
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Assessing the impact of food-derived bioactives on digestive proteases by in vitro and in silico approaches
Sara Margherita
Borgonovi†
a,
Florinda
Perugino†
 bc,
Luca
Dellafiora
b,
Francesca
Annunziata
a,
Lorenzo
Pedroni
b,
Gianni
Galaverna
b,
Andrea
Pinto
a,
Sabrina
Dallavalle
a,
Stefania
Iametti
a and
Mattia
Di Nunzio
*a
bc,
Luca
Dellafiora
b,
Francesca
Annunziata
a,
Lorenzo
Pedroni
b,
Gianni
Galaverna
b,
Andrea
Pinto
a,
Sabrina
Dallavalle
a,
Stefania
Iametti
a and
Mattia
Di Nunzio
*a
aDepartment of Food, Environmental and Nutritional Sciences (DeFENS), University of Milan, via Celoria 2, 20133 Milan, Italy. E-mail: mattia.dinunzio@unimi.it
bDepartment of Food and Drugs, University of Parma, Parco Area delle Scienze 27/A, 43124 Parma, Italy
cDepartment of Biology, University of Naples Federico II, via Cinthia, 80126 Naples, Italy
First published on 20th March 2025
Abstract
The interactions between food components and digestive tract enzymes can affect nutrient absorption and impact an individual's health. Certain components, particularly polyphenols, are reported to inhibit digestive enzymes and are commonly referred to as anti-nutritional factors. Reports on this subject often contradict each other, highlighting the need for consistent methodologies to assess the potential impact of bioactive compounds. This study evaluated the “in vitro” activity of pepsin, trypsin, and chymotrypsin using ovalbumin, gluten, and haemoglobin as substrates in the presence or absence of twenty-five bioactive natural compounds belonging to different chemical classes at gastro-intestinal physiological concentrations (0.1 mM). The results indicate that bioactives may have opposite effects on proteolytic activity depending on the substrate/enzyme combination and bioactives structure. With ovalbumin as substrate, piceid and resveratrol were described as strong chymotrypsin activators (+1.46- and 1.17-fold change, respectively), phloridzin dihydrate as a weaker activator (+0.41-fold change), while phloretin was a strong inhibitor (−0.65-fold change). A computational approach based on molecular docking and dynamics simulations was used to investigate the interactions between selected bioactives, chymotrypsin and ovalbumin. The “in silico” study included piceid and phloridzin dihydrate, as well as their respective aglycones (resveratrol and phloretin). The results obtained through computational modelling indicate that all four bioactives can interact with chymotrypsin. However, only those bioactives that enhance in vitro proteolytic activity induce a partial unfolding of ovalbumin's structure. This suggests that the effect of bioactive compounds on protein digestion may be substrate-dependent, and may vary depending on the specific protein being digested.
1. Introduction
Natural antioxidants, such as polyphenols and terpenoids, are recognized as key determinants of the health advantages associated with a diet rich in fruits and vegetables.1,2 However, the beneficial effects of polyphenol consumption on human health have been associated with a negative impact on the breakdown of macromolecules during digestion.3 Indeed, due to the generally low absorption of this family of bioactive compounds, most of them remain in the gastrointestinal tract, ranging from 0.4 to 3 mM,4 with reported inhibitory effects on enzymes involved in the breakdown of macromolecular nutrients, such as starch and oligo- or polysaccharides.5 In particular, the inhibitory activity of a polyphenol against α-amylase is determined by molecular binding interactions such as hydrogen bonding and hydrophobic interactions. The effective inhibition of the enzyme is dependent on the presence of specific characteristics, including hydroxyl groups, galloyl substituents and conjugated systems, which are inherent to bioactive compounds.6The inhibitory effects of phenolics on the digestion of saccharides and lipids may be considered one of the mechanisms, together with the modulation of neurohormones/peptides involved in food intake,6 in the prevention of obesity and other metabolic diseases.7 However, inhibitory effects on the digestion of proteins are not desirable, as it has been also demonstrated that fermentation of non-hydrolysed proteins by colonic flora increases the risk of colon cancer.8 Furthermore, several reports have indicated the formation of complexes between proteins and polyphenols in food, which may impair the enzymatic activities associated with the involved proteins and affect digestibility.9,10
The picture is complicated also by conflicting reports on the effect of polyphenols on the activity of proteolytic enzymes in the digestive tract. For instance, He et al.11 reported an inhibitory effect of green tea polyphenols on pepsin activity using haemoglobin as a substrate, whereas Donmez et al.12 reported an increase of pepsin activity on β-lactoglobulin in the presence of epigallocatechin gallate. Aside from methodological issues related to the choice of a limited set of proteases and substrates, most of the available studies were limited to a quite narrow set of polyphenols.
To address at least some of these limitations, this study undertook a comprehensive investigation into the potential influence of phenolics on protein breakdown. Standardized approaches have been used to evaluate the impact of 25 bioactives belonging to different chemical classes (stilbenoids, dihydrochalcones, phenolic alcohols, phenolic acids, flavonoids, anthraquinone, and terpenoids) on the activity of key digestive proteolytic enzymes (pepsin, trypsin, and chymotrypsin). Stilbenoids are widely present in grapes, red wine and various berries,13 phenolic alcohols in olive oil,14 phenolic acids in red berries and citrus fruits,15 while flavonoids represent the main class in green tea, and legumes.16 Furthermore, the respective glycosylated forms were included to assess the effect of the glycoside in comparison to the aglycone alone. Each enzyme was tested on three different protein substrates (bovine haemoglobin, chicken egg albumin, and wheat gluten). Haemoglobin was selected as reference protein as one of the most common substrates for the assessment of pepsin activity,11,17–19 which inclusion may facilitate comparison of the results obtained in this study. Ovalbumin and gluten were tested as they are commonly used in real food or meals and can be considered as good food models for two extreme cases: (i) solutions of native globular proteins20 and (ii) low digestible21 supramolecular assemblies of water-insoluble intrinsically disordered proteins,22 respectively. The study also included representative species of seldom studied glycosidic derivatives of representative phenolics,23,24 as well as terpenoids and anthraquinones.25,26
To rationalise the results obtained from enzyme activity measurements, the structure/function relationship in selected polyphenol/enzyme/substrate combinations was addressed through the performance of molecular docking studies and molecular dynamics simulations. In silico analyses considered the most blatant effects observed for specific bioactive/enzyme/substrate combinations, regardless of whether the added bioactive enhanced or inhibited the proteolytic activity.
The final aim of this study is to define some of the molecular basis of the structure–activity relationship (SAR) in these systems, pointing out structural features (3D structure, substitution pattern, binding affinity) relevant to specific effects. This could represent a significant advancement in our comprehension of the digestive-modulating properties of food bioactives, with potential implications for the rational design and development of functional foods and food supplements.
2. Materials and methods
2.1 Materials
Bovine haemoglobin (H2625), chicken ovalbumin (A5503), wheat gluten (G5004), porcine pepsin (P6887; ≥3200 units per mg protein), bovine trypsin (T1426; ≥10![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 BAEE units per mg protein) and α-chymotrypsin (C4129; ≥40 units per mg protein), piceide (15721), phloretin (PHL82624), phloridzin dihydrate (274313), gallic acid (842649), ferulic acid (128708), caffeic acid (8.22029), oleuropein (12247), (+)-catechin hydrate (22110), procyanidin B1 (PHL89764), naringenin (N5893), quercetin (PHR1488), rutin (PHL89270) were from Sigma-Aldrich (St. Louis, MO, USA). Resveratrol (BD4672) and epigallocatechin gallate (BD42886) were from BLD Pharm (Reinbek, Germany). Pterostilbene (F093412) and tyrosol (F077935) were from Fluorochem (Hadfield, United Kingdom). Quercetin 3-β-O-glucoside (482-35-9) and 4-hydroxy benzyl alcohol (H20806) were from HWI Group (Appenweier, Germany) and Alfa Aesar (Haverhill, MA, USA), respectively.
000 BAEE units per mg protein) and α-chymotrypsin (C4129; ≥40 units per mg protein), piceide (15721), phloretin (PHL82624), phloridzin dihydrate (274313), gallic acid (842649), ferulic acid (128708), caffeic acid (8.22029), oleuropein (12247), (+)-catechin hydrate (22110), procyanidin B1 (PHL89764), naringenin (N5893), quercetin (PHR1488), rutin (PHL89270) were from Sigma-Aldrich (St. Louis, MO, USA). Resveratrol (BD4672) and epigallocatechin gallate (BD42886) were from BLD Pharm (Reinbek, Germany). Pterostilbene (F093412) and tyrosol (F077935) were from Fluorochem (Hadfield, United Kingdom). Quercetin 3-β-O-glucoside (482-35-9) and 4-hydroxy benzyl alcohol (H20806) were from HWI Group (Appenweier, Germany) and Alfa Aesar (Haverhill, MA, USA), respectively.
2.2 Bioactives synthesis
δ-Viniferin, ε-viniferin, and a pterostilbene dimer were prepared according to Mattio et al.27 Aloe-emodin was obtained through the oxidation of aloin as described by Dallavalle et al.28 Safranal was prepared from picrocrocin (from saffron residues) as reported by Catinella et al.29 Naringin and limonin were extracted from Citrus paradisi seeds as reported by Magurano et al.30 Stock 10 mM solutions of each compound were prepared in ethanol. Fig. 1 reports the chemical structure of the selected bioactive compounds included in the study. | ||
| Fig. 1 Chemical structure of selected bioactive compounds. Resveratrol (A), pterostilbene (B), δ-viniferin (C), ε-viniferin (D), pterostilbene dimer (E), piceide (F), phloretin (G), phloridzin dihydrate (H), 4-hydroxybenzyl alchol (I), tyrosol (J), gallic acid (K), ferulic acid (L), caffeic acid (M), oleuropein (N), (+)-catechin hydrate (O), procyanidin B1 (P), epigallocatechin gallate (Q), naringenin (R), quercetin (S), naringin (T), quercetin 3-β-O-glucoside (U), rutin (V), aloe-emodin (W), safranal (X), limonin (Y). | ||
2.3 Pepsin, trypsin, and chymotrypsin activity
Pepsin, trypsin, and chymotrypsin activities were determined according to Borgonovi et al., with slight modification.31 A very small volume (0.006 ml) of a stock 10 mM ethanolic solution of the investigated compound (or of ethanol alone, in control runs) was added to 0.494 ml of 3% solutions of haemoglobin, gluten, or ovalbumin, in water at pH 2 (for pepsin), or at pH 7 (for trypsin and chymotrypsin). The pH value was adjusted – when required – with 200 mM HCl. The reaction was started by adding 0.1 ml of a solution of the appropriate enzyme (30 mg L−1 in 0.15 M NaCl, 0.0115 M CaCl2, adjusted to pH 2 for pepsin, and pH 7 for trypsin and chymotrypsin). The final bioactives, substrates and enzymes concentration were 0.1 mM, 2.47%, and 5 mg L−1, respectively. After 10, 15, 20, and 30 min, the reaction was stopped by adding 1 mL of 20% (w/v) trichloroacetic acid (TCA). After centrifugation (12000g for 10 min), TCA-soluble peptides in the supernatant were detected at 280 nm. For this study, one proteolytic unit corresponds – regardless of the enzyme being used – to a 0.001 increase in A280 per minute. Activity was normalized for enzymes concentration and expressed as U mg−1 enzyme. The effect of a given bioactive was calculated as the change (positive or negative) with respect to control. The final ethanol concentration was 1%, which did not result in a significant alteration to enzyme activity in comparison to the absence of ethanol.
2.4 Proteins and polyphenols model design
A complete, good-resolution (2.00 Å) structure of bovine chymotrypsin was used (PDB code 1ACB; https://www.rcsb.org; last accessed September 2023).32 The UCSF Chimera software (version 1.16)33 was used to process further the structure removing water and co-crystallized ligand, and adding hydrogens before running further analysis. The 3D structure of ovalbumin was derived from the AlphaFold (https://alphafold.ebi.ac.uk/) predicted structure retrieved in .pdb format from Uniprot (code: P01012), since the crystallographic structure in the RCSB database is not complete. AlphaFold is a deep learning-based system commonly used for the prediction/analysis of the structure of proteins.34 The AlphaFold structure was compared to the best resolution structure from PDB (ID 1OVA; 1.95 Å; last database access September 2023)35 recording an RMSD of 0.49 Å, pointing to high comparability of the two structures and supporting reliability of AlphaFold structure as a valid model.The 3D structures of the polyphenols used for in silico studies (resveratrol, CID: 445154); piceid, CID: 5281718; phloretin, CID: 4788; phloridzin dihydrate, CID: (4789), were taken from the PubChem database (https://pubchem.ncbi.nlm.nih.gov; last database access September 2023)36 in the Structured Data File format (.sdf). The molecules were converted into Tripos Mol2 format using Open Babel37 for further analysis.
2.5 Molecular docking
Molecular docking was performed to assess the interaction between the selected polyphenols and chymotrypsin and ovalbumin. The simulations were carried out using GOLD software (version 2021).38 The ligands were kept fully flexible and the proteins semi-flexible, allowing polar hydrogens to rotate freely. Local docking was performed defining the chymotrypsin binding site within a 10 Å radius sphere around the centroid of the co-crystallized ligand of the 1AFQ structure. Four water molecules (#507, #539, #595, and #596, according to PDB structure 1AFQ) were keep during docking simulations as found buried within the ligand binding site and thought important to shape the space available to arrange ligands. Additionally, the arrangement of each ligand with the protein pocket was facilitated using the aromatic ‘pharmacophore’ constraint (150 units weight), set in a 0.7 Å radius sphere centred in correspondence to the centroid of aromatic ring of 1AFQ ligand. This promoted the arrangement of the aromatic rings in accordance with the crystallographic data. The ovalbumin binding site was defined based on previous studies39,40 as a 5 Å radius sphere around the pocket's centroid. Ten poses were generated for each ligand, and only the best-scoring pose, according to the internal scoring function PLPScore was used in the analysis.2.6 Molecular dynamics simulations
Molecular dynamics simulations were carried out to verify the stability of proteins-polyphenols and monitor their conformational changes over time.41 They were performed by using GROMACS (version 2019.4) with CHARMM27 all-atom force field parameters support.42 Each ligand was processed and parametrized with the SwissParam tool (https://www.swissparam.ch).43 To investigated how these molecules may interact in a real-time context, an environment as similar as possible to a real biological system was obtained solvating the input structures with SPCE water in a cubic periodic boundary condition and neutralizing the complexes by adding Na+ and Cl− as counter ions. Each system was then energetically minimized to avoid steric clashes and correct improper geometries using the steepest descent algorithm with a maximum of 5000 steps. Subsequently, each system underwent 100 ps simulations under isothermal (300 K, coupling time 2 ps) and isobaric (1 bar, coupling time 2 ps) conditions. Finally, molecular dynamics simulations were run for chymotrypsin-polyphenols and ovalbumin-polyphenols systems. The simulations were 80 ns long and were conducted at 300 K with a coupling time of 0.1 ps and 1 bar with a coupling time of 2 ps, in agreement with previous studies that successfully investigated systems like those under analysis.44,452.7 Statistical analysis
Enzyme activity in the absence of selected polyphenols was compared using one-way analysis of variance (ANOVA) followed by Tukey's post hoc test for statistical analysis. The effect of bioactives on enzyme activity was evaluated using the ANOVA followed by Dunnet's post-test. All the statistical analyses were performed using GraphPad Prism 10 (GraphPad Software Inc.; San Diego, CA, USA).3. Results and discussion
3.1 Pepsin, trypsin, and chymotrypsin activity
| Haemoglobin | Ovalbumin | Gluten | |
|---|---|---|---|
| Pepsin | 1183.6 ± 164.7A,a | 222.4 ± 22.7B,a | 142.5 ± 1.4B,a |
| Trypsin | 98.2 ± 25.3A,b | 58.7 ± 9.3B,b | 24.9 ± 4.4C,c |
| Chymotrypsin | 98.8 ± 10A,b | 47.09 ± 11.4B,b | 56.8 ± 10.6B,b |
 | ||
| Fig. 2 Modulation of pepsin activity by dietary bioactives with haemoglobin (A), ovalbumin (B), or gluten (C) as substrates. Data are means ± SD of at least four replicates and are expressed as fold change with respect to control. Statistical analysis was the ANOVA (all: p < 0.05) followed by Dunnet's post-test with respect to the vehicle (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001). | ||
Fig. 3A–C show similar experiments carried out with trypsin as the enzyme. The impact of glycoside presence on substrate type is particularly noticeable when trypsin is present as an enzyme. Naringin, phloridzin dihydrate, and piceide, which are the glycosylated molecules, are some of the most effective activators of trypsin activity when haemoglobin (Fig. 3A) and ovalbumin (Fig. 3B) are present as substrates. In contrast, the presence of gluten as substrate eliminates the proteolysis-promoting activity of naringin, phloridzin dihydrate, and piceide (Fig. 3C). Furthermore, both quercetin 3-β-O-glucoside and its aglycone demonstrated no effect in the presence of ovalbumin and gluten (Fig. 3B and C), and contrasting effects in the presence of haemoglobin (Fig. 3B). Regardless of the glycoside, the substrate-dependent effect was confirmed. Tyrosol was found to promote trypsin activity in the presence of gluten (Fig. 3C), but no effect in the presence of haemoglobin and ovalbumin (Fig. 3A and B).
 | ||
| Fig. 3 Modulation of trypsin activity by dietary bioactives with haemoglobin (A), ovalbumin (B), or gluten (C) as substrate. Data are means ± SD of at least four replicates and are expressed as fold change with respect to control. Statistical analysis was the ANOVA (all: p < 0.05) followed by Dunnet's post-test with respect to the vehicle (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001). | ||
Modulation of chymotrypsin activity by selected dietary bioactives using haemoglobin, ovalbumin, and gluten as substrates are reported in Fig. 4A–C, respectively. In the case of chymotrypsin, the presence of the glycoside has been shown to modulate enzyme activity.
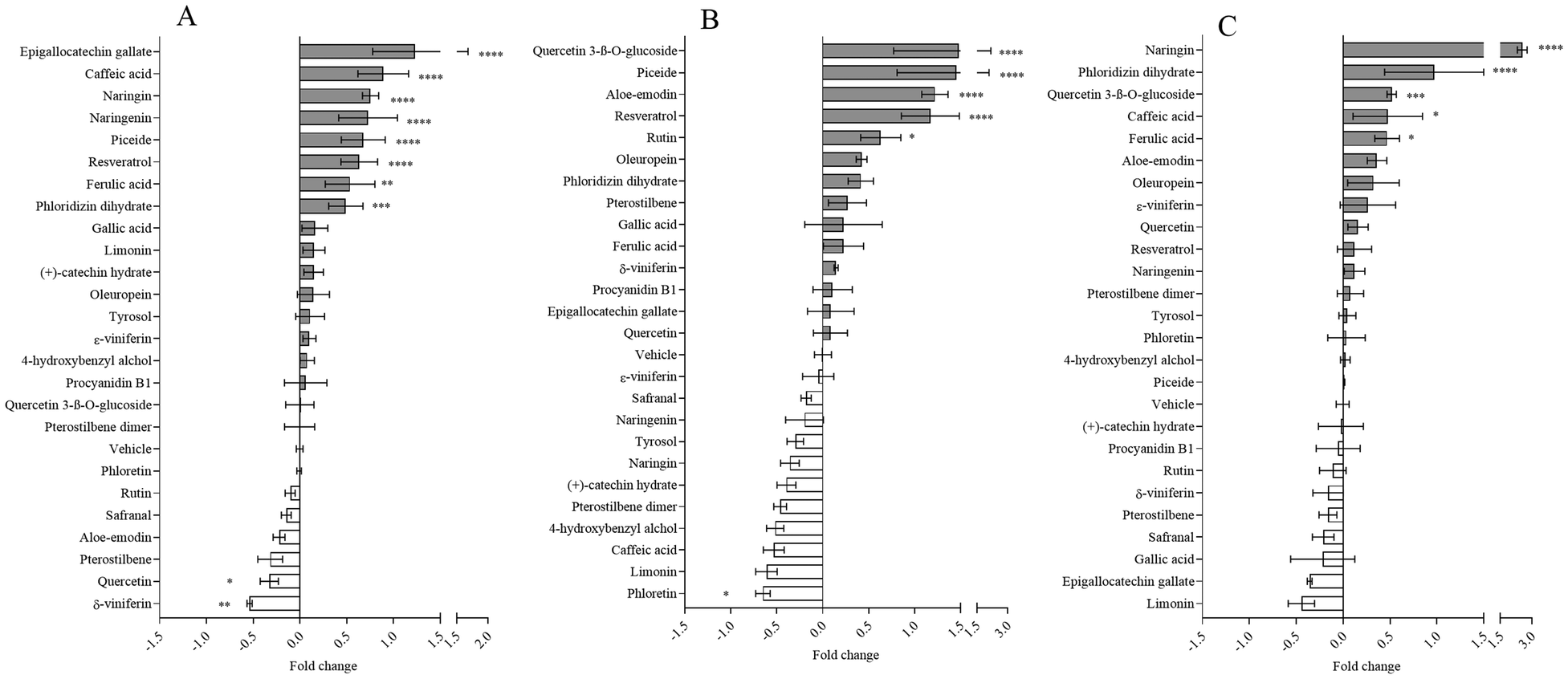 | ||
| Fig. 4 Modulation of chymotrypsin activity by dietary bioactives with haemoglobin (A), ovalbumin (B), or gluten (C) as substrates. Data are means ± SD of at least four replicates and are expressed as fold change with respect to control. Statistical analysis was the ANOVA (all: p < 0.05) followed by Dunnet's post-test with respect to the vehicle (*p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001). | ||
Specifically, the glycoside increases the catalytic activity of phloretin in haemoglobin and gluten (Fig. 4A and C) and of quercetin in ovalbumin and gluten (Fig. 4B and C) compared to the corresponding aglycone (Fig. 4A and C). However, there were no differences in the modulation of enzyme activity between resveratrol and its glycosylated form, regardless of substrate type (Fig. 4A–C). In contrast, the effect of the glycoside was more complex when bound to naringenin. Naringin positively modulated the enzyme's activity in the presence of haemoglobin and gluten (Fig. 4A and C), but had no effect in the presence of ovalbumin (Fig. 4B). The corresponding aglycone, naringenin, increased proteolysis in the presence of haemoglobin (Fig. 4A), but not in the presence of ovalbumin or gluten (Fig. 4B and C). Despite the presence of glycoside, bioactive compounds still had a substrate effect on modulation. Epigallocatechin gallate was found to be a potent promoter or inhibitor of enzyme activity in the presence of haemoglobin (Fig. 4A), respectively, but had no effect when ovalbumin and gluten were present (Fig. 4B and C). It is worth noting that bioactive compounds can have similar or opposing effects on enzyme activity, depending on the type of enzyme and regardless of the substrate. For instance, in the presence of haemoglobin, naringin has been shown to promote all enzymes activity (Fig. 2–4A). On the contrary, in the presence of ovalbumin, resveratrol has been shown to promote chymotrypsin (Fig. 4B) but have no effect on pepsin and trypsin activity (Fig. 2–3B). Regioisomerism had an opposite effect on chymotrypsin activity on haemoglobin in the presence of δ- and ε-viniferin, causing a decrease and a null effect, respectively (Fig. 4A).
A 2-dimensional coloured dataset visualization representing the magnitude of individual values inherent effect of glycosylation and regioisomerism is reported in Fig. 5.
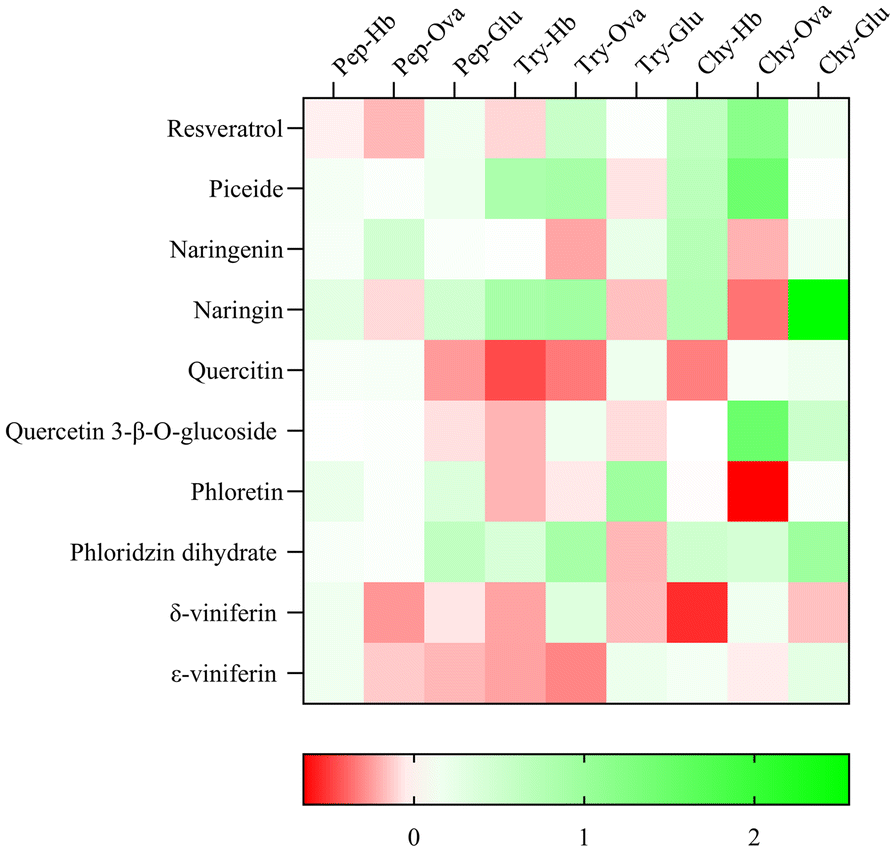 | ||
| Fig. 5 Heatmap representation of the effect of glycosidation and regioisomerism on digestive proteases activity with different substrates. Pep: pepsin; Try: trypsin; Chy: chymotrypsin; Hb: haemoglobin; Ova: ovalbumin; Glu: gluten. | ||
The current literature lacks agreement about the effect of polyphenols and polyphenols-rich food products on the activity of proteolytic enzymes in the digestive tract.17,19,48–51 Recently, Urbinati et al. indicated that the polyphenol-rich balsamic vinegar was able to decrease pepsin, but not trypsin, activity using haemoglobin and p-toluene-sulfonyl-L-arginine methyl ester as substrate.19 On the contrary, Myint et al.49 and Huang et al.48 evidenced that polyphenols from stevia and tea leaves were able to inhibit trypsin activity using the synthetic peptide benzoyl-DL arginine-p-nitroanilide, respectively. Similarly, Naz et al. reported an inhibitory effect of trypsin and chymotrypsin activity by epigallocatechin-3-gallate using N-α-benzoyl-L-arginine ethyl ester and N-acetyl-L-tyrosine ethyl ester, respectively.17 On the contrary, Tantoush et al. reported that pepsin-resistant proteins, such as β-lactoglobulin and Ara h 1 and Ara h 2, degrade faster in the presence of catechin-enriched green tea polyphenols.51 These effects were also confirmed by Tagliazucchi et al. who reported that resveratrol > quercetin > epigallocatechin-3-gallate > catechin enhanced the proteolytic activity of pepsin using haemoglobin as protein substrate by increasing the Vmax of the reaction.50 The conflicting results can be partially explained by the different experimental conditions adopted in these studies, such as bioactives concentration, substrate type and concentration, bioactives/enzyme ratio, bioactives/substrate ratio, and the way activity was assessed (SDS-PAGE, OPA assay, absorbance at 280 nm).52
Also, it is worth noting that many studies have used experimental settings that are far from normal physiological conditions, such as using different substrate types (proteins vs. synthetic peptides) and varying bioactives and substrate concentrations.52
The choice of protein substrates seems to be crucial, especially considering their resistance to digestion and frequency in the diet. Although in the present study the chosen substrates may appear less resistant to digestion compared to legume proteins, a recent article has compared the protein digestibility of cereals and legumes.21 The main findings indicate that the protein digestibility of legumes was generally higher than that of cereals, with wheat having a protein digestibility comparable to that of most of the legumes studied (adzuki bean, pea, lentil and soya bean). Furthermore, although gluten is not the primary dietary source of protein, cereals and cereal products are a fundamental part of the human diet, providing a significant proportion of dietary energy and nutrients in both developed and developing countries.53
Also the selection of bioactives concentration represents a critical factor in achieving a result that can be extrapolated to reflect the in vivo scenario, particularly considering their well-established concentration-dependent effect on the modulation of protease activity.49,50 In a recent study, Vyas et al. reported that curcumin, diosmin, morin, and 2′,3′,4′-trihydroxychalcone were able to inhibit pepsin, trypsin, and chymotrypsin activity, with the degree of inhibition being concentration-dependent.54
To circumvent these limitations, the enzyme's activity was appraised through a standardised approach at a single concentration that exclusively comprised native proteins and bioactives. The substrate concentration (2.47%) and bioactive concentration (0.1 mM) were selected in accordance with the standardised static in vitro digestion INFOGEST protocol55 and predicted gut polyphenols concentration,4 respectively. In a recent study, Tamargo et al. demonstrated that the total concentration of hydroxybenzoic acids, hydroxycinnamic acids, flavonols and flavan-3-ols in red wine after in vitro gastrointestinal digestion was approximately 0.18 mM.56 Indeed, the concentration of polyphenols within the intestine can fluctuate between 0.4 and 3 mM, contingent upon the quantity of these compounds present in the ingested food, the portion size consumed, and the extent to which they are released into the digestive tract (bioaccessibility). Consequently, it is challenging to establish a universally applicable value.
For this reason, the present study focused exclusively on a single concentration of bioactives and protein substrates, as previously reported.11,57 The addition of other concentrations would have resulted in an excess of experimental conditions, rendering it unfeasible to comprehensively explicate and discuss all the results within the confines of a single study. However, further studies are required to ascertain the impact of bioactive concentration on the diet.
The varying effects found among different bioactive compounds cannot be solely attributed to the different experimental conditions adopted in the studies. The effects on protein digestibility may be due to the binding of the bioactives compounds to digestive proteases and protein substrates, which is dependent on their chemical structure, affecting protease activity and accessibility of protein substrates. Enzyme activity may vary because of phenolic chemical binding, which generally can relax or stabilize enzyme structure. Besides competitive inhibitors targeting the substrate binding site, enzymes can be inhibited when their structure is altered to the point of disrupting the correct positioning and orientation of substrate and/or catalytic residues. For this reason, it is important to predict protein conformational changes, particularly for phenolics with bulky structures such as tannins or glycosides of phenolic compounds.58
Furthermore, the binding of phenolic compounds can hinder the catalytic or substrate binding sites, reducing the proteolytic activity 7 or causing enzyme inhibition through allosteric effects. On the contrary, polyphenols may act as allosteric effectors59 or modify enzyme conformation (stabilization) for a better fitting of protein/substrate.60 Like proteases, polyphenolic compounds can either stabilize or destabilize the structure of protein substrates, making them less or more susceptible to proteolytic attack, respectively. Phenols may specifically reduce the accessibility of proteases that prefer hydrophobic residues61 by binding to proteins and primarily interacting with their hydrophobic sites.62 On the contrary, the binding of phenolic compounds can destabilize the protein structure by causing partial unfolding, which increases the protein's accessibility to enzymes. Generally, phenolic compounds may influence protein hydrolysis through their interactions with both the protein substrate and the protease itself, making it difficult to determine the net effect of phenolic compounds on protease activity when using protein substrate. In this complicated but intriguing scenario, in silico investigations can help to study the behaviour of small molecules, within the binding site of a target protein and understand the fundamental molecular mechanisms underlying their action.63
3.2 In silico bioactives interaction with chymotrypsin and ovalbumin
Based on docking analysis and the positive scores recorded (Table 2; the higher the score, the better the interaction, as per manufacturer declaration), the selected polyphenols (i.e. resveratrol, piceid, phloretin, and phloridzin dihydrate) seemed to favourably interact with both chymotrypsin and ovalbumin.
| Chymotrypsin | Ovalbumin (binding site) | |
|---|---|---|
| Resveratrol | 201.11 | 44.95 |
| Piceid | 214.93 | 44.73 |
| Phloretin | 211.75 | 52.27 |
| Phloridzin dihydrate | 212.62 | 53.77 |
Specifically, as per manufacturer declaration (https://www.ccdc.cam.ac.uk), positive scores indicate the capability of ligands to satisfy the physicochemical requirements of the pocket and can be used to estimate the likeliness of protein–ligand complex formation. Germane to the case study, all the ligands recorded positive scores pointing to their theoretical capability to favourably interact with both proteins. Of note, the scores for chymotrypsin were around four times higher than those recorded for ovalbumin due to the constraint used to dock the ligands within the former which enhances the score when satisfied. This prevented the inter-system comparison of scores, which were instead comparable intra-system, describing the ligands considered similarly able to satisfy the requirements of each pocket.
The analysis of the docking poses revealed the capability of all the tested polyphenols to display polar interactions (hydrogen bonds) with both residues of the binding pocket and water molecules. Specifically, resveratrol (Fig. 6B) and phloretin (Fig. 6C) docked the binding site in accordance with the pharmacophore constraint. Conversely, piceid (Fig. 6D) and phloridzin dihydrate (Fig. 6E) placed the sugar moiety in the inner part of the pocket. In either case, polyphenols could theoretically interact close to the enzyme's catalytic triad (Ser195, His57, Asp102),67 suggesting their capability to compete with enzyme's substrates (Fig. 6A–D). In more detail, as reported in Fig. 6, the hydrogen bond distances between the polyphenols under investigation and the residues and water molecules were calculated at the catalytic site. The analysis revealed that the whole set of polyphenols could interact with at least one of the water molecules, which may act as a bridge to the protein. Moreover, three out of four of the considered polyphenols (i.e. phloretin, piceid and phloridzin dihydrate) also interact with at least one of the residues of the catalytic triad. Interestingly, all of them recorded an interaction with Ser195 characterized by a distance lower than that observed for the known non-covalent synthetic inhibitor co-crystallized in the 1AFQ structure68 (Fig. 6A). Conversely, resveratrol showed a slightly different mode of interaction with no direct polar contacts with the catalytic triad, though it was found interacting close to the active pocket contacting Ser217 (data not shown). As a general comment, different interaction networks were observed for the considered polyphenols, though all in line with the architecture of binding previously reported for other proteases inhibitors either in terms of polar bond length or residues involved.45,69
 | ||
| Fig. 6 Docking poses of the selected polyphenols within chymotrypsin and ovalbumin. Proteins are represented in white cartoons, while polyphenols and the amino acids of the proteins involved in polar interactions are represented in sticks. The water molecules at the chymotrypsin binding site are shown in balls-and-sticks. Yellow dashed lines indicate hydrogen bonds (interatomic distances in Å). Interaction of a known synthetic inhibitor with chymotrypsin binding site (as per PDB structure 1AFQ) (A). Interaction of resveratrol (cyan sticks) within chymotrypsin binding site (B). Interaction of phloretin (pink sticks) within chymotrypsin binding site (C). Interaction of piceid (green sticks) within chymotrypsin binding site (D). Interaction of phloridzin dihydrate (green sticks) within chymotrypsin binding site (E). Interaction of resveratrol (cyan sticks; F), phloretin (pink sticks; G), piceid (green sticks; H) and phloridzin dihydrate (yellow sticks; I) within ovalbumin binding site (F–I). | ||
Therefore, all the tested polyphenols were found to be potential chymotrypsin inhibitors. This result is consistent with previous evidence that generally describes polyphenols as inhibitors of peptidases, including chymotrypsin.7 For instance, it was demonstrated that resveratrol was capable of decreasing the activity of trypsin.70 This finding suggests a possible inhibitory effect against chymotrypsin as well, based on the high similarity between the two enzymes, which enables many inhibitors to act on both. This hypothesis has been widely and convincingly demonstrated for other inhibitors.71 With respect to ovalbumin, similarly to what observed for chymotrypsin and based on the positive docking scores recorded (Table 2), piceid, resveratrol, phloretin, and phloridzin dihydrate were predicted to interact positively with the designated binding pocket (Fig. 6F–I). Of note, the pocket was chosen based on previous evidence attesting the capability to bind a variety of ligands, including polyphenols.39,40
Taken together, the results of docking simulations could not provide a plausible explanation for the diverse impact of the selected polyphenols on the chymotrypsin activity over ovalbumin as all of them could theoretically interact with both. In particular, the interaction with chymotrypsin should have been reasonably resulted in a reduction of ovalbumin degradation, laying unexplained the enhanced degradation observed for resveratrol, piceid and phloridzin dihydrate. Therefore, the best scored docking pose of each compound when in complex with both chymotrypsin and ovalbumin was used as input for molecular dynamics simulations to check the evolution of each complex overtime. This may allow investigating further the molecular mechanisms underpinning the diverse outcome the considered polyphenols had on chymotrypsin in vitro. It arose the hypothesis that the considered polyphenols may induce a diverse dynamic of ovalbumin, making it a better substrate for hydrolysis when in complex with resveratrol, piceid and eventually with phloridzin dihydrate than when in complex with phloretin.
With respect to the complex with chymotrypsin, the outcome of molecular dynamics revealed that all the polyphenols under investigation were able to stably interact with the protease, aligning with the steady state RMSD trend of ligands and protein (Fig. 7A and B). Of note, this is a measurement able to determine whether the ligands were stably interacting at the designated binding site.72 Regardless of the ligand bound, the protein experienced a geometrical settling and reached stability from around 45 ns onward, suggesting that after an initial phase of adjustment, the systems remained stable over time. With respect to ligands, they showed a steady state interaction from around 25 ns onward, except for piceid which showed the highest mobility though within the range observed for the other ligands. These results confirmed the theoretical capability of the four polyphenols considered to have a certain degree of inhibitory activity against chymotrypsin, as discussed above and in line with previous evidence.70 This corroborated the hypothesis that they might induce differential dynamics when in complex with ovalbumin.
 | ||
| Fig. 7 Ligands and chymotrypsin RMSD variations. Resveratrol (blue), piceid (orange), phloretin (green), and phloridzin dihydrate (light blue) RMSD variations within chymotrypsin over time (A). Chymotrypsin C-alpha RMSD variations when in complex with resveratrol (blue), piceid (orange), phloretin (green), and phloridzin dihydrate (light blue) (B). | ||
Therefore, an 80 ns molecular dynamics simulation was performed for each polyphenol within the ovalbumin binding site (Fig. 8).
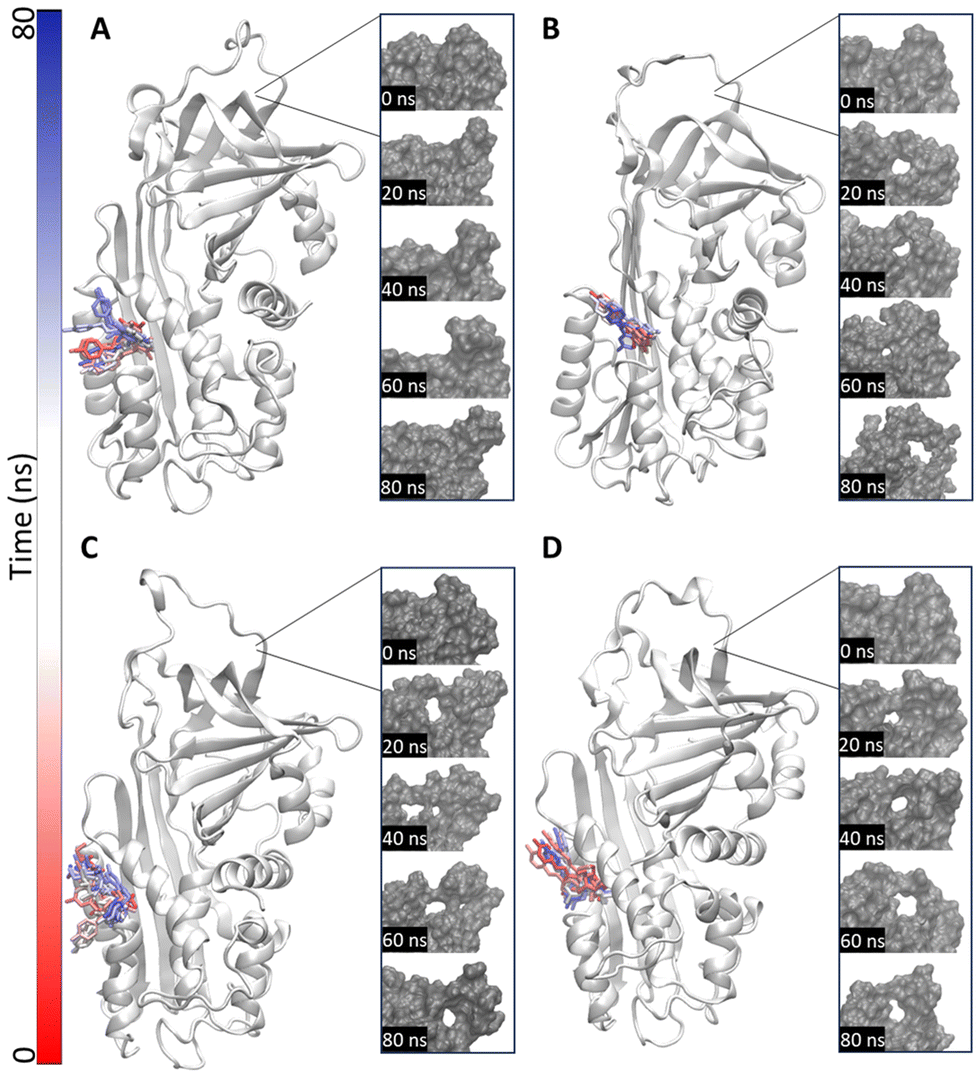 | ||
| Fig. 8 Molecular dynamics results of the selected polyphenols within ovalbumin and close-up on the newly formed pockets. Time step representation of phloretin (A), resveratrol (B), phloridzin dihydrate (C), and piceide (D) within ovalbumin and close-up of the new pocket at different times (0 ns, 20 ns, 40 ns, 60 ns, 80 ns). Protein is represented in white cartoons and in grey surface; polyphenols are represented in sticks. The red-to-blue colour switch indicates the stepwise changes of coordinates during the simulations. | ||
The results indicate that phloretin had a different effect on ovalbumin compared to other polyphenols. Indeed, during the entire simulation, ovalbumin appeared well compacted when in complex with the digestion inhibitor phloretin (Fig. 8A). In contrast, when complexed with resveratrol, piceid, or phloridzin dihydrate, ovalbumin exhibited the opening of an additional surface cleft (Fig. 8B–D), which might prelude conformational changes allowing an increased accessibility to proteases and a subsequent enhanced susceptibility to proteolytic cleavage.
This phenomenon could provide a plausible though partial explanation to the diverse outcome the selected polyphenols showed in vitro.
4. Conclusion
This study demonstrates how food-bioactive compounds can affect the proteolytic activity of digestive proteases in an enzyme- and substrate-specific way. Additionally, it has been observed that glycosylation in small molecules can significantly alter reaction kinetics. These differences may be due to non-covalent interactions – mainly hydrogen bonds – at the substrate or enzyme level. In this scenario, the assessment of enzymatic activity in an assay alone is insufficient for predicting interactions among bioactives, enzymes, and substrates. To gain a deeper understanding of the mechanistic basis of in vitro outcomes from a molecular standpoint, it is necessary to conduct in silico analysis. The in silico studies reported here demonstrate that all the chosen polyphenols can interact with chymotrypsin in a way that likely impairs enzyme activity. However, the polyphenols that enhance in vitro proteolytic activity can diversely impact on the substrate structure, reasonably making it more susceptible to undergo the proteolytic cleavage. This suggests a combined substrate- and ligand-dependent relationship that may explain why polyphenols can have opposite effects on protein digestion.To conclude, this study proves the feasibility and the significance of defining – on a molecular basis – the role of phenolics in modulating the activity of proteolytic enzymes in the digestive tract. Furthermore, this study highlights the importance of specific structural characteristics associated with the observed effects of these bioactive compounds. These characteristics include their three-dimensional structure, binding affinity, orientation, and geometry. Thus, this report is expected to contribute to further evaluation of this family of bioactive compounds as “digestive modulating agents” in the formulation of functional foods targeting specific consumers’ needs. In this frame, it is worth underscoring that isolated food components may exhibit a different behaviour if compared to those occurring when they are part of a food matrix, and this ‘matrix effect’ must be duly considered when evaluating any property of the system and all the resulting nutritional and health implications. In this intricate scenario, in vitro digestion assays can be regarded as a valuable research method for investigating and comprehending alterations, interactions, and the bioaccessibility of nutrients and non-nutritive substances in food.73 Specifically, it can be utilised to evaluate digestion endpoints or the kinetics of digestion phases, such as gastric and small intestinal hydrolysis.74,75 Notwithstanding the limitations of in vitro methods in fully reflecting the intricacies of in vivo digestion, they are regarded as valuable screening tools for various diet-related concerns, including digestibility, the release of bioactive compounds, structural changes in food, and the interaction between nutrients and bioactive food components. In any case, further investigations are required, as there is a paucity of in vivo studies in animal models and human, in contrast with the promising in vitro and in silico data.
Author contributions
Conceptualization: A.P., S.D., S.I., M.D.N. Methodology: L.D., M.D.N. Software: F. P., L.D., L. P. Validation: L.D., M.D.N. Formal analysis: S.M.B., F.P., L.D., L.P., M.D.N. Investigation: S.M.B., F.P., L.P, F.A. Resources: G.G., A.P., S.D., S.I., M.D.N. Data curation: S.M.B., F.P., L.P., F.A. Writing – original draft: S.M.B., F.P., L.D., M.D.N. Writing – Review & editing: G.G., A.P., S.D., S.I., M.D.N. Visualization: S.M.B., F.P. Supervision: L.D., M.D.N. Project administration: L.D., M.D.N. Funding acquisition: G.G., A.P., S.D., S.I., M.D.N.Data availability
The authors confirm that the data supporting the findings of this study are available within the article.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This investigation was supported in part by the National Recovery and Resilience Plan (NRRP), (Mission 4, Component 2, Investment 1.3 - Call for tender No. 341 of 15/03/2022) of the Italian Ministry of University and Research, funded by the European Union–NextGenerationEU, in the frame of the project “Research and innovation network on food and nutrition: Sustainability, Safety and Security (ON Foods), and by the Italian Piano di Sostegno della Ricerca 2022 - Azione A (Linea 2), University of Milan. This research benefits from the HPC (High Performance Computing) facility of the University of Parma, Italy.References
- C. Di Lorenzo, F. Colombo, S. Biella, C. Stockley and P. Restani, Polyphenols and Human Health: The Role of Bioavailability, Nutrients, 2021, 13, 273 CrossRef PubMed.
- A. Masyita, R. Mustika Sari, A. Dwi Astuti, B. Yasir, N. Rahma Rumata, T. B. Emran, F. Nainu and J. Simal-Gandara, Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives, Food Chem.: X, 2022, 13, 100217 CAS.
- D. W. Griffiths, The inhibition of digestive enzymes by polyphenolic compounds, Adv. Exp. Med. Biol., 1986, 199, 509–516 CrossRef CAS PubMed.
- G. Williamson, Possible effects of dietary polyphenols on sugar absorption and digestion, Mol. Nutr. Food Res., 2013, 57, 48–57 CrossRef CAS PubMed.
- L. Sun and M. Miao, Dietary polyphenols modulate starch digestion and glycaemic level: a review, Crit. Rev. Food Sci. Nutr., 2020, 60, 541–555 CrossRef CAS PubMed.
- L. Sun, F. J. Warren and M. J. Gidley, Natural products for glycaemic control: Polyphenols as inhibitors of alpha-amylase, Trends Food Sci. Technol., 2019, 91, 262–273 CrossRef CAS.
- A. I. Martinez-Gonzalez, Á.G Díaz-Sánchez, L. A. Rosa, C. L. Vargas-Requena, I. Bustos-Jaimes and A. E. Alvarez-Parrilla, Polyphenolic Compounds and Digestive Enzymes: In Vitro Non-Covalent Interactions, Molecules, 2017, 22, 669 CrossRef PubMed.
- J. D. J. Rodríguez-Romero, A. C. Durán-Castañeda, A. P. Cárdenas-Castro, J. A. Sánchez-Burgos, V. M. Zamora-Gasga and S. G. Sáyago-Ayerdi, What we know about protein gut metabolites: Implications and insights for human health and diseases, Food Chem.: X, 2022, 13, 100195 Search PubMed.
- Y. H. Dai, J. R. Wei and X. Q. Chen, Interactions between tea polyphenols and nutrients in food, Compr. Rev. Food Sci. Food Saf., 2023, 22, 3130–3150 CrossRef CAS PubMed.
- T. Ozdal, E. Capanoglu and F. Altay, A review on protein–phenolic interactions and associated changes, Food Res. Int., 2013, 51, 954–970 CrossRef CAS.
- Q. He, Y. Lv and K. Yao, Effects of tea polyphenols on the activities of α-amylase, pepsin, trypsin and lipase, Food Chem., 2007, 101, 1178–1182 CrossRef CAS.
- Ö. Dönmez, B. A. Mogol, V. Gökmen, N. Tang, M. L. Andersen and D. E. W. Chatterton, Modulation of gastrointestinal digestion of β-lactoglobulin and micellar casein following binding by (−)-epigallocatechin-3-gallate (EGCG) and green tea flavanols, Food Funct., 2020, 11, 6038–6053 RSC.
- L. M. Mattio, G. Catinella, A. Pinto and S. Dallavalle, Natural and nature-inspired stilbenoids as antiviral agents, Eur. J. Med. Chem., 2020, 202, 112541 CrossRef CAS PubMed.
- E. Almanza-Aguilera, E. Davila-Cordova, D. Guiñón-Fort, M. Farràs, G. Masala, M. Santucci de Magistris, I. Baldassari, R. Tumino, L. Padroni, V. A. Katzke, M. B. Schulze, A. Scalbert and R. Zamora-Ros, Correlation Analysis between Dietary Intake of Tyrosols and Their Food Sources and Urinary Excretion of Tyrosol and Hydroxytyrosol in a European Population, Antioxidant, 2023, 12, 715 CrossRef CAS PubMed.
- K. A. Woodward, R. Draijer, D. H. J. Thijssen and D. A. Low, Polyphenols and Microvascular Function in Humans: A Systematic Review, Curr. Pharm. Des., 2018, 24, 203–226 CrossRef CAS PubMed.
- N. Shen, T. Wang, Q. Gan, S. Liu, L. Wang and B. Jin, Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity, Food Chem., 2022, 383, 132531 CrossRef CAS PubMed.
- S. Naz, R. Siddiqi, T. P. Dew and G. Williamson, Epigallocatechin-3-gallate inhibits lactase but is alleviated by salivary proline-rich proteins, J. Agric. Food Chem., 2011, 59, 2734–2738 CrossRef CAS PubMed.
- M. Ramm, B. Alarcón-Zapata, J. Monsalves and L. Bustamante, A Simple Validated Method for the Estimation of Pepsin Activity in Microtiter Array for the INFOGEST Protocol, Foods, 2023, 12, 3851 CrossRef CAS PubMed.
- E. Urbinati, M. Di Nunzio, G. Picone, E. Chiarello, A. Bordoni and F. Capozzi, The Effect of Balsamic Vinegar Dressing on Protein and Carbohydrate Digestibility is Dependent on the Food Matrix, Foods, 2021, 10, 411 CrossRef CAS PubMed.
- A. Vesković, D. Bajuk-Bogdanović, V. B. Arion and A. Popović Bijelić, Spectroscopic Characterization of the Binding and Release of Hydrophilic, Hydrophobic and Amphiphilic Molecules from Ovalbumin Supramolecular Hydrogels, Gels, 2022, 9, 14 CrossRef PubMed.
- X. Xu, J. Tao, Q. Wang, J. Ge, J. Li, F. Gao, S. Gao, Q. Yang, B. Feng and J. Gao, A comparative study: Functional, thermal and digestive properties of cereal and leguminous proteins in ten crop varieties, LWT, 2023, 187, 115288 CrossRef CAS.
- K. Kłosok, R. Welc, E. Fornal and A. Nawrocka, Effects of Physical and Chemical Factors on the Structure of Gluten, Gliadins and Glutenins as Studied with Spectroscopic Methods, Molecules, 2021, 26, 508 CrossRef PubMed.
- Y. Feng, M. Lv, Y. Lu, K. Liu, L. Liu, Z. He, K. Wu, X. Wang, B. Zhang and X. Wu, Characterization of binding interactions between selected phenylpropanoid glycosides and trypsin, Food Chem., 2018, 243, 118–124 CrossRef CAS PubMed.
- X. Wu, W. Wang, T. Zhu, T. Liang, F. Lu, W. He, H. Zhang, Z. Liu, S. He, K. Gao and Z. He, Phenylpropanoid glycoside inhibition of pepsin, trypsin and α-chymotrypsin enzyme activity in Kudingcha leaves from Ligustrum purpurascens, Food Res. Int., 2013, 54, 1376–1382 CrossRef CAS.
- N. Dehdasht-Heidari, B. Shareghi, S. Farhadian and L. Momeni, Investigation on the interaction behavior between safranal and pepsin by spectral and MD simulation studies, J. Mol. Liq., 2021, 344, 117903 CrossRef CAS.
- G. Ren, H. Sun, G. Li, J. Fan, L. Du and G. Cui, Interaction mechanism of aloe-emodin with trypsin: molecular structure–affinity relationship and effect on biological activities, RSC Adv., 2020, 10, 20862–20871 RSC.
- L. M. Mattio, S. Dallavalle, L. Musso, R. Filardi, L. Franzetti, L. Pellegrino, P. D'Incecco, D. Mora, A. Pinto and S. Arioli, Antimicrobial activity of resveratrol-derived monomers and dimers against foodborne pathogens, Sci. Rep., 2019, 9, 19525 CrossRef CAS PubMed.
- S. Dallavalle, R. Artali, S. Princiotto, L. Musso, G. Borgonovo and S. Mazzini, Investigation of the Interaction between Aloe vera Anthraquinone Metabolites and c-Myc and C-Kit G-Quadruplex DNA Structures, Int. J. Mol. Sci., 2022, 23, 16018 CrossRef CAS PubMed.
- G. Catinella, G. Borgonovo, S. Dallavalle, M. L. Contente and A. Pinto, From saffron residues to natural safranal: Valorization of waste through a β-glucosidase, Food Bioprod. Process., 2022, 131, 144–148 CrossRef CAS.
- F. Magurano, M. Sucameli, P. Picone, M. Micucci, M. Baggieri, A. Marchi, P. Bucci, S. Gioacchini, G. Catinella, G. Borgonovo, S. Dallavalle, D. Nuzzo and A. Pinto, Antioxidant Activity of Citrus Limonoids and Investigation of Their Virucidal Potential against SARS-CoV-2 in Cellular Models, Antioxidants, 2021, 10, 1794 CrossRef CAS PubMed.
- S. M. Borgonovi, E. Chiarello, F. Pasini, G. Picone, S. Marzocchi, F. Capozzi, A. Bordoni, A. Barbiroli, A. Marti, S. Iametti and M. Di Nunzio, Effect of Sprouting on Biomolecular and Antioxidant Features of Common Buckwheat (Fagopyrum esculentum), Foods, 2023, 12, 2047 CrossRef CAS PubMed.
- F. Frigerio, A. Coda, L. Pugliese, C. Lionetti, E. Menegatti, G. Amiconi, H. P. Schnebli, P. Ascenzi and M. Bolognesi, Crystal and molecular structure of the bovine alpha-chymotrypsin-eglin c complex at 2.0 A resolution, J. Mol. Biol., 1992, 225, 107–123 CrossRef CAS PubMed.
- E. F. Pettersen, T. D. Goddard, C. C. Huang, G. S. Couch, D. M. Greenblatt, E. C. Meng and T. E. Ferrin, UCSF Chimera–a visualization system for exploratory research and analysis, J. Comput. Chem., 2004, 25, 1605–1612 CrossRef CAS PubMed.
- J. Jumper, R. Evans, A. Pritzel, T. Green, M. Figurnov, O. Ronneberger, K. Tunyasuvunakool, R. Bates, A. Žídek, A. Potapenko, A. Bridgland, C. Meyer, S. A. A. Kohl, A. J. Ballard, A. Cowie, B. Romera-Paredes, S. Nikolov, R. Jain, J. Adler, T. Back, S. Petersen, D. Reiman, E. Clancy, M. Zielinski, M. Steinegger, M. Pacholska, T. Berghammer, S. Bodenstein, D. Silver, O. Vinyals, A. W. Senior, K. Kavukcuoglu, P. Kohli and D. Hassabis, Highly accurate protein structure prediction with AlphaFold, Nature, 2021, 596, 583–589 CrossRef CAS PubMed.
- P. E. Stein, A. G. W. Leslie, J. T. Finch and R. W. Carrell, Crystal structure of uncleaved ovalbumin at 1·95 Å resolution, J. Mol. Biol., 1991, 221, 941–959 CrossRef CAS PubMed.
- S. Kim, J. Chen, T. Cheng, A. Gindulyte, J. He, S. He, Q. Li, B. A. Shoemaker, P. A. Thiessen, B. Yu, L. Zaslavsky, J. Zhang and E. E. Bolton, PubChem 2023 update, Nucleic Acids Res., 2022, 51, D1373–D1380 CrossRef PubMed.
- N. M. O'Boyle, M. Banck, C. A. James, C. Morley, T. Vandermeersch and G. R. Hutchison, Open Babel: An open chemical toolbox, J. Cheminf., 2011, 3, 33 Search PubMed.
- L. Dellafiora, G. Galaverna, C. Dall'Asta and P. Cozzini, Hazard identification of cis/trans-zearalenone through the looking-glass, Food Chem. Toxicol., 2015, 86, 65–71 CrossRef CAS PubMed.
- P. Manivel, P. Marimuthu, S. Yu and X. Chen, Multispectroscopic and Computational Investigations on the Binding Mechanism of Dicaffeoylquinic Acids with Ovalbumin, J. Chem. Inf. Model., 2022, 62, 6133–6147 CrossRef CAS PubMed.
- W. Zhou, C. Peng, D. Wang, J. Li, Z. Tu and L. Zhang, Interaction Mechanism between OVA and Flavonoids with Different Hydroxyl Groups on B-Ring and Effect on Antioxidant Activity, Foods, 2022, 11, 1302 CrossRef CAS PubMed.
- M. J. Abraham, T. Murtola, R. Schulz, S. Páll, J. C. Smith, B. Hess and E. Lindahl, GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers, SoftwareX, 2015, 1–2, 19–25 CrossRef.
- R. B. Best, X. Zhu, J. Shim, P. E. Lopes, J. Mittal, M. Feig and A. D. Mackerell Jr, Optimization of the additive CHARMM all-atom protein force field targeting improved sampling of the backbone φ, ψ and side-chain χ(1) and χ(2) dihedral angles, J. Chem. Theory Comput., 2012, 8, 3257–3273 CrossRef CAS PubMed.
- V. Zoete, M. A. Cuendet, A. Grosdidier and O. Michielin, SwissParam: a fast force field generation tool for small organic molecules, J. Comput. Chem., 2011, 32, 2359–2368 CrossRef CAS PubMed.
- W. Cheng, M. Wang, C. Li, F. Xiao, J. He, L. Liu, H. Niu and J. Ma, Study of the weak interaction mechanism of ovalbumin and caffeic acid using fluorescence spectroscopy and molecular dynamics simulation, Spectrochim. Acta, Part A, 2023, 301, 122966 CrossRef CAS PubMed.
- F. Cui, K. Yang and Y. Li, Investigate the binding of catechins to trypsin using docking and molecular dynamics simulation, PLoS One, 2015, 10, e0125848 CrossRef PubMed.
- L. Salelles, J. Floury and S. Le Feunteun, Pepsin activity as a function of pH and digestion time on caseins and egg white proteins under static in vitro conditions, Food Funct., 2021, 12, 12468–12478 RSC.
- M. Smith and D. Morton, in Digestion and Absorption, Elsevier, 2010, pp. 129–152, DOI:10.1016/B978-0-7020-3367-4.00008-6.
- H. Huang, K.-C. Kwok and H. Liang, Effects of tea polyphenols on the activities of soybean trypsin inhibitors and trypsin, J. Sci. Food Agric., 2004, 84, 121–126 CrossRef CAS.
- K. Z. Myint, Z. Zhou, Q. Shi, J. Chen, X. Dong and Y. Xia, Stevia Polyphenols, Their Antimicrobial and Anti-Inflammatory Properties, and Inhibitory Effect on Digestive Enzymes, Molecules, 2023, 28, 7572 CrossRef CAS PubMed.
- D. Tagliazucchi, E. Verzelloni and A. Conte, Effect of some phenolic compounds and beverages on pepsin activity during simulated gastric digestion, J. Agric. Food Chem., 2005, 53, 8706–8713 CrossRef CAS PubMed.
- Z. Tantoush, D. Apostolović, B. N. Kravic, I. Prodić, L. Mihajlovic, D. Stanić-Vučinić and T. C. Velickovic, Green tea catechins of food supplements facilitate pepsin digestion of major food allergens, but hampers their digestion if oxidized by phenol oxidase, J. Funct. Foods, 2012, 4, 650–660 CrossRef CAS.
- T. D. Cirkovic Velickovic and D. J. Stanic-Vucinic, The Role of Dietary Phenolic Compounds in Protein Digestion and Processing Technologies to Improve Their Antinutritive Properties, Compr. Rev. Food Sci. Food Saf., 2018, 17, 82–103 CrossRef CAS PubMed.
- W. Laskowski, H. Górska-Warsewicz, K. Rejman, M. Czeczotko and J. Zwolińska, How Important are Cereals and Cereal Products in the Average Polish Diet?, Nutrients, 2019, 11, 679 CrossRef CAS PubMed.
- K. Vyas, S. Prabaker, D. Prabhu, M. Sakthivelu, S. Rajamanikandan, P. Velusamy, C. H. Su, S. C. B. Gopinath and R. Pachaiappan, Study of an inhibitory effect of plant polyphenolic compounds against digestive enzymes using bench-working experimental evidence predicted by molecular docking and dynamics, Int. J. Biol. Macromol., 2024, 259, 129222 CrossRef CAS PubMed.
- M. Minekus, M. Alminger, P. Alvito, S. Ballance, T. Bohn, C. Bourlieu, F. Carrière, R. Boutrou, M. Corredig, D. Dupont, C. Dufour, L. Egger, M. Golding, S. Karakaya, B. Kirkhus, S. Le Feunteun, U. Lesmes, A. Macierzanka, A. Mackie, S. Marze, D. J. McClements, O. Ménard, I. Recio, C. N. Santos, R. P. Singh, G. E. Vegarud, M. S. J. Wickham, W. Weitschies and A. Brodkorb, A standardised static in vitro digestion method suitable for food – an international consensus, Food Funct., 2014, 5, 1113–1124 RSC.
- A. Tamargo, C. Cueva, M. Silva, N. Molinero, B. Miralles, B. Bartolomé and M. V. Moreno-Arribas, Gastrointestinal co-digestion of wine polyphenols with glucose/whey proteins affects their bioaccessibility and impact on colonic microbiota, Food Res. Int., 2022, 155, 111010 CrossRef CAS PubMed.
- Q. Li, Q. Wei, E. Yuan, J. Yang and Z. Ning, Interaction between four flavonoids and trypsin: effect on the characteristics of trypsin and antioxidant activity of flavonoids, Int. J. Food Sci. Technol., 2014, 49, 1063–1069 CrossRef CAS.
- G. Giuberti, G. Rocchetti and L. Lucini, Interactions between phenolic compounds, amylolytic enzymes and starch: an updated overview, Curr. Opin. Food Sci., 2020, 31, 102–113 Search PubMed.
- I. Grabska-Kobyłecka, P. Szpakowski, A. Król, D. Książek-Winiarek, A. Kobyłecki, A. Głąbiński and D. Nowak, Polyphenols and Their Impact on the Prevention of Neurodegenerative Diseases and Development, Nutrients, 2023, 15, 3454 Search PubMed.
- F. Shahidi and C. S. Dissanayaka, Phenolic-protein interactions: insight from in-silico analyses – a review, Food Prod., Process. Nutr., 2023, 5, 2 CAS.
- A. I. Petushkova, L. V. Savvateeva and A. A. Zamyatnin Jr, Structure determinants defining the specificity of papain-like cysteine proteases, Comput. Struct. Biotechnol. J., 2022, 20, 6552–6569 CAS.
- M. Li, C. Ritzoulis, Q. Du, Y. Liu, Y. Ding, W. Liu and J. Liu, Recent Progress on Protein-Polyphenol Complexes: Effect on Stability and Nutrients Delivery of Oil-in-Water Emulsion System, Front. Nutr., 2021, 8, 765589 Search PubMed.
- P. C. Agu, C. A. Afiukwa, O. U. Orji, E. M. Ezeh, I. H. Ofoke, C. O. Ogbu, E. I. Ugwuja and P. M. Aja, Molecular docking as a tool for the discovery of molecular targets of nutraceuticals in diseases management, Sci. Rep., 2023, 13, 13398 CAS.
- L. Pedroni, F. Perugino, G. Galaverna, C. Dall’Asta and L. Dellafiora, An In Silico Framework to Mine Bioactive Peptides from Annotated Proteomes: A Case Study on Pancreatic Alpha Amylase Inhibitory Peptides from Algae and Cyanobacteria, Nutrients, 2022, 14, 4680 CAS.
- M. Zakłos-Szyda, N. Pietrzyk, A. Kowalska-Baron, A. Nowak, K. Chałaśkiewicz, M. Ratajewski, G. Budryn and M. Koziołkiewicz, Phenolics-Rich Extracts of Dietary Plants as Regulators of Fructose Uptake in Caco-2 Cells via GLUT5 Involvement, Molecules, 2021, 26, 4745 Search PubMed.
- S. A. Hollingsworth and R. O. Dror, Molecular Dynamics Simulation for All, Neuron, 2018, 99, 1129–1143 CAS.
- G. J. C. Vreeke, J. P. Vincken and P. A. Wierenga, The path of proteolysis by bovine chymotrypsin, Food Res. Int., 2023, 165, 112485 CAS.
- A. Kashima, Y. Inoue, S. Sugio, I. Maeda, T. Nose and Y. Shimohigashi, X-ray crystal structure of a dipeptide-chymotrypsin complex in an inhibitory interaction, Eur. J. Biochem., 1998, 255, 12–23 CAS.
- H. Xiong, R. R. Li, S. Y. Liu, F. X. Wu, W. C. Yang and G. F. Yang, Discovery of Specific Nonpeptide Probe for Chymotrypsin via Molecular Docking-Based Virtual Screening and the Application, ACS Appl. Bio Mater., 2018, 1, 310–317 CAS.
- G. Ren, H. Sun, J. Guo, J. Fan, G. Li and S. Xu, Molecular mechanism of the interaction between resveratrol and trypsin via spectroscopy and molecular docking, Food Funct., 2019, 10, 3291–3302 CAS.
- M. Samtiya, R. E. Aluko and T. Dhewa, Plant food anti-nutritional factors and their reduction strategies: an overview, Food Prod., Process. Nutr., 2020, 2, 6 Search PubMed.
- T. Fukutani, K. Miyazawa, S. Iwata and H. Satoh, G-RMSD: Root Mean Square Deviation Based Method for Three-Dimensional Molecular Similarity Determination, Bull. Chem. Soc. Jpn., 2020, 94, 655–665 Search PubMed.
- T. Bohn, F. Carriere, L. Day, A. Deglaire, L. Egger, D. Freitas, M. Golding, S. Le Feunteun, A. Macierzanka, O. Menard, B. Miralles, A. Moscovici, R. Portmann, I. Recio, D. Rémond, V. Santé-Lhoutelier, T. J. Wooster, U. Lesmes, A. R. Mackie and D. Dupont, Correlation between in vitro and in vivo data on food digestion. What can we predict with static in vitro digestion models?, Crit. Rev. Food Sci. Nutr., 2018, 58, 2239–2261 CrossRef CAS PubMed.
- K. Pälchen, D. Michels, D. Duijsens, S. Gwala, A. Pallares Pallares, M. Hendrickx, A. Van Loey and T. Grauwet, In vitro protein and starch digestion kinetics of individual chickpea cells: from static to more complex in vitro digestion approaches, Food Funct., 2021, 12, 7787–7804 RSC.
- M. Di Nunzio, C. Loffi, E. Chiarello, L. Dellafiora, G. Picone, G. Antonelli, C. Di Gregorio, F. Capozzi, T. Tedeschi, G. Galaverna and A. Bordoni, Impact of a Shorter Brine Soaking Time on Nutrient Bioaccessibility and Peptide Formation in 30-Months-Ripened Parmigiano Reggiano Cheese, Molecules, 2022, 27, 664 CrossRef CAS PubMed.
Footnote |
| † Both authors contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2025 |