
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceProfile of steroid metabolites in human breast milk in different stages of lactation†
Olha
Khymenets
 *a,
Oscar
Vilarroya
bc,
Georgina
Benet
c,
Georgina
Feixas
d,
Angela
Arranz Betegon
d,
Malcolm D.
McLeod
e and
Oscar J.
Pozo
*a
*a,
Oscar
Vilarroya
bc,
Georgina
Benet
c,
Georgina
Feixas
d,
Angela
Arranz Betegon
d,
Malcolm D.
McLeod
e and
Oscar J.
Pozo
*a
aApplied Metabolomics Research Group, Neurosciences Research Programme, Hospital del Mar Research Institute, Barcelona, Spain. E-mail: okhymenets@researchmar.net; opozo@researchmar.net
bDepartment of Psychiatry and Legal Medicine, Autonomous University of Barcelona, Barcelona, Spain
cNeuroimaging Research Group, Neurosciences Programme, Hospital del Mar Research Institute, Barcelona, Spain
dBCNatal – Barcelona Center for Maternal-Fetal and Neonatal Medicine, Hospital Clínic and Hospital Sant Joan de Deu, Institut Clinic de Ginecologia, Obstetricia i Neonatologia, IDIBAPS, University of Barcelona, Barcelona, Spain
eResearch School of Chemistry, Australian National University, Canberra, Australia
First published on 25th April 2025
Abstract
Breast milk (BM), as an optimal food, provides the newborn with a variety of minor compounds relevant for health and wellbeing. Endogenous steroids, also minor constituents, are mainly secreted in BM as conjugated metabolites. Recent research has revealed the relevance of steroid conjugates in many physiological processes. Thus, their presence in BM appears to be very intriguing, especially in relation to breastfeeding. The objective of our study was to profile conjugated steroid metabolites present in BM in relation to the lactation stage, and to promote further evaluation of their importance in breastfeeding. For this purpose, we developed and used a direct UHPLC-MS/MS metabolomics approach capable to detect more than 60 conjugated metabolites (mono-sulfated, mono-glucuronylated, bis-sulfated and sulfate-glucuronylated) from all steroid families. We compared the occurrence of these metabolites in samples collected from breastfeeding mothers and stratified by lactation stages: colostrum, transitional and mature milk. Our results showed that many biologically relevant conjugated steroids are secreted in BM. Their concentrations were highest in colostrum, decreased remarkably in transitional and were much lower in mature milk, with some exceptions. The profile of metabolites also differed considerably between lactation stages. The approximate daily secretion in BM indicated that infants are exposed to significant oral doses of steroid conjugates during the first week of lactation. The supply of these metabolites in BM declined and became constant after the second week postpartum. Overall, our data provide a foundation for further investigation on the physiological relevance of BM secreted steroid metabolites in relation to both mother and child.
Introduction
Breast milk (BM) is one of the most intriguing biological fluids from a physiological perspective. The complexity and dynamic nature of its composition are directly related to its biological action: it is not only an age-adjusted supply of nutritive factors,1,2 but also a rich source of bioactive molecules, components and other constituents with known and anticipated biological relevance, altogether responsible for the foundation of the health and wellbeing of the baby.1,3–5 Recent research underscores the importance of studying and identifying the roles of BM's minor components.6,7 In this context, endogenous steroids, which are important players in many biological processes, merit special attention.Steroid hormones play a crucial role in many physiological processes and their presence as minor components in BM has been occasionally reported over the past decades.8–12 Most steroids exist in biological tissues and fluids, not as free compounds, but as phase II biotransformed molecules. Phase II metabolism involves the conjugation of steroids with highly polar sulfate or glucuronide groups. Among steroids, conjugated metabolites, especially sulfates, serve as transportable and deployable units in the turnover of steroid hormones, and can even act as bioactive molecules.13–16 Additionally, other conjugates, such as glucuronylated metabolites, are indicators of steroid hormonal fluctuations related to different physiological stages.17–19
BM composition, including its minor components, changes in relation to maturation of the mammary epithelium throughout lactation.1,4 A few studies have provided very partial information on the secretion of steroid metabolites in different physiological stages of lactation, mainly revealing that BM steroid content related to plasma levels of breastfeeding mothers.8,20,21 Despite the fact that recent studies on steroids indicate that some conjugated metabolites may have relevant biological activities,13,14,16 the role of these compounds in relation to BM feeding benefits has not been properly investigated, primarily due to the lack of information on the secretion of conjugate steroids in BM.
Based on this, we aimed to explore in detail the profile of conjugated steroids secreted in BM and how it varies across different lactation stages. Recent studies in steroid detection indicated that steroid metabolite profiling can be more informative than analysis focused on limited targets.22,23 Additionally, the indirect detection of conjugated steroids based on the enzymatic deconjugation of the phase II metabolite and the determination of the free steroid presents methodological challenges.22,24 For these reasons, we aimed to apply a cutting-edge methodological approach based on the direct detection of a large set of conjugated steroids using UHPLC-MS/MS to advance the exploration of the time- and the content-dependent steroidal composition of BM. The findings from our study could provide evidence on the complexity of BM as physiological fluid and contribute to ongoing research into the relevance of BM minor constituents, including the steroid component, in mother-child relationship and the baby's wellbeing.7
Experimental
Reagents and chemicals
Aqueous ammonia solution (25%), methanol, acetonitrile and formic acid (LC-MS grade) were purchased from Merck (Darmstadt, Germany). Phosphoric acid and ammonium formate were provided by Sigma-Aldrich (Louis, MO, USA) and VWR Prolabs Chemicals (Leuven, Belgium), respectively. Ultrapure water was obtained using a Milli-Q purification system (Millipore Ibérica, Barcelona, Spain).Standards
The general representation of the studied steroid metabolites is given in Fig. 1, while specific structural characteristics of the conjugated metabolites can be found in Table S1 (ESI†). For the determinations of these steroid metabolites, three types of analytical standards were used in this study: quantitative purchased, quantitative on-customer demand synthetized and qualitative in-house synthetized standards. | ||
| Fig. 1 Diagram representing diversity of steroid metabolites identified in BM in the present study. These metabolites belong to 6 steroid families: estrogen (ESTR), 4-androsten (4A), 5-androsten (5A), 5-pregnen or pregnenolone (5P), 4-pregnen or progesterone (4P) and corticoid or corticosteroid (CORT), backbone structures of which are presented in the coloured spaces. The numbers on the structures represent the steroid carbons, subjects to metabolic modifications specific for each steroid family. The text under the structures gives short descriptions of the main groups of steroid metabolites according to modified carbon bounds, where group-specific modifications are highlighted in bold. Thus, ESTR family is represented by three metabolic groups: E1 – estrone; E2 – estradiol and E3 – estriol; 4A family by four groups: AN – androsterones; Etio – etiocholanolones; epiAN – epiandrosterones; T/epiT – (epi)testosterone; 5A family by two groups: DHEA – dehydroepiandrosterone and 5AED – 5-androstenediol; 5P family by two groups: 5P – pregnenolone and 5PED – 5-pregnendiol; 4P family by four groups: alloP – allopregnanolone; P – pregnanolone; EAP – epiallopregnanolone and epiP – epipregnanolone; CORT family by three groups: E – cortisone; F – cortisol and B – corticosterone. All these steroid metabolites are conjugated either with residues of sulfate(s) (S) or glucuronic acid (G) on the positions indicated on the arrows directed to each steroid family. For individual structures of each conjugated metabolite detected in the present study, please see ESI Table S1.† | ||
Quantitative conjugated steroid standards were obtained as indicated in Table S2 (ESI†). In-house synthesis of qualitative sulfated standards was performed as previously described25 with small modifications. Details on synthesis procedure, reagents and standards can be found in the ESI.† All standards, either purchased or synthesized, were diluted in 100% methanol and stored at −20 °C prior to use.
Due to the lack of commercially available standards and/or complexity of the synthesis, several metabolites were putatively assigned on the basis of their resemblance in chromatographic and mass spectrometric behaviour to structurally and metabolically similar but known compounds (i.e. quantitative and qualitative standards). The LC-MS characteristics of studied compounds and corresponding internal standards (ISTD) are given in Table S2 (ESI†).
Study population
Sixty-two healthy lactating mothers (aged 20–50) kindly donated a part of their BM for the present study. This research was approved by the Ethics Committee at the Hospital del Mar Research Institute (Ref: 2017/7450/I). Participants were enrolled in the study in a period between 03/11/2021 and 14/12/2021, after signing a consent form. Two volunteers were posteriorly excluded from the study due to analytical problems. Detailed information on the volunteers and breast feeding is disclosed in the Table S3 (ESI†).BM samples collection
Milk was collected from one breast, either totally (whole milk = foremilk + hind milk), or partially, when only first portion of secreted milk (foremilk sample) was used as a sample. In the case of using whole milk, an aliquot was separated for the study while the rest was returned for baby feeding. The volume of collected BM aliquots was accurately measured.Samples were labelled and stored at −20 °C until analysis. According to the collection date, collected BM samples were classified in three types: colostrum (1–6 days postpartum), transitional (7–19 days postpartum) and mature (20–50 days postpartum). Two samples (one colostrum and one transitional) were lost during processing, therefore 60 samples, grouped by 18, 17 and 25 samples corresponding to colostrum, transitional and mature milks respectively, were available for data analysis.
Samples preparation
BM thawed at room temperature was well vortexed and an aliquot of 500 μL was spiked with 10 μL of internal standard mix (containing a mixture stably labelled conjugated steroids listed in Table S1, ESI†) and proteins were precipitated using 3 volumes of 3![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :7 (v:v) methanol/acetonitrile. The mixture was vigorously vortexed, left for 30 min at room temperature, and then centrifuged at 10000g for 5 min. The supernatant was separated from pellet and dried under N2 at 40 °C. The residue was reconstituted in 1 mL of water:acetonitrile (9:1), acidified with 1 mL solution of 4% aqueous phosphoric acid and proceeded to solid phase extraction using Oasis HLB 3cc cartridges (Waters Associates, Milford, MA, USA) previously pre-conditioned with 100% methanol and 2% formic acid in water. After a washing step with 2 mL of 2% formic acid in water, steroid conjugates were eluted into two steps: firstly, with 1 mL of 2% formic acid in methanol following with 1 mL 100% methanol, then, secondly, with 1 mL 5% ammonia in methanol following with 1 mL 100% methanol. Both eluates (acidic and basic) were evaporated separately under N2 at 40 °C. After evaporation, each residue was reconstituted in 1 mL methanol, and both extracts, corresponding to the same sample, were combined. The obtained mixture was evaporated under N2 at 40 °C, reconstituted in 100 μL of water:acetonitrile (9:1), and 10 μL were injected into the UHPLC-MS/MS system.
:7 (v:v) methanol/acetonitrile. The mixture was vigorously vortexed, left for 30 min at room temperature, and then centrifuged at 10000g for 5 min. The supernatant was separated from pellet and dried under N2 at 40 °C. The residue was reconstituted in 1 mL of water:acetonitrile (9:1), acidified with 1 mL solution of 4% aqueous phosphoric acid and proceeded to solid phase extraction using Oasis HLB 3cc cartridges (Waters Associates, Milford, MA, USA) previously pre-conditioned with 100% methanol and 2% formic acid in water. After a washing step with 2 mL of 2% formic acid in water, steroid conjugates were eluted into two steps: firstly, with 1 mL of 2% formic acid in methanol following with 1 mL 100% methanol, then, secondly, with 1 mL 5% ammonia in methanol following with 1 mL 100% methanol. Both eluates (acidic and basic) were evaporated separately under N2 at 40 °C. After evaporation, each residue was reconstituted in 1 mL methanol, and both extracts, corresponding to the same sample, were combined. The obtained mixture was evaporated under N2 at 40 °C, reconstituted in 100 μL of water:acetonitrile (9:1), and 10 μL were injected into the UHPLC-MS/MS system.
The method provided satisfactory recoveries for different types of conjugates in all three types of breast milk (commonly >75%, see Table S4, ESI†).
UHPLC-MS/MS instrumentation
The separation of BM extracts was performed at flow rate of 400 μL min−1 at 30 °C on an Acquity UPLC Class I chromatographic system (all from Waters Associates) using an Acquity UPLC CSH C18 column (2.1 × 100 mm i.d., 1.7 μm) fixed to Acquity UPLC CSH C18 VanGuard Pre-Column (2.1 × 5 mm i.d., 1.7 μm) (all from Waters Associates). Acetonitrile:water (9:1, v/v) and water, both with ammonium formate (25 mM), were used as organic and aqueous mobile phase solvents, respectively. The gradient elution program was applied, where the percentage of organic solvent was linearly changed as follows: 0 min, 10%; 0.5 min, 10%; 15 min, 47%; 15.5 min, 100%; 18.5 min, 100%; 19 min, 10%; 20 min, 10%. In total, the chromatographic method lasted 20 min and allowed for the separation of isomeric steroid conjugates.
MS/MS analysis was carried out on a triple quadrupole (XEVO TQ-S micro) mass spectrometer equipped with an orthogonal Z-spray-electrospray ionization source from Waters Associates. Nitrogen was used as drying and nebulizing gas. The desolvation gas flow was 1200 L h−1, and the cone gas flow was 50 L h−1. A cone voltage of 20 V and a capillary voltage of 2 kV were used in negative ionization mode. The nitrogen desolvation temperature was 600 °C, and the source temperature was 150 °C. The monitoring and quantification of all analytes was performed using selective reaction monitoring mode. The specific transitions and optimum collision energy are summarized in Table S2 (ESI†) and an example of chromatogram for each detected steroid metabolite is given in Fig. S1 (ESI†).
Quantitative evaluation of metabolites
To overcome heterogeneous matrix impact of different types of milk (colostrum, transitional and mature) on quantitative analysis of steroid metabolites, three pools of corresponding BM samples were created by mixing equal volumes of each individual sample corresponding to the same type of BM. These milk-type adjusted pools were used as biological matrices to build matrix-matched calibration curves using available quantitative standards (Table S4, ESI†). Reference calibration curves were also built in reconstituting solvent.The response of the analyte was calculated as the ratio between the areas of analyte and corresponding internal standard (see Table S4, ESI†). The concentration of steroid metabolite was calculated using a matrix-matched calibration curve built in duplicate. Endogenous metabolites, for which quantitative standards were not available were relatively quantified using calibration curves of available standards structurally similar (see Table S4, ESI†), assuming equal response for used transitions. Details on the method performance (e.g. estimation of limit of detection, recoveries and matrix effect) for the determination of phase II steroid conjugates in BM are given Table S4 (ESI†).
Estimation of daily steroid consumption
The daily consumption of conjugated steroids through BM was estimated on the basis of reported amount of milk secreted (mL 24 h−1) on average over the postpartum period26 in a specific period (1–53 days). Concentration data (nmol L−1) of detected steroid metabolites from BM samples grouped by postpartum days were normalized by the average volume of BM reported to be produced on those days.26 Estimated amounts of 24 h consumed steroid phase II metabolites via BM were expressed in μmol 24 h−1.Data analysis
MassLynx V4.1, TargetLynx XS software (Waters) and Excel (Microsoft) were used for data management, qualitative and quantitative analysis. An open source RawGraphs platform,27 PowerPoint and Excel (Microsoft) were used for data visualization purposes.Given that the steroid metabolite concentration levels in BM samples were very widespread, data were expressed as ranges for descriptive and comparative purposes, whereas zero-adjusted (substituting zero values by one half of the lowest values detected for each metabolite) and log-transformed data were used in statistical analysis and for global imaging. Percentages and daily secretions were calculated using mean values of zero-adjusted concentrations and were used for graphical data visualization and assessment.
A nonparametric Krustal–Wallis test on independent samples was applied on log-transformed data to compare concentrations of steroid conjugated metabolites between three types of BM. Statistical significance was considered when p < 0.05.
Results
Phase II steroid metabolites identified in BM
Owing to the developed methodology, we could detect within one instrumental analysis more than 60 conjugated metabolites of steroids in a relatively small volume of BM sample (0.5 mL). The detected metabolites correspond to various steroid families: 11 estrogens (ESTR), 5 pregnenolone derived metabolites (5P), 22 progesterone derived metabolites (4P), 9 5-androstene derived metabolites (5A), 13 4-androstene derived metabolites (4A), and 6 corticosteroids (CORT). They were represented by a wide spectrum of mono-conjugated (34 sulfates and 12 glucuronides) and bis-conjugated (12 bis-sulfates and 8 sulfate–glucuronide) metabolites (Fig. S1, ESI†). Mono-sulfated, mono-glucuronylated and to lesser extent bis-sulfated metabolites were omnipresent in BM samples, whereas sulfate–glucuronide metabolites were rarely detected, predominantly within colostrum samples.Levels of the steroid phase II conjugates in BM
Concentrations of phase II steroid metabolites detected in BM are given in Table S5 (ESI†). A comprehensive representation on concentrations of detected steroid conjugates in breast milk samples of the present study is presented in Fig. 2. | ||
| Fig. 2 Levels of endogenous steroid phase II conjugates (expressed in logarithmic scale because of the large differences in concentrations between samples and between metabolites) detected in tree types of BM from our population study: colostrum samples are marked as the darkest dots, transitional as intermediate intensity and mature as the lightest dots, coloured in correspondence to six main steroid families presented in the figure legend. For details on detected steroid conjugates, please refer to the ESI.† Metabolites that did not correspond to the metabolic groups (AD-SS, PD-G_1 and a set of PD-SGs) are not presented on this figure. | ||
In general, the highest concentrations of steroid phase II metabolites were detected in colostrum samples (range of 41.71–11602.65 nmol L−1). In contrast, significantly lower concentrations were observed in samples corresponding to transitional (32.78–1669.70 nmol L−1 range) and mature (24.32–152.70 nmol L−1 range) BM. Such decreases, sometimes over ten-fold, was characteristic for all types of conjugates with the exception of some androgen metabolites (e.g. androstenediol bis-sulfates) for which no differences were observed between BM types according to Krustal–Wallis test (p-values ≥ 0.05) (Table S5, ESI†). Levels of specific steroid metabolites in BM samples were of diverse concentration ranges and, in addition, they varied highly between individual samples (Table S5, ESI†).
Regarding phase II conjugation, the most abundant in BM samples were sulfated forms of steroids, with mono-sulfates the most copious metabolites irrespective of the BM type (Table S5 (ESI†) and Fig. 3). They were detected in samples within 37.78–10218.74 nmol L−1, 16.21–1588.79 nmol L−1 and 18.040–135.73 nmol L−1 concentration ranges, corresponding to colostrum, transitional and mature BM, accounting for about 90%, 87% and 79% of all quantified steroid metabolites, respectively. Bis-sulfates were also found to be abundant with concentrations ranging 1.11–578.23 nmol L−1, 3.80–62.60 nmol L−1 and 3.06–25.49 nmol L−1 which account for about 5%, 9% and 16% of total detected metabolites in colostrum, transitional and mature BM, respectively. In contrast to mono-sulfated steroid metabolites, overall concentrations of mono-glucuronides were much lower, nearly tenfold, within 1.21–804.56 nmol L−1, representing only up to 5% of detected metabolites in all three BM groups. The sulfate–glucuronide metabolites were a minor fraction of steroid conjugates observed in BM, not exceeding 10.72 nmol L−1 and accounting for less than 0.1% of all quantified conjugates. They were principally detected within colostrum samples. In contrast, bis-sulfated metabolites were commonly detected in the majority of samples irrespective of the type of BM (Table S5, (ESI†), and Fig. 3).
 | ||
| Fig. 3 Proportion of phase II conjugates (S – mono-sulfates, SS – bis-sulfates, G – mono-glucuronides and SG – sulfate–glucuronides) in three types of BM (expressed as % of total detected in each type of milk) and contributions of each steroid family to the pools of specific conjugated metabolites. | ||
Incidence of phase II metabolites from different steroid families in BM
The contribution of steroid families to the pools of specifically conjugated metabolites changed over the period of lactation and is represented in the Fig. 3. Additionally, the impact of individual metabolites in relation to input of its family and metabolic pathway (as a % of total detected metabolites) was evaluated in different stages of lactation and represented on the Fig. 4.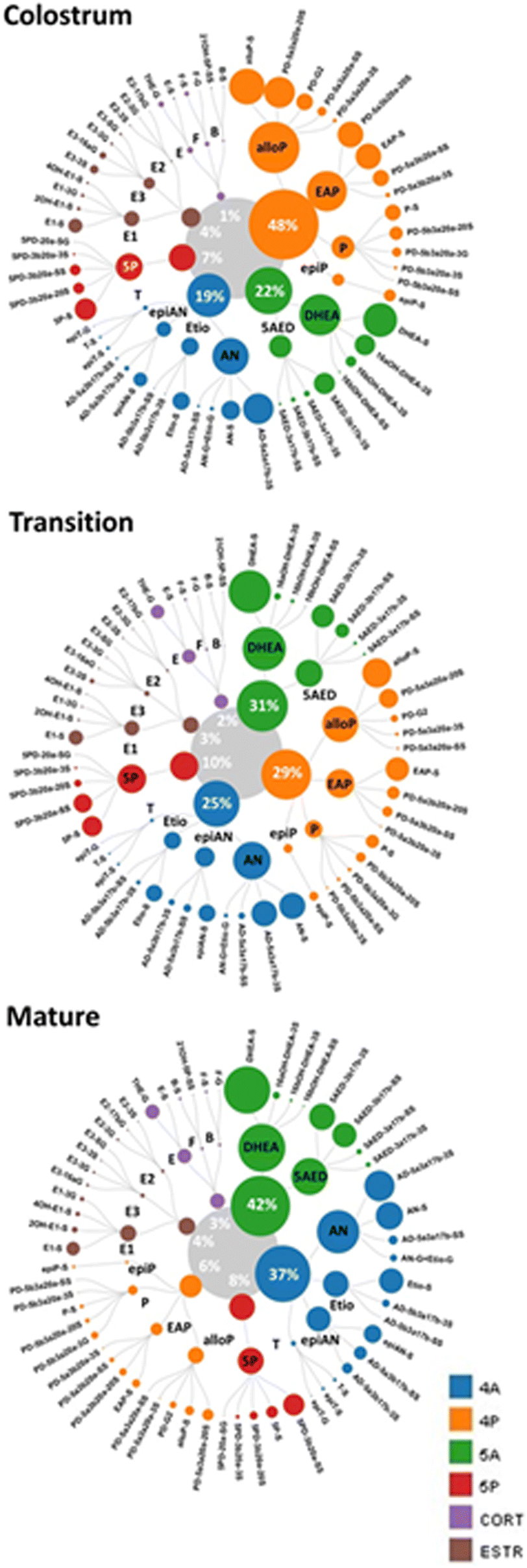 | ||
| Fig. 4 Proportion of steroid phase II conjugates in three types of BM (expressed as % of total detected in each type of milk). The contribution of each steroid family is given in numbers on the figure. The conjugated metabolites within each family are assembled according to the corresponding metabolic group (for details see Fig. 1). Steroid families, metabolic groups and steroid metabolites are presented according to their relevance (the size of the corresponding circle) in three type of BM in descending order. | ||
Regarding 5A metabolites, the relative amounts of dehydroepiandrosterone sulfate (DHEA-S), a core androgen, increased through lactation, positioning it as a principal steroid found in transitional and mature BM. The concentrations and the incidence of 16-hydroxilated conjugated metabolites of DHEA decreased drastically already from colostrum to transitional BM (Table S5, (ESI†)). Along with this, 5-androstenediol (5AED) metabolites incidence markedly increased in BM from later stages of lactation (Fig. 4). Interestingly, 5AED-3β17β-SS was the only steroid conjugate clearly maintaining concentration levels in all three types of BM, and which positioned as one of the principal steroid metabolites in mature BM (Table S5, (ESI†) and Fig. 4).
The specific contribution of steroid families to the proportion of the sulfate–glucuronide fraction in the different types of BM could not be properly characterized due to the limited data obtained on these conjugates Table S5, (ESI†). So far, it can be mentioned that estriol, 5-pregnenediol and pregnanediol metabolites were the only identified steroids contributing to this minor fraction regardless of type of BM (Fig. 3).
Daily consumption of the steroid metabolites through BM
A summary on estimated daily consumption of phase II steroid in BM over first months of lactation (1–53 postpartum days) is summarized in Fig. 5. | ||
| Fig. 5 Approximated 24 h secretion of phase II endogenous steroid metabolites (μmol 24 h−1) in BM over different stages of lactation accordingly to the detected concentrations of metabolites in BM samples from study population and reported excretion volumes (mL 24 h−1) of BM (according to Neville26) from first till 53-rd day of postpartum period. Corresponding numerical data can be found in Table S6 (ESI†). | ||
According to our approximation, in total steroid metabolites were consumed via BM on the average 674–937 μmol 24 h−1 during the first week of lactation (Table S6, (ESI†)). With an onset of lactation, concentrations of steroids decrease drastically, while daily BM consumption volume markedly increases (in general exceeding 500 mL per day (ref. 26)). Steroid daily consumption via BM decreases almost ten times when lactation becomes established. According to our approximation, the peak of daily consumption of steroid metabolites with BM by new-borns would correspond to days 4–6 postpartum (Fig. 5). The details on daily consumption of each family metabolite through lactation establishment are presented in Table S6 (ESI†).
As stated above, progesterone metabolites accounted for the largest part of daily consumed steroids during onset of lactation (first week) reaching, in some cases, 2417 μmol per day (Table S6, ESI†), which for a normal weight new-born (approximately 3 kg weight) would correspond to a daily dose of more than 800 μmol kg−1 consumed with BM. Daily BM consumption of estrogens and pregnenolone metabolites decreased ten times after the first week of lactation, dropping to the unit μmol 24 h−1 level. After onset of lactation, both androgen family metabolites were predominantly secreted on daily bases in BM in the dozen μmol 24 h−1 range. Daily consumption of corticosteroids metabolites did not vary very much during lactation with concentrations in unit μmol 24 h−1 level (Fig. 5 and Table S6, ESI†).
Discussion
In our study, the concentrations of endogenous steroid conjugates in BM were estimated in relation to the stages of lactation. By applying a developed targeted metabolomics approach for the direct detection of a wide variety of phase II conjugated steroids, we report time- and content-dependent differences in the steroidal components of BM at different stages of lactation. We also approximate the evolution of daily intake of the detected steroid metabolites by breastfed infants.The detection of steroids in BM, especially those with known physiological actions, has encouraged researchers to further explore their biological relevance and potential consequences for both maternal and infant health.12,28–33 Although these studies focused primarily on well-known steroid hormones, they collectively suggest that BM comprises a complex mixture of steroids and their metabolites, which remains largely underexplored. A comprehensive description of the BM steroidal pool has only been reported once, in a very small sample set by Sahlberg and Axelson,20 which highlighted that steroids in BM are primarily present as phase II conjugated metabolites. Therefore, our study represents a step forward in advancing knowledge on the presence of steroids in BM opening new avenues for exploring the role of these metabolites in maternal and infant health.
Our study is the first to report the concentration of steroid conjugates in BM including mono-sulfates, bis-sulfates, glucuronides and sulfate–glucuronide conjugates. Recent research has highlighted the importance of these conjugates not only as markers of steroid turnover within an organism, but as primary players in establishing and maintaining various fundamental biological processes.16,22,34 Our results were obtained using by a targeted analytical approach based on the direct detection of more than 60 steroid metabolites. To our knowledge, this is the first LC-MS method with such separation power capable of detecting a wide range of steroid metabolites from diverse families and with different types of conjugation, including highly polar bis-conjugated metabolites. In contrast, most previous studies have analysed steroid metabolites indirectly by monitoring deconjugated species after hydrolysis. Hydrolysis itself can introduce bias in steroid detection, while also compromising compound stability and recovery.24,35 Additionally, the exact nature of some conjugated species cannot be precisely identified through indirect approaches. Therefore, our study provides unique information regarding steroid disposition in BM.
Occurrence and dynamics of steroid conjugates in BM
Our results show an abrupt reduction in the concentrations of the majority of targeted steroid metabolites, with the exception of some androgens, after the first postpartum weeks. Changes in both total and individual content of steroid conjugated metabolites distinguish colostrum from transitional and mature BM. Such a decline in concentration through lactation was reported earlier for 17-keto steroids and estrogens,36–38 and was also observed for conjugated metabolites of estrogens21,33 and other steroids20 already during the first week of lactation. Our findings confirm that throughout all phases of lactation, the vast majority of BM steroids are sulfate-conjugated (over 90% of metabolites detected in the present study), mono-sulfates being the most prevalent ones, followed by bis-sulfated steroids. It should be mentioned that a higher proportion of bis-sulfates and lower proportion of mono-sulfates are found in the later stages of lactation, compared to colostrum milk. This aligns with earlier published data,20 despite the use of indirect determination, very small sample size and some differences in steroid conjugates analysed. In contrast to sulfated steroids, glucuronylated metabolites represent a smaller but relatively stable fraction of milk steroids. The predominant excretion of sulfates is interesting since some steroid mono-sulfates can exert specific biological activities themselves or as locally liberated bioactive steroids following sulfatase hydrolysis.15,39 Additionally, recent studies show that bis-sulfates conjugates seem to be products of advanced steroid metabolism with yet undefined biological impact.40 Given that the lactation is evolutionarily a well-established process, it would be of great interest to understand the physiological meaning of the consumption of potentially active sulfated steroids with BM by newborns in the first postnatal weeks, as the most crucial period for establishing the foundation for long-term infant health and well-being.Our results also show that variations of the BM steroid composition throughout lactation substantially depend not only on the steroid family but also on specific metabolic pathway and/or origin of synthesis.
The decline in phase II progesterone metabolite concentrations and their shifting relative importance among BM conjugated metabolites over lactation aligns with the decrease observed in serum and BM postpartum progesterone.8 This may reflect changes occurring in the mother in progesterone metabolism after delivery. Specifically, the high concentration of sulfated 5α-reduced progesterone metabolites (alloP-S and EAP-S and corresponding pregnanediols) in colostrum, also reported earlier,20 could be the result of accumulation of these metabolites in breast tissue/milk during the initial phase of lactogenesis, which begins in late pregnancy.41 Alternatively, this could result from the prolonged secretion and, therefore, metabolism of the pregnancy-related progesterone.8,41,42 After placental elimination, which removes the principal source of progesterone along with its associated metabolic machinery (involving intense 5α-reductase and sulfation transformations42,43), the female body slowly returns to basal levels of progesterone, now primarily produced by the adrenals and later by the corpus lutea, following corresponding metabolic pathways. In relation to that, our results show how pregnanediol glucuronide (PD-5β3α20α-3G), originated from 5β-reduced pregnanolone (P), becomes a relevant metabolite of progesterone in mature BM. This finding highlights that the phase II metabolism (conjugation with sulfate and glucuronide) is tightly linked to progesterone metabolic pathway specified by the origin of its synthesis.
In contrast to progesterone metabolites, the proportion of pregnenolone metabolites in the BM steroidal pool does not change throughout lactation. Our study demonstrates that pregnenolone sulfate (5P-S), a key component of placental steroidogenic machinery42 and the sulfate-conjugated steroid with very interesting biological activities,14 is gradually replaced from its dominant position in colostrum by its advanced bis-sulfated metabolite (5PD-3β20α-SS) in mature BM samples. This bis-conjugate is a well-known and apparently inactive urinary44 and plasmatic45 metabolite of the hormone pregnenolone originating in the adrenal gland. These changes reflect a shift in the site of synthesis and primary metabolism of pregnenolone.
Androgen metabolites, according to our data, are principal steroid metabolites in mature BM, and are chiefly represented by 4- and 5-androstene sulfated conjugates. According to our data, the dehydroepiandrosterone sulfate (DHEA-S), a core androgen, is the principal steroid metabolite in BM at all stages of lactation. Its concentrations vary widely from a few to a several hundred nmol L−1 ranges, especially within colostrum samples, illustrating high interindividual variability in accordance with earlier observed data.20 The disappearance of 16-hydroxylated DHEA metabolites alongside an increase in 5-androstanediol metabolites in BM during later stages of lactation suggests that the former were of fetal-placental origin, while the latter originate from maternal adrenals. In relation to 4-androstene metabolites, our data show that androstanediol (AD-5α3α17β-3S), androsterone sulfate (AN-S), etiocholanolone (Etio-S) and epiandrosterone (epiAN-S) sulfates are main testosterone metabolites in BM. These results contrast with the ones reported by Sahlberg and Axelson,20 which reported androstanediol bis-sulfate (AD-5α3α17β-SS) as the major metabolite in BM. The prior use of hydrolysis for steroid determination in their study may explain these discrepancies.
Our study reveals that conjugated metabolites of estrogens are the minor steroids, but they are present in BM in all stages of lactation. As it was stated in recent review,33 several studies reported on very low estrogen concentrations detected in human breast milk, among them only a few had considered the impact of estrogen conjugated metabolites.20,21 The methodological differences in sampling and in indirect analytical technics altogether provided inconsistent results regarding the presence and prevalence of conjugated estrogens in BM through lactation.33 According to our data, estrone sulfate (E1-S) is the most abundant estrogen in BM, regardless of lactation stage. In comparison to previous studies,20,21 we report broader concentration range (over 0.1–100 nmol L−1) and higher mean concentrations (tens of nmol L−1) in colostrum samples. Although all estrogen metabolites detected in colostrum are clearly pregnancy-related—specifically, placentally derived estrone and estradiol, as well as fetal-derived estriol metabolites42,43—their origins in mature BM remain uncertain, particularly for estriol metabolites.
The cortisone metabolite tetrahydrocortisone glucuronide (THE-3G) is the principal corticosteroid metabolite detected in all BM samples. Detection of other conjugated metabolites was limited to only six compounds due to the absence of corresponding standards. Therefore, a comprehensive analysis of this metabolite fraction in BM was not possible. Whether THE-3G, along with other detected cortisone and cortisol conjugates, reflects cortisol–cortisone turnover in BM throughout lactation remains an open question, one that could be explored by analysing free, unconjugated steroids in BM.
In summary, our findings are in line with an earlier published indirect profiling of BM steroids in early lactation20 regarding the distribution for steroids from various families and with different types of conjugation. Our data support the idea that steroid metabolite levels in milk reflect the average hormone content in maternal plasma, as previously reported for progesterone, estrogen46 or glucocorticoids.30,47 This resemblance, along with correlations observed between specific conjugated steroid levels20,21 suggests that conjugated steroids are transported either by direct transfer to BM from maternal serum (non-polar hormones) or by active transport (polar conjugates), as it was suggested for other appearing in BM hormones.46 Nonetheless, it should be mentioned that breast tissue by itself can exhibit metabolic activity towards steroids, therefore impacting the BM steroid profile. As an example, cortisone is much more abundant than cortisol in BM compared to serum, most probably due to the 11β-HSD2 activity in breast tissue, which by converting cortisol to cortisone protects the baby from elevated cortisol levels in BM.12 In this context, the levels of BM steroids are of interest in relation to new-born health and wellbeing.12 Therefore, it would be of great interest to compare the relative distribution of conjugated metabolites in both plasma and BM in order to establish the role of the steroid metabolic machinery of the breast tissue.
BM steroid phase II metabolites in relation to infant wellbeing
The present study confirmed that infants are exposed to a wide variety of steroids right after birth via consumption of BM. According to our findings, consumption of colostrum is characterized by the highest postnatal ingestion of steroid metabolites. In line with this, elevated excretion of some steroid metabolites detected in infant urine during the first week of life has been directly linked to BM consumption.48The exact role of BM steroids remains unclear—whether they function as a means of transmitting regulatory signals from mother to child or play a crucial physiological role in postnatal development and health.49 However, studies suggest that some BM-transmitted steroids, including cortisol, have physiological relevance for breastfed infants.5,29,31,33,50,51 Our results indicate the prevalence of conjugated steroids in BM. For a long time, conjugated steroids were considered to be inactive metabolites. However, recent studies have revealed that sulfation plays an important regulatory role in steroid metabolism, where conjugated metabolites can either serve as reservoirs for the active form of steroids—allowing for their later activation, particularly in tissues rich in steroid sulfatase (e.g., placenta, blood, brain, skin, and breast tissue)—or exert distinct biological activities themselves.15,16 In this context, our results showed that mono-sulfate metabolites are among the most detected steroids in colostrum. Therefore, BM may be considered as a substantial source of steroids with interesting molecular and physiological activities which could be of special interest in relation to newborn development and health.13,15,16,52 Throughout lactation, mono-sulfates decrease relative to bis-sulfates and glucuronides. Both conjugations, being inactivating metabolic transformations toward the parent molecule, may have a prominent role in controlling the levels and, thus, the biological activity of unconjugated steroids.16,53,54 Glucuronide concentrations in BM may serve as indicators of steroid metabolic turnover or parent steroid levels, which are often challenging to assess due to methodological limitations or difficulty in accessing the target of evaluation,55 and be used to anticipate its biological relevance. Numerous studies have emphasized the health relevance of glucuronylated steroids, particularly their potential for reactivation via deconjugation by intestinal microbiota.56,57 However, whether this process occurs in the newborn intestine remains unknown, as research on microbial colonization in neonatal intestines is still evolving.58,59
Strengths and limitations
The main strengths of our study are (i) the direct measurement of conjugates in BM, (ii) the large number of metabolites including non-commonly detected bis-sulfates and sulfate–glucuronide conjugates and (iii) the wide coverage of the cohort including samples from the first days to over one-month of the lactation period, allowing the evaluation of colostrum, transitional and mature BM.Our study has also several limitations. First, we did not analyse longitudinal intraindividual samples of BM collected from the same breastfeeding mother through period of lactation. Analysis of such samples would provide valuable information on intraindividual variability in steroid BM composition. Second, the total amount of BM produced in a 24 h period was not monitored. In absence of this “gold standard” normalization60 we normalized our results based on the average production at the day of sampling. Due to practical difficulties in obtaining this gold-standard,61 the application of this approximation is common, with and some studies of steroid BM concentrations even reported without normalization.20,21 Third, despite determining a large number of steroid metabolites, the panel of conjugates was selected on the basis of available standards (either commercially available conjugates or available starting materials for synthesis). Additional steroid conjugates may be detected by increasing the panel as soon as appropriate standards become accessible. Among them, it is remarkable the absence of more corticosteroids especially considering that the metabolome of corticosteroids is very extensive.62 Finally, our study may be complemented by the analysis of unconjugated forms of steroids in BM. For some families like the corticosteroids a relatively high proportion of unconjugated steroids in BM has been reported.63 The simultaneous determination of both conjugated and free forms of steroids could be of special interest for studies investigating the biological significance of steroid conjugation in respect to mother and baby wellbeing and health.12,64
Conclusions
In summary, this study provides direct evidence of the secretion and compositional changes of steroid conjugates in human BM throughout lactation. Our findings demonstrate that many pregnancy-related and biologically significant phase II conjugated steroids are secreted in BM, with their highest prevalence observed in colostrum, followed by a notable decline as lactation progresses.The observed secretion dynamics and metabolite profile shifts align with the maternal physiological adaptations occurring in the postpartum period. The confirmation that BM serves as a source of conjugated steroids for new-borns opens new research avenues for exploring the physiological significance of these metabolites in maternal and infant health.
Author contributions
Conceptualization: OK, OP and OV. Data curation: OK, AA and GF. Formal analysis: OK and OP. Funding acquisition: OP and OV. Investigation: OK, AA, GF, OV, MML and OP. Methodology: OK and OP. Project administration: OK, AA, GF, OV and OP. Supervision: OP. Validation and visualization: OK. Writing original draft: OK and OP. Writing and editing: OK, AA, GF, OV, MML and OP. All authors read and approved the submitted version.Data availability
The data supporting this article have been included as part of the ESI.†Conflicts of interest
There are no conflicts to declare.Acknowledgements
We are very grateful to all volunteers participating in the study and whose breast milk samples were used for the present publication. This work has received funding from the European Research Council under the European Union's Horizon 2020 research and innovation programme (Grant Agreement No. 883069). Strategic Plan for Research and Innovation in Health (PERIS) of the Catalan Government is acknowledged for the OK contract (SLT002/16/00007). We are thankful to Cristina Montero Domínguez (from Servicio de Bioquímica Clínica, Hospital General Universitario Gregorio Marañón, Madrid) for valuable technician support during her training stage in our Applied Metabolomics Research Group.References
- N. J. Andreas, B. Kampmann and K. Mehring Le-Doare, Human breast milk: A review on its composition and bioactivity, Early Hum. Dev., 2015, 91, 629–635 CrossRef CAS PubMed.
- D. K. Dror and L. H. Allen, Overview of Nutrients in Human Milk, Adv. Nutr., 2018, 9, 278S–294S CrossRef PubMed.
- C. E. Grosvenor, M. F. Picciano and C. R. Baumrucker, Hormones and growth factors in milk, Endocr. Rev., 1993, 14, 710–728 CrossRef CAS PubMed.
- O. Ballard and A. L. Morrow, Human Milk Composition: Nutrients and Bioactive Factors, Pediatr. Clin. North Am., 2013, 60, 49–74 CrossRef PubMed.
- S. Eisha, I. Joarder, S. Wijenayake and P. O. McGowan, Non-nutritive bioactive components in maternal milk and offspring development: a scoping review, J. Dev. Origins Health Dis., 2022, 13, 665–673 CrossRef CAS PubMed.
- M. Hamosh, Bioactive factors in human milk, Pediatr. Clin. North Am., 2001, 48, 69–86 CrossRef CAS PubMed.
- P. Christian, E. R. Smith, S. E. Lee, A. J. Vargas, A. A. Bremer and D. J. Raiten, The need to study human milk as a biological system, Am. J. Clin. Nutr., 2021, 113, 1063–1072 CrossRef CAS PubMed.
- S. S. Betrabet, Z. Shikary, S. Patel, V. S. Toddywalla, D. N. Patel and B. N. Saxena, Endogenous steroid in breast milk and serum of lactating women from parturition to one year postpartum, J. Obstet. Gynecol. India, 1993, 641–644 Search PubMed.
- J. F. Murphy, I. Hughes, E. R. Verrier Jones, S. Gaskell and A. W. Pike, Pregnanediols and breast milk jaundice, Arch. Dis. Child., 1981, 56, 474–476 CrossRef CAS PubMed.
- S. Pundir, C. R. Wall, C. J. Mitchell, E. B. Thorstensen, C. T. Lai, D. T. Geddes and D. Cameron-Smith, Variation of Human Milk Glucocorticoids over 24-hour Period, J. Mammary Gland Biol. Neoplasia, 2017, 22, 85–92 CrossRef PubMed.
- M. Lu, H. Xiao, K. Li, J. Jiang, K. Wu and D. Li, Concentrations of estrogen and progesterone in breast milk and their relationship with the mother's diet, Food Funct., 2017, 8, 3306–3310 RSC.
- M. Muelbert, T. Alexander, M. H. Vickers, J. E. Harding, L. Galante and F. H. Bloomfield, DIAMOND study group, Glucocorticoids in preterm human milk, Front. Nutr., 2022, 27, 965654 CrossRef PubMed.
- M. Park-Chung, A. Malayev, R. H. Purdy, T. T. Gibbs and D. H. Farb, Sulfated and unsulfated steroids modulate gamma-aminobutyric acidA receptor function through distinct sites, Brain Res., 1999, 29, 72–87 CrossRef PubMed.
- C. Harteneck, Pregnenolone sulfate: from steroid metabolite to TRP channel ligand, Molecules, 2013, 27, 12012–12028 CrossRef PubMed.
- J. W. Mueller, L. C. Gilligan, J. Idkowiak, W. Arlt and P. A. Foster, The Regulation of Steroid Action by Sulfation and Desulfation, Endocr. Rev., 2015, 36, 526–563 CrossRef CAS PubMed.
- J. Vítků and R. Hampl, Steroid Conjugates and Their Physiological Role, Physiol. Res., 2023, 72, S317–S322 Search PubMed.
- J. Roos, S. Johnson, S. Weddell, E. Godehardt, J. Schiffner, G. Freundl and C. Gnoth, Monitoring the menstrual cycle: Comparison of urinary and serum reproductive hormones referenced to true ovulation, Eur. J. Contracept. Reprod. Health Care, 2015, 20, 438–450 CrossRef CAS PubMed.
- L. F. Blackwell, D. G. Cooke and S. Brown, The Use of Estrone-3-Glucuronide and Pregnanediol-3-Glucuronide Excretion Rates to Navigate the Continuum of Ovarian Activity, Front. Public Health, 2018, 31, 153 CrossRef PubMed.
- L. Meng, S. Su, L. Li, S. Liu, Y. Li, Y. Liu, Y. Lu, Z. Xu, L. Liu, Q. He, Y. Zheng, X. Liu, Y. Cong, Y. Zhai, Z. Zhao and Z. Cao, Delivery prediction by quantitative analysis of four steroid metabolites with liquid chromatography tandem mass spectrometry in asymptomatic pregnant women, Ann. Med., 2022, 54, 1150–1159 CrossRef CAS PubMed.
- B. L. Sahlberg and M. Axelson, Identification and quantitation of free and conjugated steroids in milk from lactating women, J. Steroid Biochem., 1986, 25, 379–391 CrossRef CAS PubMed.
- H. H. McGarrigle and G. C. Lachelin, Oestrone, oestradiol and oestriol glucosiduronates and sulfates in human puerperal plasma and milk, J. Steroid Biochem., 1983, 18, 607–611 CrossRef CAS PubMed.
- E. Olesti, J. Boccard, G. Visconti, V. González-Ruiz and S. Rudaz, From a single steroid to the steroidome: Trends and analytical challenges, J. Steroid Biochem. Mol. Biol., 2021, 206, 105797 CrossRef CAS PubMed.
- M. Hill, A. Pašková, R. Kančeva, M. Velíková, J. Kubátová, L. Kancheva, K. Adamcová, M. Mikešová, Z. Žižka, M. Koucký, H. Šarapatková, V. Kačer, P. Matucha, M. Meloun and A. Pařízek, Steroid profiling in pregnancy: a focus on the human fetus, J. Steroid Biochem. Mol. Biol., 2014, 139, 201–222 CrossRef CAS PubMed.
- R. L. Gomes, W. Meredith, C. E. Snape and M. A. Sephton, Conjugated steroids: analytical approaches and applications, Anal. Bioanal. Chem., 2009, 393, 453–458 CrossRef CAS PubMed.
- M. D. McLeod, C. C. Waller, A. Esquivel, G. Balcells, R. Ventura, J. Segura and O. J. Pozo, Constant Ion Loss Method for the Untargeted Detection of Bis-sulfate Metabolites, Anal. Chem., 2017, 89, 1602–1609 CrossRef CAS PubMed.
- M. C. Neville, R. Keller, J. Seacat, V. Lutes, M. Neifert, C. Casey, J. Allen and P. Archer, Studies in human lactation: milk volumes in lactating women during the onset of lactation and full lactation, Am. J. Clin. Nutr., 1988, 48, 1375–1386 CrossRef CAS PubMed.
- M. Mauri, T. Elli, G. Caviglia, G. Uboldi and M. Azzi, RAWGraphs: A Visualisation Platform to Create Open Outputs, in Proceedings of the 12th Biannual Conference on Italian SIGCHI Chapter, New York, NY, USA: ACM, 2017, pp. 28:1–28:5.
- L. M. Glynn, E. P. Davis, C. D. Schetter, A. Chicz-Demet, C. J. Hobel and C. A. Sandman, Postnatal maternal cortisol levels predict temperament in healthy breastfed infants, Early Hum. Dev., 2007, 83, 675–681 CrossRef CAS PubMed.
- K. R. Grey, E. P. Davis, C. A. Sandman and L. M. Glynn, Human milk cortisol is associated with infant temperament, Psychoneuroendocrinology., 2013, 38, 1178–1185 CrossRef CAS PubMed.
- J. J. Hollanders, A. C. Heijboer, B. van der Voorn, J. Rotteveel and M. J. J. Finken, Nutritional programming by glucocorticoids in breast milk: Targets, mechanisms and possible implications, Best Pract. Res., Clin. Endocrinol. Metab., 2017, 31, 397–408 CrossRef CAS PubMed.
- S. Nolvi, H. M. Uusitupa, D. J. Bridgett, H. Pesonen, A. K. Aatsinki, E. L. Kataja, R. Korja, H. Karlsson and L. Karlsson, Human milk cortisol concentration predicts experimentally induced infant fear reactivity: moderation by infant sex, Dev. Sci., 2018, 21, e12625 CrossRef PubMed.
- L. Petrullo, K. Hinde and A. Lu, Steroid hormone concentrations in milk predict sex-specific offspring growth in a nonhuman primate, Am. J. Hum. Biol., 2019, 31, e23315 CrossRef PubMed.
- X. Jin, S. L. Perrella, C. T. Lai, N. L. Taylor and D. T. Geddes, Oestrogens and progesterone in human milk and their effects on infant health outcomes: A narrative review, Food Chem., 2023, 30, 136375 CrossRef PubMed.
- C. Shackleton, O. J. Pozo and J. Marcos, GC/MS in Recent Years Has Defined the Normal and Clinically Disordered Steroidome: Will It Soon Be Surpassed by LC/Tandem MS in This Role?, J. Endocr. Soc., 2018, 2, 974–996 CrossRef CAS PubMed.
- H. L. J. Makin, J. W. Honour, C. H. L. Shackleton and W. J. Griffiths, General Methods for the Extraction, Purification, and Measurement of Steroids by Chromatography and Mass Spectrometry, in Steroid Analysis, ed. H. Makin and D. Gower, Springer, Dordrecht, 2010 Search PubMed.
- M. Sas, S. Viska and J. Gellen, Der Steroidgehalt der Frauenmilch, Arch. Gynecol., 1969, 207, 452–459 CAS.
- S. T. Wolford and C. J. Argoudelis, Measurement of estrogen in cow's milk, human milk, and dietary products, J. Dairy Sci., 1979, 62, 1458–1463 CrossRef CAS PubMed.
- L. Xu, L. Zhang, Y. Zhang, Q. Sheng and A. Zhao, Qualitative and quantitative comparison of hormone contents between bovine and human colostrums, Int. Dairy J., 2011, 21, 54–57 CrossRef CAS.
- P. A. Foster and J. W. Mueller, SULFATION PATHWAYS: Insights into steroid sulfation and desulfation pathways, J. Mol. Endocrinol., 2018, 61, T271–T283 CAS.
- T. A. Lightning, T. F. Gesteira and J. W. Mueller, Steroid disulfates – Sulfation double trouble, Mol. Cell. Endocrinol., 2021, 524, 111–161 CrossRef PubMed.
- A. Alex, E. Bhandary and K. P. McGuire, Anatomy and Physiology of the Breast during Pregnancy and Lactation in Diseases of the Breast during Pregnancy and Lactation, in Adv Exp Med Biol., ed. S. Alipour and R. Omranipour, Springer Nature, Switzerland AG, 2020, pp. 1252 Search PubMed.
- J. R. Pasqualini, Enzymes involved in the formation and transformation of steroid hormones in the fetal and placental compartments, J. Steroid Biochem. Mol. Biol., 2005, 97, 401–415 CrossRef CAS PubMed.
- J. R. Pasqualini and G. S. Chetrite, The formation and transformation of hormones in maternal, placental and fetal compartments: biological implications, Horm. Mol. Biol. Clin. Invest., 2016, 27, 11–28 CAS.
- O. J. Pozo, J. Marcos, O. Khymenets, A. Pranata, C. C. Fitzgerald, M. D. McLeod and C. Shackleton, SULFATION PATHWAYS: Alternate steroid sulfation pathways targeted by LC-MS/MS analysis of disulfates: application to prenatal diagnosis of steroid synthesis disorders, J. Mol. Endocrinol., 2018, 61, M1–M12 CAS.
- J. Marín-Corral, J. Rodríguez-Morató, A. Gomez-Gomez, S. Pascual-Guardia, R. Muñoz-Bermúdez, A. Salazar-Degracia, P. Pérez-Terán, M. I. Restrepo, O. Khymenets, N. Haro, J. R. Masclans and O. J. Pozo, Metabolic Signatures Associated with Severity in Hospitalized COVID-19 Patients, Int. J. Mol. Sci., 2021, 22, 4794 CrossRef PubMed.
- D. Schams and H. Karg, Hormones in milk, Ann. N. Y. Acad. Sci., 1986, 464, 75–86 CrossRef CAS PubMed.
- B. van der Voorn, M. de Waard, J. B. van Goudoever, J. Rotteveel, A. C. Heijboer and M. J. Finken, Breast-Milk Cortisol and Cortisone Concentrations Follow the Diurnal Rhythm of Maternal Hypothalamus-Pituitary-Adrenal Axis Activity, J. Nutr., 2016, 146, 2174–2179 CrossRef CAS PubMed.
- B. Ferris, Pregnanediol Excretion by Newly Born Infants, Arch. Pediatr. Adolesc. Med., 1968, 115, 693 CrossRef CAS PubMed.
- M. Peaker and M. C. Neville, Hormones in milk: chemical signals to the offspring?, J. Endocrinol., 1991, 131, 1–3 CAS.
- K. M. Bernt and W. A. Walker, Human milk as a carrier of biochemical messages, Acta Paediatr., Suppl., 1999, 88, 27–41 CrossRef CAS PubMed.
- J. D. Piñeiro-Ramos, A. Parra-Llorca, I. Ten-Doménech, M. Gormaz, A. Ramón-Beltrán, M. Cernada, G. Quintás, M. C. Collado, J. Kuligowski and M. Vento, Effect of donor human milk on host-gut microbiota and metabolic interactions in preterm infants, Clin. Nutr., 2021, 40, 1296–1309 CrossRef PubMed.
- L. Stárka, M. Dušková and M. Hill, Dehydroepiandrosterone: a neuroactive steroid, J. Steroid Biochem. Mol. Biol., 2015, 145, 254–260 CrossRef PubMed.
- P. Wang, Y. Jia, R. Wu, Z. Chen and R. Yan, Human gut bacterial β-glucuronidase inhibition: An emerging approach to manage medication therapy, Biochem. Pharmacol., 2021, 190, 114566 CrossRef CAS PubMed.
- E. Kalogera, C. Pistos, X. Provatopoulou, S. Athanaselis, C. Spiliopoulou and A. Gounaris, Androgen glucuronides analysis by liquid chromatography tandem-mass spectrometry: could it raise new perspectives in the diagnostic field of hormone-dependent malignancies?, J. Chromatogr. B:Anal. Technol. Biomed. Life Sci., 2013, 940, 24–34 CrossRef CAS PubMed.
- B. Trabert, K. A. Michels, G. L. Anderson, L. A. Brinton, R. T. Falk, A. M. Geczik, H. R. Harris, K. Pan, R. M. Pfeiffer, L. Qi, T. Rohan, N. Wentzensen and X. Xu, Circulating androgens and postmenopausal ovarian cancer risk in the Women's Health Initiative Observational Study, Int. J. Cancer, 2019, 145, 2051–2060 CrossRef CAS PubMed.
- H. Colldén, A. Landin, V. Wallenius, E. Elebring, L. Fändriks, M. E. Nilsson, H. Ryberg, M. Poutanen, K. Sjögren, L. Vandenput and C. Ohlsson, The gut microbiota is a major regulator of androgen metabolism in intestinal contents, Am. J. Physiol.: Endocrinol. Metab., 2019, 317, E1182–E1192 CrossRef PubMed.
- S. M. Ervin, H. Li, L. Lim, L. R. Roberts, X. Liang, S. Mani and M. R. Redinbo, Gut microbial β-glucuronidases reactivate estrogens as components of the estrobolome that reactivate estrogens, J. Biol. Chem., 2019, 294, 18586–18599 CrossRef CAS PubMed.
- A. Singh and M. Mittal, Neonatal microbiome – a brief review, J. Matern.-Fetal Neonat. Med., 2020, 33, 3841–3848 CrossRef PubMed.
- R. Cheddadi, V. Yeramilli and C. Martin, From Mother to Infant, from Placenta to Gut: Understanding Varied Microbiome Profiles in Neonates, Metabolites, 2023, 13, 1184 CrossRef CAS PubMed.
- A. E. Mohr, K. E. Senkus, J. M. McDermid, P. K. Berger, M. T. Perrin and D. Handu, Human Milk Nutrient Composition Data is Critically Lacking in the United States and Canada: Results from a Systematic Scoping Review of 2017–2022, Adv. Nutr., 2023, 14, 1617–1632 CrossRef PubMed.
- L. Galante, M. H. Vickers, A. M. Milan, C. M. Reynolds, T. Alexander, F. H. Bloomfield and S. Pundir, Feasibility of Standardized Human Milk Collection in Neonatal Care Units, Sci. Rep., 2019, 9, 14343 CrossRef PubMed.
- L. Schiffer, L. Barnard, E. S. Baranowski, L. C. Gilligan, A. E. Taylor, W. Arlt, C. H. L. Shackleton and K. H. Storbeck, Human steroid biosynthesis, metabolism and excretion are differentially reflected by serum and urine steroid metabolomes: A comprehensive review, J. Steroid Biochem. Mol. Biol., 2019, 194, 105439 CrossRef CAS PubMed.
- A. Gomez-Gomez, J. Miranda, G. Feixas, A. Arranz Betegon, F. Crispi, E. Gratacós and O. J. Pozo, Determination of the steroid profile in alternative matrices by liquid chromatography tandem mass spectrometry, J. Steroid Biochem. Mol. Biol., 2020, 197, 105520 CrossRef CAS PubMed.
- S. Wijenayake, J. Martz, H. E. Lapp, J. A. Storm, F. A. Champagne and A. C. Kentner, The contributions of parental lactation on offspring development: It's not udder nonsense!, Horm. Behav., 2023, 153, 105375 CrossRef PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4fo05713a |
| This journal is © The Royal Society of Chemistry 2025 |