
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceThe role of water in mediating DNA structures with epigenetic modifications, higher-order conformations and drug–DNA interactions
Roshan
Satange
a and
Ming-Hon
Hou
 *abc
*abc
aGraduate Institute of Genomics and Bioinformatics, National Chung Hsing University, Taichung, 402, Taiwan. E-mail: mhho@nchu.edu.tw; Fax: +886 4 2285 9329; Tel: +886 4 2284 0338 ext. 7011
bDoctoral Program in Medical Biotechnology, National Chung Hsing University, Taichung 402, Taiwan
cBiotechnology Center, National Chung Hsing University, Taichung 402, Taiwan
First published on 14th March 2025
Abstract
Water is arguably one of the most important chemicals essential for the functioning of biological molecules. In the context of DNA, it plays a crucial role in stabilizing and modulating its structure and function. The discovery of water-bound motifs in crystal structures has greatly improved our understanding of the interactions between structured water molecules and DNA. In this manuscript, we review the role of water in mediating biologically relevant DNA structures, in particular those arising from epigenetic modifications and higher-order structures such as G-quadruplexes and i-motifs. We also examine water-mediated interactions between DNA and various small molecules, including groove binders and intercalators, and emphasize their importance for DNA function and therapeutic development. Finally, we discuss recent advances in tools and techniques for predicting water interactions in nucleic acid structures. By offering a fresh perspective on the role of water, this review underscores its importance as a molecular modulator of DNA structure and function.
 Roshan Satange | Roshan Satange received his PhD and conducted postdoctoral research under Prof. Ming-Hon Hou at National Chung Hsing University, Taiwan. His research focuses on unusual nucleic acid structures and their implications in drug development for cancer and neurological diseases. With expertise in structural biology, drug design, and biophysical studies, he explores DNA and RNA-targeting therapeutics. |
 Ming-Hon Hou | Ming-Hon Hou received his PhD in Biochemical Sciences from National Taiwan University. He conducted postdoctoral research at Academia Sinica and continued his work in drug design and biophysics at National Chung Hsing University in Taiwan. Currently, he is a Distinguished Professor at the Graduate Institute of Genomics and Bioinformatics and serves as the Director of the Biotechnology Center at National Chung Hsing University. Additionally, he holds a joint appointment as an Adjunct Faculty member in the Molecular and Biological Agricultural Sciences program of the International Graduate Program at Academia Sinica, Taiwan. His research interests span various aspects of structural biology and drug development, including protein structures, unusual DNA structures, DNA–drug interactions, and protein–drug interactions. |
1. Introduction
Water exhibits unique properties that are essential for biological processes. Its polarity and ability to form hydrogen bonds make it a critical solvent. Despite its simple molecular structure, water's remarkable ability to form hydrogen bonds with other molecules is crucial to its life-supporting characteristics.1,2 In water, hydrogen bonds form when each partially positively charged hydrogen atom is covalently bonded to a partially negative oxygen atom in one water molecule while also being attracted to the oxygen atom of a neighboring water molecule (Fig. 1A).3 The hydrogen bonds in water molecules undergo continuous breaking and reformation. This dynamic behavior has been explained by the “jump model” mechanism.4,5 When a water hydroxyl group switches its hydrogen bond acceptor, it undergoes large-amplitude angular jumps (Fig. 1B). These jumps regulate the reorientation dynamics of water molecules, whether they are free or interacting with other molecules, such as ions, DNA, and proteins.6–8 This remarkable ability of water to form dynamic intermolecular hydrogen bonds, both within itself and with other molecules, is a key factor in enabling various biological functions. | ||
| Fig. 1 (A) Schematic representation of hydrogen bonding between water molecules. (B) Illustration of the jump model depicting the reorientation of a water molecule (water 1) that forms a hydrogen bond with a nearby water molecule (water 2) and simultaneously breaks the hydrogen bonds with other molecules, such as DNA and proteins. The figure is adapted with permission from J. Am. Chem. Soc., 2016, 138(24), 7610–7620. (C) Diagram showing ordered water molecules in the hydration shell interacting with DNA. A range of ordered water extending up to 18 Å is depicted. (D) The linear spine of hydration (red spheres) observed in the AT-rich region of a B-DNA (PDB ID: 4C64). Enlarged view shows an extensive interconnected network of hydrogen bonds formed by water molecules in the first hydration shell (red spheres) interacting with DNA bases, the backbone, and water molecules in the second hydration shell (green spheres). | ||
Of particular interest, water plays a crucial role in influencing the stability and functionality of biomolecules, including proteins and nucleic acids. At protein interfaces, water forms a solvation layer that modulates intermolecular interactions and functions.7,9,10 In the nucleosome, for example, water promotes interactions between histones and DNA, thereby stabilizing the entire structure.11 During DNA–protein recognition, water-mediated polar contacts contribute to binding specificity.12,13 Water at protein interfaces also assists in the binding of small molecules.14 Conserved water molecules in the vicinity of proteins have been shown to play a guiding role in the structure-based design of novel protease inhibitors, a promising approach in drug discovery.15 Moreover, water can act as a channel for proton transport and plays a crucial role in the catalytic functions of proteins.16 In addition to the more dynamic water molecules on the protein surface, Takano et al. demonstrated that water molecules within the interior cavities of proteins play a crucial role in enhancing structural stability by examining the crystal structures and conducting thermodynamic studies on a series of human lysozyme mutants.17 This internal water provides stability and ensures the correct folding and unfolding of proteins. The dynamic role of water in maintaining protein structures and functions has been the subject of numerous reviews.10,18–23
Similar to proteins, the role of water in DNA structures has long been the subject of research.24–28 Since DNA lacks a distinct interior hydrophobic surface like proteins, the majority of its atoms tend to interact with surrounding water molecules. DNA can transition between its B- and A-form conformations under varying hydration conditions.29–31 This reversible conformational shift alters the DNA backbone and the groove widths, exposing buried residues to form contacts with proteins and small molecule ligands. The formation of A-DNA is also important for cellular defense mechanisms under harsh conditions.32 Thus, solvation and desolvation in DNA not only maintain its structural integrity but also drive biochemical functions. Moreover, water plays an essential role in mediating interactions between DNA and small molecule ligands. Numerous studies highlighted the importance of water interactions in drug–DNA complexes.33–35 Unlike DNA, RNA typically exists as a single strand and can fold into complex three-dimensional structures necessary for its biological functions.36,37 Water is critical to this folding process.38 Water also mediates interactions between RNA and other molecules, improving the specificity and stability of RNA–protein and RNA–drug complexes.39–42 In catalytic RNA, water plays a key role by participating in enzymatic reactions as a proton donor or acceptor.43 Thus, water acts as an integral part of the molecular architecture in biological systems, essential for driving molecular interactions and biochemical functions.
Advances in structural biology have provided significant insights into the fundamental interactions between water and DNA. The increasing number of high-resolution structural motifs has yielded extensive data emphasizing the essential role of water in DNA. A detailed analysis of literature on water-mediated interactions within nucleic acids deepens our understanding on the role of water and its possible implications in nucleic acid functions and therapeutic development. In this review, we provide an overview of the importance of water interacting with nucleic acids, with a particular focus on its role in mediating biologically relevant DNA structures. Subsequently, we discuss water-mediated interactions between small molecules and DNA. We review advancements in new tools and techniques for predicting water interactions within nucleic acid structures. This review offers a new perspective on the critical role of water in molecular and chemical biology of DNA.
2. Ordered water molecules around DNA are essential for its structural stability and function
Water maintains the structural integrity of DNA by forming a robust, interconnected network of immobilized, ordered molecules known as the “hydration shell.” Hydration shells are essential for stabilizing double-helical structures and facilitating the formation of the grooves. The hydration shell consists of multiple layers of water molecules (Fig. 1C). Water molecules directly interacting with DNA up to ∼3 Å typically form the first hydration shell, establishing direct hydrogen bonds with phosphate groups, sugar moieties, and nitrogenous bases. Water molecules located ∼3–8 Å from the DNA surface constitute the second hydration shell; these are less tightly bound and act as a bridge between the first hydration shell and bulk water. Beyond the second hydration shell, water molecules gradually transition to bulk-like behavior while still experiencing some influence from DNA's electrostatic field.While the hydration shell around biomolecules has been studied for a long time,44,45 recent experimental and computational advances have uncovered new insights into the dynamics and structures of ordered water, which play a critical role in DNA conformation and functionality. For example, using a combination of dielectric relaxation, terahertz spectroscopy and all-atom molecular dynamics simulations, Singh et al. comprehensively analyzed the collective motions of hydrated DNA molecules.46 Their results demonstrated that the hydration structure extends as far as ∼18 Å from the DNA surface, with the intensity of water molecule interactions determined by their proximity to DNA; i.e., the water molecules exhibit different relaxation behavior depending on their interactions with the DNA. For example, water molecules closer to the minor grooves exhibit the slowest dynamics, followed by those interacting directly with the major grooves and phosphate groups. Meanwhile, water molecules farther from the DNA surface are more loosely bound and have faster dynamics.46 These observations imply that DNA influences water molecules beyond just its immediate surroundings, i.e., the first and second hydration shells. Interestingly, even such distant waters can interact with Watson–Crick base pairs but not with mismatched base pairs in DNA.47 These studies suggest that ordered water interactions with Watson–Crick base pairing are important for the long-distance recognition of DNA hybridization processes and that these interactions could be affected by environmental conditions, such as pH, salt concentration, and temperature.
The properties of water interacting with biological molecules in hydration shells differ significantly from those of free bulk water. Laage et al. compared and summarized these dynamic properties across various biomolecules, including DNA, proteins, and phospholipids.48 Briefly, unlike free bulk water, the hydration shell surrounding DNA consists of strongly bound water, an increased number of hydrogen bonds, and a higher degree of molecular ordering. These water molecules often act as bridges, mediating both intermolecular and intramolecular interactions within DNA or between DNA and other biomolecules. One of the most prominent features of water–DNA interactions is the “spine of hydration” in the minor groove (Fig. 1D), first reported in the crystal structure of the B-DNA dodecamer, d(CGCGAATTCGCG).49 The water molecules in the hydration spine form a complex, interconnected network involving the first- and second-shell water molecules and their interactions with the base pairs lining the groove. These water sites are interdependent, exhibit cooperative behavior, and are less affected by modifications in DNA bases.50,51 Notably, the hydration spine can also form in solution under ambient conditions.52,53 These water interactions are crucial for maintaining integrity, stability, and functionality of DNA. In the following sections, we will explore the interactions between water and both canonical and non-canonical DNA structures, including those associated with epigenetic modifications, gene regulation, and disease-related conformations. Using examples primarily from X-ray crystallography studies, we will highlight these interactions and their critical roles in regulating DNA structure and function.
3. Water interactions in DNA structures
3.1. Water-mediated interactions in canonical DNA structures with epigenetic modifications
The B-form of the DNA duplex is the most biologically relevant structure within the genome, featuring a well-characterized network of water molecules. In canonical structures including B-DNA, water molecules tend to form hydrogen-bonded interactions with base pairs, backbone, and the grooves.26,54–56 The spine of hydration is a well-studied hydration aspect in B-DNA. Particularly in A–T rich regions, water in the minor groove creates the spine of hydration due to the narrow width of the groove, which facilitates close water-mediated interactions.57 On the other hand, in the A-DNA conformation, water molecules exhibit more interactions with the interstrand phosphate oxygen atoms. Phosphate groups in A-DNA are positioned close enough for water molecules to bridge adjacent phosphates via their free oxygen atoms—a configuration not possible in B-DNA due to the larger spacing between phosphates.26 Using in silico sampling methods and an in-house-developed thermodynamic parameter analysis, Ghosh et al. studied the dynamic and thermodynamic responses of local hydration in a variety of base pair sequences. They demonstrated that the structure, sequence, and composition of the DNA groove influence local hydration characteristics during the B-to-A transition, a process crucial for biological functions.58In addition to interacting with DNA grooves and the backbone, water-mediated interactions with base pairs play a crucial role in maintaining DNA structure. The guanine base can form hydrogen bonds with water molecules using the 2-amino and N3 atoms in the minor groove and the 6-keto group and N7 atoms in the major groove.59 Cytosine can form hydrogen-bonded interactions with water using the 2-keto group from the minor groove and the 4-amino group from the major groove. Similar to guanine, adenine can use the 6-amino group from the major groove and the free ring nitrogen atoms N3 and N7 in the minor groove to form water-mediated hydrogen bonds. Thymine forms hydrogen-bonded interactions with water using the 2-keto and 4-keto groups from the minor and major grooves, respectively. Interestingly, crystallographic analysis of the hydration sites around DNA bases showed that these hydration sites in purine bases are planar to the plane of the nucleobases and exhibit a consistent pattern, whereas those around pyrimidine bases are non-planar.60 While the positions of hydration sites around DNA bases are generally consistent, local deformations in DNA can alter the distribution of ordered water molecules surrounding the base pairs. This redistribution of water is crucial for DNA repair proteins to distinguish canonical Watson–Crick base pairs from mismatched ones. For example, the formation of a semi-open conformation of the U:G mismatch, resulted in the reordering of water molecules around uracil in such a way that water acts as a hydrogen bond donor.61 This contrasts with Watson–Crick C:G base pairing, which usually adopts a closed conformation, where water acts as a hydrogen bond acceptor to the cytosine and guanine bases. These differences in water interactions between mismatched and Watson–Crick base pairs create differences in electrostatic fields, which are likely to affect the base pair discrimination ability of DNA repair proteins.
DNA methylation is essential for regulating gene expression and maintaining genome stability.62 In an all-atom molecular dynamics simulation study, Teng et al. found that cytosine methylation reorganizes the arrangement of water molecules around DNA bases and strengthens their interactions with water.63 Recently, Lin et al. reported the crystal structures of several native and ligand-bound methylated DNA sequences using X-ray crystallography.64 Analysis of these structures revealed that cytosine methylation at non-CpG sites in native DNA can promote the formation of water-mediated Hoogsteen base pairs in methylated bases (Fig. 2A). The propensity for such water-mediated base pairing increases upon ligand binding. These findings were further corroborated in solution using NMR spectroscopy. This suggests that the positioning of water and its interactions in methylated base pairs are critical for recognition by external ligands. Thus, these types of water-mediated interactions might play an important role in the recognition of methylated bases by regulatory proteins.
 | ||
| Fig. 2 Water-mediated interactions in epigenetically modified DNA structures. (A) Induction of water-mediated Hoogsteen base pairing upon cytosine methylation in unliganded native DNA (PDB ID: 8XPA) and upon ligand binding (PDB ID: 8XP8). (B) Water-mediated interactions in 5-hydroxymethylcytosine (5hmC)-modified DNA in the A-form conformation (PDB ID: 5CH0) and in the B-form conformation (PDB ID: 5CJY). (C) Views of the major and minor groove of DNA with 5-formylcytosine (5fC) modification (PDB ID: 4QKK). A network of water in the major groove is shown, with the water molecules shown as red spheres and the modified 5-formyl group at the cytosine bases highlighted in green. On the right, an enlarged view of unusual Watson–Crick base pairing is shown. | ||
Using X-ray diffraction, Renciuk et al. solved a series of crystal structures of B-DNA containing the epigenetic modifications 5-hydroxymethylcytosine or 5-methylcytosine.65 Based on the analysis of these structures, they proposed that replacing a hydrophobic methyl group with a hydrophilic hydroxymethyl group in epigenetically modified cytosine creates a conserved network of water molecules that interact with the O5 atom of 5-hydroxymethylcytosine bases. The O5 atom of hydroxymethyl groups forms bifurcated hydrogen bonds with water molecules in the vicinity of approximately 3 Å. Similar water-mediated interactions were also observed in an independent study on 5-hydroxymethylation in B-DNA.66 In another X-ray crystallography study, Wang et al. found that, regardless of DNA conformations, 5-hydroxymethylcytosine forms strong water-bridged hydrogen bonds with neighboring bases that stabilizes overall modified structures (Fig. 2B).67
Given the fact that hydroxymethylation of cytosine is an intermediate step in the methylation and demethylation processes, and it also plays a significant role in gene regulation, pluripotency, and development,68,69 the occurrence of a conserved hydration network is supposed to have an active role in epigenetic regulation. Water-bridging interactions have also been observed in another cytosine modification, formylcytosine. In the crystal structure of DNA containing 5-formylcytosine, a water network linking the formyl group of modified cytosine to the phosphate backbone and adjacent nucleobases is identified (Fig. 2C).70 This resulted in alteration in Watson–Crick base pairing, creating an unusual geometry in which two water molecules bridge the formyl group of cytosine with its complementary guanine. In addition, the extensive hydration of modified bases also plays a crucial role in their interactions with DNA-processing enzymes. For example, in the crystal structures of ten-eleven translocation (TET) proteins complexed with hydroxymethylcytosine- and formylcytosine-containing DNA, Hu et al. reported water-mediated hydrogen bonded interactions between the exocyclic amino (N4) nitrogen of the modified bases and threonine (T1393) residue of TET2 proteins suggesting a regulatory role of water in epigenetically modified DNA and its functions.71
3.2. Water-mediated interactions in non-canonical higher-order DNA structures
In non-canonical DNA structures, such as G-quadruplexes and i-motifs, the role of water becomes even more pronounced. These alternative conformations are essential not only for regulating gene expression processes, but also have significant potential as drug targets.72–74 In conventional G-quadruplexes, water often forms a clustered network around the four grooves, which results in an increased complexity of the grooves and the overall quadruplex diversity.75–77 The presence of water molecules around different G-quadruplex topologies is important for the stabilization of overall quadruplex arrangements by small molecule ligands.78–80 In recent years, a novel topology of G-quadruplexes has been identified, called left-handed G-quadruplexes.81–83 These left-handed structures are characterized by a single, circular groove. The arrangement of water molecules around both right- and left-handed G-quadruplex structures has recently been surveyed using crystallographic analysis (Fig. 3A and B).84 Interestingly, there is significant distinction between the hydration pattern of these two conformations. In right-handed helical G-quadruplexes, the hydration tends to follow a linear pattern, similar to that found in right-handed DNA duplexes. In contrast, the left-handed G-quadruplexes display a unique, circular hydration arrangement within a single groove, termed the “wheel of hydration”. Although biological implications of left-handed G-quadruplex structures are still unclear, the distinct and consistent hydration pattern associated with these structures can be considered as their integral component which may play a crucial role in their functions.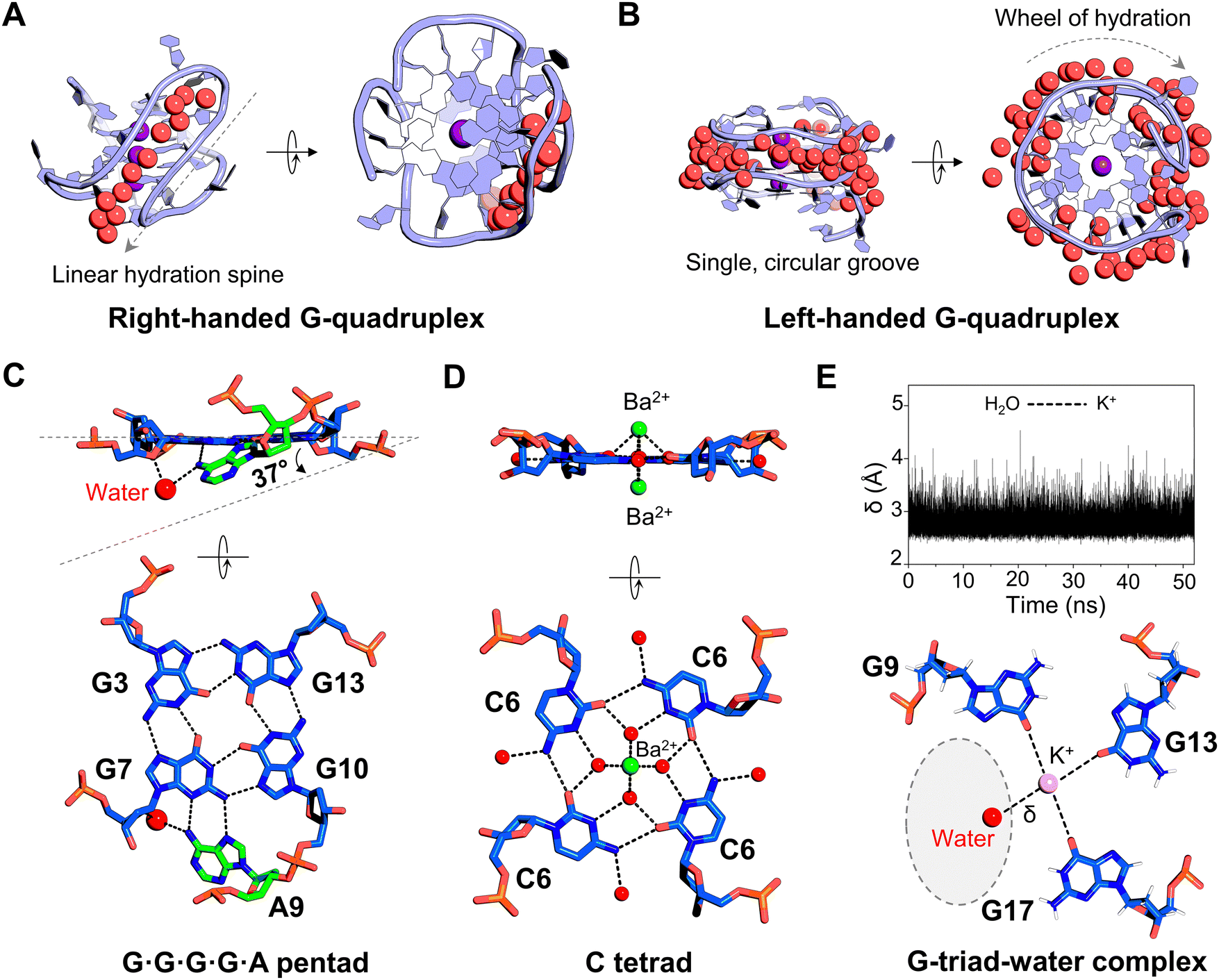 | ||
| Fig. 3 Water interactions in DNA G-quadruplexes. (A) Representative view of a narrow groove in a right-handed G-quadruplex structure, illustrating a linear arrangement of water molecules (PDB ID: 6JKN). (B) View of a single circular groove with water molecules in a left-handed G-quadruplex structure (PDB ID: 4U5M). (C) Formation of a water-mediated pentad base pair arrangement in the G-quadruplex structure of an HIV-1 integrase aptamer (PDB ID: 7XDH). (D) Water-mediated cytosine tetrad (C-tetrad) surrounding the central Ba2+ ion channel (green sphere) in the G-quadruplex structure (PDB ID: 4U92). (E) Formation of a G-triad–water complex in the G-quadruplex structure. Top: The distance (δ) between water molecules and K+ ions (pink sphere) during the simulation. Bottom: The orientation of three guanine bases and their interactions with water molecules. A dashed oval highlights a potential region for the binding of an external ligand. The figure is adapted with permission from Nucleic Acids Res., 2016, 44(2), 910–916. In all panels, the water molecules are shown as red spheres and the hydrogen bonds as dashed black lines. | ||
In addition to the grooves, water molecules also mediate planar arrangements of the bases, which is essential for the integrity and structural framework of G-quadruplex structures. For example, in an interlocked G-quadruplex structure, a conserved water-mediated pentad base-pair arrangement forms between four planar guanines and ca. 37° tilted adenine (Fig. 3C).85 This water-mediated pairing is crucial for interactions and functions of this G-quadruplex as an HIV-1 integrase inhibitor. Zhang et al. studied a biologically relevant G-quadruplex structure induced by the incorporation of a nucleoside analog, cyanovinylcarbazole, into DNA.86 They solved the 1.50 Å crystal structure of the DNA decamer containing cyanovinylcarbazole modification. This G-quadruplex structure formed a planar arrangement of four cytosines (called a C-tetrad) with water molecules mediating contacts between a divalent cation and the C-tetrad (Fig. 3D). These water molecules can donate protons to nearby cytosine bases, which in turn increases the electron density of the lone pair on water oxygen atoms. Consequently, the water oxygen can shield the charges between adjacent divalent metal ions. Thus, the presence of water has a favourable effect on stabilizing the unusual C-tetrad pairing and in forming continuous stacking interactions of the other guanine tetrads, thereby maintaining the overall integrity of the G-quadruplex structure.86 Water-mediated tetrad base pairs are also found in G-quadruplex RNA structures.87 Using NMR spectroscopy and molecular dynamics simulations, Heddi et al. demonstrated that in parallel G-quadruplexes, if one of the guanines is missing from guanine tetrads, water molecules can mimic the position of the missing guanine and form a G-triad–water complex at that site (Fig. 3E).88 These water-mediated sites can serve as binding sites for various metabolites, with implications in metabolite sensing, regulation and drug development. In an antiparallel human-telomeric G-quadruplex structure, water-mediated intra-strand interactions between two thymine bases were found to enhance stacking on the G-tetrad, further contributing to the stability of the quadruplex.89 Thus, in addition to stabilizing the grooves, water also aids in interactions with base-pair arrangements in quadruplexes and facilitates the formation of unique structural motifs, which can further influence ligand binding by stabilizing DNA conformations, enhancing specificity, and modulating binding affinities.
Similar to G-quadruplexes, i-motifs, another type of four-stranded non-canonical structure, can form in cytosine-rich sequences in the regulatory regions of the human genome.90–92 In the formation of i-motif structures, water plays an important role. It has been shown that variations in hydration conditions can regulate the folding and unfolding of i-motif structures.93 Although fewer i-motif structures have been reported compared to G-quadruplexes, the available structures provide valuable insights into the potential role of water in stabilizing and influencing the i-motif structures. The i-motifs are characterized by two wide grooves and two narrow grooves, with the majority of water molecules concentrated in the wider grooves, as the narrow grooves are too restricted to accommodate water molecules.94–96 In i-motif structures, water-mediated bridge interactions typically occur between phosphate oxygens and the N4 amino group of cytosine, playing a crucial role in stabilizing these structures. In a recently reported i-motif structure derived from the HRAS oncogene sequence, a robust network of water molecules is observed within the wider groove (Fig. 4A).97 This water network forms hydrogen bonds with the phosphate backbone, with N4 atoms in cytosine bases, or with other water molecules, similar to previous i-motif structures. In addition to these interactions, the current structure displays two high-electron density water positions: the first coordinated with two extrahelical guanine bases and another water molecule, while the second is located in the loop region of the i-motif. A similar water position is present in the loop of another HRAS i-motif solved in the same study, highlighting the possibility that water in these regions is conserved in HRAS i-motifs which might have important functional roles. Another biologically relevant intramolecular i-motif structure from the promoter region of the insulin gene has been recently reported.98 In this structure, water molecules formed bridging hydrogen-bonded interactions with the N4 atoms of cytosine bases and phosphate oxygens. Several water molecules were observed in the loop region of the i-motif, indicating potential sites for hydrogen-bonding. Such water-bound structure motifs can also provide crucial information for designing gene-selective ligands.
 | ||
| Fig. 4 Water interactions in DNA i-motifs. (A) Overall structure of the HRAS oncogene i-motif (PDB ID: 8CXF). On the right, detailed views of water-mediated interactions in the loop and core regions are shown. (B) Inter-strand coordination of water molecules in the i-motif structure of CCG repeats (PDB ID: 4PZQ). In the figure, the two strands are colored yellow and blue. In all panels, water molecules are shown as red spheres. | ||
The structures of i-motifs associated with specific tandem repeat sequences have been studied, revealing several important water-mediated interactions. Chen et al. reported the crystal structure of an i-motif formed from the CCG repeat sequence, which is linked to fragile X syndrome.99 This structure features a network of bridging water molecules mediating inter-strand interactions within the i-motif. In this structure, water molecules bridge interactions between two strands of the asymmetric units and stabilize the i-motif core (Fig. 4B). A similar water interaction network is found in another CCG-repeat-related i-motif, suggesting its importance in maintaining the i-motif conformation.100 This conserved network of water further plays an important role in stabilizing the base-extruded duplex induced by a small-molecule ligand.100 Sato et al. reported a higher-order structure formed by d(GCGA[A]1AGC), which is associated with d(GAAA)n tandem repeats in the non-coding region near telomeres.101 This structure adopts an octaplex conformation, incorporating an i-motif with water-mediated adenine quartets (A-quartets). Based on the analysis of this structure and the other octaplex structure formed by the d(GCGA[G]1AGC) repeat sequence,102 it is clear that the water-mediated hydrogen bonding network is critical in stabilizing the octaplex conformation. They further hypothesized a potential mechanism for sequence slippage during DNA replication in d(GAAA)n tandem repeats. The presence of water-mediated A-quartet structures likely stabilizes transient misaligned base-pairing events and facilitates structural rearrangements during slippage. Understanding these intricate structural details could facilitate therapeutic intervention. Since d(GAAA)n repeat expansion has been linked to approximately 34% of renal cell carcinomas,103,104 small molecules that selectively stabilize or destabilize this structure could modulate replication-associated instability, potentially offering a novel strategy for cancer treatment. Targeting the water-mediated hydrogen bonding network could allow for the rational design of drugs that either prevent pathological repeat expansion or induce structural changes that disrupt oncogenic pathways.
4. Water interactions in small molecule–DNA complexes
Water is crucial for the formation of complexes between small molecules and DNA. Its ability to form hydrogen bonds and modulate the electrostatic and structural environment of DNA binding sites has established water molecules as a key focus in drug–DNA binding mechanisms.105–107 In this section, we will examine the role of water in mediating small molecule–DNA interactions and their significance in influencing DNA structure and function.4.1. Water-mediated interactions between DNA and groove-binding agents
DNA targeting molecules typically bind DNA through two major modes: minor groove binding and intercalation.108,109 Structured water molecules are critical in mediating interactions between small molecule ligands and the DNA minor groove, improving binding affinity and biological activity.35 DNA groove-binding agents like netropsin and distamycin interact with the minor groove by displacing ordered water molecules to achieve strong binding.110 However, these molecules exhibit different binding affinities due to their distinct interactions with water.111 Distamycin, being more hydrophobic, interacts with DNA-associated water to a lesser extent than netropsin. This resulted into a more pronounced loss of distamycin mobility upon DNA binding which led to its more unfavourable entropy to the binding free energy and consequently to lower affinity.111 These examples explain how water-mediated interactions modulate the binding specificity and affinity of small molecule compounds to DNA, with implications for their biological activity.Moreover, groove-binding agents usually exhibit either shape-specific or sequence-specific interactions with DNA. In shape-specific binding, the molecules often achieve strong interactions by being isohelical, i.e. they complement the shape of the minor groove. However, even non-isohelical molecules can achieve strong minor groove binding through water-mediated interactions. For example, the non-isohelical linear heterocyclic compounds DB921, DB1055, and CGP 40215A utilize water molecules to establish hydrogen bonds that compensate for their lack of curved shape (Fig. 5A).112–115 In a X-ray crystallography-based study, Wei et al. showed that in addition to the well-reported spine of hydration in the minor groove, a conserved network of 11 ordered water molecules is present within the minor groove (Fig. 5B).116 This water-network mediates non-covalent interactions with linear heterocyclic groove-binding agents. The water-mediated interactions complete the molecular curvature required for high-affinity binding to DNA. These findings emphasize the significance of water in stabilizing the linear geometry containing small molecules in interacting with the DNA minor groove.
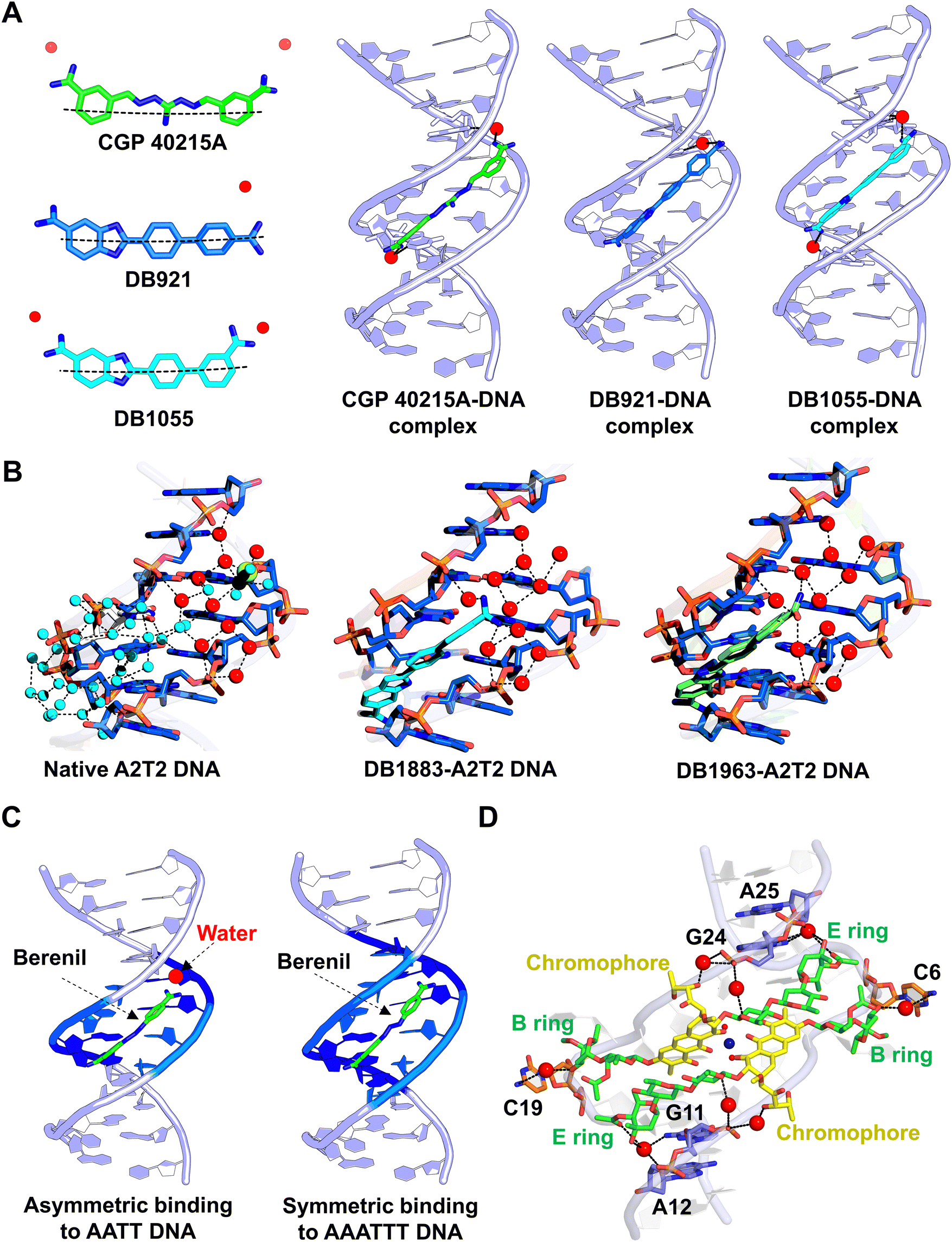 | ||
| Fig. 5 Water interactions in DNA groove binders. (A) Water-mediated binding of the linear-shaped groove binders CGP 4-215A (PDB ID: 1M6F), DB921 (PDB ID: 2B0K), and DB1055 (PDB ID: 2I5A) to DNA. The water molecules are shown as red spheres. (B) Water cluster with water molecules of the first shell (cyan spheres) and a network of 11 conserved water molecules (red spheres) in native DNA (PDB ID: 3U2N). After binding of the ligands DB1883 (PDB ID: 3U0U) and DB1963 (PDB ID: 3U08), the conserved water cluster contributes to the stabilization of the ligand–DNA complex. (C) Binding of berenil to DNA in the asymmetric mode (PDB ID: 2DBE) involves water-mediated interactions, whereas water-mediated stabilization is not required for symmetric binding (PDB ID: 1D63). (D) Water-mediated bridging interactions stabilize the flipped-out residues in chromomycin-induced CCG repeat DNA (PDB ID: 5XEW). The water molecules are shown as red spheres. | ||
Pyrrole-imidazole (Py-Im) polyamides are a class of synthetic compounds designed to target the DNA minor groove.117,118 These compounds are excellent examples of shape complementarity, as they can recognize specific DNA sequences by matching the minor groove curvature. Chenoweth and Dervan elucidated the crystal structure of an eight-ring cyclic Py-Im polyamide bound to DNA, showing the compound bridge interactions between adenine and guanine base pairs via water-mediated hydrogen bonds.119 In another X-ray crystallographic study, Oh et al. demonstrated how hairpin Py-Im polyamides block transcription by trapping DNA and RNA polymerase II.120 In this structure, two ordered water molecules were found to mediate hydrogen bonding interactions between the polyamide and the DNA minor groove, underscoring the role of water in driving the biological activity of Py-Im minor groove binding ligands.
Structured water molecules also play a critical role enabling the sequence specificity to the DNA minor groove binders. Many X-ray crystallographic studies have been reported on sequence-specific groove binders, highlighting how water molecules bridge the interactions between DNA and groove-binding agents. For example, the antitrypanosomal agent berenil has been shown to bind asymmetrically to AT-rich DNA sequences (–AATT– sequence) through water-mediated bridging interactions (Fig. 5C).121 In the asymmetric binding mode, one of the amidinium groups of the berenil forms a hydrogen-bonded interaction with the N3 atom of the adenine base, while the other amidinium group does not directly interact with the complementary strand adenine in DNA. Instead, the amidinium nitrogen atom interacts with the 5′ end of the adenine base through a water-mediated interaction. Interestingly, another independent crystal structure showed that when berenil binds symmetrically to another AT-rich sequence (–AAATTT–), the water mediation is not required.122 In the symmetric binding mode, the two amidinium groups of berenil interact directly with two adenine bases on each strand through hydrogen bonding, without requiring a bridging water molecule. These studies demonstrate that water-mediated interactions can vary depending on the DNA sequence and the drug binding orientation. Similarly, propamidine, another minor groove binder with a longer linker than berenil, relies on water-mediated interactions to recognize AT-rich sequences.123,124 Recent studies have shed light on the water-mediated interactions between the minor groove binders and biologically important DNA sequences. For example, the X-ray diffraction structure of diamidinobenzimidazole compounds shows that they selectively bind to AT-rich DNA sequences from the promoter regions of human genes.125 These compounds adopt two possible binding orientations, with a preference for water-assisted hydrogen bonding at their cationic ends.125 Using long-timescale molecular dynamics simulations, Carter et al. demonstrated the structural basis of a heterocyclic diamidine, DB2277, which exhibits a preference for binding to GC sites in DNA.126 This minor-groove binder displaces the ordered water molecules from the groove and forms several water-mediated contacts with the bases and the backbone of the DNA. These studies suggest the potential for designing a new class of minor groove binders that incorporate bound water molecules into their DNA complexes.
Moreover, water-mediated interactions are crucial for achieving optimal binding of some groove-binding antibiotics. For example, antibiotics such as chromomycin and mithramycin often adopt an octahedrally coordinated, metal-chelated dimer conformation to bind DNA.127,128 In these complexes, water molecules serve as the fifth and sixth ligands to complete the coordination. In addition, the other bridging water molecules play a key role in mediating the interactions between the drugs and disease-associated DNA structures. For example, the binding of metal-chromomycin A3 complexes to the CCG repeat DNA associated with fragile X syndrome induces large-scale DNA deformations, including the extrusion of cytosine bases and distortions in the backbone.100,129 In this unusual DNA conformation, structured water molecules stabilize the interactions between the extruded bases and drug moieties (Fig. 5D). The N4 amine atom on the extruded cytosines forms water-mediated hydrogen bonds with the O3 oxygen atoms of the disaccharide B ring of chromomycin, while the N2 atoms of the terminal guanines form water-mediated hydrogen bonds with the O3 oxygen atoms of the trisaccharide E ring. Two bridging water molecules further mediate interactions between the O1 oxygen atoms of the trisaccharide C ring of chromomycin and guanine bases. These water-mediated interactions contribute to the specific recognition of (CCG)n repeats by chromomycin complexes. Furthermore, in a G-G mismatched DNA structure stabilized by chromomycin A3, water molecules are shown to stabilize an unusual anti–syn geometry.130 The interaction of water with mismatched bases pushes the guanines toward the exterior of the helix, creating a single hydrogen bond between them. The stability of this more distant anti–syn pairing is compensated by water-mediated interactions between the mismatched bases. The study further demonstrated that this unusual water-mediated mismatch geometry serves as a hotspot for G–G mismatch recognition by another DNA-intercalating drug. Thus, water-mediated interactions are essential for stabilizing antibiotic–DNA complexes by completing metal coordination, bridging drug and DNA interactions, and facilitating the specific recognition of disease-associated DNA structures, such as tandem repeats and mismatches, through hydrogen bonding networks that induce large-scale conformational changes and structural stabilization.
4.2. Water-mediated interactions between DNA and intercalating agents
DNA intercalation is another important binding mode for many DNA-targeting agents, in which intercalators insert their planar aromatic ring structures between DNA base pairs.131 Unlike groove binders, the role of water in DNA intercalation remains less well-defined, possibly because intercalators primarily rely on stacking interactions rather than direct hydrogen bonding with DNA. However, studies have revealed a significant relationship between water and intercalating agents, emphasizing their unique roles in the functionality and structural stability of DNA. For example, ruthenium-based compounds exhibit photophysical properties upon DNA intercalation.132–134 These compounds interact with DNA through the photooxidation of guanine bases and also display luminescence properties.135,136 Hall et al. collected seven independent X-ray data sets from a single DNA crystal at room temperature under varying relative humidity conditions to understand the effects of dehydration on DNA structures.33 Their results demonstrated that the controlled dehydration and rehydration of a complex containing the ruthenium compound [Ru(TAP)2(dppz)]2+ (TAP = 1,4,5,8-tetraazaphenanthrene, dppz = dipyrido[3,2-a:2′,3′-c]phenazine), intercalated in DNA, induces reversible DNA kinking and a B-to-A transition (Fig. 6A). In its hydrated form, the complex structure shows a single water-mediated bridging interaction. Upon dehydration, however, three water-mediated interstrand cross-links form per Ba2+ ion. This enhanced water network arises from the adoption of an A-DNA-like conformation and a kink induced by the ligand intercalation. These findings show promise for the application of water-controlled ruthenium intercalators in DNA self-assembly and molecular engineering. Moreover, the chemical properties of ruthenium compounds are influenced by the orientation of their dppz moiety and its interactions with surrounding water molecules.137–139 When the dppz ligand is intercalated deeply within DNA base pairs, its nitrogen atoms become inaccessible for interaction with water. This phenomenon can be exemplified by the crystal structure of [Ru(phen)2(dppz)]2+ (phen = phenanthroline) in complex with the d(ATGCAT)2 DNA duplex.139 In this structure, the two enantiomers, Δ and Λ, of [Ru(phen)2(dppz)]2+ exhibit different orientations when simultaneously intercalated into DNA. The distinct binding modes of the two enantiomers result in the formation of differently structured water monolayers. Upon comparing these two structural halves, it was found that the Δ enantiomer intercalates at an angled orientation, exposing only one dppz nitrogen atom to mobile water molecules (Fig. 6B). In contrast, the perpendicular intercalation of the Λ enantiomer allows mobile water to access both phenazine nitrogen atoms. Since water access to the dppz phenazine nitrogen determines the photochemical lifetime of ruthenium complexes, luminescence measurements show that the Λ enantiomer exhibits lower emission intensity upon DNA interaction. This highlights the critical role of water in modulating the function of ruthenium-based DNA intercalating agents. | ||
| Fig. 6 Water-mediated interactions in DNA intercalators. (A) Controlled hydration and dehydration of the d(TCGGCGCCGA) duplex in the presence of Λ-[Ru(TAP)2(dppz)]2+ (TAP = 1,4,5,8-tetraazaphenanthrene; dppz = dipyrido[3,2-a:2′,3′-c]phenazine) induce conformational changes in DNA (PDB IDs: 4LTF and 4LTG). The enlarged view shows the water-mediated cross-linking between the strands in the major groove. (B) A water network formed by ordered water molecules in the major groove of the d(ATGCAT)2 DNA duplex containing Δ- and Λ-enantiomers of rac-[Ru(phen)2dppz]2+ (phen = phenanthroline; dppz = dipyridophenazine) (PDB ID: 4JD8). (C) Water-mediated hydrogen bonding stabilizes flipped-out guanine residues and thus contributes to the overall stability of actinomycin D bound to the DNA duplex (PDB ID: 6J0H). (D) Water-mediated stabilization of central core residues in the DNA structure induced by the triamino–triazine–acridine conjugate Z1 (PDB ID: 6M4T). In all panels, the water molecules are shown as red spheres and the hydrogen bonds as dashed black lines. | ||
Intercalators often induce stronger structural changes in DNA compared to groove binders.140–142 These changes result in the formation of unusual structural motifs, such as a kinked backbone or base pair flipping. Base pair flipping occurs when an intercalator inserts its planar rings between DNA base pairs, causing the nucleobases to rotate about their glycosidic bonds and protrude out of the helical axis.138 This disruption in base stacking and hydrogen bonding interactions exposes the flipped-out bases to the external environment, including proteins or solvent molecules. Water-mediated interactions play a crucial role in stabilizing such unusual structures. For example, the binding of actinomycin D to G–G mismatch in DNA induces a kinked right-handed Z–DNA conformation, in which the guanine bases are flipped out of the duplex.130 In this structure, a cluster of water molecules plays a key structural role in stabilizing the kinked structure with the flipped-out guanines (Fig. 6C). This water cluster holds and stabilizes the entire kinked DNA backbone structure toward the major groove side. These water-mediated stabilizing interactions of flipped bases are also observed in other drug–DNA complexes and protein–DNA complexes.129,143 Moreover, in an X-ray crystallography structure, Chien et al. reported the formation of a U-shaped head-to-head four-way junction structure upon intercalation by the triaminotriazine–acridine conjugate Z1 (Fig. 6D).144 In this highly hydrated complex structure, water molecules mediate significant interactions to stabilize the residues at the central crossover site of the junction conformation. In addition, three conserved water molecules further stabilize the Z1–DNA complex by directly mediating interactions between the ligand Z1 and DNA bases. In a recent structure of a DNA side-by-side junction formed due to the intercalation of diaminoacridine bis-intercalator DA5, a specific water-mediated interaction at the inter-duplex contact has been observed.145 In addition to bridging direct interactions between intercalators and DNA, water often supports intercalator–DNA complexes through several indirect interactions. For example, in the structure of echinomycin bound to T–T mismatched DNA, Wu et al. showed water-mediated stabilization of a drug–drug interaction and the mismatch.146 Water-mediated mismatch stabilization has also been observed in other intercalators and DNA structures.147 In general, intercalating agents induce significant structural changes in DNA, often leading to unusual motifs stabilized by water-mediated interactions, such as kinked Z–DNA conformations, four-way junctions, and inter-duplex contacts. The water interactions not only support ligand–DNA interactions but also facilitate cooperative drug binding, enhancing structural integrity and biological activity.
4.3. Water-mediated interactions between DNA G-quadruplexes and small molecule ligands
DNA G-quadruplexes are increasingly recognized as important molecular targets for small molecule compounds in drug development for treating cancer and neurological diseases.148–150 Studies on G-quadruplexes and their interactions with small molecules have identified several common features of quadruplex binders that are essential for defining their binding mechanisms. In general, G-quadruplex-binding ligands possess planar heteroaromatic groups which enabled them to stack effectively on G-quartet surfaces through π–π interactions. Additionally, these ligands often feature cationic side chains that interact with the DNA grooves and bases. Structural studies by Neidle and colleagues have highlighted the critical role of water-mediated interactions in G-quadruplex-ligand binding.151–157 Water molecules usually occupy conserved sites near the ligand-binding region and the grooves in quadruplex structures. These water molecules form hydrogen bonds with both the ligand and the DNA. The crystal structure of an acridine-based ligand, BSU6039, in complex with a telomeric G-quadruplex, revealed that water molecules from the spine of hydration in the groove interact with the protonated nitrogen atom of the pyrrolidino group and the amide nitrogen of BSU6039.151 Another well-studied quadruplex binder, BRACO-19, also employs extensive water-mediated hydrogen bonding interactions to stabilize the human G-quadruplex structure (Fig. 7A).153 Naphthalene diimide derivatives show conserved water-mediated interactions, where water molecules bridge DNA phosphate groups and the nitrogen atom of the N-methyl-piperazine ring of the ligands.155,156 Natural compounds such as berberine have shown potential for binding to G-quadruplex structures. In the crystal structure with a 2![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 berberine-to-G-quadruplex molar ratio, a water molecule occupies the central channel, mediating a conventional hydrogen bond with the carbonyl oxygen of guanine and a CH⋯O interaction with berberine (Fig. 7B).158 This specific interaction stabilizes the orientation of berberine molecules within the G-quadruplex structure. Similarly, a ruthenium polypyridyl complex has been shown to bind to a unimolecular chair G-quadruplex conformation. In this, extensive structured water molecules form a network that stabilizes the compact globular ligand-G-quadruplex structure.159
:1 berberine-to-G-quadruplex molar ratio, a water molecule occupies the central channel, mediating a conventional hydrogen bond with the carbonyl oxygen of guanine and a CH⋯O interaction with berberine (Fig. 7B).158 This specific interaction stabilizes the orientation of berberine molecules within the G-quadruplex structure. Similarly, a ruthenium polypyridyl complex has been shown to bind to a unimolecular chair G-quadruplex conformation. In this, extensive structured water molecules form a network that stabilizes the compact globular ligand-G-quadruplex structure.159
 | ||
| Fig. 7 Water-mediated interactions between small molecule ligands and G-quadruplexes. (A) Binding of the experimental anticancer drug BRACO-19 to human telomeric G-quadruplex DNA involves water-mediated interactions that stabilize the ligand-DNA complex (PDB ID: 3CE5). (B) In the berberine G-quadruplex complex, a water molecule at the 3′-end of the binding site forms a hydrogen bond that establishes a critical interaction between the drug and DNA (PDB ID: 3R6R). | ||
In an analysis of X-ray crystal structures of G-quadruplexes in both native and ligand-bound states, Neidle observed clusters of conserved water molecules.34 These water molecules serve as platforms for ligand binding and are crucial for establishing the structure–activity relationship of G-quadruplex-targeting ligands. This analysis led to an important conclusion about G-quadruplex binders: when identifying potential G-quadruplex binding ligands using high-throughput computational methods, it is imperative to account for solvent-mediated interactions in G-quadruplex binding. Neglecting water-mediated interactions increases the risk of inaccurate predictions of low-energy ligand-binding states and their interactions. Incorporating solvent-mediated considerations can not only enhances the binding affinity of these ligands but also improves their selectivity for G-quadruplexes over canonical duplex DNA structures.
5. Emerging approaches for predicting water interactions with nucleic acids
A large number of techniques have been used to study the dynamic interactions between water and nucleic acids.53,160–163 Structural techniques, including X-ray and neutron crystallography, remain the gold standards for studying these interactions, providing detailed spatial and dynamic insights into the water shell surrounding nucleic acid surfaces. While these methods yield the most valuable structural information, they have limitations in fully capturing the solvent and nucleic acid interactions. For example, resolution constraints or incomplete electron density in crystallographic structures can prevent the precise localization of water near biomolecules. Moreover, hydration networks and water positioning are significantly affected by artifacts introduced during the cryo-cooling process in biomolecular crystallography.164,165 For instance, ultrahigh-resolution structures of the protein crambin have been reported at various temperatures ranging from 100 K to 293 K.166 Teeter et al. observed that at temperatures above 200 K, the extensive hydrogen-bonding network of water molecules, identified as crystal packing artifacts, was disrupted.166 To mitigate these artifacts and obtain more detailed insights into atomic motions and solvent interactions, room-temperature X-ray crystallography has been proposed.167 However, despite its advantages, room-temperature data collection remains relatively uncommon, with cryogenically collected structures dominating the Protein Data Bank. These challenges can impede a comprehensive understanding of the role of water in nucleic acids. With the growing repository of high-resolution nucleic acid structural motifs, researchers are focusing on developing approaches and tools to accurately predict water positions and interactions with nucleic acid. Biedermannová et al. proposed a Fourier averaging procedure that analyzes hydrated nucleic acid dinucleotide conformers, known as NtC blocks, as a method to predict DNA hydration structures.168 Using data from approximately 41000 NtC blocks associated with 316265 water molecules, they developed a strategy to predict the hydration patterns within the DNA structures. This work led to the creation of a web server, the WatNA atlas (https://www.watlas.datmos.org/watna/), which provides insights into DNA hydration (Fig. 8A and B). This is a promising approach to creating fully automated prediction servers, which could aid in the interpretation of experimental data. Meisburger et al. introduced methods for decomposing X-ray scattering profiles of DNA, water, and metal ions using small-angle X-ray scattering and validated it through anomalous scattering measurements.169 This approach provides an additional means of studying hydration patterns around nucleic acids. In addition, computational tools have become essential for studying nucleic acid dynamics and interactions. Giambaşu et al. introduced a toolset based on the three-dimensional reference interaction site model (3D-RISM) and molecular solvation theory to predict cation and water binding sites in G-quadruplex nucleic acids (Fig. 8C).170 By refining force field parameters using structural and thermodynamic data, they successfully modeled cation and water interactions in G-quadruplexes with 40% accuracy in placing water molecules within 1 Å of crystallographic water positions in experimental structures. Wei et al. developed an improved method called Solvation Potential Laid around Statistical Hydration on Entire Macromolecules (SPLASH’EM), which accurately places water molecules in nucleic acid cavities (Fig. 8D).171 By employing a particle waters force field with an angular contribution function and a hybrid scoring function based on a statistical analysis of crystal structures, their method enhances water placement predictions in nucleic acids with 62% accuracy. Such computational studies not only provide valuable insights into water-binding properties of nucleic acids, but also offer medicinal chemists enhanced tools for designing water-mediated nucleic acid-binding ligands with greater precision.
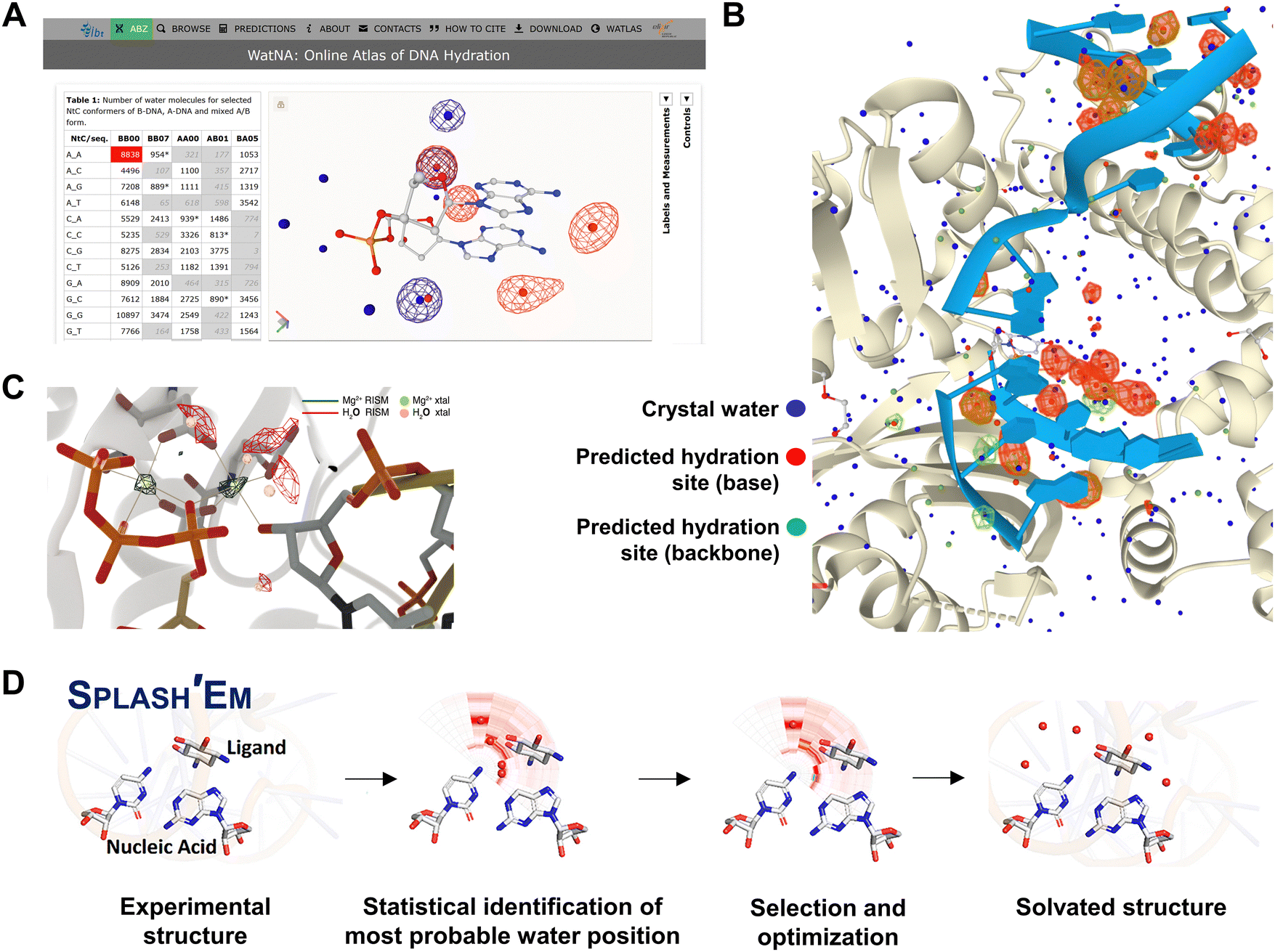 | ||
| Fig. 8 Tools and techniques for predicting water interactions in nucleic acid structures. (A) Screenshot of the WatNA online atlas of DNA hydration showing the predicted water positions in an AA dinucleotide step. (B) Screenshot of the predicted water positions represented by green and red spheres, together with the crystal waters shown as blue spheres in the PDB structure 6WID. (C) Predicted positions of metal ions and water densities (in dark green mesh) together with crystallographically resolved positions (in light green and pink mesh), illustrating the prediction accuracy of the three-dimensional reference interaction site model (3D-RISM) and molecular solvation theory. The figure is adapted with permission from J. Am. Chem. Soc., 2019, 141(6), 2435–2445. (D) The procedure for predicting water sites in nucleic acid–drug complex structures as proposed for the SPLASH’EM program. The figure is adapted with permission from J. Chem. Inf. Model., 2019, 59(6), 2941–2951. | ||
Spectroscopic techniques can also significantly contribute to advancing our understanding of the relationship between hydration and nucleic acids. For example, Franck et al. used the Overhauser effect dynamic nuclear polarization technique to demonstrate that water at DNA surfaces exhibits fast, bulk-like behavior, moving much more rapidly than water at protein or lipid surfaces.172 Terahertz time-domain spectroscopy has been employed to illustrate how the hydration shells of nucleic acids differ from free bulk water.173 McDermott et al., using second-order nonlinear optical spectroscopy, discovered that the DNA helical structure is surrounded by a network of chiral water molecules, particularly in the minor groove.52 More recently, by combining chiral-selective sum frequency generation spectroscopy (chiral-SFG) with computational analysis, Perets et al. demonstrated that the chiral water structures around DNA are confined to the first hydration shell.174 Notably, the population of chiral water molecules varies among the minor groove, major groove, and phosphate backbone. These findings contrast with earlier observations, which suggested that chiral water was confined solely to the minor groove. This updated understanding underscores a more intricate distribution of water across different regions of the DNA structure. Thus, the spectroscopy-based studies deepen our understanding of interactions of water with nucleic acids. Exploring the biological relevance of these findings could provide valuable insights into DNA function and aid in designing next-generation nucleic acid binding agents.
6. Conclusions and perspectives
Water plays a critical role in stabilizing biomolecular structures and supporting their functions.48,175,176 In this review, we have emphasized the importance of water in DNA structures by examining structural studies focused on water interactions. Through various examples, we have highlighted how water is essential in maintaining biologically relevant DNA structures and how water–nucleic acid interactions are vital for their stability and functionality. We also explored how water-bound motifs in DNA can act as binding sites for small molecules, including DNA-targeting drugs. These analyses reinforce the idea of water as a molecular modulator, not only for nucleic acid structures but also for their biological functions. Most of the water-mediated structures discussed in this review were determined through X-ray crystallography, which typically provides time-averaged, static representations of water interactions with DNA. This raises the question of whether the observations from crystal structures are consistent with in-cell conditions and accurately represent the real-time dynamic interactions between water and biomolecules. Lang et al. studied water structures within live cells and observed the hydration shells around soluble proteins.177 Tros et al. studied the random orientational motion of water molecules inside living cells using ultrafast vibrational spectroscopy and dielectric-relaxation spectroscopy.178 Interestingly, they found that slow water bound mostly to proteins and, to a lesser extent, to other biomolecules. The FTIR spectra of live cells have demonstrated DNA conformational changes upon hydration and dehydration, suggesting that the conformational changes observed in in vitro experimental conditions can also happen inside live cells.179 Although there is no direct tool to investigate the atomic-level details of water interactions with genomic DNA within cells, the current structural data on water-mediated DNA complexes are limited. However, the analysis of these structures has offered valuable insights into the broader role of water in biology and drug discovery.Similar to DNA, water plays a crucial role in mediating different RNA structures. The structures of RNA containing repeat expansions have been extensively studied, revealing significant insights. For example, in CGG and CAG repeat RNA structures, the non-canonical geometries of G–G and A–A mismatches are heavily hydrated within the major groove.180,181 These solvent-exposed base pair geometries attract charged species and bridge the interactions between mismatched pairs, suggesting a potential role for water in ligand–RNA interactions. Wang et al. reported the structures of myotonic dystrophy type 1-related CUG repeat RNA, where water molecules specifically bridge U–U mismatches through both the major and minor grooves.182 Their study demonstrated that water enables the interchangeable conformations of U–U mismatches in these RNA structures, which may be critical for the recognition by external ligands. Moreover, water has been shown to play an important role in the formation of tertiary structures and in facilitating specific interactions between RNA and other molecules in functional nucleic acids, such as riboswitches and aptamers.183–186 The formation of water-mediated interactions in RNA structures highlights the role of water, where it not only stabilizes non-canonical pairings but may also serve as a critical factor for the binding of RNA-binding proteins or small-molecule ligands. While a detailed discussion of water interactions in RNA structures is beyond the scope of this review, it is an important topic deserving a separate, dedicated analysis.
Meanwhile, advancing our understanding of water–nucleic acid interactions requires addressing the limitations of current methods. Structural techniques such as X-ray crystallography and NMR are instrumental in elucidating nucleic acid structures at the atomic level. However, these methods have limitations in capturing the dynamic and transient nature of water around nucleic acids.187 For example, while X-ray crystallography is useful for identifying structured water molecules bound to DNA, it cannot reliably depict the behavior of bulk or dynamic water molecules. Similarly, NMR spectroscopy provides valuable information about hydration patterns but may not be able to capture fast-exchanging water molecules. To overcome these limitations, more sophisticated methods need to be developed by combining experimental and computational approaches.188 Advanced techniques such as two-dimensional infrared spectroscopy and neutron scattering offer promising opportunities to study hydration shell dynamics, providing real-time, high-resolution data on water interactions.189–192 On the computational front, MD simulations are powerful tools for modeling water behavior around nucleic acids. Enhancing these simulations with improved force fields and incorporating quantum mechanical calculations can significantly boost the accuracy of predictions related to water-mediated interactions. Emerging fields such as artificial intelligence and deep learning also hold great potential for predicting water–biomolecule interactions. For example, a deep learning-based model, HydraProt, has recently been developed to accurately predict water positions within protein structures.193 Such approaches could pave the way for predictive models of water interactions with nucleic acids that would be faster and more accurate than traditional methods alone. The integration of new water-mediated structural motifs with experimental and computational methods has the potential to revolutionize our understanding of the effect of water on nucleic acid structures and functions. This, in turn, could open new opportunities in drug discovery by enabling the design of novel molecules that specifically target water-mediated nucleic acid structures.
Author contributions
Roshan Satange: writing – original draft, writing – review & editing, and visualization. Ming-Hon Hou: conceptualization, writing – original draft, writing – review & editing, supervision, and funding acquisition.Data availability
No primary research results have been included and no new data were generated as part of this review.Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors of this review article would like to acknowledge the National Synchrotron Radiation Research Center (NSRRC), Taiwan, for its continuing support in the field of nucleic acids research. This work was supported by the National Science and Technology Council, Taiwan, ROC [113-2311-B-005-006-MY3, 113-2113-M-005-016-MY3 to Ming-Hon Hou].References
- E. Brini, C. J. Fennell, M. Fernandez-Serra, B. Hribar-Lee, M. Lukšič and K. A. Dill, How water's poperties are encoded in its molecular structure and energies, Chem. Rev., 2017, 117(19), 12385–12414 CrossRef CAS PubMed.
- P. Ball, Water is an active matrix of life for cell and molecular biology, Proc. Natl. Acad. Sci. U. S. A., 2017, 114(51), 13327–13335 CrossRef CAS PubMed.
- B. Chen, I. Ivanov, M. L. Klein and M. Parrinello, Hydrogen bonding in water, Phys. Rev. Lett., 2003, 91(21), 215503 CrossRef PubMed.
- D. Laage, G. Stirnemann, F. Sterpone, R. Rey and J. T. Hynes, Reorientation and allied dynamics in water and aqueous solutions, Annu. Rev. Phys. Chem., 2011, 62, 395–416 CrossRef CAS PubMed.
- D. Laage and J. T. Hynes, A molecular jump mechanism of water reorientation, Science, 2006, 311(5762), 832–835 CAS.
- D. Laage and J. T. Hynes, Reorientional dynamics of water molecules in anionic hydration shells, Proc. Natl. Acad. Sci. U. S. A., 2007, 104(27), 11167–11172 CAS.
- E. Duboué-Dijon, A. C. Fogarty, J. T. Hynes and D. Laage, Dynamical disorder in the DNA hydration shell, J. Am. Chem. Soc., 2016, 138(24), 7610–7620 Search PubMed.
- F. Sterpone, G. Stirnemann and D. Laage, Magnitude and molecular origin of water slowdown next to a protein, J. Am. Chem. Soc., 2012, 134(9), 4116–4119 CAS.
- S. K. Pal and A. H. Zewail, Dynamics of water in biological recognition, Chem. Rev., 2004, 104(4), 2099–2124 CAS.
- M.-C. Bellissent-Funel, A. Hassanali, M. Havenith, R. Henchman, P. Pohl and F. Sterpone, et al., Water determines the structure and dynamics of proteins, Chem. Rev., 2016, 116(13), 7673–7697 CAS.
- C. A. Davey, D. F. Sargent, K. Luger, A. W. Maeder and T. J. Richmond, Solvent mediated interactions in the structure of the nucleosome core particle at 1.9 Å resolution, J. Mol. Biol., 2002, 319(5), 1097–1113 CAS.
- Z. Otwinowski, R. W. Schevitz, R. G. Zhang, C. L. Lawson, A. Joachimiak and R. Q. Marmorstein, et al., Crystal structure of trp represser/operator complex at atomic resolution, Nature, 1988, 335(6188), 321–329 CAS.
- P. Setny and M. D. Wiśniewska, Water-mediated conformational preselection mechanism in substrate binding cooperativity to protein kinase A, Proc. Natl. Acad. Sci. U. S. A., 2018, 115(15), 3852–3857 CrossRef CAS PubMed.
- W. Chen, H. He, J. Wang, J. Wang and C.-E. A. Chang, Uncovering water effects in protein–ligand recognition: importance in the second hydration shell and binding kinetics, Phys. Chem. Chem. Phys., 2023, 25(3), 2098–2109 CAS.
- M. L. Barreca, N. Iraci, G. Manfroni, R. Gaetani, C. Guercini and S. Sabatini, et al., Accounting for target flexibility and water molecules by docking to ensembles of target structures: the HCV NS5B palm site I inhibitors case study, J. Chem. Inf. Model., 2014, 54(2), 481–497 CAS.
- M. J. Fink and P.-O. Syrén, Redesign of water networks for efficient biocatalysis, Curr. Opin. Chem. Biol., 2017, 37, 107–114 CAS.
- K. Takano, J. Funahashi, Y. Yamagata, S. Fujii and K. Yutani, Contribution of water molecules in the interior of a protein to the conformational stability, J. Mol. Biol., 1997, 274(1), 132–142 CAS.
- C. J. Crilly, J. E. Eicher, O. Warmuth, J. M. Atkin and G. J. Pielak, Water's variable role in protein stability uncovered by liquid-observed vapor exchange NMR, Biochemistry, 2021, 60(41), 3041–3045 CAS.
- S.-H. Chong and S. Ham, Dynamics of hydration water plays a key role in determining the binding thermodynamics of protein complexes, Sci. Rep., 2017, 7(1), 8744 Search PubMed.
- K. Gerwert, E. Freier and S. Wolf, The role of protein-bound water molecules in microbial rhodopsins, Biochim. Biophys. Acta, Bioenerg., 2014, 1837(5), 606–613 CrossRef CAS PubMed.
- T. Khan, B. Halder, N. Das and P. Sen, Role of associated water dynamics on protein stability and activity in crowded milieu, J. Phys. Chem. B, 2024, 128(36), 8672–8686 CrossRef CAS PubMed.
- F. Mallamace, C. Corsaro, D. Mallamace, S. Vasi, C. Vasi and G. Dugo, The role of water in protein's behavior: the two dynamical crossovers studied by NMR and FTIR techniques, Comput. Struct. Biotechnol. J., 2015, 13, 33–37 CrossRef CAS PubMed.
- G. Schirò and M. Weik, Role of hydration water in the onset of protein structural dynamics, J. Phys.: Condens. Matter, 2019, 31(46), 463002 CrossRef PubMed.
- S. Neidle, H. M. Berman and H. S. Shieh, Highly structured water network in crystals of a deoxydinucleoside–drug complex, Nature, 1980, 288(5787), 129–133 CrossRef CAS PubMed.
- E. Westhof, Water: an integral part of nucleic acid structure, Annu. Rev. Biophys., 1988, 17, 125–144 CAS.
- H. M. Berman, Hydration of DNA, Curr. Opin. Struct. Biol., 1991, 1(3), 423–427 CrossRef CAS.
- M. Egli, S. Portmann and N. Usman, RNA hydration: a detailed look, Biochemistry, 1996, 35(26), 8489–8494 CrossRef CAS PubMed.
- H. M. Berman, Hydration of DNA: take 2, Curr. Opin. Struct. Biol., 1994, 4(3), 345–350 CrossRef CAS.
- H. Zhang, H. Fu, X. Shao, F. Dehez, C. Chipot and W. Cai, Changes in microenvironment modulate the B- to A-DNA transition, J. Chem. Inf. Model., 2019, 59(5), 2324–2330 CAS.
- M. Kulkarni and A. Mukherjee, Understanding B-DNA to A-DNA transition in the right-handed DNA helix: perspective from a local to global transition, Prog. Biophys. Mol. Biol., 2017, 128, 63–73 CAS.
- D. M. Gray, S. P. Edmondson, D. Lang, M. Vaughan and C. Nave, The circular dichroism and X-ray diffraction of DNA condensed from ethanolic solutions, Nucleic Acids Res., 1979, 6(6), 2089–2107 CAS.
- K. S. Lee, D. Bumbaca, J. Kosman, P. Setlow and M. J. Jedrzejas, Structure of a protein–DNA complex essential for DNA protection in spores of Bacillus species, Proc. Natl. Acad. Sci. U. S. A., 2008, 105(8), 2806–2811 CAS.
- J. P. Hall, J. Sanchez-Weatherby, C. Alberti, C. H. Quimper, K. O’Sullivan and J. A. Brazier, et al., Controlled dehydration of a ruthenium complex–DNA crystal induces reversible DNA kinking, J. Am. Chem. Soc., 2014, 136(50), 17505–17512 CAS.
- S. Neidle, Structured waters mediate small molecule binding to G-quadruplex nucleic acids, Pharmaceuticals, 2022, 15(1), 7 CAS.
- B. Nguyen, S. Neidle and W. D. Wilson, A Role for water molecules in DNA–ligand minor groove recognition, Acc. Chem. Res., 2009, 42(1), 11–21 CAS.
- M. Wu and I. Tinoco, RNA folding causes secondary structure rearrangement, Proc. Natl. Acad. Sci. U. S. A., 1998, 95(20), 11555–11560 CAS.
- M. L. Ken, R. Roy, A. Geng, L. R. Ganser, A. Manghrani and B. R. Cullen, et al., RNA conformational propensities determine cellular activity, Nature, 2023, 617(7962), 835–841 CAS.
- C. Templeton and R. Elber, Why does RNA collapse? The importance of water in a simulation study of helix–junction–helix systems, J. Am. Chem. Soc., 2018, 140(49), 16948–16951 CAS.
- S. Jones, D. T. A. Daley, N. M. Luscombe, H. M. Berman and J. M. Thornton, Protein–RNA interactions: a structural analysis, Nucleic Acids Res., 2001, 29(4), 943–954 CAS.
- B. P. Fingerhut, The mutual interactions of RNA, counterions and water – quantifying the electrostatics at the phosphate–water interface, Chem. Commun., 2021, 57(96), 12880–12897 CAS.
- A. Barik and R. P. Bahadur, Hydration of protein–RNA recognition sites, Nucleic Acids Res., 2014, 42(15), 10148–10160 CAS.
- Q. Vicens and E. Westhof, Crystal structure of paromomycin docked into the eubacterial ribosomal decoding A site, Structure, 2001, 9(8), 647–658 CAS.
- M. M. Rhodes, K. Réblová, J. Šponer and N. G. Walter, Trapped water molecules are essential to structural dynamics and function of a ribozyme, Proc. Natl. Acad. Sci. U. S. A., 2006, 103(36), 13380–13385 CAS.
- A. C. Fogarty and D. Laage, Water dynamics in protein hydration shells: the molecular origins of the dynamical perturbation, J. Phys. Chem. B, 2014, 118(28), 7715–7729 CAS.
- M. Kochoyan and J. L. Leroy, Hydration and solution structure of nucleic acids, Curr. Opin. Struct. Biol., 1995, 5(3), 329–333 CrossRef CAS PubMed.
- A. K. Singh, C. Wen, S. Cheng and N. Q. Vinh, Long-range DNA–water interactions, Biophys. J., 2021, 120(22), 4966–4979 CrossRef CAS PubMed.
- Z. Luo, H. Xiao, X. Peng, Y. Li, Z. Zhu and Y. Tian, et al., Long-range ordered water correlations between A–T/C–G nucleotides, Matter, 2020, 3(3), 794–804 CrossRef.
- D. Laage, T. Elsaesser and J. T. Hynes, Water dynamics in the hydration shells of biomolecules, Chem. Rev., 2017, 117(16), 10694–10725 CrossRef CAS PubMed.
- H. R. Drew and R. E. Dickerson, Structure of a B-DNA dodecamer: III. Geometry of hydration, J. Mol. Biol., 1981, 151(3), 535–556 CrossRef CAS PubMed.
- X. Shui, C. C. Sines, L. McFail-Isom, D. VanDerveer and L. D. Williams, Structure of the potassium form of CGCGAATTCGCG: DNA deformation by electrostatic collapse around inorganic cations, Biochemistry, 1998, 37(48), 16877–16887 CrossRef CAS PubMed.
- K. K. Woods, T. Lan, L. W. McLaughlin and L. D. Williams, The role of minor groove functional groups in DNA hydration, Nucleic Acids Res., 2003, 31(5), 1536–1540 CrossRef CAS PubMed.
- M. L. McDermott, H. Vanselous, S. A. Corcelli and P. B. Petersen, DNA's chiral spine of hydration, ACS Cent. Sci., 2017, 3(7), 708–714 CrossRef CAS PubMed.
- E. Liepinsh, G. Otting and K. Wüthrich, NMR observation of individual molecules of hydration water bound to DNA duplexes: direct evidence for a spine of hydration water present in aqueous solution, Nucleic Acids Res., 1992, 20(24), 6549–6553 CrossRef CAS PubMed.
- D. Vlieghe, J. P. Turkenburg and L. Van Meervelt, B-DNA at atomic resolution reveals extended hydration patterns, Acta Crystallogr., Sect. D: Biol. Crystallogr., 1999, 55(Pt 9), 1495–1502 CAS.
- M. Soler-López, L. Malinina, J. Liu, T. Huynh-Dinh and J. A. Subirana, Water and ions in a high resolution structure of B-DNA, J. Biol. Chem., 1999, 274(34), 23683–23686 CrossRef PubMed.
- T. Maehigashi, C. Hsiao, K. Kruger Woods, T. Moulaei, N. V. Hud and L. Dean Williams, B-DNA structure is intrinsically polymorphic: even at the level of base pair positions, Nucleic Acids Res., 2011, 40(8), 3714–3722 CrossRef PubMed.
- A. Madhumalar and M. Bansal, Structural insights into the effect of hydration and ions on A-tract DNA: a molecular dynamics study, Biophys. J., 2003, 85(3), 1805–1816 CrossRef CAS PubMed.
- B. Ghosh, S. Layek, D. Bhattacharyya and N. Sengupta, Base pair compositional variability influences DNA structural stability and tunes hydration thermodynamics and dynamics, J. Chem. Phys., 2023, 159(9), 095101 CrossRef CAS PubMed.
- S. Pal, P. K. Maiti and B. Bagchi, Exploring DNA groove water dynamics through hydrogen bond lifetime and orientational relaxation, J. Chem. Phys., 2006, 125(23), 234903 CrossRef PubMed.
- B. Schneider and H. M. Berman, Hydration of the DNA bases is local, Biophys J., 1995, 69(6), 2661–2669 CAS.
- A. Mardt, R. F. Gorriz, F. Ferraro, P. Ulrich, M. Zahran and P. Imhof, Effect of a U:G mispair on the water around DNA, Biophys. Chem., 2022, 283, 106779 CAS.
- L. D. Moore, T. Le and G. Fan, DNA methylation and its basic function, Neuropsychopharmacology, 2013, 38(1), 23–38 CAS.
- X. Teng and W. Hwang, Effect of methylation on local mechanics and hydration structure of DNA, Biophys. J., 2018, 114(8), 1791–1803 CAS.
- S.-M. Lin, H.-T. Huang, P.-J. Fang, C.-F. Chang, R. Satange and C.-K. Chang, et al., Structural basis of water-mediated cis Watson–Crick/Hoogsteen base-pair formation in non-CpG methylation, Nucleic Acids Res., 2024, 52(14), 8566–8579 CAS.
- D. Renciuk, O. Blacque, M. Vorlickova and B. Spingler, Crystal structures of B-DNA dodecamer containing the epigenetic modifications 5-hydroxymethylcytosine or 5-methylcytosine, Nucleic Acids Res., 2013, 41(21), 9891–9900 CAS.
- L. Lercher, M. A. McDonough, A. H. El-Sagheer, A. Thalhammer, S. Kriaucionis and T. Brown, et al., Structural insights into how 5-hydroxymethylation influences transcription factor binding, Chem. Commun., 2014, 50(15), 1794–1796 CAS.
- R. Wang, S. V. Ranganathan, V. A. Valsangkar, S. M. Magliocco, F. Shen and A. Chen, et al., Water-bridged hydrogen bond formation between 5-hydroxylmethylcytosine (5-hmC) and its 3′-neighbouring bases in A- and B-form DNA duplexes, Chem. Commun., 2015, 51(91), 16389–16392 CAS.
- T. Wang, Q. Pan, L. Lin, K. E. Szulwach, C.-X. Song and C. He, et al., Genome-wide DNA hydroxymethylation changes are associated with neurodevelopmental genes in the developing human cerebellum, Hum. Mol. Genet., 2012, 21(26), 5500–5510 CAS.
- J. A. Hackett, R. Sengupta, J. J. Zylicz, K. Murakami, C. Lee and T. A. Down, et al., Germline DNA demethylation dynamics and imprint erasure through 5-hydroxymethylcytosine, Science, 2013, 339(6118), 448–452 CAS.
- E.-A. Raiber, P. Murat, D. Y. Chirgadze, D. Beraldi, B. F. Luisi and S. Balasubramanian, 5-Formylcytosine alters the structure of the DNA double helix, Nat. Struct. Mol. Biol., 2015, 22(1), 44–49 CAS.
- L. Hu, J. Lu, J. Cheng, Q. Rao, Z. Li and H. Hou, et al., Structural insight into substrate preference for TET-mediated oxidation, Nature, 2015, 527(7576), 118–122 CrossRef CAS PubMed.
- F.-Y. Teng, Z.-Z. Jiang, M. Guo, X.-Z. Tan, F. Chen and X.-G. Xi, et al., G-quadruplex DNA: a novel target for drug design, Cell. Mol. Life Sci., 2021, 78(19), 6557–6583 CrossRef CAS PubMed.
- D. Varshney, J. Spiegel, K. Zyner, D. Tannahill and S. Balasubramanian, The regulation and functions of DNA and RNA G-quadruplexes, Nat. Rev. Mol. Cell Biol., 2020, 21(8), 459–474 CrossRef CAS PubMed.
- M. Farag and L. Mouawad, Comprehensive analysis of intramolecular G-quadruplex structures: furthering the understanding of their formalism, Nucleic Acids Res., 2024, 52(7), 3522–3546 CrossRef CAS PubMed.
- G. N. Parkinson, M. P. H. Lee and S. Neidle, Crystal structure of parallel quadruplexes from human telomeric DNA, Nature, 2002, 417(6891), 876–880 CrossRef CAS PubMed.
- G. Laughlan, A. I. H. Murchie, D. G. Norman, M. H. Moore, P. C. E. Moody and D. M. J. Lilley, et al., The high-resolution crystal structure of a parallel-stranded guanine tetraplex, Science, 1994, 265(5171), 520–524 CrossRef CAS PubMed.
- K. Li, L. Yatsunyk and S. Neidle, Water spines and networks in G-quadruplex structures, Nucleic Acids Res., 2020, 49(1), 519–528 Search PubMed.
- G. R. Clark, P. D. Pytel and C. J. Squire, The high-resolution crystal structure of a parallel intermolecular DNA G-4 quadruplex/drug complex employing syn glycosyl linkages, Nucleic Acids Res., 2012, 40(12), 5731–5738 CAS.
- D. Wei, G. N. Parkinson, A. P. Reszka and S. Neidle, Crystal structure of a c-kit promoter quadruplex reveals the structural role of metal ions and water molecules in maintaining loop conformation, Nucleic Acids Res., 2012, 40(10), 4691–4700 CAS.
- A. Ou, J. W. Schmidberger, K. A. Wilson, C. W. Evans, J. A. Hargreaves and M. Grigg, et al., High resolution crystal structure of a KRAS promoter G-quadruplex reveals a dimer with extensive poly-A π-stacking interactions for small-molecule recognition, Nucleic Acids Res., 2020, 48(10), 5766–5776 CAS.
- F. R. Winnerdy, B. Bakalar, A. Maity, J. J. Vandana, Y. Mechulam and E. Schmitt, et al., NMR solution and X-ray crystal structures of a DNA molecule containing both right- and left-handed parallel-stranded G-quadruplexes, Nucleic Acids Res., 2019, 47(15), 8272–8281 CAS.
- W. J. Chung, B. Heddi, E. Schmitt, K. W. Lim, Y. Mechulam and A. T. Phan, Structure of a left-handed DNA G-quadruplex, Proc. Natl. Acad. Sci. U. S. A., 2015, 112(9), 2729–2733 CAS.
- P. Das, F. R. Winnerdy, A. Maity, Y. Mechulam and A. T. Phan, A novel minimal motif for left-handed G-quadruplex formation, Chem. Commun., 2021, 57(20), 2527–2530 CAS.
- L. A. Yatsunyk and S. Neidle, On water arrangements in right- and left-handed DNA structures, Molecules, 2024, 29(2), 505 CAS.
- K. H. Ngo, C. W. Liew, S. Lattmann, F. R. Winnerdy and A. T. Phan, Crystal structures of an HIV-1 integrase aptamer: formation of a water-mediated A·G·G·G·G pentad in an interlocked G-quadruplex, Biochem. Biophys. Res. Commun., 2022, 613, 153–158 CAS.
- D. Zhang, T. Huang, P. S. Lukeman and P. J. Paukstelis, Crystal structure of a DNA/Ba2+ G-quadruplex containing a water-mediated C-tetrad, Nucleic Acids Res., 2014, 42(21), 13422–13429 CAS.
- B. Pan, Y. Xiong, K. Shi, J. Deng and M. Sundaralingam, Crystal structure of an RNA purine-rich tetraplex containing adenine tetrads: implications for specific binding in RNA tetraplexes, Structure, 2003, 11(7), 815–823 CAS.
- B. Heddi, N. Martín-Pintado, Z. Serimbetov, A. Kari Teuku Mahfuzh and A. T. Phan, G-quadruplexes with (4n − 1) guanines in the G-tetrad core: formation of a G-triad·water complex and implication for small-molecule binding, Nucleic Acids Res., 2015, 44(2), 910–916 Search PubMed.
- Y. Geng, C. Liu, B. Zhou, Q. Cai, H. Miao and X. Shi, et al., The crystal structure of an antiparallel chair-type G-quadruplex formed by Bromo-substituted human telomeric DNA, Nucleic Acids Res., 2019, 47(10), 5395–5404 CAS.
- K. Gehring, J.-L. Leroy and M. Guéron, A tetrameric DNA structure with protonated cytosine-cytosine base pairs, Nature, 1993, 363(6429), 561–565 CAS.
- M. Zeraati, D. B. Langley, P. Schofield, A. L. Moye, R. Rouet and W. E. Hughes, et al., I-motif DNA structures are formed in the nuclei of human cells, Nat. Chem., 2018, 10(6), 631–637 CrossRef CAS PubMed.
- H. Abou Assi, M. Garavís, C. González and M. J. Damha, i-Motif DNA: structural features and significance to cell biology, Nucleic Acids Res., 2018, 46(16), 8038–8056 CrossRef PubMed.
- Y. P. Bhavsar-Jog, E. Van Dornshuld, T. A. Brooks, G. S. Tschumper and R. M. Wadkins, Epigenetic modification, dehydration, and molecular crowding effects on the thermodynamics of i-Motif structure formation from C-Rich DNA, Biochemistry, 2014, 53(10), 1586–1594 CrossRef CAS PubMed.
- L. Cai, L. Chen, S. Raghavan, A. Rich, R. Ratliff and R. Moyzis, Intercalated cytosine motif and novel adenine clusters in the crystal structure of the Tetrahymena telomere, Nucleic Acids Res., 1998, 26(20), 4696–4705 CrossRef CAS PubMed.
- C. H. Kang, I. Berger, C. Lockshin, R. Ratliff, R. Moyzis and A. Rich, Crystal structure of intercalated four-stranded d(C3T) at 1.4 A resolution, Proc. Natl. Acad. Sci. U. S. A., 1994, 91(24), 11636–11640 CrossRef CAS PubMed.
- C. Kang, I. Berger, C. Lockshin, R. Ratliff, R. Moyzis and A. Rich, Stable loop in the crystal structure of the intercalated four-stranded cytosine-rich metazoan telomere, Proc. Natl. Acad. Sci. U. S. A., 1995, 92(9), 3874–3878 CrossRef CAS PubMed.
- K. S. Li, D. Jordan, L. Y. Lin, S. E. McCarthy, J. S. Schneekloth Jr. and L. A. Yatsunyk, Crystal structure of an i-motif from the HRAS oncogene promoter, Angew. Chem., Int. Ed., 2023, 62(26), e202301666 CAS.
- D. Guneri, E. Alexandrou, K. El Omari, Z. Dvořáková, R. V. Chikhale and D. T. S. Pike, et al., Structural insights into i-motif DNA structures in sequences from the insulin-linked polymorphic region, Nat. Commun., 2024, 15(1), 7119 CAS.
- Y.-W. Chen, C.-R. Jhan, S. Neidle and M.-H. Hou, Structural basis for the identification of an i-motif tetraplex core with a parallel-duplex junction as a structural motif in CCG triplet repeats, Angew. Chem., Int. Ed., 2014, 53(40), 10682–10686 CAS.
- Y.-W. Chen, R. Satange, P.-C. Wu, C.-R. Jhan, C.-K. Chang and K.-R. Chung, et al., CoII(Chromomycin)2 complex induces a conformational change of CCG repeats from i-motif to base-extruded DNA duplex, Int. J. Mol. Sci., 2018, 19(9), 2796 Search PubMed.
- Y. Sato, K. Mitomi, T. Sunami, J. Kondo and A. Takénaka, DNA octaplex formation with an I-motif of water-mediated A-quartets: reinterpretation of the crystal structure of d(GCGAAAGC), J. Biochem., 2006, 140(6), 759–762 CAS.
- J. Kondo, W. Adachi, S. Umeda, T. Sunami and A. Takénaka, Crystal structures of a DNA octaplex with I-motif of G-quartets and its splitting into two quadruplexes suggest a folding mechanism of eight tandem repeats, Nucleic Acids Res., 2004, 32(8), 2541–2549 CAS.
- G. S. Erwin, G. Gürsoy, R. Al-Abri, A. Suriyaprakash, E. Dolzhenko and K. Zhu, et al., Recurrent repeat expansions in human cancer genomes, Nature, 2023, 613(7942), 96–102 CAS.
- K. Usdin and E. Grabczyk, DNA repeat expansions and human disease., Cell. Mol. Life Sci., 2000, 57(6), 914–931 CAS.
- D. Sardana, K. Yadav, H. Shweta, N. S. Clovis, P. Alam and S. Sen, Origin of Slow Solvation dynamics in DNA: DAPI in minor groove of Dickerson-Drew DNA, J. Phys. Chem. B, 2019, 123(48), 10202–10216 CAS.
- N. K. Harika and W. D. Wilson, Bound compound, interfacial water, and phenyl ring rotation dynamics of a compound in the DNA minor groove, Biochemistry, 2018, 57(33), 5050–5057 CAS.
- C.-R. Jhan, R. Satange, S.-C. Wang, J.-Y. Zeng, Y.-C. Horng and P. Jin, et al., Targeting the ALS/FTD-associated A-DNA kink with anthracene-based metal complex causes DNA backbone straightening and groove contraction, Nucleic Acids Res., 2021, 49(16), 9526–9538 CAS.
- R. Satange, A. B. Rode and M.-H. Hou, Revisiting recent unusual drug–DNA complex structures: implications for cancer and neurological disease diagnostics and therapeutics, Bioorg. Med. Chem., 2022, 76, 117094 CAS.
- Á. Sánchez-González, T. G. Castro, M. Melle-Franco and A. Gil, From groove binding to intercalation: unravelling the weak interactions and other factors modulating the modes of interaction between methylated phenanthroline-based drugs and duplex DNA, Phys. Chem. Chem. Phys., 2021, 23(47), 26680–26695 Search PubMed.
- M. L. Kopka, C. Yoon, D. Goodsell, P. Pjura and R. E. Dickerson, The molecular origin of DNA-drug specificity in netropsin and distamycin, Proc. Natl. Acad. Sci. U. S. A., 1985, 82(5), 1376–1380 CAS.
- J. Dolenc, C. Oostenbrink, J. Koller and W. F. van Gunsteren, Molecular dynamics simulations and free energy calculations of netropsin and distamycin binding to an AAAAA DNA binding site, Nucleic Acids Res., 2005, 33(2), 725–733 CAS.
- Y. Liu, A. Kumar, S. Depauw, R. Nhili, M.-H. David-Cordonnier and M. P. Lee, et al., Water-mediated binding of agents that target the DNA minor groove, J. Am. Chem. Soc., 2011, 133(26), 10171–10183 CAS.
- B. Nguyen, M. P. H. Lee, D. Hamelberg, A. Joubert, C. Bailly and R. Brun, et al., Strong binding in the DNA minor groove by an aromatic diamidine with a shape that does not match the curvature of the groove, J. Am. Chem. Soc., 2002, 124(46), 13680–13681 CAS.
- P. Athri and W. D. Wilson, Molecular dynamics of water-mediated interactions of a linear benzimidazole–biphenyl diamidine with the DNA minor groove, J. Am. Chem. Soc., 2009, 131(22), 7618–7625 CrossRef CAS PubMed.
- Y. Miao, M. P. H. Lee, G. N. Parkinson, A. Batista-Parra, M. A. Ismail and S. Neidle,
et al., Out-of-shape DNA minor groove binders: Induced fit interactions of heterocyclic dications with the DNA minor groove, Biochemistry, 2005, 44(45), 14701–14708 CrossRef CAS PubMed.
- D. Wei, W. D. Wilson and S. Neidle, Small-molecule binding to the DNA minor groove is mediated by a conserved water cluster, J. Am. Chem. Soc., 2013, 135(4), 1369–1377 CAS.
- P. B. Dervan and B. S. Edelson, Recognition of the DNA minor groove by pyrrole-imidazole polyamides, Curr. Opin. Struct. Biol., 2003, 13(3), 284–299 CrossRef CAS PubMed.
- Y. Kawamoto, T. Bando and H. Sugiyama, Sequence-specific DNA binding Pyrrole–imidazole polyamides and their applications, Bioorg. Med. Chem., 2018, 26(8), 1393–1411 CrossRef CAS PubMed.
- D. M. Chenoweth and P. B. Dervan, Structural basis for cyclic Py-Im polyamide allosteric inhibition of nuclear receptor binding, J. Am. Chem. Soc., 2010, 132(41), 14521–14529 CrossRef CAS PubMed.
- J. Oh, T. Jia, J. Xu, J. Chong, P. B. Dervan and D. Wang, RNA polymerase II trapped on a molecular treadmill: structural basis of persistent transcriptional arrest by a minor groove DNA binder, Proc. Natl. Acad. Sci. U. S. A., 2022, 119(3), e2114065119 CrossRef CAS PubMed.
- D. G. Brown, M. R. Sanderson, J. V. Skelly, T. C. Jenkins, T. Brown and E. Garman, et al., Crystal structure of a berenil-dodecanucleotide complex: the role of water in sequence-specific ligand binding, EMBO J., 1990, 9(4), 1329–1334 CrossRef CAS PubMed.
- D. G. Brown, M. R. Sanderson, E. Garman and S. Neidle, Crystal structure of a berenil-d(CGCAAATTTGCG) complex: an example of drug–DNA recognition based on sequence-dependent structural features, J. Mol. Biol., 1992, 226(2), 481–490 CrossRef CAS PubMed.
- C. M. Nunn and S. Neidle, Sequence-dependent drug binding to the minor groove of DNA: crystal structure of the DNA dodecamer d(CGCAAATTTGCG)2 complexed with propamidine, J. Med. Chem., 1995, 38(13), 2317–2325 CrossRef CAS PubMed.
- T. C. Jenkins and A. N. Lane, AT selectivity and DNA minor groove binding: modelling, NMR and structural studies of the interactions of propamidine and pentamidine with d(CGCGAATTCGCG)2, Biochim. Biophys. Acta, Gene Struct. Expression, 1997, 1350(2), 189–204 CrossRef CAS PubMed.
- E. N. Ogbonna, A. Paul, A. A. Farahat, J. R. Terrell, E. Mineva and V. Ogbonna, et al., X-ray structure characterization of the selective recognition of AT base pair sequences, ACS Bio Med Chem Au, 2023, 3(4), 335–348 CrossRef CAS PubMed.
- E. K. Carter, S. Laughlin-Toth, T. Dodd, W. D. Wilson and I. Ivanov, Small molecule binders recognize DNA microstructural variations via an induced fit mechanism, Phys. Chem. Chem. Phys., 2019, 21(4), 1841–1851 RSC.
- M. H. Hou, H. Robinson, Y. G. Gao and A. H. J. Wang, Crystal structure of the [Mg2+-(chromomycin A3)2]–d(TTGGCCAA)2 complex reveals GGCC binding specificity of the drug dimer chelated by a metal ion, Nucleic Acids Res., 2004, 32(7), 2214–2222 CrossRef CAS PubMed.
- C. Hou, S. Weidenbach, K. E. Cano, Z. Wang, P. Mitra and D. N. Ivanov, et al., Structures of mithramycin analogues bound to DNA and implications for targeting transcription factor FLI1, Nucleic Acids Res., 2016, 44(18), 8990–9004 CrossRef CAS PubMed.
- W.-H. Tseng, Wu. P.-C. Chang C-k, N.-J. Hu, G.-H. Lee and C.-C. Tzeng, et al., Induced-fit recognition of CCG trinucleotide repeats by a nickel–chromomycin complex resulting in large-scale DNA deformation, Angew. Chem., Int. Ed., 2017, 56(30), 8761–8765 CAS.
- R. Satange, C.-Y. Chuang, S. Neidle and M.-H. Hou, Polymorphic G:G mismatches act as hotspots for inducing right-handed Z DNA by DNA intercalation, Nucleic Acids Res., 2019, 47(16), 8899–8912 CrossRef CAS PubMed.
- S. Venugopal, V. Sharma, A. Mehra, I. Singh and G. Singh, DNA intercalators as anticancer agents, Chem. Biol. Drug Des., 2022, 100(4), 580–598 CAS.
- J. Karges, S. Kuang, F. Maschietto, O. Blacque, I. Ciofini and H. Chao, et al., Rationally designed ruthenium complexes for 1- and 2-photon photodynamic therapy, Nat. Commun., 2020, 11(1), 3262 CrossRef CAS PubMed.
- A. C. Komor and J. K. Barton, The path for metal complexes to a DNA target, Chem. Commun., 2013, 49(35), 3617–3630 RSC.
- C. V. Kumar, J. K. Barton and N. J. Turro, Photophysics of ruthenium complexes bound to double helical DNA, J. Am. Chem. Soc., 1985, 107(19), 5518–5523 CrossRef CAS.
- I. Ortmans, B. Elias, J. M. Kelly, C. Moucheron and A. Kirsch-DeMesmaeker, [Ru(TAP)2(dppz)]2+: a DNA intercalating complex, which luminesces strongly in water and undergoes photo-induced proton-coupled electron transfer with guanosine-5′-monophosphate, Dalton Trans., 2004,(4), 668–676 RSC.
- A. E. Friedman, J. C. Chambron, J. P. Sauvage, N. J. Turro and J. K. Barton, A molecular light switch for DNA: Ru(bpy)2(dppz)2+, J. Am. Chem. Soc., 1990, 112(12), 4960–4962 CrossRef CAS.
- J. P. Hall, P. M. Keane, H. Beer, K. Buchner, G. Winter and T. L. Sorensen, et al., Delta chirality ruthenium ‘light-switch’ complexes can bind in the minor groove of DNA with five different binding modes, Nucleic Acids Res., 2016, 44(19), 9472–9482 CrossRef CAS PubMed.
- H. Song, J. T. Kaiser and J. K. Barton, Crystal structure of Δ-[Ru(bpy)2dppz]2+ bound to mismatched DNA reveals side-by-side metalloinsertion and intercalation, Nat. Chem., 2012, 4(8), 615–620 CrossRef CAS PubMed.
- J. P. Hall, D. Cook, S. R. Morte, P. McIntyre, K. Buchner and H. Beer, et al., X-ray crystal structure of rac-[Ru(phen)2dppz]2+ with d(ATGCAT)2 shows enantiomer orientations and water ordering, J. Am. Chem. Soc., 2013, 135(34), 12652–12659 CrossRef CAS PubMed.
- J. Sartorius and H.-J. Schneider, Intercalation mechanisms with ds-DNA: binding modes and energy contributions with benzene, naphthalene, quinoline and indole derivatives including some antimalarials, J. Chem. Soc., Perkin Trans. 2, 1997,(11), 2319–2328 RSC.
- R. Satange, S.-H. Kao, C.-M. Chien, S.-H. Chou, C.-C. Lin and S. Neidle, et al., Staggered intercalation of DNA duplexes with base-pair modulation by two distinct drug molecules induces asymmetric backbone twisting and structure polymorphism, Nucleic Acids Res., 2022, 50(15), 8867–8881 CrossRef CAS PubMed.
- Y.-S. Lo, W.-H. Tseng, C.-Y. Chuang and M.-H. Hou, The structural basis of actinomycin D–binding induces nucleotide flipping out, a sharp bend and a left-handed twist in CGG triplet repeats, Nucleic Acids Res., 2013, 41(7), 4284–4294 CrossRef CAS PubMed.
- L.-T. Da and J. Yu, Base-flipping dynamics from an intrahelical to an extrahelical state exerted by thymine DNA glycosylase during DNA repair process, Nucleic Acids Res., 2018, 46(11), 5410–5425 CrossRef CAS PubMed.
- C.-M. Chien, P.-C. Wu, R. Satange, C.-C. Chang, Z.-L. Lai and L. D. Hagler, et al., Structural basis for targeting T:T mismatch with triaminotriazine-acridine conjugate induces a U-shaped head-to-head four-way junction in CTG repeat DNA, J. Am. Chem. Soc., 2020, 142(25), 11165–11172 CrossRef CAS PubMed.
- S.-C. Huang, C.-W. Chen, R. Satange, C.-C. Hsieh, C.-C. Chang and S.-C. Wang, et al., Targeting DNA junction sites by bis-intercalators induces topological changes with potent antitumor effects, Nucleic Acids Res., 2024, 52(15), 9303–9316 CrossRef CAS PubMed.
- P.-C. Wu, S.-L. Tzeng, C.-K. Chang, Y.-F. Kao, M. J. Waring and M.-H. Hou, Cooperative recognition of T:T mismatch by echinomycin causes structural distortions in DNA duplex, Nucleic Acids Res., 2018, 46(14), 7396–7404 CrossRef CAS PubMed.
- R. Satange, C.-C. Chang, L. Y. Li, S.-H. Lin, S. Neidle and M.-H. Hou, Synergistic binding of actinomycin D and echinomycin to DNA mismatch sites and their combined anti-tumour effects, Nucleic Acids Res., 2023, 51(8), 3540–3555 CrossRef CAS PubMed.
- G. Qin, Z. Liu, J. Yang, X. Liao, C. Zhao and J. Ren, et al., Targeting specific DNA G-quadruplexes with CRISPR-guided G-quadruplex-binding proteins and ligands, Nat. Cell Biol., 2024, 26(7), 1212–1224 CrossRef CAS PubMed.
- S. Balasubramanian, L. H. Hurley and S. Neidle, Targeting G-quadruplexes in gene promoters: a novel anticancer strategy?, Nat. Rev. Drug Discovery, 2011, 10(4), 261–275 CrossRef CAS PubMed.
- N. Kosiol, S. Juranek, P. Brossart, A. Heine and K. Paeschke, G-quadruplexes: a promising target for cancer therapy, Mol. Cancer, 2021, 20(1), 40 CrossRef CAS PubMed.
- S. M. Haider, G. N. Parkinson and S. Neidle, Structure of a G-quadruplex–ligand complex, J. Mol. Biol., 2003, 326(1), 117–125 CrossRef CAS PubMed.
- G. N. Parkinson, R. Ghosh and S. Neidle, Structural basis for binding of porphyrin to human telomeres, Biochemistry, 2007, 46(9), 2390–2397 CrossRef CAS PubMed.
- N. H. Campbell, G. N. Parkinson, A. P. Reszka and S. Neidle, Structural basis of DNA quadruplex recognition by an acridine drug, J. Am. Chem. Soc., 2008, 130(21), 6722–6724 CrossRef CAS PubMed.
- N. H. Campbell, M. Patel, A. B. Tofa, R. Ghosh, G. N. Parkinson and S. Neidle, Selectivity in ligand recognition of G-quadruplex loops, Biochemistry, 2009, 48(8), 1675–1680 CrossRef CAS PubMed.
- M. Micco, G. W. Collie, A. G. Dale, S. A. Ohnmacht, I. Pazitna and M. Gunaratnam, et al., Structure-based design and evaluation of naphthalene diimide G-quadruplex ligands as telomere targeting agents in pancreatic cancer cells, J. Med. Chem., 2013, 56(7), 2959–2974 CrossRef CAS PubMed.
- G. W. Collie, R. Promontorio, S. M. Hampel, M. Micco, S. Neidle and G. N. Parkinson, Structural basis for telomeric G-quadruplex targeting by naphthalene diimide ligands, J. Am. Chem. Soc., 2012, 134(5), 2723–2731 CAS.
- N. H. Campbell, D. L. Smith, A. P. Reszka, S. Neidle and D. O'Hagan, Fluorine in medicinal chemistry: β-fluorination of peripheral pyrrolidines attached to acridine ligands affects their interactions with G-quadruplex DNA, Org. Biomol. Chem., 2011, 9(5), 1328–1331 CAS.
- C. Bazzicalupi, M. Ferraroni, A. R. Bilia, F. Scheggi and P. Gratteri, The crystal structure of human telomeric DNA complexed with berberine: an interesting case of stacked ligand to G-tetrad ratio higher than 1:1, Nucleic Acids Res., 2012, 41(1), 632–638 Search PubMed.
- K. T. McQuaid, S. Takahashi, L. Baumgaertner, D. J. Cardin, N. G. Paterson and J. P. Hall, et al., Ruthenium polypyridyl complex bound to a unimolecular chair-form G-quadruplex, J. Am. Chem. Soc., 2022, 144(13), 5956–5964 CAS.
- S. K. Pal, L. Zhao, T. Xia and A. H. Zewail, Site- and sequence-selective ultrafast hydration of DNA, Proc. Natl. Acad. Sci. U. S. A., 2003, 100(24), 13746–13751 CAS.
- D. Andreatta, J. L. Pérez Lustres, S. A. Kovalenko, N. P. Ernsting, C. J. Murphy and R. S. Coleman, et al., Power-law solvation dynamics in DNA over six decades in time, J. Am. Chem. Soc., 2005, 127(20), 7270–7271 CAS.
- V. P. Denisov, G. Carlström, K. Venu and B. Halle, Kinetics of DNA hydration, J. Mol. Biol., 1997, 268(1), 118–136 CAS.
- M. Bastos, V. Castro, G. Mrevlishvili and J. Teixeira, Hydration of ds-DNA and ss-DNA by neutron quasielastic scattering, Biophys. J., 2004, 86(6), 3822–3827 CrossRef CAS PubMed.
- B. Halle, Biomolecular cryocrystallography: structural changes during flash-cooling, Proc. Natl. Acad. Sci. U. S. A., 2004, 101(14), 4793–4798 CrossRef CAS PubMed.
- G. M. Daubner, A. Cléry and F. H. T. Allain, RRM–RNA recognition: NMR or crystallography…and new findings, Curr. Opin. Struct. Biol., 2013, 23(1), 100–108 CrossRef CAS PubMed.
- M. M. Teeter, A. Yamano, B. Stec and U. Mohanty, On the nature of a glassy state of matter in a hydrated protein: relation to protein function, Proc. Natl. Acad. Sci. U. S. A., 2001, 98(20), 11242–11247 CrossRef CAS PubMed.
- J. S. Fraser, H. van den Bedem, A. J. Samelson, P. T. Lang, J. M. Holton and N. Echols, et al., Accessing protein conformational ensembles using room-temperature X-ray crystallography, Proc. Natl. Acad. Sci. U. S. A., 2011, 108(39), 16247–16252 CrossRef CAS PubMed.
- L. Biedermannova, J. Cerny, M. Maly, M. Nekardova and B. Schneider, Knowledge-based prediction of DNA hydration using hydrated dinucleotides as building blocks, Acta Crystallogr., Sect. D: Struct. Biol., 2022, 78(8), 1032–1045 CrossRef CAS PubMed.
- P. Meisburger Steve, A. Pabit Suzette and L. Pollack, Determining the locations of ions and water around DNA from X-ray scattering measurements, Biophys. J., 2015, 108(12), 2886–2895 CrossRef CAS PubMed.
- G. M. Giambaşu, D. A. Case and D. M. York, Predicting site-binding modes of ions and water to nucleic acids using molecular solvation theory, J. Am. Chem. Soc., 2019, 141(6), 2435–2445 CrossRef PubMed.
- W. Wei, J. Luo, J. Waldispühl and N. Moitessier, Predicting positions of bridging water molecules in nucleic acid–ligand complexes, J. Chem. Inf. Model., 2019, 59(6), 2941–2951 CrossRef CAS PubMed.
- J. M. Franck, Y. Ding, K. Stone, P. Z. Qin and S. Han, Anomalously rapid hydration water diffusion dynamics near DNA surfaces, J. Am. Chem. Soc., 2015, 137(37), 12013–12023 CrossRef CAS PubMed.
- N. A. Penkova, M. G. Sharapov and N. V. Penkov, Hydration shells of DNA from the point of view of terahertz time-domain spectroscopy, Int. J. Mol. Sci., 2021, 22(20), 11089 CrossRef CAS PubMed.
- E. A. Perets, D. Konstantinovsky, T. Santiago, P. E. Videla, M. Tremblay and L. Velarde, et al., Beyond the “spine of hydration”: Chiral SFG spectroscopy detects DNA first hydration shell and base pair structures, J. Chem. Phys., 2024, 161(9), 095104 CrossRef CAS PubMed.
- K. Mitusińska, A. Raczyńska, M. Bzówka, W. Bagrowska and A. Góra, Applications of water molecules for analysis of macromolecule properties, Comput. Struct. Biotechnol. J., 2020, 18, 355–365 CrossRef PubMed.
- N. Watanabe, K. Suga and H. Umakoshi, Functional hydration behavior: interrelation between hydration and molecular properties at lipid membrane interfaces, J. Chem., 2019, 2019(1), 4867327 Search PubMed.
- X. Lang, L. Shi, Z. Zhao and W. Min, Probing the structure of water in individual living cells, Nat. Commun., 2024, 15(1), 5271 CrossRef CAS PubMed.
- M. Tros, L. Zheng, J. Hunger, M. Bonn, D. Bonn and G. J. Smits, et al., Picosecond orientational dynamics of water in living cells, Nat. Commun., 2017, 8(1), 904 CrossRef PubMed.
- B. R. Wood, The importance of hydration and DNA conformation in interpreting infrared spectra of cells and tissues, Chem. Soc. Rev., 2016, 45(7), 1980–1998 RSC.
- A. Kiliszek, R. Kierzek, W. J. Krzyzosiak and W. Rypniewski, Crystal structures of CGG RNA repeats with implications for fragile X-associated tremor ataxia syndrome, Nucleic Acids Res., 2011, 39(16), 7308–7315 CrossRef CAS PubMed.
- A. Kiliszek, R. Kierzek, W. J. Krzyzosiak and W. Rypniewski, Atomic resolution structure of CAG RNA repeats: structural insights and implications for the trinucleotide repeat expansion diseases, Nucleic Acids Res., 2010, 38(22), 8370–8376 CrossRef CAS PubMed.
- S.-C. Wang, Y.-T. Chen, R. Satange, J.-W. Chu and M.-H. Hou, Structural basis for water modulating RNA duplex formation in the CUG repeats of myotonic dystrophy type 1, J. Biol. Chem., 2023, 299(7), 104864 CrossRef CAS PubMed.
- D. Sussman and C. Wilson, A water channel in the core of the vitamin B12 RNA aptamer, Structure, 2000, 8(7), 719–727 CrossRef CAS PubMed.
- K. D. Warner, L. Sjekloća, W. Song, G. S. Filonov, S. R. Jaffrey and A. R. Ferré-D'Amaré, A homodimer interface without base pairs in an RNA mimic of red fluorescent protein, Nat. Chem. Biol., 2017, 13(11), 1195–1201 CrossRef CAS PubMed.
- A. Sun, C. Gasser, F. Li, H. Chen, S. Mair and O. Krasheninina, et al., SAM-VI riboswitch structure and signature for ligand discrimination, Nat. Commun., 2019, 10(1), 5728 CrossRef CAS PubMed.
- X. Xu, M. Egger, H. Chen, K. Bartosik, R. Micura and A. Ren, Insights into xanthine riboswitch structure and metal ion-mediated ligand recognition, Nucleic Acids Res., 2021, 49(12), 7139–7153 CrossRef CAS PubMed.
- B. Z. Zsidó, B. Bayarsaikhan, R. Börzsei, V. Szél, V. Mohos and C. Hetényi, The advances and limitations of the determination and applications of water structure in molecular engineering, Int. J. Mol. Sci., 2023, 24(14), 11784 CrossRef PubMed.
- L. Biedermannová and B. Schneider, Hydration of proteins and nucleic acids: advances in experiment and theory. A review, Biochim. Biophys. Acta, Gen. Subj., 2016, 1860(9), 1821–1835 CrossRef PubMed.
- J. M. Harp, L. Coates, B. Sullivan and M. Egli, Water structure around a left-handed Z-DNA fragment analyzed by cryo neutron crystallography, Nucleic Acids Res., 2021, 49(8), 4782–4792 CAS.
- M. Yang, Ł. Szyc and T. Elsaesser, Vibrational dynamics of the water shell of DNA studied by femtosecond two-dimensional infrared spectroscopy, J. Photochem. Photobiol., A, 2012, 234, 49–56 CAS.
- I. Michalarias, I. A. Beta, J. C. Li, S. Ruffle and R. Ford, The interaction of water with DNA—a combined inelastic neutron scattering and infrared spectroscopic study, J. Mol. Liq., 2002, 101(1), 19–26 CAS.
- S. Hume, G. M. Greetham, P. M. Donaldson, M. Towrie, A. W. Parker and M. J. Baker, et al., 2D-infrared spectroscopy of proteins in water: using the solvent thermal response as an internal standard, Anal. Chem., 2020, 92(4), 3463–3469 CAS.
- A. Zamanos, G. Ioannakis and I. Z. Emiris, HydraProt: a new deep learning tool for fast and accurate prediction of water molecule positions for protein structures, J. Chem. Inf. Model., 2024, 64(7), 2594–2611 CAS.
| This journal is © The Royal Society of Chemistry 2025 |