
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceBioactive secondary metabolites from fungal endophytes, Penicillium oxalicum and Phoma herbarum, associated with Morus nigra and Ficus sycomorus: an in silico study†
Mohamed M. M. AbdelRazek a,
Ahmed M. Elissawybc,
Nada M. Mostafab,
Ashaimaa Y. Moussab,
Mohamed A. Elshanawanya and
Abdel Nasser B. Singab*bc
a,
Ahmed M. Elissawybc,
Nada M. Mostafab,
Ashaimaa Y. Moussab,
Mohamed A. Elshanawanya and
Abdel Nasser B. Singab*bc
aDepartment of Pharmacognosy, Faculty of Pharmacy, Badr University in Cairo (BUC), Cairo 11829, Egypt
bDepartment of Pharmacognosy, Faculty of Pharmacy, Ain-Shams University, Cairo 11566, Egypt. E-mail: vpr.nassersingab@asu.edu.eg
cCenter of Drug Discovery Research and Development, Ain Shams University, Cairo 11566, Egypt
First published on 14th November 2024
Abstract
Two pure fungal strains were isolated and identified from Ficus sycomorus and Morus nigra, namely, Penicillium oxalicum (OR673586) and Phoma herbarum (OR673589), respectively. The extract and fractions of secondary metabolites of each fungus were evaluated for antioxidant, anti-inflammatory, antimicrobial, antibiofilm, antidiabetic, and cytotoxic activities. The chloroform fraction of P. oxalicum showed potent cytotoxic activity (IC50 = 7.695 μg mL−1) against Hep-G2 cell line, alongside moderate antioxidant and anti-inflammatory activities. On the other hand, the P. herbarum chloroform fraction showed potent antioxidant (DPPH IC50 = 5.649 μg mL−1) and antidiabetic activities (IC50 = 14.91 μg mL−1) against inhibition of α-glucosidase, in addition to moderate cytotoxicity, anti-inflammatory, and antimicrobial activities. Guided cytotoxic fractionation leads to identifying bioactive compounds using hyphenated techniques. LC-MS identified fourteen compounds for P. herbarum and thirteen compounds for P. oxalicum. Three known compounds, mevalolactone (1), glycerol monolinoleate (3), and ergosterol (7) in addition to one new compound, barcelonyl acetate (2), were isolated from P. herbarum. On the other hand, four known compounds, 4-hydroxyphenyl acetic acid (4), secalonic acid D (5), altersolanol A (6), and ergosterol (7), were isolated from P. oxalicum. Altersolanol A (6) and secalonic acid D (7) exhibited outstanding cytotoxic activity against Hep-G2 and Caco-2 cell lines, with IC50 values ranging from 0.00038 to 0.208 μM. In silico study findings showed altersolanol A (6), 4-hydroxyphenyl acetic acid (4), glycerol monolinoleate (3), and barcelonyl acetate (2) displayed significant potential but may benefit from further optimization as lead for developing potent c-Jun N-terminal kinase 2 (JNK2, PDB: 3NPC) inhibitors, potentially leading to novel therapeutic strategies targeting cancer therapy.
1. Introduction
Ficus sycomorus and Morus nigra are medicinal plants used traditionally to treat different diseases in many parts of Africa and Asia.1,2 Endophytic fungi proved a promising source for optimizing new resources in drug discovery. Endophytic fungi contain a wealth of chemical classes, such as polyketides, alkaloids, coumarins, xanthones, anthraquinones, phenols, phenolic acids, terpenoids, steroids, sterols, and miscellaneous compounds.3–5 These are promising metabolites for developing potential medication candidates with bioactivities such as cytotoxic, antibiofilm, antiviral, anti-inflammatory, anthelmintic, antidiabetic, antioxidant, antibacterial, antifungal, and immune-modulating activities.6,7Through the last two decades, many reports have described the endophytic fungi associated with medicinal plants belonging to the family Moraceae. Many endophytic fungi isolates were reported from Morus alba from various tissues (leaves, stems, and roots), with Fusarium being the most prevalent genus. Among these isolates, a majority were successfully identified as genera Alternaria, Phoma, Colletotrichum, Aspergillus, Macrophomina, Penicillium, and Scytalidium, while the remaining isolates were classified as undefined.8–14 However, one endophytic fungal strain was reported of Morus plants as Botryosphaeria sp. for M. nigra, Phomopsis sp. for M. cathayana, and seven undefined strains for M. macroura.15–17
A limited number of articles reported the biological activities of endophytic fungal extracts associated with Morus plants evaluated for antifungal, antioxidant, antiviral, antidiabetic, and antibiofilm potential. In addition, few articles reported the isolation of compounds from endophytic fungi associated with Morus plants. The reported compounds belong to diverse chemical classes, including anthraquinones, quinones, pyrones, naphthoquinones, polyketides, oxazole derivatives, and furoic acid derivatives that were tested for various biological potentials of neuroprotective, antioxidant, antimicrobial, antimalarial, glucose inhibitory, hemolytic, and cytotoxic activities.7,8,10,11,16,18
However, Ficus endophytic fungal strains were reported of the genera Penicillium and Aspergillus as the most frequently identified endophytes. Few works reported the endophytic fungi associated with Ficus plants: F. elastica, F. carica, F. microcarpa, F. hirta, F. religiosa, and F. sphenophyllum.19–27 The reported studies on Ficus-associated endophytes showed cytotoxic, antioxidant, antidiabetic, antibacterial, and antifungal potential of the fungal extracts associated with F. elastica and F. religiosa.28,29 Endophytic fungi associated with F. elastica, F. carica, and F. hirta were reported to have different chemical classes of isolated compounds, including xanthones, alkaloids, chromones, isochromenones, cyclopentenones, polyketides, and triterpenoids. The activities of these compounds were evaluated for antioxidant, antibacterial, and antifungal to neuraminidase inhibitory and herbicidal potential.19–28
To our knowledge, the present study aims to characterize the endophytic fungi associated with F. sycomorus for the first time in addition to M. nigra of limited reported work. The study aims to evaluate the biological activities and chemical constituents of their active extracts and fractions as a potential source for further exploration of bioactive molecules for drug discovery and development. This study is part of an ongoing research project to investigate the active metabolites of endophytic fungi associated with Moraceae plants as an alternative eco-friendly source for active molecules.
2. Experimental section
2.1. Purification of the fungal endophytes
Morus nigra was collected from a farm in El-Monofia governorate, Egypt, while Ficus sycomorus was collected from El-Kanater Horticulture Research Station, El-Kalubia governorate, Egypt, in November 2020, identified by Prof. Abd El-Halim Abd El-Magly Mohamed (Horticulture Research Institute, Flora and Phytotaxonomy Research Unit, Egypt), and vouchered samples were kept at Ain Shams University under the codes PHG-P-MN-333 for M. nigra and PHG-P-FS-332 for F. sycomorus. The isolation and identification workup of the fungal strains were performed as previously reported.302.2. Cultivation, extraction, and isolation
Cultivation and fermentation of the fungal strains were performed as previously reported.30 The crude extract yield of twelve 1 L flasks for each fungal strain was 35.614 g for Penicillium oxalicum and 35.614 g for Phoma herbarum. The crude extracts were partitioned between n-hexane and 90% aqueous methanol (MeOH). Then liquid/liquid fractionation was performed using a separating funnel multiple times with varying gradients, involving the following combinations: n-hexane: MeOH (90%), chloroform (CHCl3): MeOH (60%), and ethyl acetate (EtOAc): MeOH (60%).31,32 Three fractions were collected for each fungal strain. P. herbarum fractions yield 14.8211 g for the n-hexane fraction (PH1), 6.1490 g for the CHCl3 fraction (PH2), 12.7969 g for the EtOAc fraction (PH3). While P. oxalicum yields 18.6911 g for the n-hexane fraction (PO1), 2.6845 g for the CHCl3 fraction (PO2), 5.5356 g for the ethyl acetate fraction (PO3). Vacuum liquid chromatography (VLC) was applied for the fungal CHCl3 fractions (PH2) and (PO2) on silica gel 60 eluting with gradient mobile phase (n-hexane-EtOAc 100![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :0 to 0:100, CHCl3–MeOH 100:0 to 0:100) to give thirteen subfractions of each CHCl3 fungal fraction. Various chromatographic techniques were employed to sequentially separate and refine the entire extract, such as VLC (Vacuum Liquid Chromatography), normal pressure CC (Column Chromatography), and semi-preparative HPLC (High-Performance Liquid Chromatography). Pure compounds were isolated and successively identified using diverse spectroscopic techniques.
:0 to 0:100, CHCl3–MeOH 100:0 to 0:100) to give thirteen subfractions of each CHCl3 fungal fraction. Various chromatographic techniques were employed to sequentially separate and refine the entire extract, such as VLC (Vacuum Liquid Chromatography), normal pressure CC (Column Chromatography), and semi-preparative HPLC (High-Performance Liquid Chromatography). Pure compounds were isolated and successively identified using diverse spectroscopic techniques.
2.3. Antioxidant evaluation
Antioxidant activity using the DPPH assay was evaluated as defined by.33,34 The FRAP assay was performed as defined by.35 Tested samples in the FRAP assay were of 1 mg mL−1 concentration. A calibration curve of Trolox was used, and results were expressed in μM Trolox equivalent per mL by substitution in the linear regression equation of the Trolox calibration curve (Y = 0.0018X − 0.0340, R2 = 0.9982).2.4. Cytotoxic evaluation
Cytotoxic activity was evaluated from the crude extracts and their fractions using the SRB assay as defined by,36,37 against the liver cancer (Hep-G2) cell line, while the isolated fungal metabolites were tested against colon cancer (Caco-2) and liver cancer (Hep-G2) cell lines, and doxorubicin was used as a positive control. Crude extracts and their fractions were prepared in methanol of serial concentrations 1000, 100, 10, 0.1, and 0.01 μg mL−1, while for isolated fungal metabolites of serial concentrations 1000, 100, 10, 0.1, 0.01, and 0.001 μg mL−1. Absorbance was measured at 540 nm using a microplate reader (BMG Labtech-FLUO star Omega, Ortenberg, Germany).2.5. Anti-inflammatory evaluation
The anti-inflammatory activity was performed by measuring the nitric oxide inhibitory activity of murine macrophage RAW264.7 cells using the assay as defined.38,39 L-NAME was used as a positive control. The nitric oxide (NO) inhibitory activity was measured at 540 nm using a microplate reader (BMG Labtech-FLUO star Omega, Ortenberg, Germany).2.6. Antimicrobial evaluation
The agar well diffusion method was performed to evaluate the antimicrobial activity as defined by.40 Samples were prepared for the agar well diffusion assay of 20 mg mL−1 concentration. While the minimum inhibitory concentration was done using a microtiter plate turbidity assay as defined by41–43 against three microorganisms: Staphylococcus aureus (ATCC-6538), Pseudomonas aeruginosa (ATCC-9027), and Candida albicans (ATCC-10231). Gentamicin (MEMPHIS, Egypt) was used as positive control.2.7. Antibiofilm inhibitory assay
The biofilm inhibitory activity was performed against Pseudomonas aeruginosa (ATCC-9027) using the assay as defined by.44 Samples were dissolved in 10% DMSO and prepared in serial concentrations of 10, 5, 2.5, 1.25, and 0.625 mg mL−1. The reaction mixture was read spectrophotometrically after gently shaking on a microplate reader (BioTek 800 TS, Agilent, USA) at 630 nm wavelength.2.8. Antidiabetic evaluation
The antidiabetic activity was tested using α-glucosidase inhibitory assay as defined by.45,46 Acarbose was used as a positive control. Samples were dissolved in DMSO, and serial dilutions were prepared. The enzymatic activity was measured at 405 nm using a microplate reader (BMG Labtech-FLUO star Omega, Ortenberg, Germany).2.9. Liquid chromatography-mass spectrometry (LC-MS)
The LC-MS experiments were conducted using a triple quadruple instrument, C-18 column: 1.7 μm, particle size 2.1 × 50 mm (XEVO TQD, Waters Corporation, Milford, USA) located at the Center for Drug Discovery, Research, and Development, Faculty of Pharmacy, Ain Shams University.47,482.10. Nuclear magnetic resonance (NMR) spectroscopy
The NMR experiments were conducted using a Bruker Ascend 400/R spectrophotometer (Bruker®, AVANCE III HD, 400 MHz, Switzerland) located at the Center for Drug Discovery, Research and Development, Faculty of Pharmacy, Ain Shams University.442.11. Protein and ligand preparation
The library of natural compounds was downloaded from the purchasable “natural products” library from the Zinc server,49 a freely accessible web-based server. The LigPrep module of Maestro was used to prepare the ligands at the standard setting and under the OPLS4 forcefield. The crystal structure of JNK2 complexed with the co-crystal BIRB796 was obtained from the protein data bank (PDB: 3NPC, 2.35 angstrom (Å)).50 The protein was prepared by removing water molecules and the AZD3229 analog, along with the addition of any missing residues and hydrogen atoms. Following that, the prepared protein structure was protonated at physiological pH. Finally, the energy was minimized by employing a conjugate gradient method based on the OPLS-2005 force field, which produced a unique low-energy minimum of the structure.512.12. Molecular docking
A molecular docking study was carried out from the hits identified in the virtual screening and fragment-based studies. To avoid false positives, each ligand was docked in the binding site using Glide's extra precision module, yielding 32 poses for each ligand. The 2D interaction diagrams were generated employing the BIOVIA Discovery Studio Visualizer 2022 package, and the docking scores were ranked, with the most negative scores chosen for each complex. Docking validation was carried out by redocking the BIRB796 analog (co-crystal) and calculating the RMSD value (root-mean-square deviation).3. Results and discussion
3.1. Morphological identification of M. nigra and F. sycomorus -associated endophytes
Two endophytic fungi were isolated from plants of the family Moraceae. The first strain encoded MN14 was isolated from the stem tissue of Morus nigra and exhibited distinctive characteristics, including ellipsoidal-shaped conidia measuring 4 × 3.5 μm, swollen phialides with a diameter of 23.5 μm and a length of 4.5 μm, and swollen metulae measuring 12.0 μm in diameter and 3.8 μm in length. These microscopic features and the observation of conidiophores reaching a diameter of 4.5 μm confirmed the strain's classification as Phoma sp. (Fig. S1, see ESI†). The second strain encoded FS12 was isolated from the stem tissue of Ficus sycomorus and displayed characteristic features, including bi-verticillate hyphae resembling Penicillium. Microscopic examination revealed ellipsoidal-shaped conidia measuring 4 × 3.5 μm in breadth. The presence of swollen phialides and metulae, measuring 23.5 × 4.5 μm and 12.0 × 3.8 μm in diameter, respectively. Conidiophores were observed to reach a diameter of 4.5 μm. Further supporting its classification as Penicillium sp. (Fig. S2, see ESI†).3.2. Molecular identification of M. nigra and F. sycomorus -associated endophytes
Phoma sp. MN14 fungal isolate from Morus nigra stem tissue and the Penicillium sp. FS12 fungal isolates from Ficus sycomorus stem tissue were sequenced using the genetic regions of the 18 s rRNA gene. When compared to similar sequences in the National Center for Biotechnology Information (NCBI) database, the Phoma isolate showed a 100% identity and 100% query coverage with P. herbarum BZYB, while the Penicillium isolates exhibited a 100% identity and 100% query coverage with P. oxalicum SL2. Alignment analysis using MEGA10 software revealed no differences in the 1653 base pairs analyzed for the Phoma isolate and 1649 for the Penicillium isolate. The evolutionary relationships of taxa were examined through Neighbour-Joining tree analysis and phylogenetic tree bootstrap analysis, confirming the alignment results (Fig. S3 and S4, see ESI†). The sequences of P. herbarum MN14 isolate and P. oxalicum FS12 isolate were deposited in GenBank under the accession numbers OR673589 and OR673586, respectively.3.3. Antioxidant activity
P. herbarum CHCl3 fraction (PH2) showed potent antioxidant activity with DPPH IC50 values of 5.649 ± 0.47 μg mL−1 compared to Trolox as a positive control with IC50 value of 7.217 ± 0.309 μg mL−1. The result was confirmed by the FRAP assay that showed value of 2926.48 ± 142.654 μM Trolox equivalent per mL. P. herbarum total extract (PHT) and its ethyl acetate fraction (PH3) in addition to P. oxalicum CHCl3 fraction (PO2) showed moderate antioxidant activity with IC50 values ranging from 38.95 ± 1.79 to 198 ± 7.76 μg mL−1. However, P. herbarium n-hexane fraction (PH1) in addition to P. oxalicum total extract (POT) and its n-hexane fraction (PO1) showed weak antioxidant activity with DPPH IC50 greater than 200 μg mL−1 (Table S1, see ESI†).3.4. Cytotoxic activity
The total extracts and their fractions were tested for cytotoxic activity. The most potent cytotoxic activity against liver cancer (Hep-G2) cell line was P. oxalicum CHCl3 fraction (PO2) with IC50value of 7.695 μg mL−1, followed by moderate activity of P. herbarum CHCl3 fraction (PH2) that showed an IC50 value of 77.8 μg mL−1. P. oxalicum and P. herbarum total extracts and their other fractions showed weak cytotoxic activity with IC50 values greater than 100 μg mL−1 against Hep-G2 compared to doxorubicin as a positive control with IC50 value of 0.87 μg mL−1 (Table S2, see ESI†).The isolated compounds were tested against colon cancer cell line (Caco-2), and liver cancer (Hep-G2), while doxorubicin was used as a control drug as shown in Table 1. Altersolanol A (6) from P. oxalicum (PO) showed potent cytotoxic activity with IC50 values of 0.00038 ± 0.0003 μM against Hep-G2, and 0.044 ± 0.004 μM against Caco-2. Secalonic acid D (5), also isolated from PO, displayed a potent cytotoxic activity with IC50 values of 0.032 ± 0.0003 μM against Hep-G2, and 0.007 ± 0.001 μM against Caco-2. Another compound from PO, 4-hydroxyphenyl acetic acid (4), showed a moderate cytotoxic activity with IC50 values of 15.51 ± 2.03 μM against Hep-G2 cells, and 38.27 ± 2.20 μM against Caco-2. On the other hand, P. herbarum (PH) isolated compound, mevalonolactone (1) showed weak cytotoxic activity with IC50 values of 919.47 ± 79.76 μM against Hep-G2, and 872.14 ± 126.71 μM against Caco-2 cells. Barcelonyl acetate (2) isolated from PH showed moderate cytotoxic activity with IC50 values of 163.44 ± 12.23 μM against Hep-G2, and 110.90 ± 7.37 μM against Caco-2. Additionally, glycerol monolinoleate (3) from PH showed moderate cytotoxic activity with IC50 values of 166.53 ± 18.23 μM against Hep-G2, and 70.63 ± 8.50 μM against Caco-2. The results were compared with the control drug, doxorubicin, on the same cell lines that exhibited IC50 values of 0.2226 ± 0.06 μM against Hep-G2, and 0.401 ± 0.02 μM against Caco-2. These findings provide significant insights into the potential anti-cancer activity of the isolated compounds, highlighting their varying effectiveness against different cell lines.
| SN | Isolated compound | IC50 (μM) | |
|---|---|---|---|
| Hep-G2 | Caco-2 | ||
| a Hep-G2: Liver Cancer Cell line, Caco-2: Colon Cancer Cell line, Dox.: Doxorubicin was used as a positive control drug. | |||
| 1 | Mevalonolactone | 919.47 ± 79.76 | 872.14 ± 126.71 |
| 2 | Barcelonyl acetate | 163.44 ± 12.23 | 110.90 ± 7.37 |
| 3 | Glycerol monolinoleate | 166.53 ± 18.23 | 70.63 ± 8.50 |
| 4 | 4-Hydroxyphenyl acetic acid | 15.51 ± 2.03 | 38.27 ± 2.20 |
| 5 | Secalonic acid D | 0.032 ± 0.0003 | 0.007 ± 0.001 |
| 6 | Altersolanol A | 0.00038 ± 0.0003 | 0.044 ± 0.004 |
| — | Dox | 0.2226 ± 0.06 | 0.401 ± 0.02 |
3.5. Anti-inflammatory activity
P. herbarium CHCl3 fraction (PH2), n-hexane fraction (PH1), and P. oxalicum CHCl3 fraction (PO2) showed potent nitric oxide inhibitory activity with IC50 values of 26.51, 17.82, and 14.68 μg mL−1, respectively. P. herbarum EtOAc fraction (PH3), in addition to P. oxalicum n-hexane fraction (PO1), showed moderate activity with IC50 values of 76.09 and 65.99 μg mL−1, respectively. However, P. herbarum total extract (PHT) in addition to P. oxalicum total extract (POT) showed weak nitric oxide inhibitory activity with IC50 values greater than 100 μg mL−1 compared to nitro-L-arginine methyl ester hydrochloride (L-NAME) as a positive control that showed IC50 value of 7.316 μg mL−1 (Table S3, see ESI†).3.6. Antimicrobial activity screening
P. herbarum total extract (PHT) and its CHCl3 fraction (PH2) showed antimicrobial activity using the agar well diffusion assay of the tested sample concentration of 20 mg mL−1 that showed inhibition zones ranging from 22 to 38 mm against the tested Gram-positive Staphylococcus aureus ATCC-6538, Gram-negative bacteria Pseudomonas aeruginosa ATCC-9027, and the tested fungal strain Candida albicans ATCC-10231. Moreover, the microtiter plate dilution assay was conducted to confirm the activity that displayed minimum inhibitory concentration (MIC) values ranging from 2 to 4 mg mL−1. However, P. herbarium n-hexane fraction (PH1) showed weak antimicrobial activity with inhibition zones ranging from 12 to 15 mm against the tested organisms and MIC values ranging from 3 to 8 compared to the negative control. While P. oxalicum total extract and all its fractions showed selective moderate antimicrobial activity against Gram-positive S. aureus with inhibition zones ranging from 17 to 29 mm. However, they displayed weak activity against Gram-negative bacteria P. aeruginosa and the tested fungal strain C. albicans, with inhibition zones ranging from 10 to 21 mm of the agar well diffusion assay and MIC values ranging from 4 to 12 mg mL−1 of the microtiter plate dilution assay. Gentamicin was used as a positive control and showed inhibition zones ranging from 34 to 40 mg mL−1 in the agar well diffusion assay and MIC values of 0.003125 to 0.0125 mg mL−1 in the microtiter plate turbidity assay (Table S4, see ESI†).3.7. Antibiofilm activity
P. herbarum n-hexane fraction (PH1) and CHCl3 (PH2) fraction, in addition to P. oxalicum n-hexane fraction (PO1), showed antibiofilm inhibitory activity against Pseudomonas aeruginosa ATCC-9027 in a dose-dependent manner and showed minimum biofilm inhibitory concentration (MBIC) values of 1.25, 5, and 2.5 mg mL−1. However, the total extracts of P. herbarum and P. oxalicum showed weak activity with MBIC values greater than 10 mg mL−1 (Table S4, see ESI†).3.8. Antidiabetic activity
P. herbarium CHCl3 fraction (PH2) showed α-glucosidase enzyme inhibitory activity in a dose-dependent manner and displayed IC50 values of 14.91 μg mL−1. Followed by the moderate activity of P. herbarum total extract (PHT) and its EtOAc fraction (PO3) that showed IC50 of 79.17 and 148.5 μg mL−1, respectively. P. herbarum n-hexane fraction in addition to P. oxalicum total extract (POT) and all its thereof fractions showed weak activity with IC50 values greater than 1000 μg mL−1 compared to acarbose as a positive control that showed IC50 value of 224.0 μg mL−1. However, the CHCl3 fraction (PO2) of P. oxalicum showed an inhibitory percentage of 46.16 ± 1.32% at the test's highest concentration of 1000 μg mL−1 (Table S5, see ESI†).3.9. Chemical characterization of P. herbarum chloroform fraction (PH2) and P. oxalicum chloroform fraction (PO2)
P. herbarum CHCl3 fraction (PH2) and chemical contestants were tentatively identified using LC/MS-positive-ESI analysis and compared with spectral data in the literature (Table S6 and Fig. S5, see ESI†). Thirteen compounds were identified as mevalonolactone (1), phomasparapyrone A, 8-hydroxy-pregaliellalactone B, phomactin B, tersone F, phomaether A, barcelonyl acetate (2), barceloneic acid C, terezine N, terezine L, alterporriol S, phomalide, glycerol monolinoleate (3), and ergosterol (7). The identified compounds exhibit a broad range of chemical structures, encompassing lactones, pyrone derivatives, polyketides, diterpenes, pyridine alkaloids, diphenyl ethers, terazine derivatives, pyrazine derivatives, anthraquinone derivatives, cyclic polypeptide derivatives, glycerides, and sterol. On the other hand, the investigation of the chemical constituents of P. oxalicum CHCl3 fraction (PO2) showed thirteen identified compounds as mevalonolactone (1), 4-hydroxyphenyl acetic acid (4), 1,3-dihydroxypropan-2-yl 2,4-dihydroxy-6-methylbenzoate, 2-(4-hydroxybenzyl) quinazolin-4(3H)-one, penipanoid A, 2,5-dimethyl-7-hydroxychromone, coniochaetone J, meleagrin, meleagrin A; Me ether, penioxamide A, secalonic acid D (5), altersolanol A (6), and ergosterol (7). The compounds represented a wide range of structural classes including pyrone derivatives, phenolic acids, alkaloids, chromanone derivatives, xanthone derivatives, sterols, and anthraquinone derivatives (Table S7 and Fig. S5, see ESI†). This diversity suggests a wide array of biosynthetic pathways operating within these fungi. The structures of the major identified fungal metabolites are displayed in Fig. 1 and 2.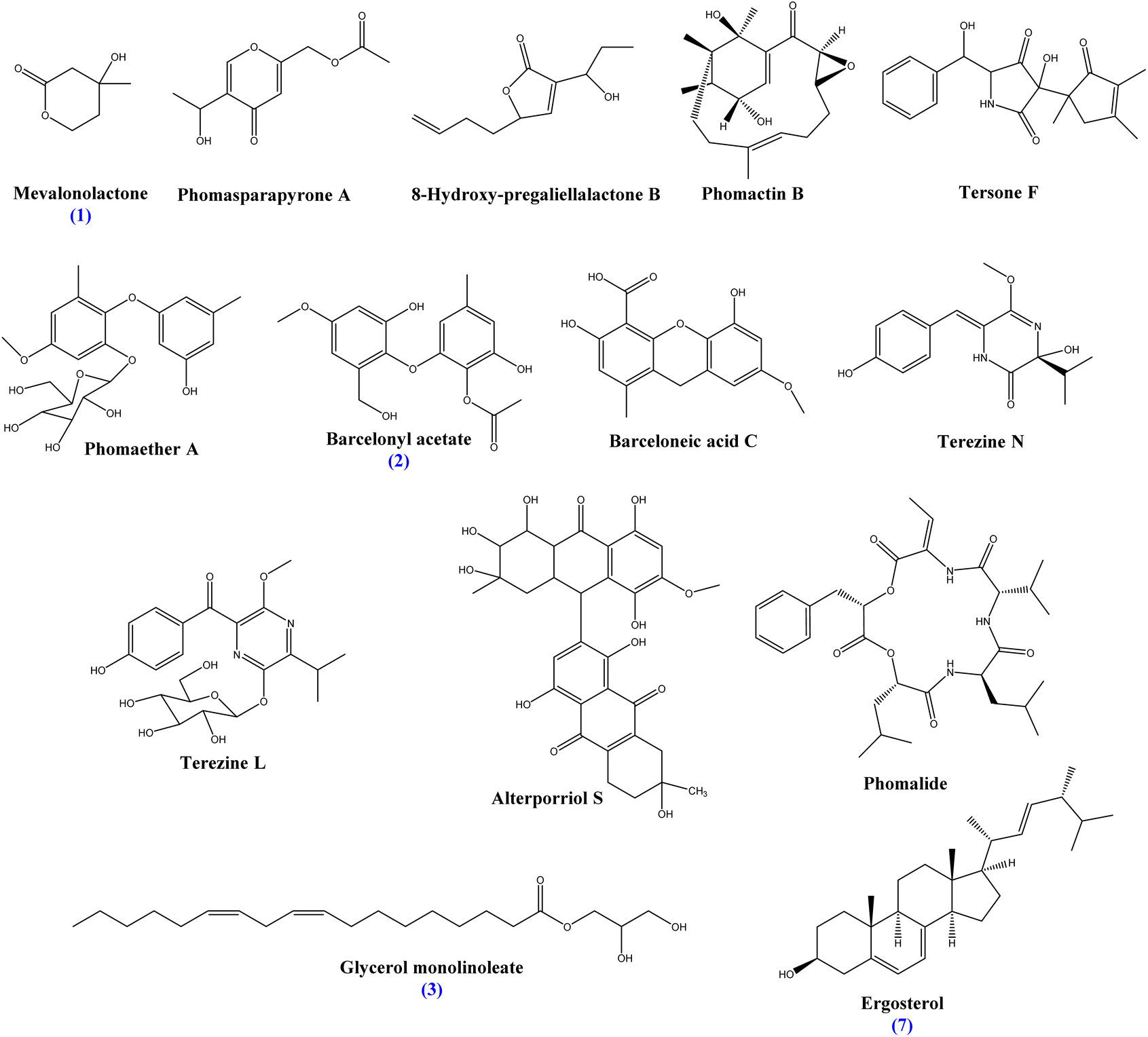 | ||
| Fig. 1 Structure of the main identified constituents of P. herbarum chloroform fraction. | ||
 | ||
| Fig. 2 Structure of the main identified constituents of P. oxalicum chloroform fraction. | ||
3.10. Identification of isolated compounds of fungal chloroform fractions
To explore the active metabolites present in the identified fungi, the total extracts were purified, isolating three known compounds: mevalolactone (1), glycerol monolinoleate (3), and ergosterol, in addition to one new barcelonic acid derivative namely barcelonyl acetate (2) from P. herbarum chloroform fraction, and four known compounds: 4-hydroxyphenyl acetic acid (4), secalonic acid D (5), altersolanol A (6), and ergosterol (7) from P. oxalicum. These compounds were identified based on their spectral NMR data, and a comparison was made with compounds previously reported in the literature.Mevalonolactone (1): was separated as a yellow oil from P. herbarum chloroform fraction, yielding 14.5 mg of pale yellow oil, which was confirmed by reported NMR data52,53 (ESI: Fig. S6–S9†). Mevalonolactone was previously reported from Pestalotiopsis sp. & Penicillium solitum.54,55 This is the first time to isolate mevalonolactone from P. herbarum associated with M. nigra.
Barcelonyl acetate (2): was separated as a yellow oil from P. herbarum chloroform fraction, yielding 10 mg as yellowish oil, and was confirmed by the reported data of a closer structure, namely 8-O-methyl barceloneate, which are both derived from barceloneic acid A as described in Table 2.56,57 The acetyl group was confirmed by the HMBC correlation of C-8 at δc = 170 Hz with the 3H protons of carbon 9 at δH = 2.0 Hz. The 1HNMR, attached proton test (APT), and 2D NMR data confirmed the structure is a new compound, namely, 3-hydroxy-1-(2′-hydroxy-6′-(hydroxymethyl)-4′-methoxyphenoxy)-5-methylphenyl acetate, which is abbreviated to barcelonyl acetate (ESI: Fig. S10–S14†). This is the first time to report barcelonyl acetate from P. herbarum associated with M. nigra. Moreover, this is the first time to report this compound from nature (Fig. 3).
| Position | Barcelonyl acetate (2) | 8-O-Methyl barceloneate56 | ||
|---|---|---|---|---|
| δC (CDCl3, 100 MHz) | δH (CDCl3, 400 MHz, J in Hz) | δC (125 MHz, CDCl3) | δH (500 MHz, CDCl3) | |
| 1 | 158.62 | — | 158.1 | — |
| 2 | 99.43 | — | 100.9 | — |
| 3 | 163.96 | — | 162.8 | — |
| 4 | 113.62 | 6.65 (d, J = 4.5, 1H) | 112.6 | 6.51, d, (1.5) |
| 5 | 147.60 | — | 147.2 | — |
| 6 | 104.60 | 5.94 (d, J = 7.1, 1H) | 105.8 | 5.94, d, (1.5) |
| 7 | 22.19 | 2.21, (s, 3H) | 22.1 | 2.17, s |
| 8 | 170.45 | — | 170.2 | — |
| 9 | 55.73 | 2.0, (s, 3H) | 52.8 | 3.99, s |
| 1′ | 130.36 | — | 132.7 | — |
| 2′ | 149.59 | — | 149.3 | — |
| 3′ | 103.37 | 6.57 (d, J = 11.0, 1H) | 101.8 | 6.57, d, (3.0) |
| 4′ | 157.31 | — | 157.9 | — |
| 5′ | 107.91 | 6.57 (d, J = 11.0, 1H) | 105.8 | 6.59, d, (3.0) |
| 6′ | 131.76 | — | 135.0 | — |
| 7′ | 61.61 | 4.94 (s, 2H) | 61.0 | 4.50, s |
| 8′ | 55.73 | 3.84 (s, 3H) | 55.6 | 3.82, s |
 | ||
| Fig. 3 Structure of the new isolated compound barcelonyl acetate (2) compared to closer molecules barceloneic acid A and 8-O-methyl barceloneate. | ||
Glycerol monolinoleate (3): was separated as a colorless oil from P. herbarum chloroform fraction of yield 45.9 mg, which was confirmed by reported NMR data58 (ESI: Fig. S15–S19†).
4-Hydroxyphenyl acetic acid (4): was separated as a white crystalline powder of P. oxalicum chloroform fraction of yield 39.7 mg, which was confirmed by reported NMR data on 4-hydroxyphenyl acetic acid59 (ESI: Fig. S20–S24†).
Secalonic acid D (5): it was separated as yellow gum of P. oxalicum chloroform fraction of yield 24.8 mg, which was confirmed by reported NMR data on secalonic acid D60 (ESI: Fig. S25–S29†).
Altersolanol A (6): it was separated as orange crystals of P. oxalicum chloroform fraction of yield 8 mg, which was confirmed by reported NMR data61 (ESI: Fig. S30–S34†). This compound was previously reported from Stemphylium globuliferum.61,62 However, to our knowledge, it is the first time to report altersolanol A from P. oxalicum.
Ergosterol (7): was separated as colorless crystals from both P. herbarum and P. oxalicum chloroform fractions of yield 83.6 mg and 15.2 mg, respectively, which was confirmed by reported NMR data on ergosterol (ESI: Fig. S35–S36†).63,64
3.11. Molecular docking
JNK2 plays a multifaceted role in both cancer and inflammation. In some cancers, JNK2 acts as a tumor suppressor, promoting cell death and hindering the spread of cancer cells. However, in other cancers, JNK2 can encourage tumor growth. This complex relationship highlights the need for further research to understand the specific context in which JNK2 functions in different cancers. Similarly, JNK2 can be both anti-inflammatory and pro-inflammatory depending on the tissue and cell type. It can help maintain gut health and reduce inflammation, but also contribute to inflammatory processes in immune cells. Due to this complex interplay, targeting JNK2 with inhibitors holds promise for cancer and inflammatory disease treatment. If researchers can develop inhibitors that specifically target JNK2's pro-cancerous or pro-inflammatory functions, it could lead to novel therapeutic strategies. However, the challenge lies in creating inhibitors that are specific and don't disrupt JNK2's potentially beneficial functions in other contexts.This molecular docking investigation studied the potential of several naturally isolated compounds as inhibitors of c-Jun N-terminal kinase 2 (JNK2) using a docking approach. The co-crystal inhibitor, BIRB796, served as a reference for evaluating the interaction patterns with JNK2. Identifying compounds that mimic BIRB796's interactions is crucial for discovering potent JNK2 inhibitors. BIRB796 forms a well-defined interaction profile with JNK2, including hydrogen bonds with key residues Met111, Glu73, and Asp169. It also engages in extensive pi–pi interactions with aromatic amino acids like Val40, Ala53, and Phe170, and a pi–sulfur interaction with Met108. These interactions provide a benchmark for assessing the binding potential of the natural compounds.
Among the evaluated compounds, 4-hydroxyphenyl acetic acid (4) displayed common interactions with BIRB796. It forms a hydrophobic interaction with Val40 and Lys55, suggesting favourable binding affinity. Additionally, altersolanol A (6) presented a compelling profile with hydrogen bonds matching BIRB796 and additional interactions with Lys55 and Gly171, potentially enhancing binding. These findings highlight altersolanol A (6) as a promising candidate for further development as JNK2 inhibitors. Secalonic aid D (5) failed in the docking as it couldn't fit the pocket of the receptor for molecular docking as it is a large molecule of molecular weight 638. Meanwhile, mevalonolactone (1) exhibited fewer interactions compared to other compounds. While it formed hydrogen bonds with Asp169 and Lys55, it lacked extensive pi–pi interactions, suggesting a need for optimization to improve its binding affinity and efficacy as a JNK2 inhibitor. Barcelonyl acetate (2) displayed robust interaction capabilities, mimicking BIRB796's pi–anion bonds with Asp169, and Arg72. It also shared key pi–alkyl interactions with Arg150 and pi–pi interactions, suggesting a strong binding potential similar to BIRB796. These findings warrant further investigation and development of barcelonyl acetate (2) as a JNK2 inhibitor. Meanwhile, glycerol monolinoleate (3) presented the most comprehensive interaction profile. It formed a hydrogen bond with Met111 and engaged in multiple pi–pi interactions with residues crucial for BIRB796 binding. This extensive overlap suggests a high binding affinity and strong potential for JNK2 inhibition. The molecular docking interactions of the investigated compounds are outlined in Table 3. The 2D and 3D interaction figures are provided in (ESI Table S8†).
| Compound | Docking score | Interaction type | Interacting residues |
|---|---|---|---|
| Co crystal (BIRB796) | −9.687 | Hydrogen bond | Met111, Glu73, Asp169 |
| Pi–pi | Val40, Ala53, Lys55, Arg69, Leu77, Ile85, Leu110, Leu142, Ile147, His149, Val158, Phe170 | ||
| Pi–sulfur | Met108 | ||
| Mevalonolactone (1) | −5.998 | Hydrogen bond | Asp169, Lys55 |
| Pi–pi | Leu168, Ile86 | ||
| Barcelonyl acetate (2) | −6.769 | Pi–anion | Asp169, Arg72 |
| Pi–alkyl | Arg150 | ||
| Pi–pi | Ala173, Gln37, Gly171, Arg69, Arg192, Asp151, His149, Ile147, Ile148, Leu76 | ||
| Glycerol monolinoleate (3) | −6.719 | Hydrogen bond | Met111 |
| Pi–pi | Val40, Leu77, Lys55, Ile85, Ile86, Val80, Leu168, Phe170 | ||
| 4-Hydroxyphenyl acetic acid (4) | −5.919 | Pi–alkyl | Lys55, Leu168, Val40, Met108 |
| Altersolanol A (6) | −7.094 | Hydrogen bond | Asp169, Glu73, Lys55, Gly171, Gln73, Arg69 |
| Pi–pi | Ile85, Leu77, Ile86 |
4. Conclusions
The study reports for the first time the isolation and identification of two fungal endophytes, Phoma herbarum and Penicillium oxalicum, from Morus nigra and Ficus sycomorus, respectively. Biological evaluation of the fungal extracts and fractions revealed a range of activities, with the chloroform fractions generally exhibiting the most potent effects. P. herbarum chloroform fraction demonstrated potent antioxidant and antidiabetic potential. While P. oxalicum chloroform fraction showed potent cytotoxic activity.Through bioactivity-guided isolation, seven compounds were obtained and identified from the active chloroform fractions. Barcelonyl acetate (2) was reported for the first time from nature from P. herbarum that showed moderate cytotoxic activity. In addition, altersolanol A (6) and secalonic acid D (5) from P. oxalicum exhibited the strongest cytotoxicity against various cancer cell lines. Molecular docking identified glycerol monolinoleate as a promising JNK2 inhibitor, while altersolanol A (6) and barcelonyl acetate (3) also showed favourable interaction profiles. Key residues involved in hydrogen bonding and pi–pi interactions with the compounds were determined. The bioactive potential demonstrated by P. herbarum and P. oxalicum extracts and fractions, as well as the isolated compounds, warrants further investigation of their mechanisms and therapeutic applications. The in silico findings provide valuable leads for developing novel JNK2 inhibitors. Future work will optimize derived hits for the discovery of bioactive natural products.
Data availability
All data generated or analyzed during this study are included in this published article (and its additional ESI file†).Author contributions
Conceptualization, ANBS and MAE; methodology, MMM; validation, ANBS, MME, NMM, AYM, and AME; formal analysis, ANBS, AME; investigation, MMA and AME; resources, MMA and AME; data curation, MMA and AME; writing – original draft preparation, MMA; writing – review and editing, MMA, AME, and ANBS; visualization, ANBS and AME; supervision, ANBS, MAE, NMM, AYM, and AME; project administration, ANBS. All authors have read and agreed to the published version of the manuscript.Conflicts of interest
The authors declare that they have no conflicts of interests.Acknowledgements
Abdel Nasser Singab is thankful to Egyptian Science and Technology Development Funds (STDF) for their financial support through grant No. 46667 entitled “Sustainability of Lab. Capacities of the Center of Drug Discovery Research and Development.”Notes and references
- M. A. Hossain, J. King Saud Univ., 2019, 31, 961–965 CrossRef.
- S. H. Lim and C.-I. Choi, Nutrients, 2019, 11, 437 CrossRef CAS PubMed.
- L.-W. Wang, Y.-L. Zhang, F.-C. Lin, Y.-Z. Hu and C.-L. Zhang, Mini Rev. Med. Chem., 2011, 11, 1056–1074 CrossRef CAS.
- S. S. El-hawary, A. S. Moawad, H. S. Bahr, U. R. Abdelmohsen and R. Mohammed, RSC Adv., 2020, 10, 22058–22079 RSC.
- J. F. Imhoff, Mar. Drugs, 2016, 14, 19 CrossRef.
- A. N. B. Singab, N. M. Mostafa, Y. A. Elkhawas, E. Al-Sayed, M. M. Bishr, A. M. Elissawy, M. S. Elnaggar, I. M. Fawzy, O. M. Salama and Y.-H. Tsai, Mar. Drugs, 2022, 20, 331 CrossRef CAS.
- M. M. M. AbdelRazek, A. M. Elissawy, N. M. Mostafa, A. Y. Moussa, M. A. Elanany, M. A. Elshanawany and A. N. B. Singab, Molecules, 2023, 28, 1718 CrossRef CAS PubMed.
- J. H. Song, C. Lee, D. Lee, S. Kim, S. Bang, M.-S. Shin, J. Lee, K. S. Kang and S. H. Shim, J. Nat. Prod., 2018, 81, 1411–1416 CrossRef CAS PubMed.
- L. P. Zheng, Z. Zhang, L. Q. Xie, H. Y. Yuan and Y. Q. Zhang, Adv. Mater. Res., 2013, 642, 615–618 Search PubMed.
- S. Bang, H. Eun, J. Yun, D. Sik, S. Kim, S. Nam, D. Lee, K. Sung and S. Hee, Bioorg. Chem., 2020, 105, 104449 CrossRef CAS.
- H. G. Choi, J. H. Song, M. Park, S. Kim, C. Kim, K. S. Kang and S. H. Shim, Biomolecules, 2020, 10, 91 CrossRef CAS.
- H. Ku, J. Baek, K. S. Kang and S. H. Shim, Nat. Prod. Res., 2021, 0, 1–7 Search PubMed.
- J. Vora, S. Velhal, S. Sinha, V. Patel and N. Shrivastava, HIV Med., 2021, 22, 690–704 CrossRef CAS PubMed.
- R. Ayesha and T. Iftikhar, Pakistan J. Bot., 2010, 42, 583–592 Search PubMed.
- A. Aparecida, J. C. Polonio, A. M. Bulla, D. Polli, J. C. Castro, L. C. Soares, V. A. De, V. Elisa, P. Vicentini, A. José, B. De Oliveira, J. E. Gonçalves, R. Aparecida, C. Gonçalves, J. Lúcio, B. A. De Abreu-filho, J. A. Pamphile, A. Aparecida, J. C. Polonio, A. M. Bulla, D. Polli, J. C. Castro, L. C. Soares, V. A. De Oliveira-junior, V. Elisa, P. Vicentini, A. José, B. De Oliveira and J. E. Gonçalves, Nat. Prod. Res., 2021, 36, 3158–3162 Search PubMed.
- E. Hermawati, L. D. Juliawaty and E. H. Hakim, Rec. Nat. Prod., 2017, 11, 315–317 CAS.
- J. Lukša and E. Servienė, J. Environ. Eng. Landsc. Manag., 2022, 28, 183–191 CrossRef.
- E. Hermawati, S. D. Ellita, L. D. Juliawaty, E. H. Hakim, Y. M. Syah and H. Ishikawa, J. Nat. Med., 2021, 75, 217–222 CrossRef CAS PubMed.
- A. S. Ratnayake, W. Y. Yoshida, S. L. Mooberry and T. K. Hemscheidt, J. Org. Chem., 2001, 66, 8717 CrossRef CAS PubMed.
- A. D. Allen, B. Cheng, M. H. Fenwick, B. Givehchi, H. Henry-riyad and A. Valerij, J. Org. Chem., 2003, 68, 1640 CrossRef CAS.
- E. V. Patridge, A. Darnell, K. Kucera, G. M. Phillips, H. R. Bokesch, K. R. Gustafson, D. J. Spakowicz, L. Zhou, W. M. Hungerford, M. Plummer, D. Hoyer, A. Narváez-Trujillo, A. J. Phillips and S. A. Strobel, Nat. Prod. Commun., 2020, 10, 1649–1654 Search PubMed.
- Z. Sun, F. Liang, Y. Chen, H. Liu and H. Li, J. Asian Nat. Prod. Res., 2016, 6020, 0–6 Search PubMed.
- Z. Sun, H. Li, F. Liang, Y. Chen, H. Liu and S. Li, Molecules, 2016, 21, 1–6 Search PubMed.
- H. C. Zhang, Y. M. Ma, R. Liu and F. Zhou, Biochem. Syst. Ecol., 2012, 45, 31–33 CrossRef CAS.
- X. Liang, H. Zhang and R. Liu, J. Agric. Food Chem., 2016, 64, 7–11 Search PubMed.
- H. Zhang, Y. Ma and R. Liu, Appl. Mech. Mater., 2012, 181, 783–786 Search PubMed.
- X. A. Liang, Y. M. Ma, H. C. Zhang and R. Liu, Nat. Prod. Res., 2016, 6419, 2407–2412 CrossRef PubMed.
- Z. Ding, T. Tao, L. Wang, Y. Zhao, H. Huang, D. Zhang, M. Liu, Z. Wang and J. Han, J. Microbiol. Biotechnol., 2019, 29, 731–738 CrossRef CAS.
- K. K. Jayant and B. S. Vijayakumar, Ital. J. Mycol., 2021, 50, 10–20 Search PubMed.
- A. Y. Moussa, N. M. Mostafa and A. N. B. Singab, Nat. Prod. Res., 2019, 0, 1–8 Search PubMed.
- S. M. Shama, A. M. Elissawy, M. A. Salem, F. S. Youssef, M. S. Elnaggar, H. R. El-Seedi, S. A. M. Khalifa, K. Briki, D. I. Hamdan and A. N. B. Singab, RSC Adv., 2024, 14, 18553–18566 RSC.
- M. S. Elnaggar, A. M. Elissawy, F. S. Youssef, M. Kicsák, T. Kurtán, A. N. B. Singab and R. Kalscheuer, RSC Adv., 2023, 13, 16480–16487 RSC.
- R. Boly, T. Lamkami, M. Lompo, J. Dubois and P. Guisson, Int. J. Clin. Pharmacol. Res., 2016, 8, 29–34 Search PubMed.
- Z. Chen, R. Bertin and G. Froldi, Food Chem., 2013, 138, 414–420 CrossRef CAS PubMed.
- D. N. Jamalova, H. A. Gad, D. K. Akramov, K. S. Tojibaev, N. M. Al Musayeib, M. L. Ashour and N. Z. Mamadalieva, Plants, 2021, 10, 1529 CrossRef PubMed.
- P. Skehan, R. Storeng, D. Scudiero, A. Monks, J. McMahon, D. Vistica, J. T. Warren, H. Bokesch, S. Kenney and M. R. Boyd, JNCI, J. Natl. Cancer Inst., 1990, 82, 1107–1112 CrossRef CAS PubMed.
- R. M. Allam, A. M. Al-Abd, A. Khedr, O. A. Sharaf, S. M. Nofal, A. E. Khalifa, H. A. Mosli and A. B. Abdel-Naim, Toxicol. Lett., 2018, 291, 77–85 CrossRef CAS.
- T. Oliveira, C. A. Figueiredo, C. Brito, A. Stavroullakis, A. Prakki, E. Da Silva Velozo and G. Nogueira-Filho, Int. J. Cell Biol., 2014, 2014, 535789 Search PubMed.
- M. S. Yoo, J. S. Shin, H. E. Choi, Y. W. Cho, M. H. Bang, N. I. Baek and K. T. Lee, Food Chem., 2012, 135, 967–975 CrossRef CAS PubMed.
- M. M. M. A. Razek, A. Y. Moussa, M. A. El-Shanawany and A. B. Singab, Int. J. Pharm. Sci. Res., 2019, 11, 3289–3296 Search PubMed.
- F. Barchiesi, J. Bille, E. Chryssanthou, D. Denning, J. P. Donnelly, B. Dupont, W. Fegeler, C. Moore, M. Richardson, P. E. Verweij, K. Hospital, N. Manchester, G. Hospital and H. Hospital, Clin. Microbiol. Infect., 2003, 9, 1–8 Search PubMed.
- C. Rodr, C. Alonso-calleja, C. Garc, J. Carballo and R. Capita, Biology, 2022, 11, 46 Search PubMed.
- R. Mogana, A. Adhikari, M. N. Tzar, R. Ramliza and C. Wiart, BMC Complement. Med. Ther., 2020, 20, 1–11 CrossRef.
- M. M. M. AbdelRazek, A. Y. Moussa, M. A. El-shanawany and A. B. Singab, Chem. Biodivers., 2020, 17, 1–9 Search PubMed.
- R. Subramanian, M. Z. Asmawi and A. Sadikun, Acta Biochim. Pol., 2008, 55, 391–398 CrossRef CAS.
- H. M. Abdallah, A. T. Kashegari, A. A. Shalabi, K. M. Darwish, A. M. El-Halawany, M. M. Algandaby, S. R. M. Ibrahim, G. A. Mohamed, A. B. Abdel-Naim, A. E. Koshak, P. Proksch and S. S. Elhady, Biology, 2022, 11, 762 CrossRef CAS PubMed.
- S. Saeed Kotb, I. M. Ayoub, S. A. El-Moghazy and A. N. B. Singab, Nat. Prod. Res., 2022, 1–6 Search PubMed.
- S. H. Abdallah, N. M. Mostafa, M. A. E. H. Mohamed, A. S. Nada and A. N. B. Singab, Nat. Prod. Res., 2022, 36, 5619–5625 CrossRef CAS PubMed.
- J. J. Irwin and B. K. Shoichet, J. Chem. Inf. Model., 2005, 45, 177–182 CrossRef CAS PubMed.
- A. Kuglstatter, M. Ghate, S. Tsing, A. G. Villaseñor, D. Shaw, J. W. Barnett and M. F. Browner, Bioorg. Med. Chem. Lett., 2010, 20, 5217–5220 CrossRef CAS PubMed.
- H. Nada, S. Kim, S. Godesi, J. Lee and K. Lee, J. Biomol. Struct. Dyn., 2023, 41, 11904–11915 CrossRef CAS PubMed.
- M. Kishida, N. Yamauchi, K. Sawada, Y. Ohashi, T. Eguchi and K. Kakinuma, J. Chem. Soc., Perkin Trans. 1, 1997, 891–895 RSC.
- Y. Ying, L. Zhang, W. Shan and Z.-J. Zhan, Chem. Nat. Compd., 2014, 50, 628–629 CrossRef.
- Z. Xu, X. Wu, G. Li, Z. Feng and J. Xu, Nat. Prod. Res., 2020, 34, 1002–1007 CrossRef CAS PubMed.
- Z.-H. He, J. Wu, L. Xu, M.-Y. Hu, M.-M. Xie, Y.-J. Hao, S.-J. Li, Z.-Z. Shao and X.-W. Yang, Mar. Drugs, 2021, 19, 580 CrossRef CAS.
- D. P. Overy, T. O. Larsen, P. W. Dalsgaard, K. Frydenvang, R. Phipps, M. H. G. Munro and C. Christophersen, Mycol. Res., 2005, 109, 1243–1249 CrossRef CAS PubMed.
- Y. Tan, Y. Wang, Q. Li, X. Xing, S. Niu, B. Sun, L. Chen, R. Pan and G. Ding, Phytochemistry, 2022, 201, 113264 CrossRef CAS PubMed.
- E. Okuyama, T. H. Asegawa, T. M. Atsushita, H. F. Ujimoto and M. I. Shibashi, Chem. Pharm. Bull., 2001, 49, 154–160 CrossRef CAS PubMed.
- J. E. Milne, T. Storz, J. T. Colyer, O. R. Thiel, M. Dilmeghani Seran, R. D. Larsen and J. A. Murry, J. Org. Chem., 2011, 76, 9519–9524 CrossRef CAS PubMed.
- L. Chen, Y. X. Bi, Y. P. Li, X. X. Li, Q. Y. Liu, M. G. Ying, Q. H. Zheng, L. Du and Q. Q. Zhang, Heterocycles, 2017, 94, 1766–1774 CrossRef CAS.
- A. Aly, A. Debbab, R. Edrada-Ebel, W. Muller, M. Kubbutat, V. Wray, R. Ebel and P. Proksch, Mycosphere, 2010, 1, 153–162 Search PubMed.
- M. Teiten, F. Mack, A. Debbab, A. H. Aly, M. Dicato, P. Proksch and M. Diederich, Bioorg. Med. Chem., 2013, 21, 3850–3858 CrossRef CAS PubMed.
- A. Haque, M. S. Hossain, M. Z. Rahman, M. R. Rahman, S. Hossain, M. Mosihuzzaman, N. Nahar and S. I. Khan, J. Pharm. Sci., 2005, 4, 127–130 Search PubMed.
- R. X. Receptor, C. Xie, D. Zhang, J. Xia, C. Hu, T. Lin, Y. Lin, G. Wang, W. Tian, Z. Li, X. Zhang, X. Yang, H. Chen and M. Drugs, Mar. Drugs, 2019, 17, 178 CrossRef.
Footnote |
| † Electronic supplementary information (ESI) available: Fig. S1–S36 and Tables S1–S8. See DOI: https://doi.org/10.1039/d4ra06840h |
| This journal is © The Royal Society of Chemistry 2024 |