp. A significant health-promoting benefit of these genera is their capacity to produce short-chain fatty acids (SCFAs), which have been demonstrated to regulate a variety of functions within the gastrointestinal system, including the functions of gut epithelial cells and mucus barrier, immunity, inflammation, glucose and lipid metabolisms, energy expenditure, and satiety.4 Particularly, they have been extensively evaluated as nondigestible food ingredients,2,3,5,6 but recently, their role in the treatment of illnesses such as psoriasis, atopic dermatitis, skin health, colon cancer, COVID, bone health and calcium absorption has been identified.7–11 Thus, they have been highlighted for their relevance in the area of prebiotics.The global prebiotics market, estimated at $6 billion in 2022, is forecasted to reach a revised size of $13.8 billion by 2030, growing at a CAGR (compound annual growth rate) of 11% over the analysis period of 2022–2030. Considering the ongoing post-pandemic recovery, the growth of the galacto-oligosaccharide (GOS) segment has been adjusted to a revised CAGR of 12.1% over the next 8 years.11
In recent times, a range of compounds classified as prebiotics has expanded beyond the “established prebiotics” (inulin and galacto- and fructo-oligosaccharides (GOS and FOS, respectively)) to “emerging prebiotics”, comprising new carbohydrate-based molecules12–14 such as xylooligosaccharides (XOS),15–20 isomaltooligosaccharides (IMO),21,22 pectin oligosaccharides (POS),23–25 mannooligosaccharides (MOS),26 chitooligosaccharides (CHOS),27 lactosucrose, raffinose, epilactose, and glucomannans.12
Furthermore, as the latest definition of prebiotics includes skin microbiota as the target of their activity,3 non-carbohydrate substances, such as polyphenols and other phenolic compounds, have been identified as candidates to exert prebiotic effects on the host.3,12,28 Since they are derived from plant-based materials or by-products generated during food and beverage processing, they align well with current market trends and the principles of a circular economy. Therefore, research on emerging prebiotics, including their synthesis and/or extraction and modification, in vitro models for confirmation of their prebiotic activity, green processing and incorporation in food products, is a multidisciplinary research topic. Owing to the importance of this topic, the number of publications has increased over the last 20 years. In fact, there has been an exponential increase in the number of publications on this topic over the last 10 years (Fig. 1).
 | ||
| Fig. 1 Number of publications per year on prebiotics. Source: Scopus. | ||
Emerging prebiotics are at an earlier development stage than established prebiotics, facing challenges in their discovery, testing, and application. Ensuring resistance to gastric digestion is the key for prebiotics to reach the colon and be fermented by gut microbiota, which depends on their molecular structure. For instance, long-chained IMO is more resistant to degradation in the upper GI tract, making it more potent than short-chained IMO.29
Selectively stimulating beneficial bacteria, such as Bifidobacterium and Lactobacillus, is also crucial, given that different prebiotics can impact the microbial composition differently. For example, XOS was found to stimulate Bifidobacteria growth more effectively than FOS in rats.30 However, validating the efficacy of prebiotics is challenging due to variations in microbiomes and metabolic responses, although new evaluation methods such as dynamic in vitro digestion and fecal fermentation experiments are helping. Despite these challenges, ongoing research is uncovering novel prebiotics that show resistance to digestion and promote health benefits, both in vitro and in vivo.31
One of the most interesting aspects is the preparation of oligosaccharides, which can be produced either via the top–down approach (hydrolysis of the parent polysaccharides to oligosaccharides) or bottom-up approach (synthesis of oligosaccharides from simple sugars) (Fig. 2). The first approach makes it possible to produce these molecules from different sources by hydrolysis, especially from waste from the agri-food or marine industry, lignocellulosic waste,32 seaweeds,33 and crustacean waste34 (Fig. 2). In these cases, the production of carbohydrates involves treatment with strong acids, whereas the use of enzymes allows their preparation in aqueous media under mild conditions, and therefore a higher degree of purity with respect to other by-products.
 | ||
| Fig. 2 Advantages of the enzymatic strategy in the green synthesis of prebiotic oligosaccharides. (a) Top-down approach synthesis, (b) bottom-up approach synthesis, (c) comparison between enzymatic and chemical approaches, and (d) example of the synthesis of a tailor-made bioactive oligosaccharide using enzymatic versus chemical approach. | ||
The enzymatic hydrolysis of polysaccharides serves a dual purpose in the production of emerging prebiotics. It not only facilitates oligosaccharide generation but also acts as a tool for breaking down the cell wall in plants, enhancing the extraction of phenolic compounds, another class of emerging prebiotics, through a green method commonly known as enzyme-aided or enzyme-assisted extraction.35
The second approach, a synthetic method, starts from single building blocks (Fig. 2), where different chemical glycosylation reactions (using metal catalysts, base, etc.) have been described.36 The versatility of chemical processes allows the selective production of various molecules such as di-, tri- and tetra-saccharides, but makes the production of larger compounds more difficult.35,36
In this sense, the use of enzymes can overcome this drawback,37–42 producing different regioisomers of a given chain length with high selectivity and facilitating the formation of larger oligomers more easily (Fig. 2).
To date, most of these compounds on the market are based on a mixture and their prebiotic efficacy has been evaluated in a general way, obviously considering the question of price-performance ratio.
Therefore, several issues such as final overall yield of products, waste production, number of synthetic steps (based on protection and purification steps) can be improved using enzymes as catalysts (Fig. 2). The extraordinary selectivity and specificity of enzymes (particularly, glycosidic bond enzymes) allow these molecules to be obtained in one or a few synthesis steps in aqueous media at moderate temperature, also with high versatility, enabling the same enzyme to be used in several processes. One example is shown in Fig. 2 regarding the synthesis of a hexamer-oligosaccharide.43 The product was synthesized in 94% overall yield using glycosyltransferases (in four enzymatic steps by using cofactor-recycling43), whereas the chemical approach required multiple glycosylation steps (with a high number of previous synthesized building blocks prepared), giving a final overall yield of 2.6%.45
More recently, synthesis strategies have also been developed where enzymatic and chemical steps are combined to obtain tailor-made oligosaccharides.42,44,45
Thus, a large number of enzymatic glycosylation processes has been described, in which, in addition to taking advantage of their excellent natural properties, strategies have been applied to improve them by increasing their stability under the reaction conditions or their versatility in recognizing other similar substrates, for example through protein engineering, or by improving their properties and recyclability, thus improving the economic sustainability of the process, which is of industrial interest.
This review discusses the major advances in the enzymatic synthesis, modification and extraction of various types of emerging prebiotics, as well as contributions in the area of property improvement processing and equipment through new intensified technologies for the food and cosmetics industries.
2. Enzyme-driven development of emerging prebiotics
2.1. Emerging oligosaccharide prebiotics
Currently, GOS, FOS and lactulose are the carbohydrates with prebiotic activity confirmed by clinical studies. However, with the expanded definition of prebiotics, there is a growing demand for the identification, production and evaluation of new prebiotic carbohydrates that can affect the broader range of beneficial microorganisms and targeted hosts.46 These compounds, such as XOS, POS, MOS, IMO and CHOS (Fig. 3), are known as “emerging” prebiotics, given that evidence of their resistance to gastric digestion and selective growth stimulation of beneficial bacteria is still lacking. These compounds present a great scientific and technological challenge due to their structural diversity, arising from their complex natural substrates. The latest achievements and trend towards sustainable transformation of different biomaterials in this field will be presented next. | ||
| Fig. 3 Typical structure of emerging prebiotics, emphasizing the monomeric unit of each one. | ||
XOS are non-digestible oligomers consisting of 2 to10 linked xylose units by β-1,4 linkages (Fig. 3a). Naturally, XOS are present in small amounts in vegetables, fruits, honey and dairy products. XOS are considered to be competitive emerging prebiotics, ideal for incorporation into various products in the food and feed sectors, especially considering their excellent application properties such as high stability in a wide range of temperatures (up to 100 °C) and pH (2.5–8), good sweetening power and low caloric value.12 Depending on the xylan sources used for the production of XOS, the structures of XOS are different in terms of degree of polymerization (DP), monomeric units, and types of linkages. Generally, XOS are mixtures of oligosaccharides formed by xylose linked through β-(1 → 4)-linkages.47 Several examples of the synthesis of XOS from sugar monomers have been described. The enzyme β-xylosidase is capable of synthesising various alkyl β-xylosides through transxylosylation processes.48 The β-xylosidase from A. niger IFO 6662 has strong transxylosyl activity and has been reported to produce a novel non-reducing disaccharides. The synthesis of XOS from β-(1 → 4)-xylobiose in the presence of D-mannose by trans-xylosylation was reported, resulting in the production of two xylosylmannoses and non-reducing XOS49 (Fig. 4a).
 | ||
| Fig. 4 Synthesis of XOS. (a) Synthesis using β-xylosidase and (b) enzyme hydrolysis of xylan. | ||
However, in most cases, XOS are produced using commercially available xylan, but to enable their economically viable production, significant emphasis has been placed nowadays on the use of lignocellulosic biomass, which is an abundant (up to 34% w/w), and more importantly, cheap source of xylan.50 Lignocellulosic materials can be hydrolyzed to XOS using a combination of thermal and chemical pre-treatments to degrade their complex structure, mainly composed of cellulose, hemicellulose and lignin, followed by enzymatic hydrolysis, as shown in Fig. 4b. To date, numerous lignocellulosic materials (wheat and rice straw, sugarcane bagasse, corncob, beech and birch wood) have been studied for XOS production, with varying efficiencies depending on the amount and type of xylan present. Namely, xylan is a biopolymer with a β-1,4-linked xylose backbone with α-D-glucopyranuronic acids and/or L-arabinofuranose residues, and accordingly can be classified into one of four main groups (homoxylan, glucuronoxylan, arabinoxylan, arabinoglucuronoxylan and glucuronoarabinoxylan) based on the presence and distribution of substituents.26 Therefore, different methods of XOS production and the great diversity of potential xylan substrates result in a wide spectrum of different XOS structures with different substituents on the xylose backbone and degrees of polymerization, which consequently have a great impact on their prebiotic and other functional properties.
For example, Vieira et al.51 demonstrated that the lignocellulosic by-product of palm processing (Bactris gasipaes Kunth) can serve as an excellent substrate for XOS production. After mild alkali pretreatment of peach palm waste (inner sheath and peel), XOS were obtained by enzymatic hydrolysis using commercial xylanase from Aspergillus oryzae with XOS yields from the xylan inner sheath and xylan peel of 50.1% and 48.8%, respectively. The obtained XOS showed exceptional antioxidant capacity, significantly higher than that obtained from commercial xylan. In the last few decades, several processes for XOS production have been evaluated, but this is still an evolving field, where the best methods, catalysts and substrates are not completely clear. Therefore, special evaluation of the structure and content of xylan is of utmost importance for the development of viable XOS synthesis processes, especially in terms of enzyme selection. Endo-1,4-β-xylanases (EC 3.2.1.8) are the main enzymes responsible for the hydrolysis of xylan to XOS. Given their diversity, some of them work well on unsubstituted xylan, while others are more susceptible to substituted xylan. Nevertheless, the use of auxiliary enzymes to remove xylan substituents such as α-glucuronidases (EC 3.2.1.139), α-arabinofuranosidases (EC 3.2.1.55) and acetyl esterases (EC 3.1.1.72) has been investigated. An example of enzyme synergism is the degradation of oat spelt xylan using α-L-arabinofuranosidases from Aspergillus hortai, produced and purified from a medium containing citrus pulp and orange peel, and a previously described purified endoxylanase from the same microorganism. The experiments were carried out at 40 °C using 2% (w/v) oat spelt xylan solution in ammonium acetate buffer (pH 4.5), and the enzymes were used individually or in sequential reactions. The results showed that the prior action of the α-L-arabinofuranosidases, which removed the side arabinose substituents and made the main chain more accessible to xylanase action, resulted in a two-fold increase in the hydrolysis yields achieved.52 However, it should be noted that some xylanases show a preference for substituted xylans, where auxiliary enzymes should be included in the process to generate unsubstituted XOS and allow their better utilization by target microorganisms.53 For example, Zhou et al. carried out the hydrolysis of hardwood xylan (glucoronoxylan) using Thermotoga maritima xylanase (XynB) and α-glucuronidase (AguA) co-expressed in Escherichia coli via dual-promoter and bicistronic constructs to reduce the enzyme costs. The α-glucuronidase enabled the removal of 4-O-methyl-D-glucuronic acid residues from the branched XOS, resulting in an increase in the antioxidant capacity of the XOS mixtures produced with both XynB and AguA. In addition, these unsubstituted XOS are believed to significantly accelerate the growth of some bacteria from Bifidobacterium sp.53 Thus, xylanases are excellent tools to tailor prebiotic oligosaccharides with the intention of stimulating different types of bacteria from different niches, in congruence with the newly established definition of prebiotics.54 Recently, to achieve better XOS production, tremendous efforts have been invested in the development of new enzymes with improved activity and stability by increasing the XOS/xylose ratio.
Also, selected microorganisms have been cultivated to produce extracellular xylanases that would thereafter hydrolyze xylan to XOS. A new strain, Bacillus subtilis KCX006, was found to constitutively produce endo-xylanase and xylan debranching enzymes without β-xylosidase activity in the presence of lignocellulosic biomass.55 Therefore, this microorganism was used for the simultaneous production of xylanase and XOS from lignocellulosic biomass (wheat bran, rice bran, rice husk and sugarcane bagasse (SB)) through solid-state fermentation. To provide nitrogen sources for the growth of Bacillus, organic and inorganic nitrogen sources together with different oil-cakes (ground nut oil-cake, sunflower oil-cake, castor oil-cake, and cotton seed cake) were used. Among the various substrates, wheat bran and groundnut oil-cake supported the highest xylanase and XOS production, which under the optimized conditions yielded 3102 IU g−1 and 48 mg g−1, respectively.
POS (Fig. 3b) represent the most diverse group of functionally active carbohydrates within the group of emerging prebiotics. Numerous studies have shown the different physiological activities of POS mixtures, such as prebiotic, antibacterial, anticancer and antioxidant properties.56 The main reason for this diversity is the complexity of the structure of their molecule of origin, pectin. This heteropolysaccharide, which is the major constituent of the cell wall in higher plants, consists of four main structural components, homogalacturonan (HG), rhamnogalacturonan-I (RG-I), rhamnogalacturonan-II (RG-II) and xylogalacturonan (XG).57 The most abundant pectin substructure (more than 65%) is homogalacturonan (HG), which represents a sequence of galacturonic acid residues with α-1,4-linkages, occasionally esterified with methyl (C6) or acetyl groups (O2 and O3). The rest of the molecule is comprised of different monosaccharides (rhamnose, arabinose, galactose, and xylose) and various linkages.56 POS can be produced from pectin via several methods such as enzymatic and acid hydrolysis, hydrothermal treatment method or a combination of acid/enzymatic/hydrothermal treatment. The choice of employed methods is primarily dependent of the pectin source; however, the application of enzymes showed the greatest potential (Fig. 5).
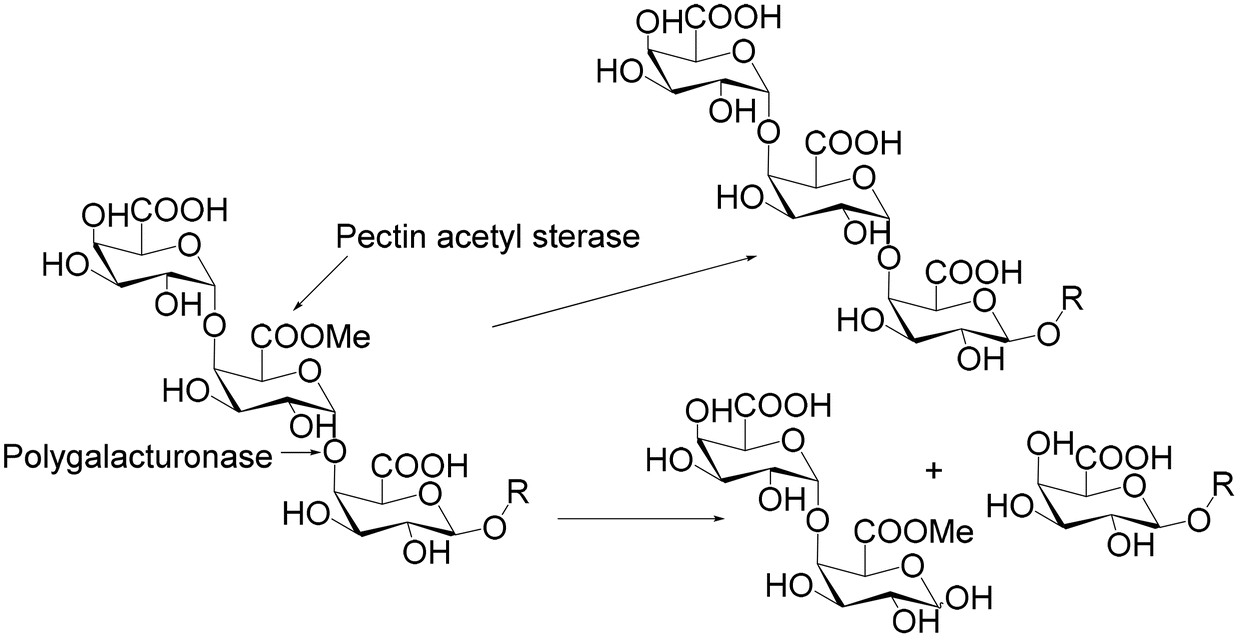 | ||
| Fig. 5 Enzymatic hydrolysis of pectin. | ||
To date, pectin from various sources has been utilized for the production of POS, especially given that various agricultural by-products (apple pomace, sugar beet pulp, citrus waste, berry pomace, etc.) have been recognized as cheap and abundant sources.56,57 The enzymatic hydrolysis procedure is highly complex and requires the synergistic action of different groups of enzymes including hydrolases, esterases and lyases.58 The enzymes responsible for pectin degradation can be majorly divided into HG degrading (polygalacturonase, EC 3.2.1.15, pectate lyase EC 4.2.2.2, and pectin lyase EC 4.2.2.10, pectin methyl esterase EC 3.1.1.11, and pectin acetyl esterase EC 3.1.1.6) and rhamnogalacturonan-I (RG-I) degrading enzymes (α-arabinofuranosidase EC 3.2.1.55, endoarabinase EC 3.2.1.99, β-galactosidase EC 3.2.1.23, endogalactanase EC 3.2.1.89, feruloyl EC 3.1.1.73 and p-coumaroyl esterases EC 3.1.1.B10).
Although there are examples of newly isolated enzymes that are utilized for the synthesis of POS,57 most of the publications presented the results of commercial hydrolytic enzyme mixtures derived from Aspergillus sp.56 For instance, Sabater and co-workers used the commercial enzyme preparation Viscozyme® L, multi-enzyme complex from Aspergillus aculeatus, for the hydrolysis of artichoke pectin into POS. The enzymatic process was optimized using an experimental design and further analyzed by the application of artificial neural networks, yielding 65.9% pectin conversion to POS under the optimal conditions. The structural elucidation of the obtained POS revealed the presence of oligosaccharides DP2–DP6 (from dimer to oligomers with 6 units of monomers), which exhibited strong radical scavenging activities.59 The same enzyme was utilized for the conversion of pectin from industrial byproducts, such as lemon peels, sugar beet pulp60 and onion skin,61 to form a functionally active POS. The crude pectic extract from onion skins, mostly made of homogalacturonan with very scarce rhamnogalacturonan regions, obtained by sodium hexametaphosphate (SHMP) extraction was processed utilizing a continuous crossflow membrane bioreactor. The enzymatic hydrolysis and in situ membrane separation were combined to obtain high yields of tailor-made POS because the membrane allows the continuous removal of products of the targeted DP simultaneously, protecting the POS from further hydrolysis and monosaccharide formation, and hence avoiding enzyme inhibition by reaction products. The reaction resulted in high POS yields (around 60%) provided that enzyme and substrate concentrations were 41.4 U mL−1 and 50 g L−1, respectively. Under these conditions, the highest POS volumetric productivity (22.0 g L−1 h−1), as well as the lowest POS/monosaccharide ratio (4.5 g g−1) were achieved. Also, it must be noted that a stable production was achieved for the whole reaction period and short POS (DP2, DP3, and DP4) were observed as the main reaction products.61
In addition to the dominant processes, which include the utilization of hydrolases, lyases are often applied for the synthesis of POS by cleaving the α-1,4-glycosidic bond of the substrate molecules via trans β-elimination reactions.62 A recent example of cold-active pectate lyases attracted particular attention due to their ability to retain high catalytic efficiency at lower temperatures, simultaneously enabling energy saving, cost reduction and potentially the preservation of physiochemical properties of the treated products. A new cold-tolerant pectate lyase (ErPelPL1) gene from Echinicola rosea was cloned, and thereafter heterologously expressed in Escherichia coli bacteria. The enzyme was purified by high-affinity Ni-charged resin FF (Ni–NTA Sepharose) and the molecular mass of 50 kDa was observed. This enzyme exhibited high catalytic activity at a low temperature (4 °C), although it exhibited optimal activity at 35 °C and pH 8.0 in the presence of 1 mM of Ca2+. It was found that wide range of oligosaccharides (DP2–6) was produced during the reaction course with the predominance of DP2–3.62
MOS (Fig. 3c) are a group of non-digestible oligosaccharides, which are comprised of 2–10 mannose units linked by two types of glycosidic bonds (α-1,6 and β-1,4). Accordingly, MOS can be further divided into two major groups, α- and β-MOS, mostly based on their substrate source. Small amounts of naturally occurring MOS can be extracted from the structural and storage parts of plants; however, the greatest amount of MOS is nowadays generated by the hydrolytic cleavage of different naturally occurring mannans (Fig. 6). Although α-MOS products from yeast cell wall (α-1,6-mannan) hydrolysis have been well established as a feed additive in the agriculture industry,63 β-MOS can be derived from mannans, which represent one of the major constituents (around 50%) of hemicellulose in a variety of plants, primarily softwood, plant seeds and legumes. Also, certain agricultural by-products (for example copra meal and palm kernel meal)64 and waste materials such as spent coffee grounds65 have been proven to be a great mannan source (up to 60% mannans). Plant mannans may appear as homo- (linear mannan) or hetero-polymers (galactomannan, glucomannan and galactoglucomannan) of mannose depending on their source, and therefore β-MOS are primarily generated through the hydrolytic activity of β-mannanase (EC 3.2.1.78), leading to the generation of β-MOS with varying degrees of polymerization (DP). Different β-mannanases showed different preferences in terms of potential substrates and potential products, and thus the achieved yields and DP of the obtained MOS proved to be highly dependent on the β-mannanase utilized. For example, using β-mannanase from Aspergillus oryzae (ManAo) under the same conditions, MOS were obtained from various agricultural byproducts (locust bean gum (LBG), guar gum (GG), konjac gum (KG), palm kernel cake (PKC) and copra meal (CM)) in wide range of yields 9–56%,46 clearly showing specificity towards particular substrates or more specifically types of mannans within these substrates. Magengelele et al. examined the potential of recombinant Aspergillus niger endo-mannanase (Man26A) expressed in Saccharomyces cerevisiae Y294 for the conversion of three substrates, ivory nut linear mannan and two galactomannan substrates with varying amounts of galactosyl substitutions (GG and LBG).66 The enzyme exhibited high substrate specificity towards locust bean gum and ivory nut mannan with the major products DP 2–4, while its specificity towards guar gum was rather low and these reactions generated MOS of higher DP. However, when considering the quantity of obtained MOS, the results showed a discrepancy, given that higher yields were achieved when using galactomannans (4.91 mg mL−1 total reducing sugars for guar gum and 3.89 mg mL−1 total reducing sugars for locust bean gum) as substrates than linear mannan (2.24 mg mL−1 total reducing sugars) during extended periods of mannan hydrolysis. Unlike these enzymes, some β-mannanases show an obvious preference towards unsubstituted regions of linear mannan. Therefore the utilization of different auxiliary enzymes such as β-glucosidase (EC 3.2.1.21), β-mannosidase (EC3.2.1.25), as main-chain mannan-degrading enzymes, and α-galactosidase (EC 3.2.1.22), and acetyl mannan esterase (EC 3.1.1.6), which can enable the removal of the side-chain substituents and break down the complex structure of substituted mannans, should be considered.67 Yang et al. showed that the hydrolysis efficiency of the β-mannanase from Trichoderma reesei is greatly affected by the side chain of galactoses, which introduces steric hindrance for enzymes. They examined the potential of α-galactosidase to enhance the hydrolysis of galactomannan from Sesbania seeds (26% w/w), by adding the enzyme separately (firstly treated with α-galactosidase), sequentially, and simultaneously. Finally, enzymatic hydrolysis by the simultaneous addition of α-galactosidase significantly improved the obtained MOS yields (from 17% to 31%).68 However, it must be noted here that besides the enzyme and substrate selection, careful optimization of the process conditions should be performed to achieve satisfactory product yields, and some of these examples are presented in several review papers.64,67 Besides the achieved yields, the determination of the DP of the obtained products plays an important part in the development of the MOS synthesis process, given that their prebiotic function, as well as their antioxidant, anti-inflammatory, cryoprotectant, anti-stress and anti-diabetic potential is highly dependent on this property. Arunrattanamook et al. used gut the microbiota model microorganism Lactobacillus reuteri to examine the prebiotic potential of synthesized β-MOS.69 Their study showed that medium-length MOS (DP 4 and DP 5) exhibited the highest prebiotic potential, given that higher MOS were poorly utilized, whereas the excessive hydrolysis of mannans resulted in the loss of selectivity toward beneficial bacteria. Therefore, in terms of obtaining the most promising MOS prebiotic mixture and increasing the selectivity toward the production of medium-length MOS, different strategies have been proposed, such as changing the sources of mannan or optimizing the enzymatic hydrolysis conditions and manipulation with β-mannanase.66–68 In the work by Arunrattanamook et al.,69 they improved the specificity of β-mannanase from Aspergillus niger (ManF3) toward the desired product size through rational-based enzyme engineering. Namely, they replaced tyrosine (Tyr 42 and Tyr 132) in the enzyme active site with glycine, which being smaller amino acid, enabled the formation of an extended substrate-binding site, and consequently increased the possibility for higher molecular weight MOS to bind to the enzyme. Finally, this mutation resulted in additional space for the enzyme to accommodate larger hydrolysis products. Mutations of tyrosine into glycine resulted with enhanced yields of medium-chain MOS (DP4 and DP5); however, these mutations had reduced hydrophobic interactions within the enzyme molecule, and thus negatively affected thermal and conformational stability of the mutant enzymes. This is why it was impossible to obtain a double mutant enzyme despite it being successfully constructed.
 | ||
| Fig. 6 Hydrolysis of mannans. | ||
To date, numerous β-mannanases have been tested for the synthesis of MOS, prevalently from genera Aspergillus and Bacillus.67 However, recent studies have brought to our knowledge a previously scarcely explored group of β-mannanolytic enzymes originating from the gut microbiota. For example, Bhattacharya et al.70 explored the potential of a cell-surface exposed β-mannanase (BoMan26B) from the abundant gut bacterium Bacteroides ovatus for the synthesis of MOS from galactomannan, which is abundant in legumes, and acetyl-galactoglucomannan, abundant in softwoods. The reaction yielded partially acetylated linear and galactosyl-containing β-MOS (MOS/GMOS) with an approximate degree of polymerization (DP) between 2 and 6, which was revealed using a newly developed high-resolution anion-exchange chromatography procedure. The abundance of MOS derived from galactomannan followed the order of prevalently DP 5, DP 6 = DP 2 and DP 4, and finally DP 3, while the profile of MOS derived from acetyl-galactoglucomannan was slightly different, i.e., prevalently DP 5, followed by DP 4 and DP 2 = DP 3, and finally DP 6. Under the optimal conditions, the yields of the obtained oligosaccharides were 33% (w/w) and 30% (w/w), respectively. The prebiotic potential of the obtained oligosaccharides was confirmed by means of measuring the production of short-chain fatty acids using the human gut bacteria Bifidobacterium adolescentis ATCC 15703 and Roseburia hominis DSMZ 6839 as acetate and butyrate producers, respectively.70
IMO (Fig. 3d) have been defined as a mixture of oligosaccharides composed of several (usually 2–10) glucose units linked by α-(l → 4) and α-(1 → 6) glycosidic bonds. They are mainly comprised of isomaltose, panose, isomaltotriose, isomaltotetraose, isopanose, and higher branched oligosaccharides. Nowadays, a great deal of IMO is synthesized throughout different chemical and enzymatic processes, but they can be found in honey and soy-based fermented products, such as miso and soy sauce, in small quantities. Besides their controversial prebiotic activity due to the existence of readily digestible α-l,4 linkages, which places them in the emerging prebiotics category, they are characterized by high temperature and pH stability, low viscosity, and low water activity. These properties enabled the approval of various health claims, making them interesting for application in the food and feed sectors.71 IMO are primarily synthesized via the enzymatic conversion of starch from natural and sustainable plant sources (Fig. 7). Cereal crops (wheat, rice, barley, and corn), pulses, and tubers (cassava and potato) are considered the major sources of starch to produce IMO.72 This process involves the multimeric enzymatic action of α-amylase (EC 3.2.1.1) and β-amylase (EC 3.2.1.2), which hydrolyze the long and internal starch branches, together with the starch debranching pullulanase (EC 3.2.1.41) and transglycosylating α-glucosidase (EC 3.2.1.20), as shown at Fig. 7. During the first phase, thermo-tolerant α-amylase liquefies the starch via random cleavage of the α-1,4 linkages, and thereafter α-amylase together with β-amylase, and pullulanase enables starch conversion to maltooligosaccharides. The subsequent reaction is catalyzed by α-glucosidase, an enzyme with primary hydrolytic function that acts on the non-reducing terminal of α-glucosides, but alternatively can transfer a glucosyl residue to another glucose, maltose, isomaltose or isomaltotriose molecule via the formation of α-1,6 linkages. Studies have shown that this step is crucial in the synthesis of IMO, and that the right choice of the α-glucosidase and reaction conditions may greatly influence the functional activity of obtained products through an increment in the panose and isomaltose share in oligosaccharides.72 For example, it has been determined that the simultaneous processes of saccharification and transglycosylation are more likely to produce an increase in the yield and productivity of IMO from starch compared to the conventional process, resulting in lower glucose accumulation through greater transglycosylation. For instance, Duong Hong et al. developed a simple two-step procedure for the production of IMO using sweet potato starch as a low-cost substrate.73 Firstly, they examined the effect of several commercial α-amylase preparations (Spezyme Xtra, Liquozyme SC DS, Spezyme Alpha, and Termamyl SC DS) on the process of starch liquefaction (25% w/v) after 30 min with aim to find the most suitable α-amylase based on the obtained mixture of oligosaccharides. The targeted oligosaccharide mixture in the first step was DP 2–6, given that it was presumed that it would shorten the simultaneous saccharification and transglycosylation time required for the maximum concentration of future IMO with DP between 2 and 4. Accordingly, the Spezyme Xtra preparation (1.0 CU g−1) was chosen for the liquefaction reaction given that the highest concentration of desired oligosaccharides (49.24% w/w) without any generation of free glucose and low generation of starch residue (1.98% w/w) was achieved.
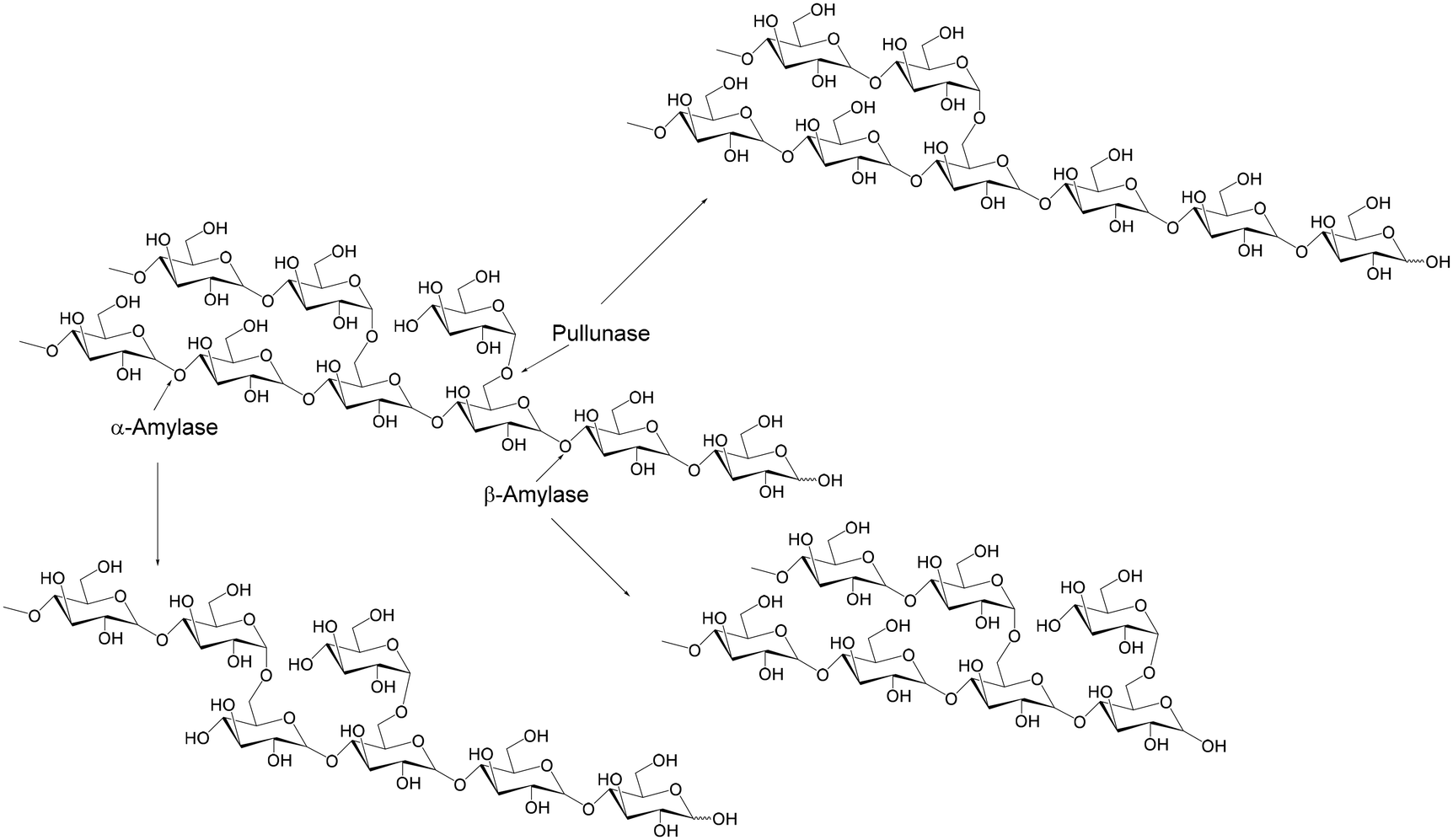 | ||
| Fig. 7 Hydrolysis of starch using different enzymes. | ||
Thereafter, barley β-amylase, pullulanase M2 from Bacillus licheniformis, and transglucosidase from Aspergillus niger were used for simultaneous saccharification and transglycosylation upon the detailed optimization of the enzyme dosage and reaction conditions. Finally, β-amylase (3 U g−1), pullulanase (0.8 U g−1), and transglucosidase (10 U g−1) were applied, leading to the generation of 68.85 g L−1 of IMO at total reaction time three times shorter than in previous studies.73
Another approach toward the development of IMO rich in panose involves the isolation of a new α-glucosidase enzyme preparation possessing advanced kinetic characteristics, primarily increasing the ratio of transglycosylation to hydrolytic activity. Kumar et al. isolated for the first time α-glucosidase from a non-niger Aspergillus isolate with high transglycosylation potential.74 The new soil isolate was identified as Aspergillus neoniger. After purification using a DEAE Sepharose-CL6B column, α-glucosidase was found to possess the molecular mass of 145 kDa and some structural similarities with the commercially available α-glucosidase from Aspergillus niger, although their gene sequences are considerably different. When comparing their activities, it must be noted that the maltose consumption profiles of both enzymes were similar, although the initial rate of consumption was slightly slower in case of the new enzyme. Likewise, the hydrolysis rate of maltose to glucose is lower compared to the commercial enzyme, which is quite important given that the low generation of undesirable by-product can be expected. Additionally, the new enzyme showed higher potential for the synthesis of panose in comparison to the commercial enzyme, as well as a reduction in the secondary hydrolysis of panose rate.74 Therefore, it can be concluded that this enzyme shows great potential for the synthesis of IMO. Successful maltose conversion into desirable IMO (panose, isomaltose and isomaltotriose) can be also achieved using Saccharomyces cerevisiae cells with α-glucosidase activity.73 Namely, the aglA gene that encodes α-glucosidase from Aspergillus niger, known for its transglycosylating activity, was expressed in Saccharomyces cerevisiae in a manner that yeast cells can be used directly as the catalytic agent. The aglA gene was fused to glycosylphosphatidylinositol anchor sequences from the yeast SED1 gene, which enabled the stable attachment of the resulting hybrid enzymes to the cell surface. Thereafter, the potential of the obtained enzyme preparation to perform maltose conversion into IMO was examined. After 8 h incubation, the only reaction product present at a significant concentration was panose. Despite the fact that panose and glucose were produced at equimolar concentrations in the transglycosylation reaction, glucose was most likely consumed by the yeast during this initial period, given that it was not detected in the reaction mixture. As the reaction progressed, the glucose concentration increased, becoming available as the acceptor to produce isomaltose. Therefore, after 24 h, the main transglycosylation product was isomaltose, with lower but significant amounts of panose and isomaltotriose, and low concentrations of higher IMO.74
An alternative method for the production of IMO is based on sucrose as the substrate for the synthesis of dextran through the activity of dextransucrase (EC 2.4.1.5), and the subsequent conversion of dextran into IMO using the activity of dextranase (EC 3.2.1.11). Similar to IMO production from starch, the highest interest from the scientific community is the development of efficient transglycosylation processes.
Chitin is a polymer composed mostly of GlcNAc units linked by β-1 → 4 bonds, together with small amounts of GlcN. It is one of the most abundant polymers in nature, together with cellulose. Natural sources of chitin include exoskeletons of arthropods, mollusks, marine crustaceans, and various microorganisms.75,76 The large quantities of shell and exoskeleton waste generated by these sources contribute to ocean and environmental pollution. As a result, numerous studies have been conducted on exploring the potential applications of chitin and its derivatives. Thus, these polymers have been used as food stabilizers, dietary fiber, and in healthcare.77,78 However, their crystalline structure, resulting from their high proportion of hydrogen bonds, makes them poorly soluble in most solvents. Therefore, the deacetylated product has also garnered interest due to its improved solubility in water and greater chemical modifiability, which often results in higher physiological activity. Chitosan is the deacetylated product of chitin. It is typically produced using chemical methods on a large scale, which consume significant amounts of acids and energy, making them environmentally unsustainable. Alternatively, enzymatic hydrolysis methods offer many advantages over chemical methods, including high efficiency under environmentally benign conditions. Biosynthetic methods are used to prepare various oligosaccharides, including those derived from natural and semi-synthetic sources. These carbohydrates, known as CHOS (Fig. 3e), possess beneficial biological properties, such as antibacterial,79 immunoprotective,80 antitumor,81 and improvements in intestinal health.82 As a result, they have significant applicability, particularly in the food industry.
CHOS can be enzymatically produced through chitin hydrolysis processes or synthesis processes from monomeric or short-chain molecules. Chitin hydrolysis processes are catalyzed by the enzyme chitinase, which has endo- and exo-chitinase activity (EC 3.2.1.14 and 3.2.1.29, respectively) (Fig. 8). Other enzymes, such as lysozyme, lipase, pectinase, glucanase, and papain, have also been described to catalyze the hydrolysis of chitin.83 Yamabhai et al. described the use of chitinase from an extract of Bacillus chitosanase, BsCsn46A, which was capable of producing oligosaccharides of 2–3 sugar units after a 48 h reaction. This study also demonstrated the anti-inflammatory activity of these oligosaccharides.84
 | ||
| Fig. 8 Hydrolysis/synthesis of chitin oligosaccharides catalyzed by chitinase/glycoside hydrolase. | ||
Oligosaccharides can be obtained from monomer units through a hydrolytic process. This reaction is typically catalyzed by the enzyme glycoside hydrolase, either through reverse hydrolysis or transglycosylation.
2.2. Emerging polyphenolic prebiotics
According to the latest definition of prebiotics by the ISAPP in 2017, a compound is considered a prebiotic if it positively impacts the structure and activity of the microbiota, thereby enhancing the host health.85 This broader definition has spurred increased and more intensive research into the synthesis and evaluation of potential prebiotics for two main reasons. Firstly, it expands the range of compounds that meet this criterion, and secondly it acknowledges that the targeted microbiota includes not only the gut but also the skin, thereby involving different microorganisms that respond to different prebiotics.Polyphenols are among the most thoroughly researched emerging prebiotics. These compounds are widely known for their beneficial bioactivities, such as antioxidant, antiatherogenic, and anti-inflammatory.86,87 Nowadays, the effect of different polyphenol structures on the human microbiota is analyzed to select those with prebiotic properties, and thereby a novel field of application. Based on reports of polyphenols as compounds that simultaneously exhibit a stimulatory effect on beneficial human microbiota and antimicrobial activity against pathogens, conferring a health benefit, a novel classification of polyphenols has been suggested, i.e., an emerging class of “duplibiotics”.88 Polyphenols act as prebiotics by influencing the gut microbiota through complex interactions with microorganisms. Some studies have shown that they uniquely support the growth of beneficial commensal microorganisms, while inhibiting the growth of pathogens.87,89 The most representative studies dealing with polyphenol-rich extracts as potential prebiotics are presented in Table 1. Evidence is available on the prebiotic activity of plant extracts rich in certain types of flavonoids; however, a straightforward relationship between specific structures and prebiotic activity has only been established in a limited number of human studies.
| Polyphenols | Microbes stimulated in the gut | Microbes inhibited in the gut | Dosage of the polyphenol | Techniques used for the analysis | Ref. |
|---|---|---|---|---|---|
| Orange juice rich in hesperidin and naringenin | Lactobacillus, Akkermansia, and Ruminococcus | — | 300 ml per day orange juice for 60 days | In vivo study using human faces, qPCR | 90 |
| Mango rich in gallotannins and gallic acid | Lactococcus lactis | Clostridium leptum, Bacteroides thetaiotaomicron | 400 g of mango per day for 6 weeks | In vivo study using human faces, qPCR | 91 |
| Blackcurrant extracts | Bifidobacteria, Lactobacilli | Bacteroides, Clostridia | 2 mL of berry extracts per day for 4 weeks | In vivo study using the cecal of rats, FISH | 92 |
| Blackcurrant extract powder | Bifidobacteria, Lactobacilli | Clostridium spp. and Bacteroides spp. | 672 mg of blackcurrant powder per d | In vivo study using human faces, FISH | 93 |
| Mixture of anthocyanins from blueberry, black currant and black rice extracts. | Bacteroidetes | Firmicutes and Actinobacteria | 215 mg anthocyanins per d for 8 weeks | In vivo study using human faces | 94 |
| Cranberry powder rich in anthocyanins | A. muciniphila, Muribaculaceae, D. newyorkensis, Angelakisella, Coriobacteriaceae, Eggerthellaceae | — | 200 mg of polyphenols per kg of body weight in mice for 8 weeks | In vivo study using the faces of HFHS-fed mice | 95 |
| Concord grape polyphenols rich in anthocyanins | Akkermansia muciniphila | reduction in the ratio of Firmicutes to Bacteroidetes | 1% Concord grape polyphenols for 13 weeks | In vivo study using the faces of C57BL/6J mice. | 96 |
| Bilberry extract rich in anthocyanins | Lactobacillus | reduction of the Firmicutes/Bacteroidetes ratio | 10, 20 or 40 mg extract per kg body weight per day for 70 days | In vivo study using the faces of Sprague-Dawley rats | 97 |
| Green tea extract rich in Epigallocatechin gallate | Lactococcus and Akkermansia | Turicibacter and Romboutsia | 5 mg per kg bodyweight per day for 4 weeks | In vivo study using the faces of C57BL/6J female mice | 98 |
| Dark tea extract rich in gallocatechin | Lactococcus and Akkermansia | Turicibacter, Parasutterella, Lachnoclostridium | |||
| Gallic, ellagic acid, chebulinic acid from triphala | Bifidobacterium spp., Lactobacillus spp. | E. coli | 2% of triphala extract for 3 weeks | In vivo study using Drosophila melanogaster; qPCR | 99 |
| Fermented papaya juice rich in gallic and caffeic acids | — | Firmicutes, Clostridium scindens/Eggerthella lenta | 9 g fermented papaya juice per day for 30 days | In vivo study using human faces, qPCR | 100 |
| Chlorogenic acid | Bacteroidaceae, Lactobacillaceae | Desulfovibrionaceae, Ruminococcaceae, Lachnospiraceae, Erysipelotrichaceae | 150 mg kg−1 day−1 for 6 weeks | In vivo study using the cecum of mice | 101 |
| Ripened Pu-erh tea extract rich in catechins | Roseburia and Akkermansia | Reduction of the Firmicutes/Bacteroidetes ratio | 0.4% of water extract for 8 weeks | In vivo study using male C57BL/6N mice | 102 |
| Dicaffeoylquinic acids from Ilex kudingcha | Bifidobacterium and Akkermansia | — | 3.3 or 10 mg per mouse for 8 weeks | In vivo study using male C57BL/6 mice | 103 |
Polyphenolic compounds can be classified into various groups based on their chemical structures, including flavonoids, phenolic acids, tannins, coumarins, quinones, stilbenes, and lignans and different types of polyphenol structures, as presented in Fig. 9.86
 | ||
| Fig. 9 Classification of polyphenolic compounds. | ||
They are commonly extracted from these raw materials by conventional extraction methods, which involve the utilization of organic solvents. However, novel environment-friendly extraction techniques, which rely on the use of microwaves, ultrasound and enzymes, together with greener media such as deep eutectic solvents, water and supercritical fluids, are nowadays being extensively examined.104,105
Enzyme-assisted extraction procedures have emerged as very promising, given that in addition to the ability of cell wall degrading enzymes to improve the polyphenol extraction efficiency and extract composition, they can also serve as a tool for modifying naturally occurring phenolic compounds, thereby altering their bioavailability and bioactivity, including prebiotic potential (Fig. 10).12,106 The following literature examples will give insight into how different enzyme preparations were previously used for obtaining high yields of specific phenolics with emerging prebiotic potential.
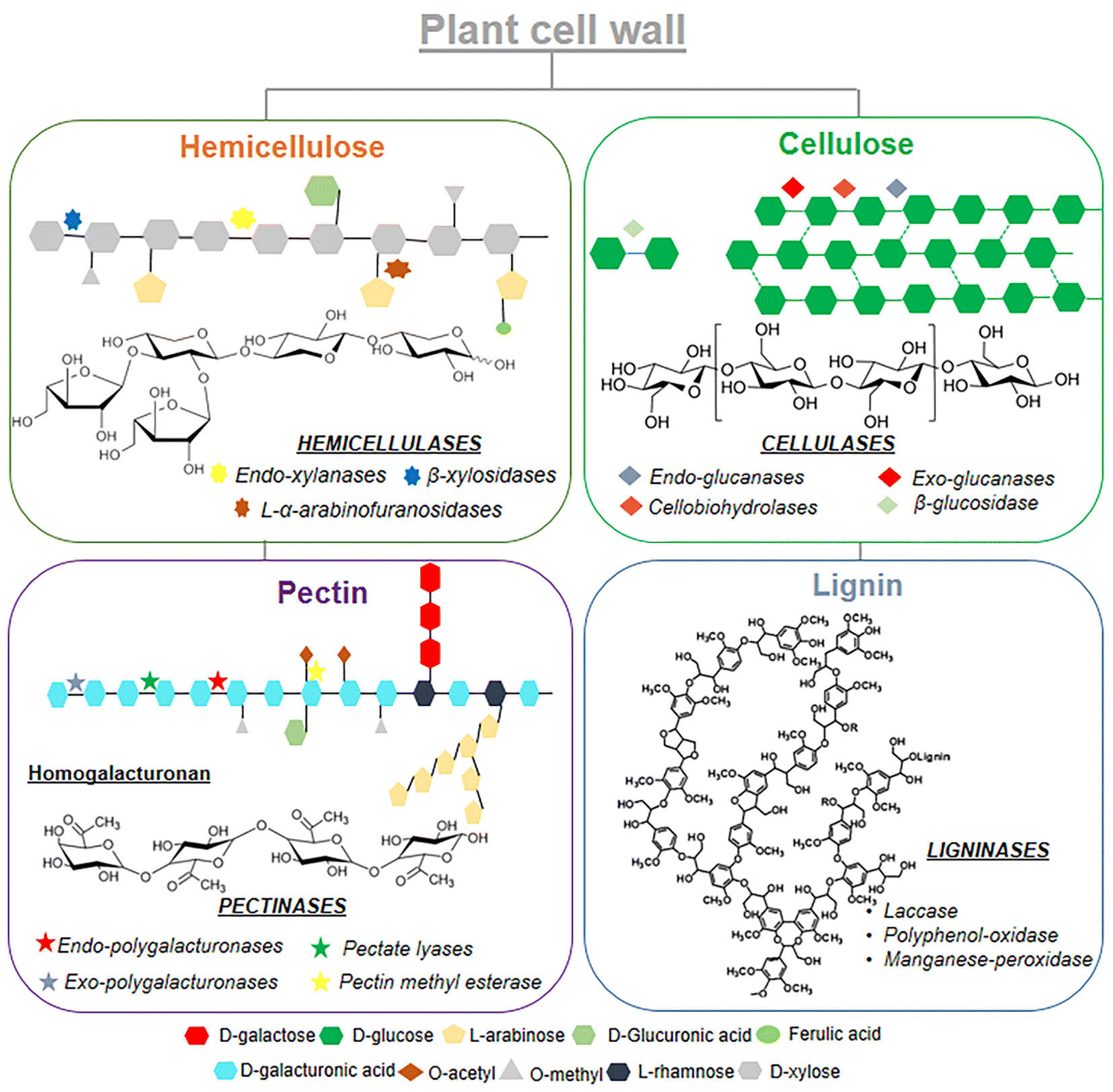 | ||
| Fig. 10 Schematic of enzyme-catalyzed hydrolysis of plant cell wall structures. | ||
One of the polyphenols most thoroughly examined as a potential prebiotic is quercetin. Several animal and in vitro studies found that quercetin had a positive effect on the intestinal microbiota composition.107–110 It is widely distributed in various fruits, vegetables, grains and leaves, usually as glycoside derivatives, and hence β-glucosidase-based modification processes have often been applied for obtaining this compound. In the study performed by Lindahl et al., the enzymatic hydrolysis of quercetin 3,4′-O-diglucoside (Fig. 11a) was combined with a continuous-flow hot water extractor, which resulted in high quercetin yields from yellow, red and shallot onions.111 Onion skin waste, a widely available industrial waste originating from the processing of onions, was also used as a raw material for the production of quercetin. For example, Choi and coworkers reported an increase in quercetin extraction yield of 1.61 times in comparison with the conventional method using a mixture of cellulase (Celluclast® 1.5 L), pectinase (Pectinex® SP-L) and xylanase (X2629 endo-1,4-β-D-xylanase).112 Other plant materials have also been used for the production of quercetin in enzyme-based processes. In a study using guava leaves as the raw material, enzyme-aided extraction produced significantly higher yields of soluble phenolics, including quercetin, compared to conventional extraction.113 Although xylanase-assisted extraction did not influence the composition and yield of extracted polyphenols, the process with cellulase or Trichoderma reesei β-glucosidase enhanced the content of soluble phenolics by around two times, while the enhancement in quercetin was even higher (3.5-fold).
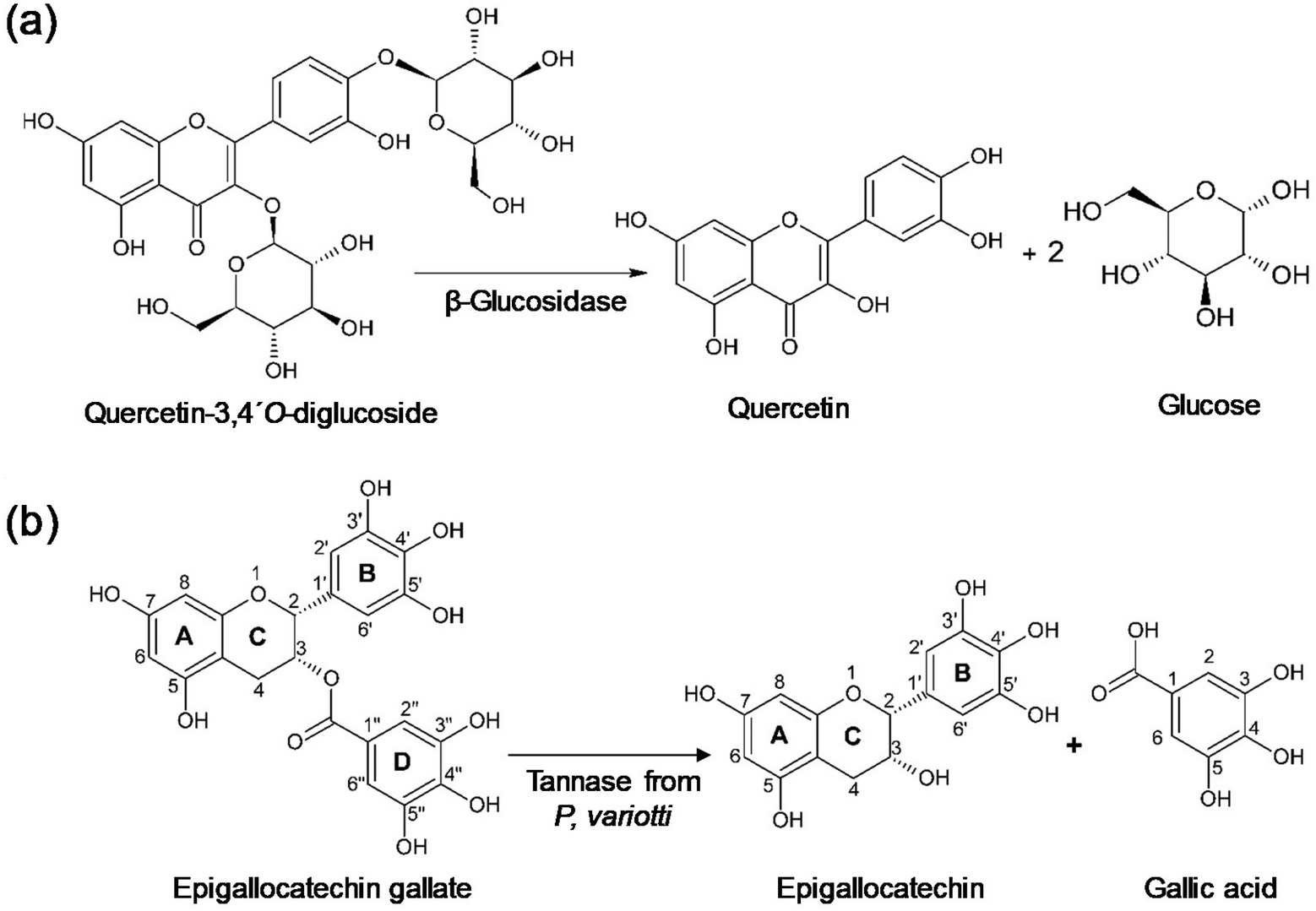 | ||
| Fig. 11 (a) Hydrolysis of quercetin 3,4′-O-diglucoside catalyzed by β-glucosidase (TnBgl1A N221S/P342L). (b) Hydrolysis of epigallocatechin gallate catalyzed by tannase from P. variotii. | ||
Generally, β-glucosidase activity is useful for transforming wide range of naturally occurring phenolic glycosides other than quercetin into their aglycons. One example is the sequential process of enzymatic hydrolysis and supercritical fluid extraction of flavonols and dihydrochalcones from apple pomace.114 The commercial enzyme mix snailase readily removed 96% of sugar moieties from quercetin glycosides, kaempferol glycosides, phloridzin and 3-hydroxyphloridzin owing to its β-glucosidase activity, enabling the production of flavonoid aglycones (quercetin, kaempferol, phloretin and 3-hydroxyphloretin) with 90% extraction yield after sCO2 extraction. The simultaneous process of enzyme-assisted supercritical fluid extraction was also successfully performed.
Phenolic acid-rich extracts (Table 1) and individual phenolic acids (e.g. sodium ferulate and gallic acid)115,116 have also been widely assessed as potential intestinal prebiotics. Enzyme-assisted extraction processes have shown good potential for obtaining increased yields of these compounds from various plant materials. For example, very high yields of three phenolic acids (p-coumaric acid, 8 times, ferulic acid, 4 times, and caffeic acid, 32 times) was achieved in the extract obtained from the extraction of unripe apples with the enzyme preparation Viscozyme® L.117 Another illustrative study focused on grape seed extract and grape pomace, which are known sources of polyphenols. By applying commercial Aspergillus ficum tannase with grape seed extract and grape pomace and the pectinolytic preparation Pektozyme® with grape pomace, the galloylated form of catechin was modified into its free form, releasing gallic acid, which led to an increase in antioxidant activity.118 Meini and coworkers applied an enzymatically assisted process for the extraction of grape pomace polyphenols and proved that the application of pectinase, cellulase and tannase not only significantly improved the extraction efficiency (by 66%) and antioxidant activity (by 80%), but also served as a tool for obtaining extracts of different compositions.119 Hence, using tannase from A. oryzae particularly influenced gallic acid extraction, while p-coumaric acid and malvidin-3-O-glucoside were extracted more efficiently with the aid of cellulase from A. niger.
Flavan-3-ols are structurally very diverse derivatives of flavans with numerous reports on the prebiotic potential of various monomeric, oligomeric and polymeric flavan-3-ols, both in the form of extracts rich in these compounds (Table 1) and as individual molecules.120–122 A. niger cellulase and pectinase were used for the extraction of catechins from black tea (Assam tea variety, S3A3 tea cultivar) leaves, increasing the content of catechin and epigallocatechin gallate content by 51.26% and 15.36%, respectively, compared to the conventional hot water extraction process.123 Another study combined enzymatic treatment using cell wall-degrading enzymes with the sequential tannase hydrolysis of extracted catechins from green tea leaves, which led to an increase in the extraction efficiency and free radical scavenging activity.124,125 Among the tested enzymes, Viscozyme® L stood out as the most effective, given that after sequential treatment with Viscozyme® L and tannase, more than 95% of epigallocatechin gallate and epicatechin gallate was hydrolyzed to epigallocatechin and epicatechin and high amounts of gallic acid were released, increasing the antioxidant activity. Battestin et al. also applied tannase-catalyzed hydrolysis for obtaining epigallocatechin and gallic acid via the degalloylation of epigallocatechin gallate green tea extract using Paecilomyces variotii tannase (Fig. 11b).126 Given that epigallocatechin gallate, unlike degalloylated derivatives and gallic acid, could not revert gut microbiota dysbiosis in previous studies, these examples show the high potential of tannase treatments for increasing not only the antioxidant, but also the prebiotic capacity of plant extracts naturally rich in galloylated catechins.
Anthocyanins have also been thoroughly investigated because not only the growth of beneficial gut microbiota is being stimulated, but also by metabolizing anthocyanins, the microbiota acts as a ‘mediator’ that increases the bioavailability and therapeutic potential of anthocyanins against different chronic diseases, such as obesity, type II diabetes, cardiovascular disease, neurodegenerative disease, inflammatory bowel disease, cancer, fatty liver disease, chronic kidney disease and osteoarthritis.127 Number relevant literature studies have shown the gut microbiota modulatory activity of anthocyanin-rich extracts and the prebiotic-like effect of extracts rich in specific anthocyanins (Table 1).127–129 The enzyme-assisted extraction of anthocyanins from mulberry wine residues and eggplant peel enabled a significant increase in total anthocyanin and polyphenol content in comparison with conventional processes. The dominant anthocyanins in mulberry wine extract were cyanidin-3-O-glucoside and cyanidin-3-O-rutinoside, while eggplant peel extract was rich in delphinidin, petunidin and malvidin.130–132
The evident effect of the type of carbohydrate moiety in polyphenols on their prebiotic properties indicates that the enzymatic modification of this part of the polyphenol molecule can be used as biotechnological tool for fine tuning their activity. Unlike β-glucosidase, which liberates aglycon forms of the corresponding phenolic glycosides, rhamnosidase was previously used for partial deglycosylation, often leading to increased prebiotic activity. A good example is the study by Pan et al., where they compared the effects of different aglycons (hesperetin (flavanon), naringenin (flavanon) and quercetin (flavonol)) and their diglycosides (hesperidin, naringin and rutin) and enzymatically derived (Fig. 12) monoglycosidic forms (hesperetin-7-O-glucoside, prunin and isoquercitrin) on the human gut microbiota, and their metabolism was analyzed by in vitro simulated fermentation.133 The results showed that among the nine tested flavonoids, the enzymatically derived monoglycosides exhibited the most significant beneficial effect on the human gut microbiota composition. The abundance of Bifidobacterium and Lactobacillus was significantly enhanced by three monoglycosides and rutin, while potential pathogens (e.g., Lachnoclostridium and Bilophila) were inhibited by three monoglycosides, naringin and rutin.
 | ||
| Fig. 12 α-L-Rhamnosidase-derived monoglycosides, hesperetin-7-O-glucoside, naringenin-7-O-glucoside, and quercetin-3-O-glucoside, demonstrating the most significant effect on the human gut microbiota composition. | ||
The importance of the transformation of polyphenols to their monoglycosidic form for prebiotic activity was observed in another study, where rutin, hesperidin, naringin and narcissin/rutin mixture were used and 14 tested probiotic strains showed substrate-specific rhamnosidase activity, which reached 27% for hesperidin and 56% for narcissin after four days and 80% for hesperidin and 97% for narcissin after 10 days.133
Besides the gut, the skin microbiome is attracting increasing attention from scientists, consumers and industry. Nowadays, it is known that certain skin commensal strains, such as coagulase negative Staphylococci, are considered probiotics against pathogenic species, primarily S. aureus.134–136 The frequent application of antibiotics, weakened immune system, and genetic and external factors often lead to the development of various skin diseases (e.g. psoriasis and atopic dermatitis), accompanied by skin microbiome disbalance, which is characterized by a decrease in microbial diversity and population of commensal bacteria. Beside oligosaccharides and certain sugar alcohols (e.g. xylitol), polyphenols were assessed for their skin prebiotic potential.137–140 A study with blackcurrant extract revealed that enzymatic processes can provide improvements in the development of novel skin prebiotics, where it was shown that the enzymatically derived extract obtained using a mixture of the commercial preparations Viscozyme® L and Rohapect® MC exhibited higher prebiotic capacity compared to the conventionally obtained extract. This is because the mixture promoted the growth of beneficial S. epidermidis to a higher extent and inhibited harmful S. aureus more strongly.141 Chromatographic analysis revealed differences in the anthocyanin composition caused by partial cyanidin rutinoside and delphinidin rutinoside hydrolysis in enzymatic extracts, implicating that the formed monoglycosides may be responsible for increased prebiotic activity, which is consistent with some of the results obtained with gut microorganisms. It should be noted that the enzymatically derived extracts were rich in cell wall polysaccharide degradation products, given that preparations with cellulolytic and pectinolytic activity were applied, which can also contribute to improved prebiotic properties.
Finally, it should be noted that although promising results were obtained with different polyphenol-rich plant extracts or their constituents obtained by conventional and enzymatic processes, the dose-dependent inhibitory effect of various polyphenols against probiotic strains and stimulation of pathogens was previously reported as well, indicating that further investigations are needed for a better understanding of their gut/skin microbiome modulatory activity and prospective oral/topical application.142–144
3. Technologies for improving enzymes for the synthesis of oligosaccharides
3.1. Protein engineering modification
Protein engineering at the genetic level has emerged as an interesting approach to improve the activity of enzymes.145 This approach has been very useful to improve the production of oligosaccharides via different methods. One approach is to introduce mutations in the sequence of hydrolytic enzymes to improve their activities. For example, in the production of xylooligosaccharides, Wang et al. proposed a method for improving the catalytic performance of XynLC9 xylanase from Bacillus subtilis via the mutation of the N-terminal residues 5-YWQN-8, leading to the development of two mutants named W6F/Q7H and N8Y, which exhibited 2.6- and 1.8-fold higher catalytic activity than wild-type XynLC9, respectively. The double mutant W6F/Q7H with the highest catalytic activity was selected to hydrolyze corncob-extracted xylan in comparison to the wild-type enzyme. During the whole reaction course, W6F/Q7H (substitutions at positions 6 and 7 with Phe and His, respectively) exhibited a superior performance in XOS synthesis. After 14 h, the concentration of XOS reached 10.6 mg mL−1, which was about 1.6-fold higher yield than that of wild-type XynLC9. Additionally, both mutants showed enhanced thermostability.146The second approach is focused on eradicating the hydrolytic capacity of the enzymes by an unique amino acid modification in the protein structure to get a synthase, which is the so-called glycosynthase. This concept is based on the exchange of the catalytic residue (usually an aspartate or glutamate) responsible for promoting nucleophilic attack on the substituted anomeric carbon (Fig. 13) by a non-functional residue (usually alanine or glycine). As a result, they become hydrolytically incompetent.147,148 Then, the enzymatic ability of the enzyme goes through the transfer of an activated glycosyl donor (typically a glycosyl fluoride in anomeric position) to a suitable acceptor to catalyze the formation of a glycosidic bond in high yields.147
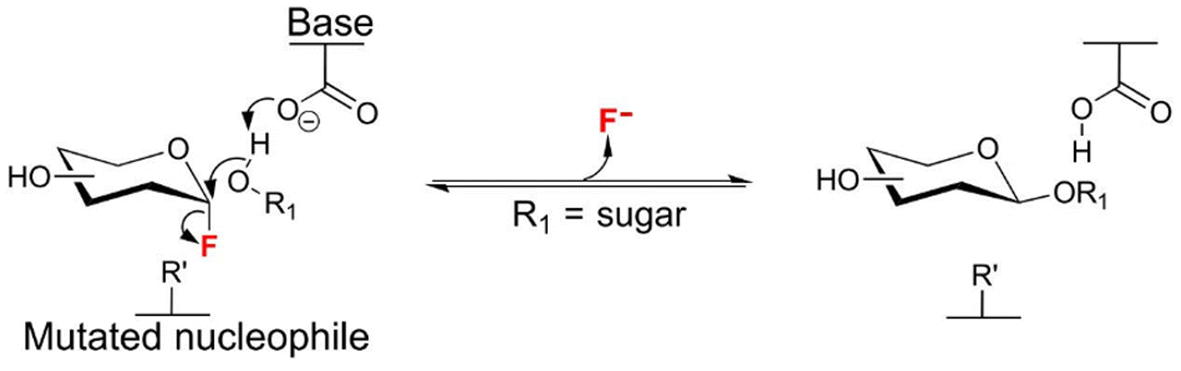 | ||
| Fig. 13 trans-Glycosylation process catalyzed by β-glycosynthases. | ||
This synthetic potential has an enormous impact in the synthesis of oligosaccharides.149 As prebiotic compounds, complex human milk oligosaccharides are gaining increasing attention.150,151 Microorganisms engineered for the synthesis of oligosaccharides from nutrients are considered the most promising systems for process development.152 Therefore, K. Schmölzer et al.153 developed a glucosynthase (variant D746E) from Bifidobacterium bifidum β-N-acetylhexosaminidase JCM1254 through the β-glycosylation of activated N-acetyl-D-glucosaminyl donors by 1,2-oxazoline (Fig. 14a). This represents an important synthetic strategy towards oligosaccharides.154 The authors emphasized enzymatic chemoselectivity given that it is decisive in the highly efficient glycosylation of lactose (∼90%) of NAG-oxa lactose, giving rise to lacto-N-triose II (LNT II), a central building block of human milk oligosaccharides (HMOs). In contrast, the wild-type enzyme hydrolyzed both the NAG-oxa donor and the trisaccharide product with significantly higher activity than glycosynthase. This makes the wild-type enzyme quite unsuitable for synthetic application. The synthesis involved the use of chemically prepared NAG-oxa in 40% yield from N-acetyl-D-glucosamine (GlcNAc). Using equivalent amounts of NAG-oxa and lactose at their solubility limit (600 mM), LNT II was obtained (515 mM; 281 mg mL−1; ∼90% yield; ≤1 h reaction time), which could be immediately recovered from the biocatalytic reaction with 85% purity. These process efficiency metrics reveal the remarkable potential of glycosynthase for chemical process application and highlight it as superior to alternative synthetic options for trisaccharide production.
 | ||
| Fig. 14 (A) trans-Glycosylation by β-glycosynthases and β-N-acetylhexosaminidases. (B) Partial sequence alignment of prokaryotic and eukaryotic GH-20 β-N-acetylhexosaminidases displaying residues critical for substrate-assisted catalysis. Critical amino acids (green) and mutation sites (arrows). (C) Close-up view of the modeled active site of BbhI. The homology model (shown in blue). Key active-site residues of BbhI used for mutagenesis are drawn as sticks (Asp746; D746, polarizing residue; Tyr827; Y827, stabilization of reaction intermediate by hydrogen bonding). The LNB-thiazoline bound in lacto-N-biosidase is shown with green-colored carbon atoms. (D) Comparison of enzymes regarding the maximum yield of LNT II (black bars) and the selectivity parameter RTH (white bars). Copyright 2019, the American Chemical Society. | ||
BbhI is an exo-acting β-N-acetylhexosaminidase and belongs to the GH-20 family of glycoside hydrolases. Enzymes of the GH-20 family use the participation of the neighboring 2-acetamido group of the substrate in catalysis. The enzymatic reaction is promoted by a highly conserved triad of residues (Glu, Asp, and Tyr; Fig. 14b) and proceeds through an intermediate 1,2-oxazolinium ion. A close-up of the active site of a modeled structure of BbhI is shown in Fig. 14c. Based on evidence for endo-β-N-acetylglycosaminidases from the GH-18 and GH-85155,156 families, a promising design for the BbhI glycosynthase was to substitute the residues (Asp746 and Tyr827) involved in the stabilization of the oxazolinium intermediate. The structure model (Fig. 14c) corroborates the evidence for the sequence alignment (Fig. 14b) by suggesting that Asp746 and Tyr827 are positionally conserved at the BbhI active site. Within the GH-20 family, there is limited precedence in the development of glycosynthases. The D313A variant of β-hexosaminidase from Streptomyces plicatus (GH-20) and Tyr470 (Phe, His, and Asn) variants of β-hexosaminidase from Talaromyces flavus157 (GH-20) reflect conceptually similar enzyme design strategies to that applied to BbhI. Four site-directed variants (D746E, D746A, D746Q, and Y827F) of BbhI were prepared (Fig. 14d). The Asp746 variants involve the loss of electrostatic stabilization (D746A) of the intermediate, a likely steric conflict in the substrate/intermediate positioning (D746E), or both (D746Q).
The Y827F variant involves the removal of a hydrogen bond for substrate binding and catalysis.158 However, although these BbhI variants are predicted to have low activity for LNT II hydrolysis, they can utilize the 1,2-oxazoline of N-acetyl-D-glucosamine (NAG-oxa) as a donor for the β-1,3-glycosylation of lactose.
3.2. Chemical modification of protein surface: controlling selectivity in the synthesis of short-oligosaccharides
Selectivity represents a key characteristic in enzymatic synthesis. Structural modifications can influence the capacity to recognize one regioisomer of a particular carbohydrate over another. Thus, modifications by introducing chemical molecules on the protein surface have been successfully developed to enhance this recently.159–163 One example demonstrated the selectivity of levansucrose in controlling the polymer size.159 The authors performed a chemical modification on the tyrosine of levansucrase from B. megaterium, which preferentially happened at residues Y196 and Y247 (Fig. 15). The modification was based on a luminol derivative containing an alkyne group, which represents an orthogonal group. Then, regioselectivity monosaccharides were incorporated on the protein via click chemistry reaction using azide-anomeric activated glucose. This modification allowed the regioselectivity of the enzyme to be altered, and the bioconjugate could produce mainly larger polymers (Fig. 15).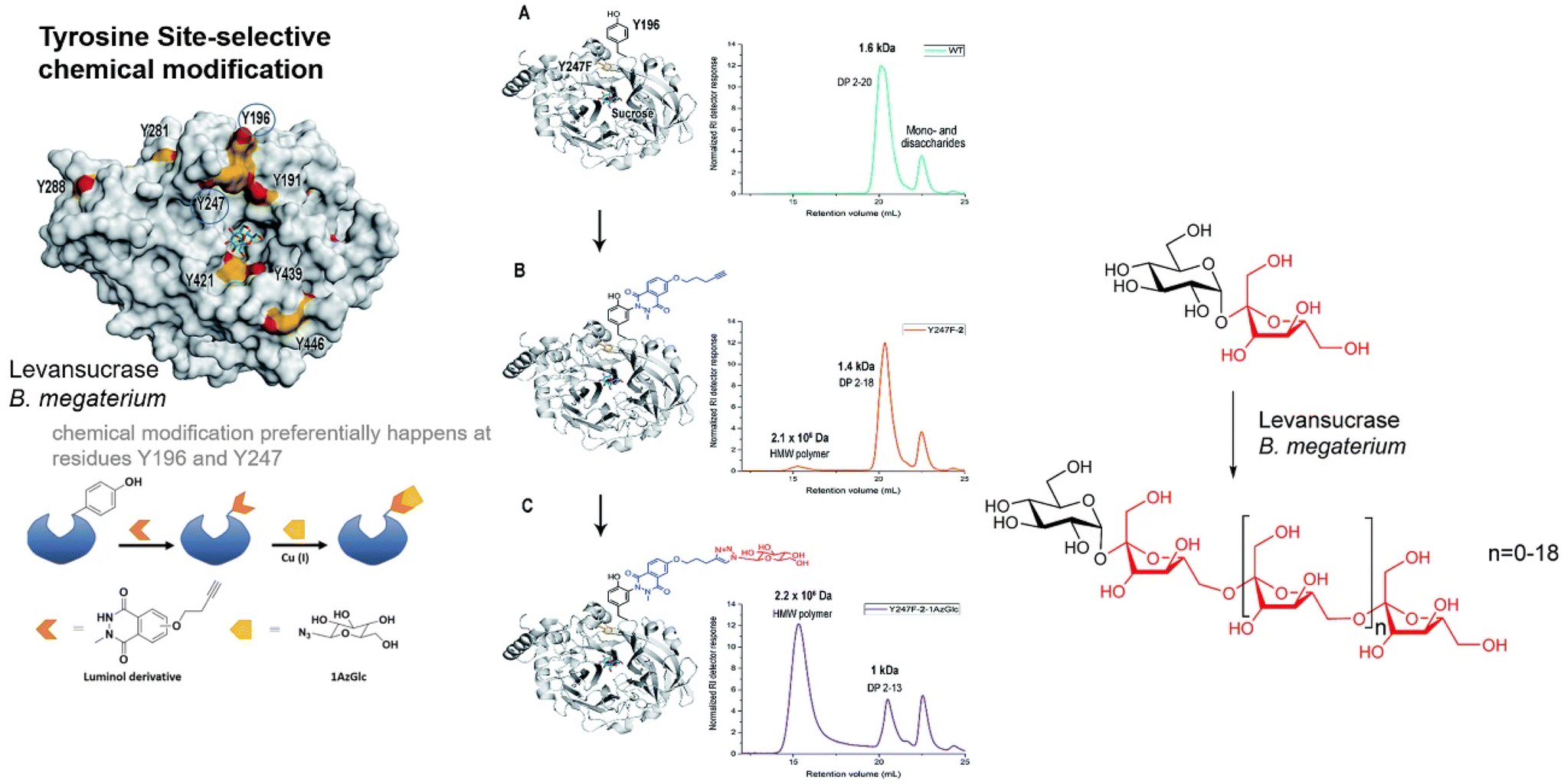 | ||
| Fig. 15 Polymer-elongation specificity via chemical modification of levansucrase from B. megaterium. Figure adapted from ref. 159 with permission from RSC Publishing, Copyright 2018. | ||
Another new alternative in the production of new carbohydrate molecules is the in situ cascade metallo-enzymatic reaction. Employing this approach, a new type of artificial metalloenzyme containing two active sites, one from the enzyme an another artificial site created by the enzyme-induced in situ generation of copper nanoparticles (CuNPs), has been developed.162,163 The green process for the production of new biocatalysts allowed the production of a previous lipase supported on graphene sheets, enabling the selective formation of CuNPs on the enzyme surface exclusively, where the process involved the coordination of the metal with the amino acids of the enzyme in aqueous media. This CuNP-modified lipase conserved the excellent enzymatic regioselectivity against monodeacetylation of peracetylated glucal at C-3 and combined the oxidative capacity of CuNPs for the synthesis of disaccharides via the formation of an epoxy intermediate (Fig. 16). This chemical modification enabled the preparation of very robust bioactive catalysts with excellent recyclability.162,163
 | ||
| Fig. 16 Synthesis of new disaccharides using bioconjugate CuNP-enzyme hybrid. Adapted from ref. 162 with permission from the American Chemical Society, Copyright 2023. | ||
3.3. Immobilization technology to obtain oligosaccharides for industrial application
The use of immobilized enzymes in chemical processes offers several advantages over processes catalyzed by soluble enzymes. One of the most significant advantages is the ability to reuse the catalyst during different reaction cycles. This eliminates the need for purification processes caused by the mixture of the soluble enzyme and different components of the reaction in the case of homogeneous catalysis processes.164 In addition to these advantages, other benefits have been described, such as the ability to use immobilization processes to enhance various enzyme properties, including activity, stability and selectivity.165–168 In the field of oligosaccharide production, enzymes are often improved to enhance their properties and enable the reuse of the catalysts in successive reaction cycles. In this case, immobilization has been shown to improve the properties of enzymes, as demonstrated by the immobilization of xylanase from Aspergillus niger on porous agarose supports using covalent attachment methods. The derivatives resulting from this process were 1100-times more stable than the soluble enzyme. This allowed the production of 60% of oligosaccharides ranging from X2 to X6 xylose units.169Polygalacturonase from Aspergillus aculeatus was immobilized by entrapment in a sol-gel system, resulting in a highly active derivative (94.6% of its initial activity) and improved stability. The immobilized enzyme retained 57% of its catalytic activity after incubation at 55 °C for 2 h, compared to only 17% for the free enzyme. Furthermore, the antibacterial activity of the obtained POS using standard methods was confirmed for POS in the range of DP2−DP4. This immobilization method allowed the reuse of the enzyme, with 65% activity retained after 6 batch reactions.170
β-Mannanase from a Konjac glucomannan preparation was immobilized by entrapment in alginate gels. The enzyme extract, with a purity of 95%, retained 68.3% of its catalytic activity after immobilization. The immobilized catalysts were more stable against extreme pH and high temperatures, with the optimal pH and temperature of 6 and 75 °C, respectively. These catalysts could be reused in the production of MOS for 8 cycles, retaining 70.3% of their catalytic activity. In the oligosaccharide production assay, MOS was generated at a concentration of 8 mg mL−1, consisting of 5 mg mL−1 of mannobiose and 3 mg mL−1 of mannotriose.171
To make the process of MOS synthesis more cost-effective and economically viable by enhancing the stability of the enzyme, Suryawanshi et al. carried out LBG hydrolysis utilizing an immobilized preparation from the newly isolated Aspergillus quadrilineatus RSNK-1.172 Namely, the mixture of hemicellulotytic enzymes comprised of endo-β-mannanase, endo-β-xylanase, β-xylosidase, β-glucosidase and α-galactosidase generated by SSF fermentation on low-cost copra meal was covalently immobilized on aluminum oxide pellets (AOP) after glutaraldehyde 1% (v/v) activation. After optimization of the immobilization procedure using the statistical method of response surface methodology, MOS production was performed in a column bioreactor. The reaction finished in 20 min, and thereafter the obtained immobilized preparation (Man-AOP) was successfully recycled 10 times without significant loss in its activity. The average MOS generation within the 10 successive cycles was 0.95 mg per cycle (1.50, 5.15 and 2.84 mg mL−1 of DP4, DP3 and DP2 MOS, respectively).
Chalane et al.173 described an endodextranase system (D8144) from Penicillium sp. immobilized on an epoxy-activated monolithic Convective Interaction Media (CIM®) disk to produce on-line IMO from Dextran T40. The system retained more than 80% of its residual activity after 5000 column volumes, demonstrating the high stability of the immobilized endodextranases.
Hooda co-immobilized chitinase and glucosaminidase on polyurethane nanoparticles coated with zinc oxide.174 The co-immobilized catalyst demonstrated greater activity under the optimal pH and temperature conditions and higher stability against temperature than soluble enzymes. After incubating the soluble enzymes at 75 °C, 75% loss in activity was observed, while that in the case of the co-immobilized catalyst was only 40%. Furthermore, the catalyst properties were improved, enabling its reuse for at least 50 reaction cycles, with a 50% loss in catalytic efficiency.
Another interesting example of immobilization was described by Ruzic et al.,175 employing the same variant, i.e., Bifidobacterium bifidum β-N-acetyl hexosaminidase variant D746E. Specifically, glucosynthase immobilized on Cu2+-agarose beads (4%) (∼30 mg g−1) packed in a fixed bed (1 mL) was utilized for the stable continuous production of LNT II (145–200 mM) with a quantitative yield of donor substrate (Fig. 17a). Wild-type β-N-acetyl-hexosaminidase used under exactly the same conditions mainly generated the hydrolysis product D-glucosamine (∼85%). By allowing a short residence time (2 min), which is difficult to establish for mixed-pot reactor types, the glycosynthase flow reactor achieved the effective uncoupling of LNT II formation (∼80–100 mM min−1) from slower side reactions (decomposition of donor substrate and enzymatic hydrolysis of LNT II) for the optimal synthetic efficiency (Fig. 17b and c). Thus, this study provides a strong case for the application of flow chemistry principles in glucosynthase reactions, and thereby reveals the important synergy between enzymes and reaction engineering for the biocatalytic synthesis of oligosaccharides.
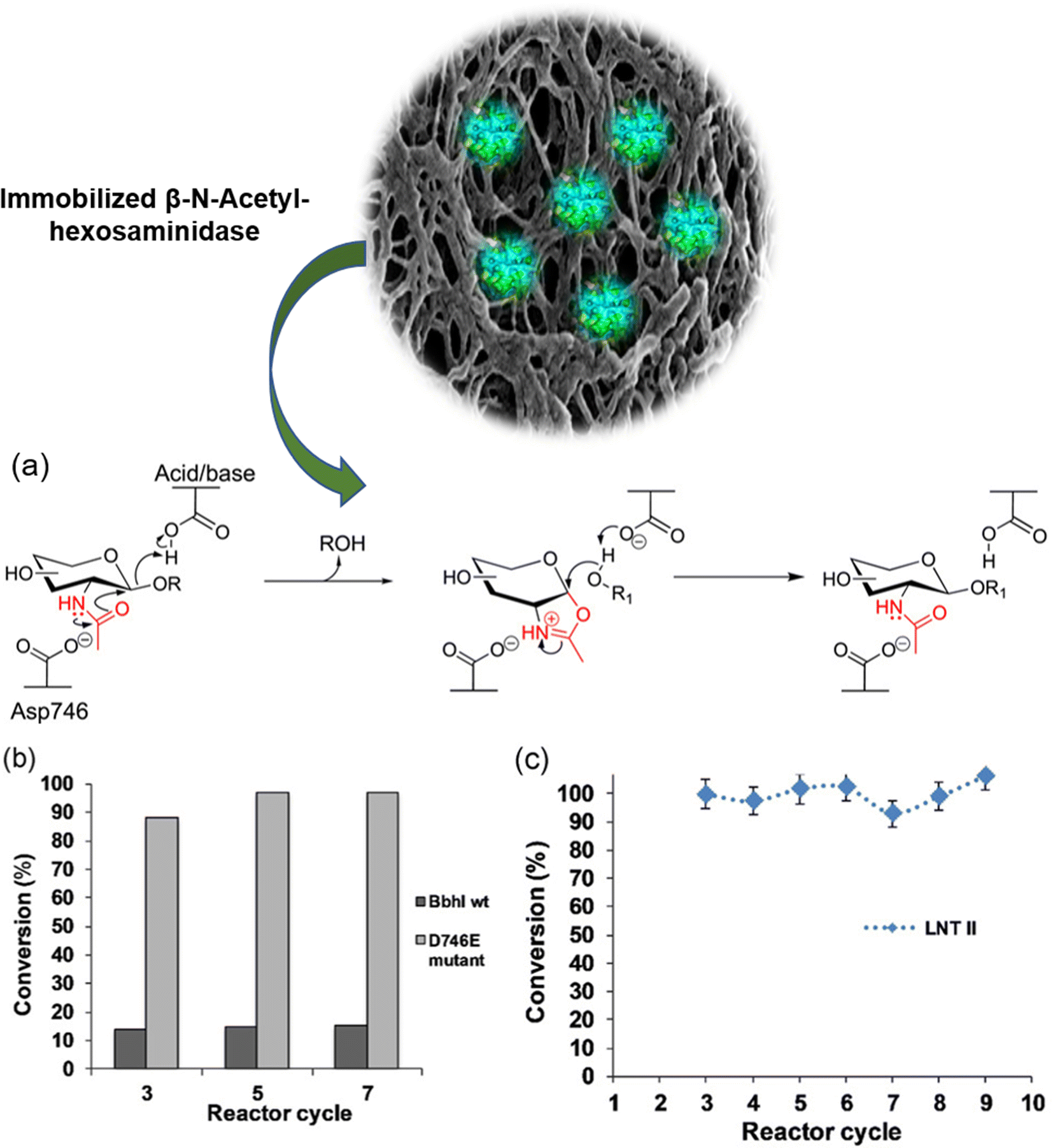 | ||
| Fig. 17 (a) Asp746 of BbhI facilitates the formation of the oxazoline intermediate in the enzymatic reaction. (b) Conversion of GlcNAc–oxa into LNT II. (c) Recyclability process using immobilized D746E glycosynthase. | ||
3.4. Process intensification in oligosaccharides synthesis
Process intensification (PI) concepts and methods can deliver significant benefits in the production of biochemicals and green chemicals. The most promising PI concepts for the synthesis of enzymatic bioproducts are transitioning from batch to continuous processing in novel types of reactors (e.g. micro/milli reactors, oscillatory flow reactors, spiral reactors, and rotating disk reactors), integration of reaction and separation in multifunctional units (e.g. membrane reactors and chromatographic reactors) and utilization of alternative energy sources (e.g. microwaves and sonication). The application of the above-mentioned methods in reactor and process design can result in significant improvements in productivity, higher desired product selectivity, reduction of reactor volume and other fixed costs, lower energy and utilities costs, better process operability and control, and reduced waste generation. Recent reviews176–178 shed light on the versatile and ample possibilities of PI in biochemical applications, proposing the new term bioprocess intensification (BPI/BI).Regarding the synthesis of emerging prebiotics, the research and utilization of PI are still in their infancy. Expectedly, more examples in the literature can be found for established prebiotics, e.g. galacto-oligosaccharides (GOS) and fructo-oligosaccharides (FOS). Fig. 18 presents the process intensification methods and devices used for the synthesis of prebiotics, including the main results achieved (Fig. 18). To transfer from batch to continuous processing, the classical tubular reactors with a fixed bed of immobilized enzymes have been investigated (Fig. 18a). Shin et al.179 used a continuous fixed-bed reactor with β-galactosidase adsorbed on chitosan particles and obtained a good GOS yield of 55% with stable operation for 15 days. Warmerdam et al.180 reported that they achieved the same GOS yield in a fixed-bed bioreactor as in a comparable batch bioreactor, but the volumetric productivity increased by 6 times in the continuous reactor. Lorenzoni et al.181 used both fixed-bed and fluidized-bed reactors for the continuous production of FOS. They reported similar FOS yields (54–59%) in both reactors, although the immobilized enzymes were more stable in the fixed-bed (no changes in activity for 40 days). Zambelli et al.182 showed for FOS synthesis that the reaction time of 72 h in batch processing can be reduced to 10 h in a fixed-bed reactor, resulting in a much higher volumetric productivity. However, the drawbacks of fixed-bed reactors include a relatively high pressure drop and possible mass transfer limitations, both external due to the low velocities used (long residence times needed) and internal due to interparticle diffusion effects (in the case of porous particles with immobilized enzymes) (Fig. 18b). These negative effects can be somewhat reduced in fluidized-bed reactors. Membrane reactors have also demonstrated potential for process improvement, given that the in situ separation of components can lead to higher conversions of substrate and yields of the prebiotic products. Petzelbauer et al.183 used a membrane reactor for GOS synthesis and achieved a GOS concentration 3- to 4-times higher than in batch mode under the same conditions. Córdova et al.184 designed and optimized an ultrafiltration membrane bioreactor. By using high concentrations of lactose, the amount of generated GOS per unit mass of catalyst increased by 2.44-times compared to the batch systems. Pottratz et al.185 investigated the production of GOS by immobilizing the enzyme β-galactosidase on a methacrylic macroporous monolith, which was used as the membrane reactor. In this reactor, the fluid flows unhindered through the pores, which reduces the mass transfer limitations (that occur when conventional supports are used). The results showed that during continuous production, the GOS yield was up to 60% higher compared to the batch production. These reactors are suitable for industrial production due to the simple process scale-up achieved by adding monoliths. Sen et al.186 compared a rotary disk membrane bioreactor (Fig. 18d) with a batch reactor in terms of GOS production. Their research showed that the product yield and purity were higher in the rotating disk membrane bioreactor, with yields 80.2% and 77%, respectively. When comparing product purity, the obtained purity was twice as high when using the rotary disk membrane bioreactor (67.4%) than the batch reactor followed by membrane filtration (32.4%). However, the disadvantages of membrane reactors are the use of high pressures, which can reduce the activity of enzymes by damaging their active centers and possibility of membrane clogging and fouling, all reducing the yield and productivity.
 | ||
| Fig. 18 Process intensification strategies for the synthesis of prebiotics. (a) Fixed bed reactors, (b) Fluidized bed reactors, (c) Membrane reactors, (d) Rotating membrane reactors, (e) Oscilatory buffle reactorz, (f) Helical oscillatory baffled reactors, (g) Table containing the processes identification results in prebiotic molecules production. Figures were adapted from ref. 179, 181, 183, 186–189. | ||
Furthermore, newer types of reactors with special constructions and operation have been proposed for the synthesis of prebiotics. These reactors are oscillatory flow reactors with baffles (OBR) (Fig. 18e), which exhibit very good mixing due to the induced and structured vortices appearing at low net flow rates (needed for longer residence times). Slavnić187 showed that OBRs can be efficiently used for GOS synthesis, both with free-flowing and immobilized enzymes, obtaining higher productivity than in the batch reactor. The shear stresses in OBRs are lower than in the classical mixing units (with propellers), and thus the enzymes maintain their stability.187 Recently, Todić et al.188,189 proposed a modified OBR, replacing the straight tube with a curved one in a helical oscillatory baffled reactor (HOBR, Fig. 18f), which displayed even better mixing due to additional Dean vortices. Pravilović et al.190 confirmed that GOS can be produced in 3D-printed SOBR with a 3-fold increase in volumetric productivity and higher selectivity per enzyme consumed in comparison to the batch reactor. This work189–191 also demonstrated how modeling, optimization and 3D printing can aid the rapid development of intensified enzymatic processes and novel reactors for the synthesis of prebiotics.
It is reasonable to assume that many of the above-presented PI methods can also be applied to the synthesis of emerging prebiotics with comparable success. One should note that the production of emerging prebiotics is more demanding, given that it commonly involves complex-structure substrates and multiphase processing. Thus, some of the above-mentioned concepts would not be suitable or would need considerable modifications. Nevertheless, several research efforts have been reported on the intensification of emerging prebiotics synthesis, e.g. XOS and POS. A flow-type microreactor (150 × 300 μm, 18.7 μL) was used to intensify and increase the efficiency of enzymatic XOS production from beechwood xylan using purified endoxylanase from Thermomyces lanuginosus. The xylan hydrolysis performance was compared with the same reaction performed under batch conditions (100 rpm). The experimental results showed that XOS synthesis was significantly improved compared to the batch reactor, namely a yield of over 80% was achieved in a residence time of less than one minute. The enzymatic xylan hydrolysis performance and xylan to XOS conversion were three times higher under the optimized flow conditions in comparison to the examined batch process.192 However, the disadvantages of micro-reactors are related to their small-size channels, which are not suitable for solid-liquid flows (often required for the synthesis of emerging prebiotics), as well as high fixed costs per unit of flowrate (costs for industrial level capacity).
Another continuous process was designed for the production of POS from sugar beet pulp in a cross flow continuous enzyme membrane reactor, achieving stable production for 28.5 h under the optimized conditions, with an average POS yield of 82.9% (w/w) combined with a volumetric productivity of 17.5 g L−1 h−1 and a specific productivity of 8.0 ± 1.0 g g−1 E h−1. This work demonstrated the stable continuous production of POS from sugar beet pulp using a reactor more suitable for upscaling.193
4. Conclusions and outlook
Emerging prebiotics are compounds with high impact, not only through the general beneficial effect of prebiotics on human health and well-being, which is very important nowadays due to the increasing occurrence of diseases related to food intake and sedentary lifestyles, but also through their contribution to a green transition given that they are compounds that can be produced by green biotechnological processes. These prebiotics, primarily oligosaccharides and polyphenols, are eco-friendly because they are sustainably sourced, given that they can be derived from raw materials such as industrial by-products or waste (e.g., meals, pomaces, bagasse, and biomass) through extraction or hydrolysis from polysaccharide constituents. This review presented the most notable chemical compounds identified as emerging prebiotics, together with evidence of their prebiotic activity. Efficient enzymatic processes are essential for the optimal use of raw materials and fine-tuning the physiological activity of prebiotics. These green catalysts are employed to produce prebiotic oligosaccharides and enhance the extraction and structural modification of polyphenolic prebiotics. Consequently, this review highlighted the significance of enzymatic glycosylation and glycolysis in the production of emerging prebiotics and described the most relevant enzymes used in these processes. However, the high cost of enzymes often poses a significant obstacle in the development of new enzyme-based green technologies, and hence this review provided a comprehensive overview of the current efforts to maximize the biocatalytic potential of enzymes using engineering tools such as protein engineering, immobilization technology, and process intensification. The share of emerging prebiotics in the rapidly growing prebiotic market is expected to increases from 4.7% to 12.9% over the next decade, and hence the joint application of the different types of scientific expertise presented in this review is necessary to commercialize processes that meet market demands, while also adhering to the green transition regulations of industry. Furthermore, life cycle assessment (LCA) studies can be considered for the quantification of the impacts of products and processes on their whole life cycle, adopting a holistic approach.194,195 This analysis can facilitate the identification of the main environmental hotspots of the production process, the comparison of the different process alternatives, and the suggestion of eco-design options for the improvement of these processes. Moreover, these findings can be re-used in other LCA studies including emerging prebiotics in the production system.Author contributions
All authors revised and edited the manuscript. All authors have read and agreed to the published version of the manuscript.Data availability
No primary research results, software or code have been included, and no new data were generated or analysed as part of this review.Conflicts of interest
The authors declare no conflict of interest, financial or otherwise.Acknowledgements
The authors thank the support from the Spanish National Research Council (CSIC) and Ministry of Science, Innovation and University of Spain. Serbian part of research is financed by the taxpayers of the Republic of Serbia (Contract No. 451-03-136/2025-03/200135 of Ministry of Technological Development and Innovations of the Republic of Serbia and programme IDEAS, project PrIntPrEnzy, no. 7750109) and carried out underconditions of legal and financial repression exerted by the Ministry of Education on the teaching staff of University of Belgrade. The authors acknowledge the funding from European Commission, project “Twinning for intensified enzymatic processes for production of prebiotic-containing functional food and bioactive cosmetics “(grant no. 101060130), HORIZON-WIDERA-2021-ACCESS-02-01. N. L-G. thanks the funding by the European Union (BioNanoAct, GA 101153145). Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union or European Research Executive Agency (REA). Neither the European Union nor the REA can be held responsible for them. We acknowledge the support of the publication fee by the CSIC Open Access Publication Support Initiative through its Unit of Information Resources for Research (URICI).References
- R. Todd, M. Kleerebezem, M. V. Kopp and M. Rescigno, Nat. Rev. Immunol., 2012, 12, 728–734 CrossRef PubMed.
- G. R. Gibson and M. B. Roberfroid, J. Nutr., 1995, 125, 1401–1412 CrossRef CAS PubMed.
- G. R. Gibson, R. Hutkins, M. E. Sanders, S. L. Prescott, R. A. Reimer, S. J. Salminen, K. Scott, C. Stanton, K. S. Swanson, P. D. Cani, K. Verbeke and G. Reid, Nat. Rev. Gastroenterol. Hepatol., 2017, 14, 491–502 CrossRef PubMed.
- E. E. Blaak, E. E. Canfora, S. Theis, G. Frost, A. K. Groen, G. Mithieux, A. Nauta, K. Scott, B. Stahl, J. van Harsselaar, R. van Tol, E. E. Vaughan and K. Verbeke, Benefic. Microbes, 2020, 11, 411–455 CAS.
- K. Swennen, C. M. Courtin and J. A. Delcour, Crit. Rev. Food Sci. Nutr., 2006, 46, 459–471 CrossRef CAS PubMed.
- R. Y. Wu, P. Määttänen, S. Napper, E. Scruten, B. Li, Y. Koike, K. C. Johnson-Henry, A. Pierro, L. Rossi, S. R. Botts, M. G. Surette and P. M. Sherman, Microbiome, 2017, 5, 135 CrossRef PubMed.
- V. Lolou and M. I. Panayiotidis, Fermentation, 2019, 5, 41 CrossRef CAS.
- Y. H. Lee, N. K. Verma and T. Thanabalu, J. Funct. Foods, 2021, 78, 104352 CrossRef CAS.
- P. Mishra, V. M. Badiyani, S. Jain, S. Subramanian, S. V. Maharaj, A. Kumar and B. N. Singh, Cancer Rep., 2023, 6, e1870 Search PubMed.
- D. Xavier-Santos, M. Padilha, G. A. Fabiano, G. Vinderola, A. G. Cruz, K. Sivieri and A. E. Costa Antunes, Trends Food Sci. Technol., 2022, 120, 174–192 CrossRef CAS PubMed.
- Prebiotics: Global Strategic Business Report, Region: Global Industry Analysts, Inc., ID: 1206755, 2023 Search PubMed.
- B. B. Cardoso, C. Amorim, S. C. Silvério and L. R. Rodrigues, Adv. Food Nutr. Res., 2021, 95, 41–95 CAS.
- Y. Nakazawa, M. Kageyama, M. Matsuzawa, T. Z. Liang, K. Kobayashi, H. Shimizu, K. Maeda, M. Masuhiro, S. Motouchi, S. Kumano, N. Tanaka, K. Kuramochi, H. Nakai, H. Taguchi and M. Nakajima, Commun. Biol., 2025, 8, 66 CrossRef CAS PubMed.
- I. Ignatova, A. Arsov, P. Petrova and K. Petrov, Molecules, 2025, 30, 803 CrossRef CAS PubMed.
- M. Martins, P. F. Avila, C. C. P. de Andrade and R. Goldbeck, Biocatal. Agric. Biotechnol., 2020, 28, 101747 CrossRef.
- M. Martins, M. F. Silva, T. M. Dinamarco and R. Goldbeck, Process Biochem., 2022, 122, 331–340 CrossRef CAS.
- P. Katapodis and P. Christakopoulo, LWT, 2008, 41, 1239–1243 CrossRef CAS.
- L. Khaleghipour, J. A. Linares-Pasten, H. Rashedi, S. O. R. Siadat, A. Jasilionis, S. Al-Hamimi, R. R. R. Sardari and E. N. Karlsson, Biotechnol. Biofuels, 2021, 14, 153 CrossRef CAS PubMed.
- B. Han, J. Gao, X. Han, H. Deng, T. Wu, C. Li, J. Zhan, W. Huang and Y. You, Food Res. Int., 2022, 162, 112019 CrossRef PubMed.
- G. Precup, J. Venus, M. Heiermann, R. Schneider, I. D. Pop and D. C. Vodnar, Polymers, 2022, 14, 1336 CrossRef CAS PubMed.
- C. Berrocal, H. Chico, E. Carranza and R. Vega, Biochem. Eng. J., 2021, 171, 108003 CrossRef CAS.
- S. Rengarajan and R. Palanivel, Process Biochem., 2020, 98, 93 CrossRef CAS.
- J. Kang, X. Sun, S. Yu, Z. Wang, J. Zhang, Y. Zhao, S. Wang and Q. Guo, Int. J. Biol. Macromol., 2024, 281, 136302 CrossRef PubMed.
- D. Orrego, M. L. Olivares-Tenorio, L. V. Hoyos, C. Alvarez-Vasco, B. K. Ceberio and N. Caicedo, LWT, 2024, 207, 116681 CrossRef CAS.
- Y. Wandee, D. Uttapap, P. Mischnick and V. Rungsardthong, Food Chem., 2021, 348, 129078 CrossRef CAS PubMed.
- J. Chen, J. Yin, H. Xie, W. Lu, H. Wang, J. Zhao and J. Zhu, Food Funct., 2024, 15, 3810–3823 RSC.
- S. A. Ismail, H. S. El-Sayed and B. Fayed, Carbohydr. Polym., 2020, 234, 115941 CrossRef CAS PubMed.
- M. Cunningham, G. Vinderola, D. Charalampopoulos, S. Lebeer, M. E. Sanders and R. Grimaldi, Trends Food Sci. Technol., 2021, 112, 495 CrossRef CAS.
- J. Kaulpiboon, P. Rudeekulthamrong, S. Watanasatitarpa, K. Itoand and P. Pongsawasdi, J. Mol. Catal. B:Enzym., 2015, 120, 127 CrossRef CAS.
- C.-K. Hsu, J.-W. Liao, Y.-C. Chung, C.-P. Hsieh and Y.-C. Chan, J. Nutr., 2004, 134, 1523–1528 CrossRef CAS PubMed.
- J. C. Gomes de Alencar, G. T. da Silva Pinto, K. F. C. Silva, J. M. S. Santos, M. D. Hubinger, J. L. Bicas, M. R. Marostica, C. L. de Oliveira Petkowicz and B. N. Paulino, Trends Food Sci. Technol., 2025, 155, 104808 CrossRef.
- R. Saini, A. K. Patel, J. K. Saini, C.-W. Chen, S. Varjani, R. R. Singhania and C. D. Dong, Bioengineered, 2022, 13, 2139–2172 CrossRef CAS PubMed.
- P. K. Perumal, C.-Y. Huang, C.-W. Chen, G. S. Anisha, R. R. Singhania, C.-D. Dong and A. K. Patel, Bioengineered, 2023, 14, 2252659 CrossRef PubMed.
- H. Amiri, M. Aghbashlo, M. Sharma, J. Gaffey, L. Manning, S. M. M. Basri, J. F. Kennedy, V. K. Gupta and M. Tabatabaei, Nat. Food, 2022, 3, 822–828 CrossRef PubMed.
- C. J. Crawford and P. H. Seeberger, Chem. Soc. Rev., 2023, 52, 7773–7801 RSC.
- O. Gligor, A. Mocan, C. Moldovan, M. Locatelli, G. Crişan and I. C. F. R. Ferreira, Trends Food Sci. Technol., 2019, 88, 302–315 CrossRef CAS.
- M. Huerta, A. San Martín, B. Arancibia, F. A. Cornejo, F. Arenas, A. Illanes, C. Guerrero and C. Vera, Food Bioprod. Process., 2024, 147, 474–482 CrossRef CAS.
- C. Vera, A. Illanes and C. Guerrero, Curr. Opin. Food Sci., 2021, 37, 160–170 CrossRef CAS.
- M. Filice, J. M. Guisan, M. Terreni and J. M. Palomo, Nat. Protoc., 2012, 7, 1783–1796 CrossRef CAS PubMed.
- H.-W. Tseng, H.-K. Tseng, K.-E. Ooi, C.-E. You, H.-K. Wang, W.-H. Kuo, C.-K. Ni, Y. Manabe and C.-C. Lin, JACS Au, 2024, 4, 4496–4506 CrossRef CAS PubMed.
- J. Zhen, H. Xu, J. Fang and X. Zhang, Carbohydr. Polym., 2022, 291, 119564 CrossRef PubMed.
- M. Filice and J. M. Palomo, RSC Adv., 2012, 2, 1729–1742 RSC.
- T.-I. Tsai, H.-Y. Lee, S.-H. Chang, C.-H. Wang, Y.-C. Tu, Y.-C. Lin, D.-R. Hwang, C.-Y. Wu and C.-H. Wong, J. Am. Chem. Soc., 2013, 135, 14831–14839 CrossRef CAS PubMed.
- S. S. Mandal, G. Liao and Z. Guo, RSC Adv., 2015, 5, 23311–23319 RSC.
- R. Zhong, L. Gao, Z. Chen, S. Yuan, X. Chen and C. Zhao, Food Chem.: X, 2021, 12, 100152 CAS.
- L. Santibáñez, C. Henríquez, R. Corro-Tejeda, S. Bernal, B. Armijo and O. Salazar, Carbohydr. Polym., 2021, 251, 117118 CrossRef PubMed.
- A. A. Aachary and S. G. Prapulla, J. Agric. Food Chem., 2008, 56, 3981–3988 CrossRef CAS PubMed.
- H. Shinoyama, Y. Kamiyama and T. Yasui, Agric. Biol. Chem., 1988, 52, 2197–2202 CAS.
- H. Kizawa, H. Shinoyama and T. Yasui, Agric. Biol. Chem., 1991, 55, 671–678 CAS.
- A. Palaniappan, U. Antony and M. N. Emmambux, Trends Food Sci. Technol., 2021, 111, 506–519 CrossRef CAS.
- T. F. Vieira, R. C. G. Corrêa, R. d. F. P. M. Moreira, R. A. Peralta, E. A. de Lima, C. V. Helm, J. A. A. Garcia, A. Bracht and R. M. Peralta, Waste Biomass Valorization, 2021, 12, 6727–6740 CrossRef CAS.
- C. C. Terrone, J. M. F. Nascimento, C. R. F. Terrasan, M. Brienzo and E. C. Carmona, Biocatal. Agric. Biotechnol., 2020, 23, 101460 CrossRef.
- T. Zhou, Y. Xue, F. Ren and Y. Dong, J. Carbohydr. Chem., 2018, 37, 210–224 CrossRef CAS.
- E. N. Karlsson, E. Schmitz, J. A. Linares-Pastén and P. Adlercreutz, Appl. Microbiol. Biotechnol., 2018, 102, 9081–9088 CrossRef PubMed.
- S. S. Reddy and C. Krishnan, Prep. Biochem. Biotechnol., 2016, 46, 49–55 CrossRef CAS PubMed.
- R. P. Singh and J. M. R. Tingirikari, Biocatal. Agric. Biotechnol., 2021, 31, 101910 CrossRef CAS.
- N. Babbar, W. Dejonghe, M. Gatti, S. Sforza and K. Elst, Crit. Rev. Biotechnol., 2016, 36, 594–606 CrossRef CAS PubMed.
- J.-L. Xia and P.-J. Li, Pectic Enzymes, in Encyclopedia of Food Chemistry, ed. L. Melton, F. Shahidi and P. Varelis, Academic Press, Oxford, 2019, pp. 270–276 Search PubMed.
- C. Sabater, A. Blanco-Doval, A. Montilla and N. Corzo, Food Hydrocolloids, 2021, 110, 106161 CrossRef CAS.
- B. Gómez, C. Peláez, M. C. Martínez-Cuesta, J. C. Parajó, J. L. Alonso and T. Requena, LWT, 2019, 109, 17–25 CrossRef.
- S. Baldassarre, N. Babbar, S. Van Roy, W. Dejonghe, M. Maesen, S. Sforza and K. Elst, Food Chem., 2018, 267, 101–110 CrossRef CAS PubMed.
- L. Zheng, Z. Guo, S. Cao and B. Zhu, Bioresour. Bioprocess., 2021, 8, 121 CrossRef PubMed.
- M. Faustino, J. Durão, C. F. Pereira, M. E. Pintado and A. P. Carvalho, Carbohydr. Polym., 2021, 272, 118467 CrossRef CAS PubMed.
- N. Hlalukana, M. Magengelele, S. Malgas and B. I. Pletschke, Foods, 2021, 10, 2010 CrossRef CAS PubMed.
- C. Wongsiridetchai, V. Jonjaroen, T. Sawangwan, T. Charoenrat and S. Chantorn, LWT, 2021, 148, 111717 CrossRef CAS.
- M. Magengelele, N. Hlalukana, S. Malgas, S. H. Rose, W. H. van Zyl and B. I. Pletschke, Enzyme Microb. Technol., 2021, 150, 109893 CrossRef CAS PubMed.
- U. K. Jana and N. Kango, Int. J. Biol. Macromol., 2020, 149, 931–940 CrossRef CAS PubMed.
- L. Yang, G. Shi, Y. Tao, C. Lai, X. Li, M. Zhou and Q. Yong, Appl. Biochem. Biotechnol., 2021, 193, 405–416 CrossRef CAS PubMed.
- N. Arunrattanamook, R. Wansuksri, T. Uengwetwanit and V. Champreda, J. Biosci. Bioeng., 2020, 130, 443–449 CrossRef CAS PubMed.
- A. Bhattacharya, M. Wiemann and H. Stålbrand, LWT, 2021, 151, 112215 CrossRef CAS.
- A. S. Himat, S. Gautam, J. P. C. Garcia, A. X. Vidrio-Sahagún, Z. Liu, D. Bressler and T. Vasanthan, Bioact. Carbohydr. Diet. Fibre, 2021, 26, 100275 CrossRef CAS.
- C. Berrocal, H. Chico, E. Carranza and R. Vega, Biochem. Eng. J., 2021, 171, 108003 CrossRef CAS.
- Q. D. Hong, T. H. Minh, H. G. N. Thi, T. H. N. Thi, N. H. Nguyen, T. L. N. Thi, T. T. Vu and H. N. Luong, J. Food Qual., 2021, 2021, 1987219 Search PubMed.
- S. Kumar, A. Basu, K. A. Anu-Appaiah, B. S. Gnanesh Kumar and S. Mutturi, J. Appl. Microbiol., 2020, 129, 1644–1656 CrossRef CAS PubMed.
- J. L. Shamshina, P. Berton and R. D. Rogers, ACS Sustainable Chem. Eng., 2019, 7, 6444–6457 CrossRef CAS.
- C. P. Souza, B. C. Almeida, R. R. Colwell and I. N. Rivera, Mar. Biotechnol., 2011, 13, 823–830 CrossRef CAS PubMed.
- H. Wang, B. Li, F. Ding and T. Ma, Prog. Org. Coat., 2020, 149, 11 Search PubMed.
- M. Dohendou, K. Pakzad, Z. Nezafat, M. Nasrollahzadeh and M. G. Dekamin, Int. J. Biol. Macromol., 2021, 192, 771–819 CrossRef CAS PubMed.
- Z. Fan, Y. Qin, S. Liu, R. Xing, H. Yu, X. Chen, K. Li, R. Li, X. Wang and P. Li, Carbohydr. Polym., 2019, 224, 115155 CrossRef CAS PubMed.
- A. Maia, Y. C. Gloria, K. Fuchs, T.-H. Chang, P. Engels, M. Zhou, T. Hinnenthal, E. Rusch, C. Gouttefangeas and A. N. R. Weber, J. Leukocyte Biol., 2023, 114, 180–186 CrossRef CAS PubMed.
- X. Liu, Y. Zhang, Z. Liu and X. Xie, OncoTargets Ther., 2019, 12, 7581 CrossRef CAS PubMed.
- H. Zhen, Q. Yan, Y. Liu, Y. Li, S. Yang and Z. Jiang, Food Sci. Hum. Wellness, 2022, 11, 999–1009 CrossRef CAS.
- M. Guo, X. Wei, S. Chen, J. Xiao and D. Huang, Int. J. Biol. Macromol., 2022, 209, 631–641 CrossRef CAS PubMed.
- M. Yamabhai, M. Khamphio, T. T. Min, C. N. Soem, N. C. Cuong, W. R. Aprilia, K. Luesukprasert, K. Teeranitayatarn, A. Maneedaeng, T. R. Tuveng, S. B. Lorentzen, S. Antonsen, P. Jitprasertwong and V. G. H. Eijsink, Carbohydr. Polym., 2024, 324, 121546 CrossRef CAS PubMed.
- G. R. Gibson, R. Hutkins, M. E. Sanders, S. L. Prescott, R. A. Reimer, S. J. Salminen, K. Scott, C. Stanton, K. S. Swanson, P. D. Cani, K. Verbeke and G. Reid, Nat. Rev. Gastroenterol. Hepatol., 2017, 14, 491–502 CrossRef PubMed.
- A. Andreu, M. Ćorović, C. Garcia-Sanz, A. S. Santos, A. Milivojević, C. Ortega-Nieto, C. Mateo, D. Bezbradica and J. M. Palomo, Catalysts, 2023, 13, 1359 CrossRef CAS.
- F. Nazzaro, F. Fratianni, V. De Feo, A. Battistelli, A. G. Da Cruz and R. Coppola, Adv. Food Nutr. Res., 2020, 94, 35–89 CAS.
- M. C. Rodríguez-Daza, E. C. Pulido-Mateos, J. Lupien-Meilleur, D. Guyonnet, Y. Desjardins and D. Roy, Front. Nutr., 2021, 8, 689456 CrossRef PubMed.
- A. Bačić, J. Gavrilović and M. Rajilić-Stojanović, Arh. Farm., 2023, 73, 535–553 CrossRef.
- M. Fidélix, D. Milenkovic, K. Sivieri and T. Cesar, Food Funct., 2020, 11, 1599–1610 RSC.
- R. C. Barnes, H. Kim, C. Fang, W. Bennett, M. Nemec, M. A. Sirven, J. S. Suchodolski, N. Deutz, R. A. Britton and S. U. Mertens-Talcott, Mol. Nutr. Food Res., 2019, 63, 1800512 CrossRef PubMed.
- A. L. Molan, Z. Liu and M. Kruger, World J. Microbiol. Biotechnol., 2010, 26, 1735–1743 CrossRef.
- A. L. Molan, Z. Liu and G. Plimmer, Phytother. Res., 2014, 28, 416–422 CrossRef CAS PubMed.
- S. N. Hester, A. Mastaloudis, R. Gray, J. M. Antony, M. Evans and S. M. Wood, J. Nutr. Metabol., 2018, 2018, 7497260 Search PubMed.
- M. C. Rodríguez-Daza, M. Roquim, S. Dudonné, G. Pilon, E. Levy, A. Marette, D. Roy and Y. Desjardins, Front. Microbiol., 2020, 11, 2032 CrossRef PubMed.
- D. E. Roopchand, R. N. Carmody, P. Kuhn, K. Moskal, P. Rojas-Silva, P. J. Turnbaugh and I. Raskin, Diabetes, 2015, 64, 2847–2858 CrossRef CAS PubMed.
- J. Li, T. Wu, N. Li, X. Wang, G. Chen and X. Lyu, Food Funct., 2019, 10, 333–343 RSC.
- Y. Liu, L. Luo, Y. Luo, J. Zhang, X. Wang, K. Sun and L. Zeng, J. Agric. Food Chem., 2020, 68, 6368–6380 CrossRef CAS PubMed.
- S. Westfall, N. Lomis and S. Prakash, Artif. Cells, Nanomed., Biotechnol., 2018, 46, 441–455 CrossRef CAS PubMed.
- Y. Fujita, H. Tsuno and J. Nakayama, PLoS One, 2017, 12, e0169240 CrossRef PubMed.
- Z. Wang, K. L. Lam, J. Hu, S. Ge, A. Zhou, B. Zheng, S. Zeng and S. Lin, Food Sci. Nutr., 2019, 7, 579–588 CrossRef CAS PubMed.
- J. Liu, W. Hao, Z. He, E. Kwek, Y. Zhao, H. Zhu, N. Liang, K. Y. Ma, L. Lei and W. S. He, Food Funct., 2019, 10, 2847–2860 RSC.
- M. Xie, G. Chen, P. Wan, Z. Dai, X. Zeng and Y. Sun, J. Agric. Food Chem., 2018, 67, 171–183 CrossRef PubMed.
- E. B. Mojzer, M. K. Hrnčič, M. Škerget, Ž. Knez and U. Bren, Molecules, 2016, 21, 901 CrossRef PubMed.
- K. Ameer, H. M. Shahbaz and J. H. Kwon, Compr. Rev. Food Sci. Food Saf., 2017, 16, 295–315 CrossRef PubMed.
- Y. Hu, B. Yan, Z. S. Chen, L. Wang, W. Tang and C. Huang, J. Renewable Mater., 2022, 10, 1471–1490 CAS.
- U. Etxeberria, N. Arias, N. Boque, M. T. Macarulla, M. P. Portillo, F. I. Milagro and J. A. Martinez, Benefic. Microbes, 2015, 1, 97–111 Search PubMed.
- D. Porras, E. Nistal, S. Martínez-Flórez, S. Pisonero-Vaquero, J. L. Olcoz, R. Jover, J. González-Gallego, M. V. García-Mediavilla and S. Sánchez-Campos, Free Radicals Biol. Med., 2017, 102, 188–202 CrossRef CAS PubMed.
- J. Xie, W. Song, X. Liang, Q. Zhang, Y. Shi, W. Liu and X. Shi, Biomed. Pharmacother., 2020, 127, 110147 CrossRef CAS PubMed.
- S. Liu, Y. T. Loo, Y. Zhang and K. Ng, Food Chem., 2024, 434, 137508 CrossRef CAS PubMed.
- S. Lindahl, J. Liu, S. Khan, E. N. Karlsson and C. Turner, Anal. Chim. Acta, 2013, 785, 50–59 CrossRef CAS PubMed.
- I. S. Choi, E. J. Cho, J.-H. Moon and H. J. Bae, Food Chem., 2015, 188, 537–542 CrossRef CAS PubMed.
- L. Wang, Y. Wu, Y. Liu and Z. Wu, Molecules, 2017, 22, 1648 CrossRef PubMed.
- P. Mikšovsky, C. Kornpointner, Z. Parandeh, M. Goessinger, K. Bica-Schröder and H. Halbwirth, ChemSusChem, 2024, 17, e202301094 CrossRef PubMed.
- Z. Liu, Z. Ma, H. Zhang, B. S. Summah, H. Liu, D. An, Q. Zhan, W. Lai, Q. Zeng and H. Ren, Biomed. Pharmacother., 2019, 120, 109482 CrossRef CAS PubMed.
- Y. Li, Z. Xie, T. Gao, L. Li, Y. Chen, D. Xiao, W. Liu, B. Zou, B. Lu and X. Tian, Food Funct., 2019, 10, 4046–4061 RSC.
- H. Z. Zheng, I. W. Hwang and S. K. Chung, J. Zhejiang Univ., Sci., B, 2009, 10, 912–919 CrossRef CAS PubMed.
- S. Chamorro, A. Viveros, I. Alvarez, E. Vega and A. Brenes, Food Chem., 2012, 133, 308–314 CrossRef CAS PubMed.
- M.-R. Meini, I. Cabezudo, C. E. Boschetti and D. Romanini, Food Chem., 2019, 283, 257–264 CrossRef CAS PubMed.
- A. Takagaki and F. Nanjo, Biol. Pharm. Bull., 2015, 38, 325–330 CrossRef CAS.
- A. Takagaki and F. Nanjo, Biosci., Biotechnol., Biochem., 2016, 80, 199–202 CrossRef CAS PubMed.
- P. Dey, B. D. Olmstead, G. Y. Sasaki, Y. Vodovotz, Z. Yu and R. S. Bruno, J. Nutr. Biochem., 2020, 84, 108455 CrossRef CAS PubMed.
- R. Pacheco-Ordaz, A. Wall-Medrano, M. G. Goñi, G. Ramos-Clamont-Montfort, J. F. Ayala-Zavala and G. González-Aguilar, Lett. Appl. Microbiol., 2018, 66, 25–31 CrossRef CAS PubMed.
- K. N. Baruah, S. Singha, P. Mukherjee and R. V. Uppaluri, Biomass Convers. Biorefin., 2024, 14, 24407–24425 CrossRef CAS.
- Y.-H. Hong, E. Y. Jung, Y. Park, K.-S. Shin, T. Y. Kim, K.-W. Yu, U. J. Chang and H. J. Suh, Biosci., Biotechnol., Biochem., 2013, 77, 22–29 CrossRef CAS PubMed.
- V. Battestin, G. Macedo and V. De Freitas, Food Chem., 2008, 108, 228–233 CrossRef CAS.
- A. Liang, W. Leonard, J. T. Beasley, Z. Fang, P. Zhang and C. S. Ranadheera, Crit. Rev. Food Sci. Nutr., 2024, 64, 7563–7588 CrossRef CAS PubMed.
- Y. Zhu, H. Sun, S. He, Q. Lou, M. Yu, M. Tang and L. Tu, PLoS One, 2018, 13, e0195754 CrossRef PubMed.
- X. Liu, K. K. Hu and V. S. Haritos, Food Chem., 2024, 435, 137562 CrossRef CAS PubMed.
- W. Yue and F. Han, Food Chem.: X, 2022, 16, 100501 CAS.
- L. Zhang, G. Fan, M. A. Khan, Z. Yan and T. Beta, Food Chem., 2020, 323, 126714 CrossRef CAS PubMed.
- P. Amulya and R. ul Islam, Food Chem., 2023, 18, 100643 CAS.
- L. Pan, H. Ye, X. Pi, W. Liu, Z. Wang, Y. Zhang and J. Zheng, Front. Microbiol., 2023, 14, 1092729 CrossRef PubMed.
- M. Mueller, B. Zartl, A. Schleritzko, M. Stenzl, H. Viernstein and F. M. Unger, Bioprocess Biosyst. Eng., 2018, 41, 221–228 CrossRef CAS PubMed.
- A. E. Paharik, C. P. Parlet, N. Chung, D. A. Todd, E. I. Rodriguez, M. J. Van Dyke, N. B. Cech and A. R. Horswill, Cell Host Microbe, 2017, 22, 746–756 CrossRef CAS PubMed.
- K. Bier and B. Schittek, Exp. Dermatol., 2021, 30, 1442–1452 CrossRef PubMed.
- A. Stacy and Y. Belkaid, Science, 2019, 363, 227–228 CrossRef CAS PubMed.
- S. Di Lodovico, F. Gasparri, E. Di Campli, P. Di Fermo, S. D'Ercole, L. Cellini and M. Di Giulio, Microorganisms, 2020, 9, 37 CrossRef PubMed.
- C. Le Bourgot, C. Meunier, E. Gaio, V. Murat, M. Micheletto, E. Tedesco and F. Benetti, Sci. Rep., 2022, 12, 9702 CrossRef CAS.
- A. P. Ivanković, A. Milivojević, M. Ćorović, M. Simović, K. Banjanac, P. Jansen, A. Vukoičić, E. Van den Bogaard and D. Bezbradica, Chem. Biol. Technol. Agric., 2023, 10, 125 CrossRef.
- A. P. Ivanković, M. Ćorović, A. Milivojević, M. Simović, K. Banjanac, M. Veljković and D. Bezbradica, Int. J. Fruit Sci., 2024, 24, 85–101 CrossRef.
- D. Gwiazdowska, K. Juś, J. Jasnowska-Małecka and K. Kluczyńska, Acta Biochim. Pol., 2015, 62, 895–901 CrossRef CAS PubMed.
- A. Duda-Chodak, J. Physiol. Pharmacol., 2012, 63, 497–503 CAS.
- Z. Zhang, X. Peng, S. Li, N. Zhang, Y. Wang and H. Wei, PLoS One, 2014, 9, e90531 CrossRef PubMed.
- Spotlight on protein structure design, Nat. Biotechnol., 2024, 42, 157 CrossRef.
- L. Wang, K. Cao, M. M. Pedroso, B. Wu, Z. Gao, B. He and G. Schenk, J. Biol. Chem., 2021, 297, 101262 CrossRef CAS.
- S. M. Hancock, M. D. Vaughan and S. G. Withers, Curr. Opin. Chem. Biol., 2006, 10, 509–519 CrossRef CAS PubMed.
- G. S. Bulmer, P. de Andrade, R. A. Field and J. M. van Munster, Carbohydr. Res., 2021, 508, 108411 CrossRef CAS PubMed.
- Z. Mészáros, P. Nekvasilová, P. Bojarová, V. Kren and K. F. Slámová, Biotechnol. Adv., 2021, 49, 107733 CrossRef PubMed.
- L. Bode, Glycobiology, 2012, 22, 1147–1162 CrossRef CAS.
- J. T. Smilowitz, C. B. Lebrilla, D. A. Mills, J. B. German and S. L. Freeman, Annu. Rev. Nutr., 2014, 34, 143–169 CrossRef CAS PubMed.
- X. Chen, D. C. Baker and D. Horton, Adv. Carbohydr. Chem. Biochem., 2015, 72, 113–190 CrossRef PubMed.
- K. Schmölzer, M. Weingarten, K. Baldenius and B. Nidetzky, ACS Catal., 2019, 9, 5503–5514 CrossRef.
- C. André-Miral, F. M. T. Koné, C. Solleux, C. Grandjean, M. Dion, V. Tran and C. Tellier, Glycobiology, 2015, 25, 394–402 CrossRef PubMed.
- X. Tong, T. Li, C. Li and L.-X. Wang, Biochem, 2018, 57, 5239–5246 CrossRef CAS.
- T. B. Parsons, W. B. Struwe, J. Gault, K. Yamamoto, T. A. Taylor, R. Raj, K. Wals, S. Mohammed, C. V. Robinson, J. L. P. Benesch and B. G. Davis, Angew. Chem., Int. Ed., 2016, 55, 2361–2367 CrossRef CAS.
- K. Slámová, J. Krejzová, P. Marhol, L. Kalachova, N. Kulik, H. Pelantová, J. Cvačka and V. Křen, Adv. Synth. Catal., 2015, 357, 1941–1950 CrossRef.
- T. Ito, T. Katayama, M. Hattie, H. Sakurama, J. Wada, R. Suzuki, H. Ashida, T. Wakagi, K. Yamamoto, K. A. Stubbs and S. Fushinobu, J. Biol. Chem., 2013, 288, 11795–11806 CrossRef CAS PubMed.
- M. E. Ortiz-Soto, J. Ertl, J. Mut, J. Adelmann, T. A. Le, J. Shan, J. Teßmar, A. Schlosser, B. Engels and J. Seibel, Chem. Sci., 2018, 9, 5312–5321 RSC.
- J. M. Palomo, Org. Biomol. Chem., 2012, 10, 9309–9318 RSC.
- M. Filice, O. Romero, A. Aires, J. M. Guisan, A. Rumbero and J. M. Palomo, Adv. Synth. Catal., 2015, 357, 2687–2696 CrossRef CAS.
- N. Losada-Garcia, E. P. Urriolabeitia and J. M. Palomo, ACS Appl. Nano Mater., 2023, 6, 704–713 CrossRef CAS.
- N. Losada-Garcia, E. P. Urriolabeitia and J. M. Palomo, ChemCatChem, 2023, 15, e202201632 CrossRef CAS.
- C. Mateo, J. M. Palomo, G. Fernandez-Lorente, J. M. Guisan and R. Fernandez-Lafuente, Enzyme Microb. Technol., 2007, 40, 1451–1463 CrossRef CAS.
- J. M. Palomo, G. Fernández-Lorente, C. Mateo, R. Fernández-Lafuente and J. M. Guisan, Tetrahedron: Asymmetry, 2002, 13, 2375–2381 CrossRef CAS.
- R. C. Rodrigues, C. Ortiz, A. Berenguer-Murcia, R. Torres and R. Fernandez-Lafuente, Chem. Soc. Rev., 2013, 42, 6290–6307 RSC.
- J. M. Guisan, G. Fernandez-Lorente, J. Rocha-Martin and D. Moreno-Gamero, Curr. Opin. Green Sustainable Chem., 2022, 35, 100593 CrossRef CAS.
- Y. Zhang, J. Ge and Z. Liu, ACS Catal., 2015, 5, 4503–4513 CrossRef.
- C. C. Aragon, A. I. Ruiz-Matute, N. Corzo, R. Monti, J. M. Guisán and C. Mateo, Int. Food Nutr. Metabol., 2018, 5, 1–9 Search PubMed.
- L. Xue, J. Long, C. Lu, X. Li, X. Xu and Z. Jin, Food Biosci., 2021, 39, 100837 CrossRef CAS.
- X. Chen, Z. Tian, H. Zhou, G. Zhou and H. Cheng, Foods, 2023, 12, 3089 CrossRef CAS PubMed.
- R. K. Suryawanshi, U. K. Jana, B. P. Prajapati and N. Kango, Food Chem., 2019, 289, 95–102 CrossRef CAS PubMed.
- S. Chalane, C. Delattre, P. Michaud, A. Lebert, C. Gardarin, D. Kothari, C. Creuly, A. Goyal, A. Štrancar and G. Pierre, Process Biochem., 2017, 58, 105–113 CrossRef CAS.
- P. V. Hooda, Int. J. Biol. Macromol., 2018, 106, 1173–1183 CrossRef PubMed.
- L. Ruzic, J. M. Bolivar and B. Nidetzky, Biotechnol. Bioeng., 2020, 117, 1597–1602 CrossRef CAS.
- J. M. Woodley, Comput. Chem. Eng., 2017, 105, 297–307 CrossRef CAS.
- O. A. Prado-Rubio, R. Morales-Rodríguez, P. Andrade-Santacoloma and H. Hernández-Escoto, Process Intensification in Biotechnology Applications, in Process Intensification in Chemical Engineering, ed. J. Segovia-Hernández and A. Bonilla-Petriciolet, Springer, Cham, 2016 Search PubMed.
- K. V. K. Boodhoo, M. C. Flickinger, J. M. Woodley and E. A. C. Emanuelsson, Chem. Eng. Process., 2022, 172, 108793 CrossRef CAS.
- H.-J. Shin, J.-M. Park and J.-W. Yang, Process Biochem., 1998, 33, 787–792 CrossRef CAS.
- A. Warmerdam, E. Benjamins, T. Leeuv, T. Broekhuis, R. Boom and A. Janssen, Food Bioprod. Process., 2014, 92, 383–392 CrossRef CAS.
- S. G. A. Lorenzoni, L. F. Aydos, M. P. Klein, M. A. Z. Ayub, R. C. Rodrigues and P. F. Hertz, J. Mol. Catal. B: Enzym., 2015, 111, 51–55 CrossRef.
- P. Zambelli, L. Tamborini, S. Cazzamalli, A. Pinto, S. Arioli, S. Balzaretti, F. J. Plou, L. Fernandez-Arrojo, F. Molinari, P. Conti and D. Romano, Food Chem., 2016, 190, 607–613 CrossRef CAS PubMed.
- I. Petzelbauer, B. Splechtna and B. Nidetzky, Biotechnol. Bioeng., 2002, 77, 394–404 CrossRef CAS.
- A. Córdova, C. Astudillo, C. Vera, C. Guerrero and A. Illanes, J. Biotechnol., 2016, 223, 26–35 CrossRef PubMed.
- I. Pottratz, I. Müller and C. Hamel, Catalysts, 2022, 12, 7 CrossRef CAS.
- D. Sen, A. Sarkar, S. Das, R. Chowdhury and C. Bhattacharjee, Ind. Eng. Chem. Res., 2012, 51, 10671–10681 CrossRef CAS.
- D. Slavnić, Dynamics of liquid and solids flow in oscillatory flow reactors and application to biochemical reaction with immobilized enzymes, PhD thesis, University of Belgrade, 2020 Search PubMed.
- B. Todić, R. Pravilović and N. Nikačevic, Chem. Eng. Process., 2024, 203, 109895 CrossRef.
- B. Todić, R. Pravilović, D. Bezbradica and N. Nikačević, Design and simulations of a spiral oscillatory baffled reactor for biochemical reactions, 14th European Congress of Chemical Engineering and 7th European Congress of Applied Biotechnology, 17th–21st September, 2023,Berlin, Germany.
- R. Pravilović, Intensification of the continuous prebiotic oligosaccharide production process, PhD thesis, University of Belgrade, 2024 Search PubMed.
- R. Pravilović, T. Janković, C. M. C. Aimacaña, B. Todić, M. Simović, D. Bezbradica and N. Nikačević, Optimization of an intensified reactor for the continuous production of prebiotics, 14th European Congress of Chemical Engineering and 7th European Congress of Applied Biotechnology, 17–21 September, 2023, Berlin, Germany.
- D. Ghosh, A. B. Vir, G. Garnier, A. F. Patti and J. Tanner, Chem. Eng. Sci., 2021, 244, 116789 CrossRef CAS.
- K. Elst, N. Babbar, S. Van Roy, S. Baldassarre, W. Dejonghe, M. Maesen and S. Sforza, Bioprocess Biosyst. Eng., 2018, 41, 1717–1729 CrossRef CAS PubMed.
- C. Pénicaud, V. Monclus, B. Perret, S. Passot and F. Fonseca, J. Cleaner Prod., 2018, 184, 847–858 CrossRef.
- E. Gerbino, C. Quentier and C. Pénicaud, Data Brief, 2022, 43, 108478 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2025 |