
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceUnravelling the potential of yolkin for nutraceutical use: the origin, structure, and functional insights of a hen egg yolk polypeptide complex†
Aleksandra
Zambrowicz
a,
Katarzyna
Kapczyńska
b,
Paweł
Kania
c,
Jakub Stanisław
Nowak
d,
Marta
Kaszowska
b,
Katarzyna
Szymczak-Kulus
 b,
Wioletta
Kazana-Płuszka
b,
Marta
Piksa
b,
Sabina
Górska
b,
Dominika
Jakubczyk
b,
Józefa
Macała
b and
Agnieszka
Zabłocka
*b
b,
Wioletta
Kazana-Płuszka
b,
Marta
Piksa
b,
Sabina
Górska
b,
Dominika
Jakubczyk
b,
Józefa
Macała
b and
Agnieszka
Zabłocka
*b
aDepartment of Functional Food Products Development, Wroclaw University of Environmental and Life Sciences, Chelmonskiego 37, 51-640 Wroclaw, Poland
bHirszfeld Institute of Immunology and Experimental Therapy, Polish Academy of Sciences, Weigla 12, 53-114 Wroclaw, Poland. E-mail: agnieszka.zablocka@hirszfeld.pl; Tel: +48 71 337 11 72
cNanotempertech, Bobrzyńskiego 14, 30-348, Cracow, Poland
dMalopolska Centre of Biotechnology, Jagiellonian University, Gronostajowa 7a, 30-387 Cracow, Poland
First published on 10th October 2024
Abstract
Nutraceuticals can reduce the risk of many diseases, such as cardiovascular disease, immune deficiencies, neurodegeneration, and others. Their delivery remains a challenge because it depends on many factors, most notably the stability of the bioactive compounds. Yolkin is a peptide complex isolated from hen egg yolk with immunomodulatory and neuroprotective potential. However, yolkin remains relatively poorly characterized. We aimed to determine the origin and glycosylation level of yolkin, its storage conditions, its thermal stability, and its aggregation ability and to assess its antioxidant, antihypertensive, and antidiabetic potential. The peptide composition of yolkin was shown to be homologous to that of vitellogenin II and vitellogenin I. These results indicate the stability of yolkin in a lyophilized form, preferably at 4 °C, with nonaggregation, antioxidant, and antidiabetic activities. As a result, yolkin can be considered to have significant therapeutic potential and represents a valuable tool for the development of novel nutraceuticals.
1. Introduction
Hen egg yolk is a rich reservoir of lipids, proteins, minerals, and vitamins necessary for bird embryo development.1–4 Fractionation of hen egg yolk diluted with water by ultracentrifugation produces two fractions: granules and plasma.4–6 Granules contain mainly high-density lipoproteins (HDLs) such as α-lipovitellins and β-lipovitellins, phosvitins, and low-density lipoproteins (LDLs). Within plasma, there is a wide variety of proteins, including immunoglobulins (>45%), albumins (14%), glycoproteins such as yolk glycopeptide (YGP) 42 and YGP40 (41%), and the newfound protein YGP30, with 75% similarity to YGP40.5,7 The main protein precursor of the granular fraction is vitellogenin.5,8–10 In response to estrogen circulating in the blood, the liver produces vitellogenin, which is released into the bloodstream and is taken up by growing oocytes, where it is enzymatically converted into a variety of egg yolk proteins. Moreover, vitellogenin is characterized by a high degree of sequence homology and diverse biological activities, irrespective of the source organism.3,10–12Despite the well-characterized bioactive compounds found in egg white and yolk, little is known about the proteins in egg yolk plasma besides immunoglobulin Y (IgY). In 2012, Polanowski et al.12 reported that the IgY fraction obtained from chicken egg yolk is a complex with protein fragments called yolkin, characterized by immunoregulatory properties. Yolkin is a mixture of proteins, polypeptides, and peptides abundant in acidic amino acid residues with a small amount of methionine. The molecular weight of yolkin peptides varies from 1 to 35 kDa, but those in the 16–35 kDa range are the most abundant. The N-terminal amino acid sequences of the electrophoretically purified yolkin components, determined by Edman degradation, showed homology to the vitellogenin II C-terminal domain. The fractions with a molecular weight below 12 kDa corresponded to the sequence of vitellogenin II starting from position 1732 aa. In turn, the fractions with a molecular weight over 16 kDa corresponded to the amino acid sequence of vitellogenin II at position 1572.13 Notably, the presence of vitellogenin in the yolks of different bird species is not the only factor influencing the protein profile of the yolkin complex. Depending on various factors, such as the time of egg collection or the specific pen used, the ratio of polypeptides and proteins can vary significantly.14
Our studies to date have shown that yolkin is nontoxic to cells and has potent immunomodulatory, antioxidative, and neuroprotective effects. Yolkin stimulates human whole blood (ex vivo) to secrete a broad range of cytokines, both pro- and anti-inflammatory;13,15–19 downregulates the level of intracellular oxygen radicals;20 and affects the formation of the immune response and the cell phenotype in lymphoid organs.21 Recent evidence also indicates that yolkin delivered in ovo influences the leukocyte population profile and cytokine levels in broiler chickens. The most significant effect of yolkin is an elevation of the T lymphocyte populations, including both CD4+ and CD8+ subsets, particularly in the blood.22 Furthermore, yolkin stimulates neurons to express and secrete the neurotrophin brain-derived neurotrophic factor (BDNF)23 and improves cognitive function in aged rats.24 These data highlight that yolkin can act in a broad and multidirectional manner and may be involved in regulating the survival and activity of immune and neuronal cells. Therefore, it would seem that yolkin can potentially be employed in the future as a safe, bioavailable, natural nutraceutical to enhance immunity and cognition.
There is currently a trend towards consuming foods rich in bioactive compounds that may reduce the risk of acute and chronic diseases such as cardiovascular diseases, cancer, immune deficiencies, neurodegenerative diseases, obesity, and others.25–27 Compounds derived from foods and food raw materials with nutritional and therapeutic effects are defined as nutraceuticals by combining the words “nutritional” and “pharmaceutical”.28 Nutraceuticals are “food or parts of food that provide health benefits, including the prevention and treatment of diseases”.28 In China, nutraceuticals are governed by legislation and can function as an alternative and complementary strategy for treating and preventing specific diseases. Meanwhile, in the United States of America and the European Union, nutraceuticals are classified and regulated as dietary supplements.25 Nutraceuticals comprise a variety of different chemical classes including polyphenols, omega-3 fatty acids, omega-6 polyunsaturated fatty acids (PUFAs), terpenoids, alkaloids, carotenoids and nitrogenous compounds such as proteins, polypeptides, peptides and amino acids.25–27 It is necessary to demonstrate the efficacy, safety, and lack of toxic effects of nutraceuticals and dietary supplements. Nutraceutical research, therefore, focuses on testing (in vitro and in vivo) the bioactivity of potential nutraceuticals, developing and standardizing methods to assess the mechanisms involved in nutraceutical bioavailability, and selecting food matrices and nutraceutical delivery systems to improve bioavailability after oral administration.25,26 Nutraceuticals are consumed both as dietary supplements (pills, syrups, powders, capsules) and as functional foods enriched with nutraceuticals in the form of free or encapsulated bioactive compounds. The form of administration is determined by the stability of the bioactive compounds under different conditions, such as temperature, pH, and other physicochemical factors, used in the production of dietary supplements or functional foods.26,27
Yolkin is still an undercharacterized substance, particularly because of its practical use. The proper characterization and stability of nutraceuticals, especially those in the form of peptide complexes, are essential for maintaining their therapeutic efficacy. However, the exposure of nutraceuticals to low or high temperatures, either during storage or transport, could reduce their efficacy.13 The improper storage of nutraceuticals at undesirable temperatures is one of the main factors contributing to the reduced efficacy of the active ingredient. Therefore, this study aimed to determine the storage conditions of yolkin, its thermal stability, and its ability to form aggregates. The amino acid sequence of yolkin was determined by mass spectrometry (MS), and the results were compared with those obtained primarily by a simple Edman degradation method. The glycosylation profile was also determined by lectin blotting and MALDI-TOF mass spectrometry analysis. In addition, we tested whether yolkin has activities other than immunoregulatory and neuroprotective activities, such as antihypertensive, antidiabetic, or antioxidant.
2. Materials and methods
2.1. Reagents and chemicals
Reagents for SDS-PAGE were obtained from Bio-Rad (California, USA). Sephacryl S-100 HR resin was obtained from GE Healthcare (Chicago, Illinois, USA). 1,1,1,3,3,3-Hexafluoro-2-propanol (HFIP), thioflavin T (ThT), tris(hydroxymethyl)aminomethane (Tris), glycine, N,N,N′-N′-tetramethylethylenediamine (TEMED), sodium dodecyl sulfate (SDS), NH4HCO3, dithiothreitol, iodoacetamide, bovine serum albumin (BSA), bacterial lipopolysaccharide (LPS) from Escherichia coli (serotype O55:B5), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), Tween-20, angiotensin converting enzyme (ACE, EC 3.4.15.1; from rabbit lung), hippuryl-His-Leu, α-glucosidase from Saccharomyces cerevisiae, p-nitrophenyl glucopyranoside (pNPG), dipeptidyl peptidase-4 (DPP-4) from porcine kidney, Gly-Pro-p-nitroanilide, and 2,2′-azinobis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) were purchased from Sigma (New York, NY, USA). A Precision Plus protein standard (10–250 kDa) was obtained from Bio-Rad (Hercules, CA, USA). PageRuler™ Plus Prestained Protein Ladder (10–250 kDa) was obtained from Thermo Scientific (Waltham, MA, USA). Amyloid β42 (Aβ42) was obtained from Tocris (Bristol, UK). High-glucose Dulbecco's modified Eagle's medium (DMEM), a trypsin + EDTA mixture, and phosphate-buffered saline (PBS) (pH 7.4) were prepared at the Laboratory of General Chemistry of the Institute of Immunology and Experimental Therapy, PAS (Wroclaw, Poland). L-Glutamine and antibiotics (penicillin/streptomycin mixture) were purchased from BioWest (Nuaillé, France). Fetal bovine serum (FBS) was purchased from EURx Ltd (Gdansk, Poland). N-(1-Naphthyl)-ethylenediamine was purchased from Serva Feinbiochemica (Heidelberg, Germany). Sulfanilamide, sodium nitrite, orthophosphoric acid, and K2HPO4 were purchased from Avantor (Gliwice, Poland). Trifluoroacetic acid (TFA) was obtained from Fluka (Buchs, Switzerland). Acetonitrile was obtained from Merck (Darmstadt, Germany), and the trypsin solution was obtained from Promega (Madison, USA).2.2. Isolation of the yolkin polypeptide complex
Five successive isolations of yolkin were performed to show the reproducibility of the isolation method and the similarities of the resulting protein preparations. For each isolation, 10 eggs from caged hens were used. Yolkin was isolated according to the procedure described in detail by Polanowski et al.13 that consists of three main steps: fractionation of yolk proteins into granules and plasma, salting of the IgY-containing yolkin complex from the plasma with solid ammonium sulphate, and finally, separation of the yolkin from the IgY sample on a Sephacryl S-100 HR column (K50/100 Pharmacia Ltd, Kent, UK). Fractions containing yolkin were pooled, dialysed against water, and lyophilised. Particular isolations were named yolkins 3, 12, 13, 14 and 15. The concentration of protein in each of the isolated yolkin preparations was determined by the BCA method.29 The purity and peptide composition of the subsequent yolkin preparation were analysed using the SDS-PAGE technique, whereas the thermal stability was determined by nanoDSF.2.3. SDS-PAGE analysis
The purity of yolkin (numbered 3 and 12 to 15) was determined by SDS-PAGE. SDS/polyacrylamide slab gels were prepared using TXG Fast Cast acrylamide solutions (Bio-Rad, California, USA). Protein samples (20 μg perline) were diluted with a buffer containing dithiothreitol as a reducing reagent and loaded onto the gel slabs. At the end of the analysis, the gel slabs were stained with Coomassie Brilliant Blue R-250.2.4. Evaluation of the storage stability of yolkin
To test the storage stability, yolkin 3, both lyophilised and dissolved in sterile PBS (1 mg mL−1, filtered through a 0.22 μm filter into a sterile Eppendorf tube), was stored for four weeks at a refrigeration temperature of 4 °C, a freezing temperature of −20 °C, a room temperature of 25 °C, and a temperature of 37 °C. Electrophoretic separation under denaturing conditions was performed as described in section 2.3. The peptide profile obtained for each preparation was subjected to qualitative analysis. In addition, the biological activity was determined as described in section 2.5.2.5. Effect of storage conditions on the biological activity of yolkin
2.6. Evaluation of aggregation ability using ThT
Thioflavin T (ThT) was dissolved in sterile phosphate buffer (100 mM, pH 7.4) to a final concentration of 10 μM and vortexed for 10 min at RT. Aβ42 (20 μM) was used as a reference sample and was prepared as described previously.31 The fibrillation of yolkin 3 (100 μg in sterile PBS, pH 7.4) was measured in 96-well black plates. Before starting the measurement, ThT was added to the reaction mixture. The final volume of the samples was 100 μL. Fluorescence kinetics data were collected in triplicate at 37 °C using a CLARIOstar® Microplate Reader (BMG LabTech, Offenburg, Germany), and measurements were taken at 15-minute intervals for 48 hours. The excitation and emission wavelengths were set to 440 and 480 nm, respectively. The experiment was repeated three times with two repetitions each (n = 6). The plot shows a single measurement point based on the average value of the three independent repetitions, with error bars representing their standard deviation.2.7. Nanodifferential scanning fluorimetry (nanoDSF)
Thermal stability analysis of yolkins 3 and 12–15 was performed using the Prometheus NT.48 platform (NanoTemper Technologies GmbH). The standard capillaries were filled with 10 μL of sample solution containing 1 mg mL−1 protein in sterile PBS (pH 7.4). The initial fluorescence scans at 330 nm and 350 nm at 20 °C were used to determine the optimal LED excitation power of 36%. The fluorescent emission at 330 nm and 350 nm indicated that all the analyzed samples were within the optimal detection range of less than 20![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 counts. The samples were subjected to a temperature ramp of 1 °C min−1 from 20 °C to 95 °C during the melting scan, with continuous analysis and collection of the fluorescence signal at 330 nm and 350 nm. Data analysis was performed using dedicated instrument software, PR Therm Control, and PR Stability Analysis and presented as a ratio of 350/330 nm (NanoTemper Technologies GmbH).
000 counts. The samples were subjected to a temperature ramp of 1 °C min−1 from 20 °C to 95 °C during the melting scan, with continuous analysis and collection of the fluorescence signal at 330 nm and 350 nm. Data analysis was performed using dedicated instrument software, PR Therm Control, and PR Stability Analysis and presented as a ratio of 350/330 nm (NanoTemper Technologies GmbH).
2.8. LC-MS analysis
:1, v/v, Sigma, Merck). The reduction of disulfide bridges formed between thiol groups in cysteine residues was achieved by incubation for 30 minutes at 56 °C in a 10 mM solution of dithiothreitol (Sigma) in 100 mM NH4HCO3. Alkylation of the cysteine sulfhydryl groups was carried out by adding a 55 mM solution of iodoacetamide (Sigma) in 100 mM NH4HCO3 and incubating for 20 minutes in the dark at room temperature. In-gel digestion was carried out using a trypsin solution (Promega) at a concentration of 13 ng μL−1 in 10 mM NH4HCO3 containing 10% (v/v) acetonitrile at 37 °C overnight. After overnight incubation, the sample was centrifuged, and the solution was collected. Peptides were further extracted from the gel using 70% acetonitrile. All solutions were prepared using LC-MS grade reagents and Milli-Q water.
 | ||
| Fig. 1 SDS-PAGE analysis of yolkin preparations (a and b) and SDS-PAGE separation of yolkin stored at different temperatures (c). Yolkins 12–15 (20 μg per line) (a), yolkin 3 (20 μg per line) (b), and immunoglobulin Y (a, b and c) were subjected to electrophoresis followed by staining with Coomassie Brilliant Blue R-250. A Bio-Rad molecular weight marker was used. IgY from each preparation was loaded at 20 μg per line. (c) Yolkin 3 was either freeze-dried (1 mg mL−1) or dissolved in PBS (1 mg mL−1) and stored at 37 °C, 25 °C, 4 °C, or −20 °C for four weeks. Next, 20 μg of yolkin per line was electrophoresed on a 12% polyacrylamide gel under denaturing conditions. The proteins in the gel were stained with Coomassie Brilliant Blue R-250, and the gel image was captured using the Molecular Imager ChemiDoc XRS+ Imaging System (Bio-Rad). | ||
:water (90:10), 0.1% formic acid (Sigma); gradient: 5–55% eluent B; time: 150 minutes; eluent flow rate: 300 ml min−1; and injection volume: 15 μL. LC-MS additives, solvents, and Milli-Q water were used to prepare the eluents. Mass spectra were collected in positive ion mode with a range of m/z 110–2000. The data were analyzed and identified using ProteinScape 3.0 software (Bruker) and the MASCOT search engine (Matrix Science, 2.5.0) in the SwissProt database. The search criteria included taxonomy: vertebrates, mass tolerance: 10 ppm, fixed modifications: carbamidomethylation (C), and variable modifications: oxidation (M) and acetylation (protein N-terminus).
2.9. Analysis of yolkin glycosylation profiles
The released N-glycans were purified using Sep-Pak C18 cartridges (Waters Corporation, Milford, MA, USA) as described by Morelle and Michalski,33 with slight modifications. Briefly, the Sep-Pak C18 column was conditioned with 5 mL of methanol followed by 10 mL of 0.5% acetic acid. After PNGase A treatment, the sample was loaded. The elution of N-glycans was performed using 5 mL of 0.5% acetic acid. The lyophilized samples were then subjected to MALDI-TOF mass spectrometry analysis.
Samples (1 mg mL−1 in mQ) were analyzed using a MALDI-TOF Ultraflextreme III instrument (Bruker, Germany). The MALDI-TOF MS spectra were obtained in negative ion mode. AA [10 mg mL−1 in 1:1 AcN/2.5% FA (v/v)] was used as a matrix for analyses.
:5000 in TBS/1% BSA/0.05% Tween 20, for 1 hour. Finally, the glycoprotein bands were visualized using the BCIP/NBT reaction.
2.10. Determination of yolkin biological activity
| Inhibition activity (%) = [(A − B)/A] × 100% |
A − reaction blank, in which the mixture contained the same volume of the buffer solution instead of the ACE inhibitor sample, and B – reaction in the presence of both ACE and its inhibitor. The IC50 value was estimated from a dose–response curve of an inhibitor versus the percentage of ACE activity.
2.10.2.1. The α-glucosidase inhibitory activity was measured according to the method described by Yu et al.36. Briefly, 5 μL of the α-glucosidase solution (10 U mL−1 resolved in 0.1 M potassium phosphate buffer, pH 6.8) was premixed with 10 μL of the sample solution at different concentrations (in 10% DMSO) in 620 μL of 0.1 M potassium phosphate buffer (pH 6.8). After 20 min of incubation at 37.5 °C, 10 μL of pNPG (10 mM) used as a substrate was added to the mixture to start the enzymatic reaction, followed by incubation at 37 °C for 30 min. Finally, 650 μL of 1 M Na2CO3 solution was added to stop the reaction. The concentration of p-nitrophenol released was measured at λ = 410 nm, and the IC50 value was defined as described in point 2.1.10.
2.10.2.2. The DPP-4 inhibitory activity (IC50) was determined using the methods of Lacroix and Li-Chan37. The freeze-dried yolkin 3 was dissolved in 0.1 M Tris–HCl buffer, pH 8.0. The test sample (25 μL) was preincubated with an equal volume of DPP-4 (0.01 U ml−1, in 0.1 M Tris–HCl buffer, pH 8.0) at 37 °C for 10 min. Then, 50 μL of the substrate Gly-Pro-p-nitroanilide (1.6 mM) was added, and the mixture was incubated at 37 °C for 60 min. Then, 100 μL of 1 M sodium acetate buffer, pH 4.0, was added to stop the reaction. The amount of p-nitroanilide released was measured at λ = 405 nm. The IC50 value was defined as described in point 2.1.10.
The antioxidant activity was determined using the method of Re et al.38 The ABTS radical cation (ABTS˙+) was dissolved in water to an absorbance of 0.70 (60.02) at 734 nm. The tested samples (10 μL) were mixed with 990 μl of ABTS˙+ water solution. The mixtures were left at room temperature for 6 min, and then the absorbance of the resulting solutions was measured at 734 nm. For calibration, aqueous solutions of known Trolox concentrations were used. The radical scavenging activity of yolkin was expressed as μM Troloxeq per mg protein.
2.11. Data analysis and graphical visualization
Statistics and graphs were prepared using GraphPad Prism Software v 9.5.1. The data are presented as the means ± SDs. The data were analyzed using one-way ANOVA. A value of p ≤ 0.05 was considered to indicate statistical significance.3. Results
3.1. Isolation of the yolkin complex
The yolkin obtained from the IgY sample is a mixture of polypeptides and proteins with molecular weights ranging from 15 to 37 kDa, as shown by the electrophoretic analysis of five independent isolations named 3 (yolkin 3) and 12–15 (yolkins 12–15) (Fig. 1a and b). The predominant proteins had molecular weights of 35 and 37 kDa. Despite additional re-chromatography of the preparations, slight IgY contamination was observed in yolkins 12–15. An average concentration of 900 μg ml−1 protein (determined by the BCA method) was obtained from two milligrams of lyophilised yolkin preparation.3.2. Evaluation of storage stability
The stability of yolkin 3 stored in two forms, lyophilization (referred to as y100/L) and PBS solution (referred to as y100/P), was investigated. The yolkin samples were stored at different temperatures (high, 37 °C; room, 25 °C; refrigeration, 4 °C; and freezing, −20 °C) for 4 weeks. Two parameters were analyzed: the intensity of the yolkin-related bands and the biological activity determined by the effect of yolkin on NO production in BMDM cells.SDS-PAGE analysis revealed changes in the intensity of the yolkin-related band. The intensities of the effects of lyophilized yolkin stored at −20 °C, 4 °C, and 25 °C, as well as PBS-solubilized yolkin stored at −20 °C and 4 °C, were comparable. However, yolkin stored at 37 °C (both lyophilized and PBS-solubilized) showed a significant reduction in band intensity (Fig. 1c).
Analysis of the effect of storage temperature on the biological activity of yolkin showed that lyophilised yolkin stored at −20 °C, 25 °C, and 4 °C induced the production of comparable amounts of NO, which were 38.6 μM, 37.9 μM, and 37.7 μM, respectively. However, yolkin stored at 37 °C was much less active (Fig. 2e). Compared with lyophilization, the storage of yolkin in PBS solution resulted in a 50% decrease in activity. The concentrations at −20 °C and 25 °C were 21.6 μM NO and 21.5 μM NO, respectively (Fig. 2a and c). However, at 4 °C, there was no significant difference, with 38.3 μM NO for y100/P and 37.7 μM NO for y100/L (Fig. 2b). Nevertheless, 37 °C was found to be the least favourable temperature for storing yolkin, particularly as a PBS solution (Fig. 2d), resulting in a reduction of approximately 40% in NO production (23.2 μM NO for y100/L and 8.2 μM NO for y100/P) (Fig. 2e and f) compared to the other temperatures tested.
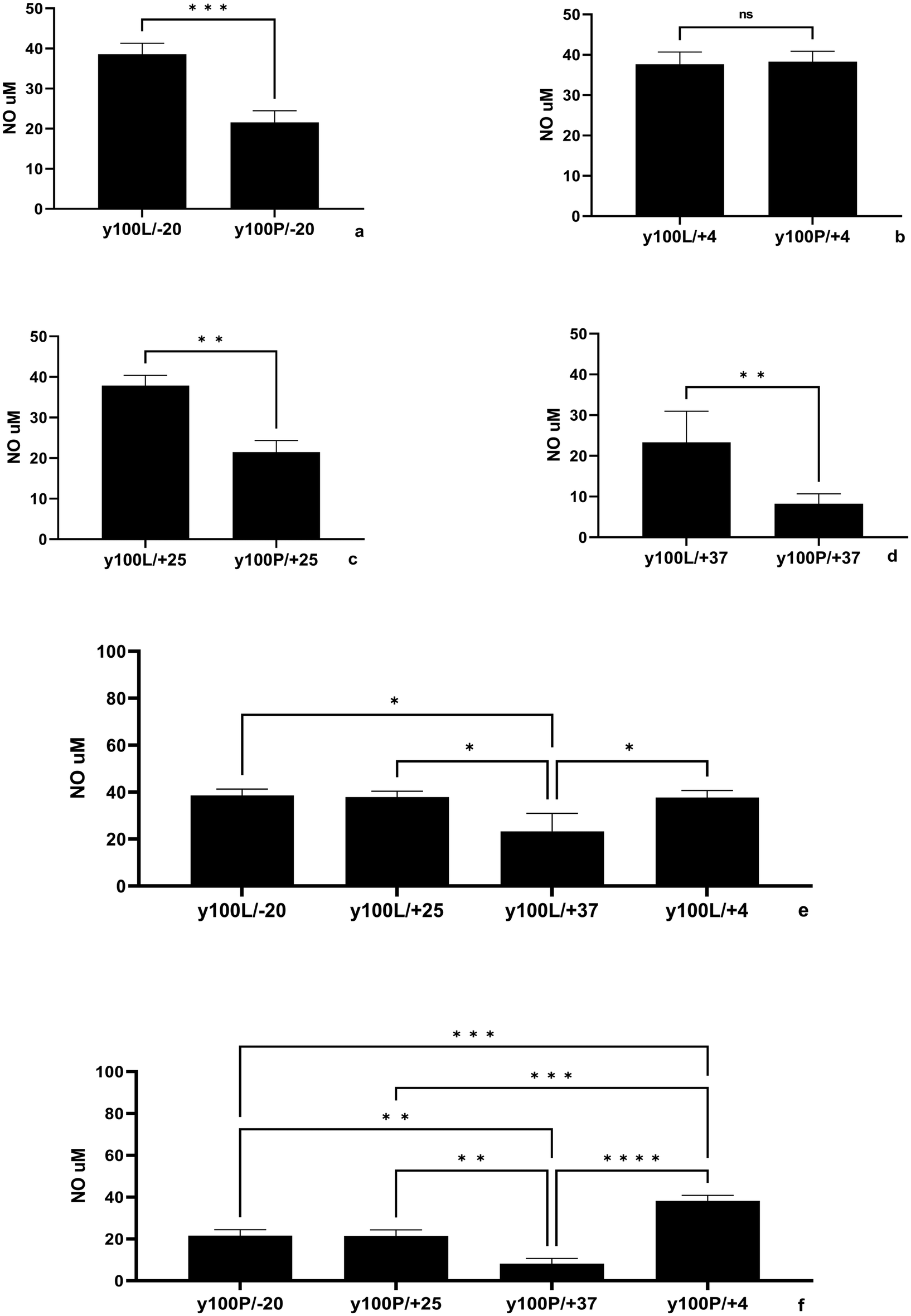 | ||
| Fig. 2 Impact of yolkin storage conditions on nitric oxide (NO) production in BMDMs. Yolkin 3 was either freeze-dried (y100/L, 1 mg mL−1) or dissolved in PBS (y100/P, 1 mg mL−1) and stored at 37 °C (d, e, f), 25 °C (c, e, f) 4 °C (b, e, f) or −20 °C (a, e, f) for 4 weeks. After the incubation period, we analyzed the effect of the yolkin preparation on nitric oxide production in BMDM cells. BMDMs (1 × 106 mL−1) were cultured with a specific yolkin sample (100 μ mL−1) under 5% CO2/95% air for 24 hours. Then, the supernatants were collected, and the level of NO was determined by the Griess reaction. The results represent three independent experiments and are presented as the mean ± SD. To examine the differences between the samples studied, we used one-way ANOVA. The significance levels are denoted as follows: *p ≤ 0.05, **p ≤ 0.001, ***p ≤ 0.0001 and ****p < 0.0001. | ||
3.3. Evaluation of the yolkin aggregation ability using ThT
Using a small molecule dye, thioflavin T (ThT), is one of the simplest and most widely used methods for monitoring the aggregation of amyloidogenic proteins. ThT exhibits a fluorescence emission maximum at a wavelength of approximately 480 nm when it binds to the beta-sheet structure of prefibrillar and fibrillar protein aggregates. Many proteins can form dimers, oligomers, and fibrils containing β-sheet and amyloid structures.39 The fluorescence measured in the presence of the yolkin complex after 36 hours of incubation at 37 °C showed no change in intensity compared to that of the amyloid β42 (10 μM) fluorescence, which was used here as a reference (Fig. 3). This indicates that yolkin does not tend to aggregate.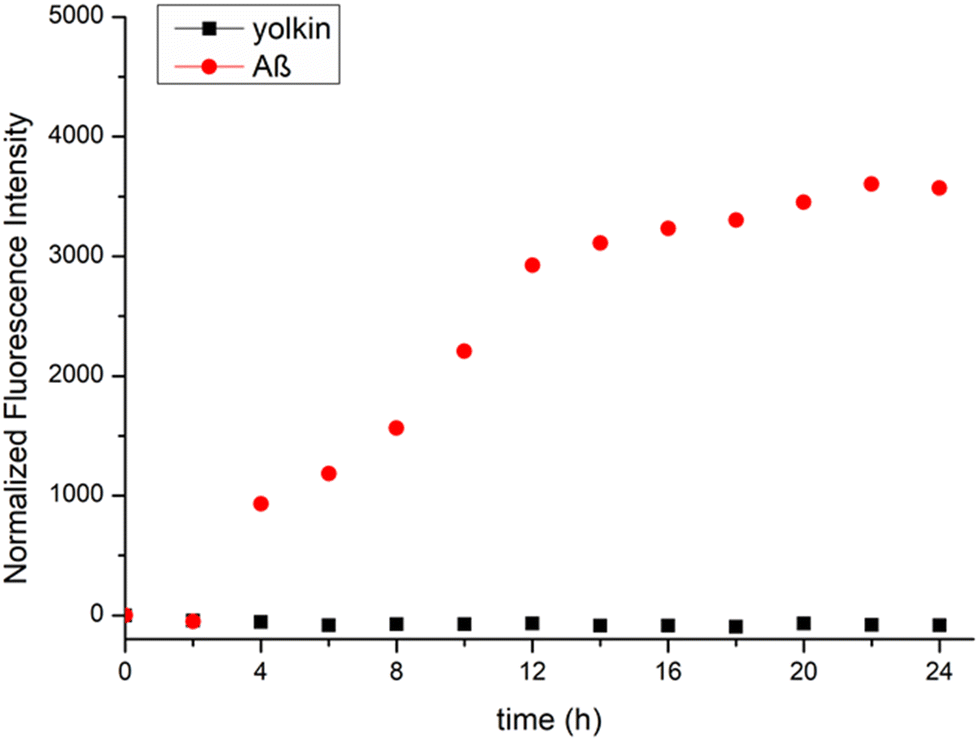 | ||
| Fig. 3 Determination of yolkin aggregation ability. The formation of yolkin 3 (100 μg mL−1) and Aβ42 (10 μM) fibrils was monitored using the ThT fluorescence assay. The fluorescence intensity was measured at an excitation/emission wavelength of 420/489 nm and is presented as a normalized fluorescence curve. The experiment was repeated twice in three independent replicates (n = 6). Pure ThT solution was used as a blank to overcome the problem of autofluorescence. | ||
3.4. Nanodifferential scanning fluorimetry (nanoDSF)
NanoDSF, performed using the Prometheus NT.48 platform (NanoTemper Technologies GmbH), was used to characterize the process of yolkin isolation. The studies focused on analysing the thermal and colloidal stability, folding quality, and conformational similarity of yolkin after the isolation process. Yolkins 12, 13, 14, and 15, which were derived from the same upstream source material, had similar unfolding profiles (Fig. 4a and b) and exhibited relatively high initial fluorescence ratios in the range of 0.82–0.86 (Fig. 4a). They exhibit two unfolding events, with the first melting temperature point (Tm) in the range of 46–48 °C and the second unfolding event with the Tm in the range of 72–73 °C (Table 4A, ESI†). The differences in the colloidal stability of the yolkin samples also indicate their tendency to aggregate (Fig. 4c). Yolkins 12 and 13 show relatively little aggregation in the presence of thermal unfolding, but yolkins 14 and 15 show a significant propensity to aggregate in association with the second unfolding event—the onset of aggregation in the range of 69–72 °C and its inflection point in the range of 75 °C. In contrast, yolkin 3, derived from different upstream sources, exhibited different melting curve characteristics (Fig. 4). Its initial fluorescence ratio is significantly lower in the range of 0.7 (Fig. 4a), and it has only one unfolding event with the Tm in the range of 71 °C, corresponding to the second Tm value of yolkins 12–15. Moreover, the unfolding profiles of IgY are similar to those of the first Tm in the range of Tm1 values of yolkins 12–15 (Fig. 4a). | ||
| Fig. 4 Thermal unfolding profiles of yolkins 12–15 compared to the unfolding signals of IgY and yolkin 3. Samples were measured over a temperature range of 20 °C–95 °C. (a) The upper part shows the ratio of integrated fluorescence at 350/330 nm. The vertical lines indicate the Tm. (b) The middle part shows the corresponding first derivative plotted against the temperature. (c) The lower part shows the aggregation profile visualized as a function of the scattering signal versus temperature. | ||
3.5. Sequence analysis of yolkin (LC-MS)
To characterize the peptides contained in yolkin 3, MS was used. Mass spectrometric analysis revealed the presence of 19 peptides whose amino acid sequences were homologous to part of the amino acid sequence of vitellogenin II (see ESI Table 2A†), with a sequence coverage of 9.70%. This metric refers to the proportion of amino acid sequence coverage of the identified yolkin peptides ranging from 7–23 amino acid residues compared to the entire vitellogenin II sequence of 1850 amino acid residues. In addition, 12 peptides homologous to vitellogenin I were also identified, with 6.60% sequence coverage, in terms of the proportion of amino acid sequence coverage of the identified yolkin peptides (see ESI Table 3A†). The identified yolkin peptides range from 6–23 amino acid residues, whereas the polypeptide chain of vitellogenin I has a length of 1912 amino acid residues.3.6. Analysis of yolkin glycosylation profiles
Lectin blot analysis using Con A, SNA, RCA and AAL (specificities listed in Table 1A, ESI†) revealed that three yolkin fractions with MWs above 16 kDa were glycosylated (Fig. 5a). Most fractions were detected by Con A staining, followed by SNA. RCA bound only to the major fractions of yolkin, and AAL stained predominantly a band corresponding to the IgY heavy chain. PNGase A treatment reduced the staining intensity of all the lectins, with some glycopeptides/glycoproteins being completely deglycosylated. These results suggest that yolkin fractions carry N-glycan chains, both of which are high-mannose and complex types capped by sialic acid residues, with a low level of fucosylation. | ||
| Fig. 5 Glycosylation profile of yolkin. (a) Lectin blot analysis of yolkin 3 glycosylation using Con A, SNA, RCA, and AAL (specificities listed in Table 1, ESI†) before and after PNGase A treatment. (b) MALDI-TOF MS mapping of 2AA-N-glycans derived from yolkin. | ||
The MALDI-TOF MS spectrum of 2AA-N-glycans from yolkin 3 is shown in Fig. 5b. The profile indicates a peak at m/z 1375.99 [Man5GlcNAc2-2AA + H + Na]+, and up to five additional Man residues were added to this structure, which are represented by ions at m/z 1538.01, 1700.01, 1862.53, 2025.09 and 2186.04. The profile also indicated that the ions at m/z 1478.78 [GalMan3GlcNAc4 + H]+, m/z 1680.48 [GalMan3GlcNAc5 + H]+, m/z 1842.03 [Gal2Man3GlcNAc5 + H]+, and m/z 2132.60 [Neu5AcGal2Man3GlcNAc4 + H]+ corresponded to non-2AA tags. The ions at m/z 1255.04 [Man3GlcNAc3-2AA + H + Na]+, m/z 1417.04 [GalMan3GlcNAc3-2AA + H + Na]+, m/z 1620.05 [GalMan3GlcNAc4-2AA + H + Na]+, m/z 1782.09 [Gal2Man3GlcNAc4-2AA + H + Na]+, m/z 1911.00 [Neu5AcGalMan3GlcNAc4-2AA + H + Na]+, m/z 2072 [Neu5AcGal2Man3GlcNAc4-2AA + H + Na]+, and m/z 2363.97 [Neu5Ac2Gal2Man3GlcNAc4-2AA + H + Na]+ represented all N-glycan structures present in yolkin 3.
3.7. Yolkin biological activity
Not much is known about peptides naturally present in egg yolks, such as yolkin. Therefore, its antidiabetic, antioxidant and antihypertensive activities were measured (Table 1). Yolkin showed antioxidant activity. The free radical-ABTS scavenging activity of yolkin for 1 milligram was equivalent to that of 2.5 μM Trolox. In addition, yolkin showed significant inhibitory activity against the enzyme α-glucosidase, reaching an IC50 = 34.84 μg. Yolkin was not shown to be a significant inhibitor of ACE (IC50 = 357.83 μg) or DPP-IV (IC50 = 325.46 μg).| Biological activity | |||
|---|---|---|---|
| Antidiabetic | Antihypertensive | Antidiabetic | Antioxidant |
| DPP-IV IC50 (μg) | ACE IC50 (μg) | α-Glucosidase IC50 (μg) | ABTS scavenging (μMTroloxeq per mg) |
| 325.46 ± 3.96 | 357.83 ± 4.80 | 34.84 ± 4.18 | 2.50 ± 0.30 |
4. Discussion
Hippocrates said, “Let food be thy medicine and medicine be thy food”. This golden phrase has inspired people to search for highly nutritious substances in food. Biologically active nutraceuticals are characterized by multiple direct effects and high potential as safe, effective, and long-term supplements, as they are promising therapeutic agents that can maintain the body in good shape and prevent various diseases (cancer, diabetes, neurodegenerative diseases or inflammation); additionally, they can improve quality of life and slow the ageing process. The abundance of food sources provides great variety. Today, people's increasing fascination with nutraceuticals is driven by a growing awareness of their dual properties, which include both nutritional and pharmaceutical functions.40 Recent studies have shown that hen eggs can be valuable materials for biomedical applications due to the high levels of biologically active compounds they contain. Most of the substances present in the egg material possess biological activity related to their antimicrobial, anticancer, and immunogenic properties.27 Yolkin deserves special attention.In 2012, Polanowski et al.12 showed that IgY, the major chicken immunoglobulin fraction, is accompanied by extra proteins and functions as a transporter of biologically active substances, as previously shown for colostral IgG.41 To date, the ability of yolkin to regulate both immunomodulatory and neuroprotective mechanisms has been demonstrated, suggesting its potent therapeutic properties to aid in the treatment or prevention of both immunological and neurodegenerative disorders.3 To establish a novel therapeutic approach based on yolkin, further research is needed to fully characterize its physical and biological properties.
In the present work, we determined the storage conditions of yolkin, its thermal stability, its potential aggregation activity, and the influence of these parameters on its biological activity. In addition, the degree of glycosylation and the amino acid sequence were determined.
Yolkin complexed with IgY was first isolated from egg yolk on Sephacryl S-100 HR by size exclusion chromatography, as described in the procedure published by Polanowski et al.12 Electrophoretic analysis of yolkin from five independent isolations revealed a mixture of several proteins and peptides with MWs ranging from over 15 to approximately 37 kDa (Fig. 1a and b), with bands at 35, 32, and 24 kDa representing the major proteins. These results are in general agreement with those of Polanowski et al.13 and Zambrowicz et al.,16 who defined yolkin as a heterogeneous set of proteins and peptides ranging in size from 1 to approximately 35 kDa. In our opinion, yolkin preparations can vary, and the particular constituents can be highly dependent on the starting material (egg yolk).
The yolkin proteins and polypeptides separated by SDS-PAGE, previously identified by the Edman technique, showed homology to the C-terminal sequence of vitellogenin II.13 Edman degradation is a classic and accurate method for protein sequencing, especially for smaller proteins, but it is limited in speed and scalability.42 Depending on the amount of the sample, it can sequence 10–50 amino acid residues. Therefore, we decided to perform an additional analysis using mass spectrometry (MS), a modern and versatile technique for protein identification and sequencing, especially for complex mixtures and larger proteins.43 A comparison of the high homology of yolkin components with vitellogenin II revealed the presence of 19 peptides with a high sequence coverage of 9.70% in the Mascot database. Interestingly, 12 peptides homologous to vitellogenin I were also detected, with a sequence coverage of 6.60%. The results obtained confirm previous studies on the peptide composition of yolkin and its homology to vitellogenin II and provide additional data on the content of peptides homologous to vitellogenin I.44
Hen eggs are abundant in glycoproteins. In particular, N-glycosylated egg proteins play an important role in maintaining their structure and stability and are important for activating the immune system to neutralize pathogens during embryonic development.45–47 The precursor of the major egg yolk proteins, vitellogenin, is enzymatically cleaved during egg processing into lipovitellins, phosvitins, and peptides YGP40 and YGP42.5,8,48,49 In addition, the latest research has shown that vitellogenin II-derived peptides and proteins, such as the yolk glycopeptide YGP40 and yolkin, whose amino acid sequence corresponds to the N-terminal region of the C-terminal domain of vitellogenin II (cysteine-rich), have immunomodulatory properties and may participate in host immune defence.3,13,15,18,19,50 In 1985, Yamamura et al.8 showed that YGP40 contains both high-mannose-type and complex-type carbohydrate chains that bind to proteins through asparagine (N-linked) residues. Considering that YGP40 can be a potential precursor of some yolkin peptides, we speculated that YGP40 can also be glycosylated to a level comparable to that of YGP40. Our study confirmed our estimations. We observed that yolkin, comparable to fractions with MWs above 16 kDa, is glycosylated and carries N-glycan chains, both of which are high in mannose and complex types capped by sialic acid residues. There is now strong evidence that yolkin constituents may be a set of peptides resulting from vitellogenin-derived YGP40 proteolytic cleavage. Certain proteins require glycosylation for their biological activity.45–47 Although glycosylation can enhance biological maintenance, as observed with IgY, a prominent avian immunoglobulin class,51 there are some exceptions. Zambrowicz et al.17 demonstrated an inverse correlation between the yolkin glycosylation level and nitric oxide production in macrophages. Compared with the glycosylated yolkin fractions, the deglycosylated yolkin fractions showed greater activity with no significant effect on cytokine induction. In contrast, recombinant YGP40 lacks carbohydrate moieties due to the lack of glycosylation activity in E. coli. Interestingly, the ability of YGP40 to upregulate iNOS expression, induce NO production, and stimulate cytokine production is comparable to that of the glycosylated yolkin complex.50 This indicates that the presence of the carbohydrate is not a critical factor in determining yolkin biological activity. On the other hand, protein glycosylation has been shown to stabilize the structure and stability of proteins and protect them from proteolytic degradation, aggregation, or damage by active oxygen radicals.45,51,52 We speculate that the presence of high-mannose-N-glycans in yolkin peptides may prevent its aggregation and decrease its sensitivity to thermal denaturation and proteolytic hydrolysis. However, further studies are needed to explain this phenomenon.
In the field of nutraceutical science, careful study of the optimal storage conditions is essential. This study aimed to elucidate specific aspects of yolkin storage, focusing on parameters that have a significant influence on yolkin stability and bioavailability. Unfortunately, the current literature on the stability of bioactive preparations during storage is very limited. Maintaining the integrity and functionality of purified proteins during prolonged storage is paramount, with the shelf life ranging from days to over a year, depending on the protein type and storage conditions. The key characteristics of protein storage conditions also depend on factors, such as formulation and temperature. Specifically, storage at surrounding temperatures can lead to protein degradation and reduced activity. A better understanding of the technological conditions is essential to optimize the storage protocols for egg yolk-based therapeutic strategies.53,54 In particular, proteins stored in solution at 4 °C may experience reduced stability within a few days or weeks, mainly due to microbial or proteolytic degradation. Whereas freezing at −20 °C is a common method for cold protein storage, the drawback is the potential compromise of protein stability due to freeze–thaw cycles. On the other hand, lyophilization is an alternative that allows the protein to be stored for longer periods, minimizing the risk of degradation.2,55 Another important consideration is the effect of storage conditions on protein aggregation, which can have a significant impact on bioavailability.56 In this study, the effects of two different yolkin formulations, lyophilised and as PBS solution, were investigated, and different storage temperatures were tested: high (37 °C), room (25 °C), refrigeration (4 °C), and freezing (−20 °C) for 4 weeks. Two parameters were analyzed: the intensity of the yolkin-related bands and the biological activity determined by the effect of yolkin on NO production in BMDM cells. SDS-PAGE analysis revealed changes in the intensity of the yolkin-related bands, indicating that the most favorable form of storage for yolkin at 4 °C is lyophilization. However, analysis of the effect of storage temperature on the biological activity of lyophilized yolkin showed no differences between temperatures of −20 °C, 4 °C and 25 °C (Fig. 2e), although there were marked differences in band intensity in the electrophoretic image (Fig. 1c). A temperature of 37 °C was found to be the most unfavorable temperature for yolkin storage. Both band intensity (Fig. 1c) and NO production (Fig. 2d, e and f) were significantly reduced (approximately 40%) compared to those under the other conditions tested.
Recent data suggest that some proteins can self-aggregate and form amyloid fibrils under certain storage conditions.57,58 These fibrils are not connected with any known amyloid disease; however, they have comparable morphological features to the amyloid fibrils of disease-associated proteins and can significantly alter their bioavailability and biological potential. For instance, the bacteriostatic and bactericidal properties of hen's egg white lysozyme are mainly attributed to its monomers and dimers. In contrast, its fibrils demonstrate notably enhanced antibacterial activity against both lysozyme-resistant S. aureus and lysozyme-insensitive E. coli,59,60 compared to its monomers. On the other hand, insulin has been shown to form polymorphic amyloid fibrils at the site of drug injections in diabetic patients, leading to harmful insulin injection amyloidosis.61
Based on the above data, we investigated whether yolkin tended to form amyloid fibrils. The recommended fluorescence method using thioflavin T (ThT) was used for this purpose.62 The assay was performed at 37 °C, the temperature at which the greatest reduction in yolkin biological activity was observed (Fig. 2e and f). ThT fluorescence in the presence of yolkin after 36 hours of incubation did not change compared to that in the presence of amyloid β42 (10 μM), which is used as a reference protein with a high aggregation capacity (Fig. 3). This indicates that yolkin is a stable complex and does not tend to self-aggregate, which may be one of the reasons for the reduced bioavailability and biological activity of yolkin.
In the next step of our study, we used the nanoDSF method to analyze the thermal and colloidal stability, folding quality, and conformational similarity of yolkin after isolation.63,64 The Prometheus NT.48 platform, employing dual-UV technology, facilitated rapid fluorescence detection, ensuring high-resolution unfolding curves and label-free evaluation of tryptophan and tyrosine residue properties. This method is ideal for efficient protein analysis, offering quick, accurate results in protein research, engineering, formulation development, and quality control, spanning R&D to downstream validation in purification and storage processes.65,66 We observed that yolkins 3 and 12–15 have similar unfolding profiles (Fig. 4a and b), with two unfolding events with the first Tm in the range of 46–48 °C and the second unfolding event with the Tm in the range of 72–73 °C. Interestingly, when we analyzed the colloidal stability of the samples, represented by their tendency to aggregate, we observed different colloidal properties of the individual samples (Fig. 4c). Yolkins 12 and 13 show relatively little aggregation in the presence of thermal unfolding. However, yolkins 14 and 15 showed a significant tendency to aggregate in association with the second unfolding event – the onset of aggregation in the high-temperature range of 69–72 °C and its inflection point in the range of 75 °C. The parameters of the aggregation event suggest that the thermal unfolding of the sample triggered this event. Yolkins 12–15 were derived from the same upstream source material. Interestingly, when we compared the thermal and colloidal stability profiles of these samples with those of yolkin 3, which were derived from different upstream sources, we observed differences in the melting curves (Fig. 4). Yolkins 12–15 all have relatively high initial fluorescence values with ratios in the range of 0.82–0.86, whereas yolkin 3 has a significantly lower initial fluorescence ratio in the range of 0.7. In addition, yolkin 3 has only one unfolding event with the Tm in the range of 71 °C, corresponding to the second Tm value of yolkins 12–15. Therefore, the dual unfolding properties of yolkins 12–15 may indicate the presence of a more complex protein mixture. It is possible that yolkins 12–15 have two protein groups, whereas yolkin 3 is more homogeneous. The protein groups were shared by yolkins 12–15 due to the high similarity of the unfolding profiles of these samples. A higher initial fluorescence ratio would suggest that the group with a lower Tm value could have more aromatic residues exposed to the environment in their native state or have more Tyr in their sequence. Furthermore, when we analyzed the unfolding profiles of IgY, which could be a common contaminant obtained during the extraction process, we observed a very similar unfolding profile with the first Tm in the range of the Tm1 value of yolkins 12–15 (Fig. 4). SDS-PAGE analysis also confirmed that yolkins 12–15 had bands corresponding to those of the IgY samples. This potentially suggests that the additional Tm value of yolkins 12–15 could be of IgY origin and is missing in yolkin 3 both when analyzed on gel and on the nanoDSF melting profile.
There is a great deal of information available on biologically active peptides produced in vitro from egg yolk proteins by enzymatic hydrolysis.67–69 However, the polypeptides and peptides naturally occurring in egg yolk have not been adequately studied. The granular fraction of egg yolk contains α-lipovitellins, β-lipovitellins, high-density lipoproteins (HDLs), phosvitins, and low-density lipoproteins (LDLs). Enzymatic hydrolysis of this fraction with pepsin after defatting yields peptides possessing antioxidant, antidiabetic, and antihypertensive activity.67 Synthetic analogues of these isolated and identified peptides showed biological activity. The peptide YIEAVNKVSPRAGQF (apovitellin-1 fragment) shows strong ACE inhibitory activity, with IC50 = 9.4 μg ml−1. The YINQMPQKSRE peptide (apolipoprotein B fragment) showed significant DPPH free radical scavenging and DPP-IV inhibitory activity, and the ACE inhibitory activity (IC50) reached 10.1 μg ml−1. The VTGRFAGHPAAQ peptide, a fragment of vitellogenin-2, shows α-glucosidase inhibitory activity (IC50 = 365.4 μg ml−1). Lipoproteins from the granular fraction of egg yolk are precursors of a potentially antihypertensive peptide with the sequence K.VQWGIIPSWIK. K.68 Phosvitin, on the other hand, is a precursor of phosphopeptides with multifunctional activity; it has good iron chelating capacity and free radical scavenging activity, as well as inhibitory activity against proinflammatory cytokines.69 The common feature of yolk granule proteins and yolkin is that they share the same precursor, vitellogenin.9 Similarly, yolkin, which is a product of vitellogenin proteolysis, may have similar activity. We have shown that yolkin has both antioxidant and antidiabetic activity (inhibition of α-glucosidase activity) (Table 1).
5. Conclusions
This research demonstrated the potential of the yolkin complex derived from hen egg yolks as natural nutraceuticals with significant health benefits. This study investigated the peptide composition, glycosylation level, storage conditions, and thermal stability of yolkins. This finding emphasizes the importance of proper storage conditions and thermal stability and showcases the credibility and expertise of yolkin as a natural nutraceutical with various health benefits. Yolkin possesses both antioxidant and antidiabetic properties, making it a highly promising natural compound for addressing various health issues in the nutraceutical landscape. The yolkin complex has demonstrated its potential as a valuable addition to the field due to its ability to reduce oxidative stress and metabolic dysregulation. However, further research is needed to determine the specific dosage forms, optimal delivery mechanisms, and potential synergistic effects of these compounds with other bioactive compounds. To fully understand the benefits of the individual components of the yolkin complex, it is critical to thoroughly explore its biological potential. Unlocking the full therapeutic potential of yolkin and facilitating its integration into personalized and preventive healthcare strategies is crucial. This research has the potential to revolutionize the field of healthcare. Therefore, it is imperative to continue investing in this area to fully understand the benefits of yolkin and its potential applications.Author contributions
Aleksandra Zambrowicz: investigation, methodology, data curation, formal analysis, funding acquisition, writing – original draft. Katarzyna Kapczyńska, Paweł Kania, Katarzyna Szymczak-Kulus, and Marta Kaszowska: investigation, methodology, data curation, formal analysis, writing – original draft. Jakub Nowak: investigation, methodology, data curation, formal analysis, funding acquisition, writing – original draft. Sabina Górska, Dominika Jakubczyk, and Marta Piksa: investigation, methodology, writing – original draft. Józefa Macała and Wioletta Kazana-Płuszka: investigation. Agnieszka Zabłocka: conceptualization, supervision, investigation, methodology, data curation, formal analysis, funding acquisition, writing – original draft, review & editing. All the authors have read and approved the final manuscript.Data availability
Data for this article, including original SDS PAGE gels, lectin gels, WB and LC-MS, and nanoDSF, are available at the University of Gdansk repository at https://doi.org/10.18150/QCYXVY.In addition, the data supporting this article have been included as part of the ESI.†
Conflicts of interest
The authors declare no competing interests.JSN acts as an external application scientist consultant at NanoTemper Technologies. The research results and opinions presented in this article were not influenced by the above affiliations in terms of their design, execution, or interpretation. The remaining authors have no competing interests to declare that they are relevant to the content of this article.
Acknowledgements
This work was partially supported by the Wroclaw University of Environmental and Life Sciences (Poland) as part of the Ph.D. research program “MISTRZ”, No. N090/0016/22, and partially by the statutory activity of IITD PAS. J. S. N. is supported by the European Research Council (ERC) under the European Union's Horizon 2020 Research and Innovation Program grant no. 101001394. We would like to thank NanoTemper for providing Prometheus NT.48 for our measurements.References
- Y. Mine, Egg proteins and peptides in human health – chemistry, bioactivity and production, Curr. Pharm. Des., 2007, 13, 875–884 CrossRef CAS PubMed.
- S. Rehault-Godbert, F. Baron, S. Mignon-Grasteau, V. Labas, M. Gautier, M. T. Hincke and Y. Nys, Effect of temperature and time of storage on protein stability and anti-salmonella activity of egg white, J. Food Prot., 2010, 73, 1604–1612 CrossRef CAS PubMed.
- A. Zambrowicz, A. Zabłocka, D. Bednarz and Ł. Bobak, Importance for humans of recently discovered protein compounds – yolkin and yolk glycopeptide 40, present in the plasma of hen egg yolk, Poult. Sci., 2023, 102, 102770 CrossRef CAS PubMed.
- J. Li, J. Zhai, L. Gu, Y. Su, L. Gong, Y. Yang and C. Chang, Hen egg yolk in food industry – A review of emerging functional modifications and applications, Trends Food Sci. Technol., 2021, 115, 12–21 CrossRef CAS.
- J. Sui, J. Xiao, X. Chang, H. Ye, Y. Xu, J. Wang and F. Geng, Research Note: Integrated proteomic analyses of chicken egg yolk granule, Poult. Sci., 2023, 102, 102711 CrossRef CAS PubMed.
- A. Laca, B. Paredes, M. Rendueles and M. Díaz, Egg yolk granules: Separation, characteristics and applications in food industry, LWT – Food Sci. Technol., 2014, 59, 1–5 CrossRef CAS.
- P. Jolivet, C. Boulard, T. Chardot and M. Anton, New insights into the structure of apolipoprotein B from low-density lipoproteins and identification of a novel YGP-like protein in hen egg yolk, J. Agric. Food Chem., 2008, 56, 5871–5879 CrossRef CAS PubMed.
- J. Yamamura, T. Adachi, N. Aoki, H. Nakajima, R. Nakamura and T. Matsuda, Precursor-product relationship between chicken vitellogenin and the yolk proteins: the 40 kDa yolk plasma glycoprotein is derived from the C-terminal cysteine-rich domain of vitellogenin II, Biochim. Biophys. Acta, Gen. Subj., 1995, 1244, 384–394 CrossRef CAS PubMed.
- R. N. Finn, Vertebrate Yolk Complexes and the Functional Implications of Phosvitins and Other Subdomains in Vitellogenins1, Biol. Reprod., 2007, 76, 926–935 CrossRef CAS PubMed.
- J. S. Chen, T. W. Sappington and A. S. Raikhel, Extensive sequence conservation among insect, nematode, and vertebrate vitellogenins reveals ancient common ancestry, J. Mol. Evol., 1997, 44, 440–451 CrossRef CAS PubMed.
- W. R. Lindsay, C. R. Friesen, C. Sihlbom, J. Bergström, E. Berger, M. R. Wilson and M. Olsson, Vitellogenin offsets oxidative costs of reproduction in female painted dragon lizards, J. Exp. Biol., 2020, 223, jeb221630 CrossRef PubMed.
- A. Polanowski, A. Zabłocka, A. Sosnowska, M. Janusz and T. Trziszka, Immunomodulatory activity accompanying chicken egg yolk immunoglobulin Y, Poult. Sci., 2012, 91, 3091–3096 CrossRef CAS PubMed.
- A. Polanowski, A. Sosnowska, A. Zabłocka, M. Janusz and T. Trziszka, Immunologically active peptides that accompany hen egg yolk immunoglobulin Y: separation and identification, Biol. Chem., 2013, 394, 879–887 CrossRef CAS PubMed.
- A. Zabłocka, Ł. Bobak, J. Macała, J. Rymaszewska, W. Kazana and A. Zambrowicz, Comparative Studies of Yolkin Preparations Isolated from Egg Yolks of Selected Bird Species, Chem. Biodivers., 2021, 18, e2100178 CrossRef PubMed.
- A. Zabłocka, A. Sosnowska, A. Urbaniak, M. Janusz and A. Polanowski, Peptides accompanying chicken egg yolk IgY – alternative methods of isolation and immunoregulatory activity, Food Funct., 2014, 5, 724–733 RSC.
- A. Zambrowicz, A. Zabłocka, Ł. Bobak, J. Macała, M. Janusz, A. Polanowski and T. Trziszka, A simple and rapid method of isolation of active polypeptide complex, yolkin, from chicken egg yolk, Food Chem., 2017, 230, 705–711 CrossRef CAS PubMed.
- A. Zambrowicz, A. Zabłocka, M. Sudoł, Ł. Bobak, P. Sosicka and T. Trziszka, The effect of carbohydrate moieties on immunoregulatory activity of yolkin polypeptides naturally occurring in egg yolk, LWT, 2018, 88, 165–173 CrossRef CAS.
- W. Kazana, M. Mitkiewicz, M. Ochnik, M. Sochocka, A. Zambrowicz, G. Piechowiak, J. Macała, P. Miernikiewicz and A. Zabłocka, Yolkin Isolated from Hen Egg Yolk as a Natural Immunoregulator, Activating Innate Immune Response in BMDM Macrophages, Oxid. Med. Cell. Longevity, 2020, 2020, 5731021 CAS.
- W. Kazana, D. Jakubczyk, K. Pacyga-Prus, K. Leszczyńska, S. Górska, J. Siednienko, J. Macała, G. Piechowiak and A. Zabłocka, A Novel Mechanism of Macrophage Activation by the Natural Yolkin Polypeptide Complex from Egg Yolk, Int. J. Mol. Sci., 2022, 23, 3125 CrossRef CAS PubMed.
- A. Zablocka, A. Zambrowicz, J. Macala, W. Kazanal and A. Polanowski, Yolkin- A Polypeptide Complex Isolated From Chicken Egg Yolk with Potential Neuroprotective and Antioxidative Activity, Neuropsychiatry (London), 2018, 8, 833–842 Search PubMed.
- B. Obmińska-Mrukowicz, M. Szczypka, M. Lis, A. Pawlak, A. Suszko-Pawłowska, A. Sysak, A. Zambrowicz, T. Burster, M. Kocięba, J. Artym, E. Zaczyńska, I. Kochanowska and M. Zimecki, Effects of yolkin on the immune response of mice and its plausible mechanism of action, Immunol. Lett., 2020, 220, 21–31 CrossRef PubMed.
- M. Szczypka, M. Lis, M. Kuczkowski, K. Bobrek, A. Pawlak, A. Zambrowicz, A. Gaweł and B. Obmińska-Mrukowicz, Yolkin, a Polypeptide Complex from Egg Yolk, Affects Cytokine Levels and Leukocyte Populations in Broiler Chicken Blood and Lymphoid Organs after In Ovo Administration, Int. J. Mol. Sci., 2023, 24, 17494 CrossRef CAS PubMed.
- W. Kazana, D. Jakubczyk, J. Siednienko, A. Zambrowicz, J. Macała and A. Zabłocka, Mechanism of Molecular Activity of Yolkin-a Polypeptide Complex Derived from Hen Egg Yolk-in PC12 Cells and Immortalized Hippocampal Precursor Cells H19-7, Mol. Neurobiol., 2023, 60, 2819–2831 CrossRef CAS PubMed.
- M. Lemieszewska, M. Jakubik-Witkowska, B. Stańczykiewicz, A. Zambrowicz, A. Zabłocka, A. Polanowski, T. Trziszka and J. Rymaszewska, Pro-Cognitive Properties of the Immunomodulatory Polypeptide Complex, Yolkin, from Chicken Egg Yolk and Colostrum-Derived Substances: Analyses Based on Animal Model of Age-Related Cognitive Deficits, Arch. Immunol. Ther. Exp., 2016, 64, 425–434 CrossRef CAS.
- C. Dima, E. Assadpour, S. Dima and S. M. Jafari, Bioavailability of nutraceuticals: Role of the food matrix, processing conditions, the gastrointestinal tract, and nanodelivery systems, Compr. Rev. Food Sci. Food Saf., 2020, 19, 954–994 CrossRef CAS PubMed.
- L. Lee, K. Samardzic, M. Wallach, L. R. Frumkin and D. Mochly-Rosen, Immunoglobulin Y for Potential Diagnostic and Therapeutic Applications in Infectious Diseases, Front. Immunol., 2021, 12, 696003 CrossRef CAS PubMed.
- T. Trziszka, H. Rozanski and A. Polanowski, Eggs as a Very Promising Source of Biomedical and Nutraceutical Preparations: A Review, J. Life Sci., 2013, 7, 862–877 CAS.
- S. L. DeFelice, The nutraceutical revolution: its impact on food industry R&D, Trends Food Sci. Technol., 1995, 6, 59–61 CrossRef CAS.
- J. M. Walker, in Basic Protein and Peptide Protocols, ed. J. M. Walker, Humana Press, Totowa, NJ, 1994, pp. 5–8 Search PubMed.
- I. Guevara, J. Iwanejko, A. Dembińska-Kieć, J. Pankiewicz, A. Wanat, P. Anna, I. Gołabek, S. Bartuś, M. Malczewska-Malec and A. Szczudlik, Determination of nitrite/nitrate in human biological material by the simple Griess reaction, Clin. Chim. Acta, 1998, 274, 177–188 CrossRef CAS PubMed.
- B. Stańczykiewicz, T. M. Goszczyński, P. Migdał, M. Piksa, K. Pawlik, J. Gburek, K. Gołąb, B. Konopska and A. Zabłocka, Effect of Ovocystatin on Amyloid β 1–42 Aggregation—In Vitro Studies, Int. J. Mol. Sci., 2023, 24, 5433 CrossRef PubMed.
- A. Shevchenko, H. Tomas, J. Havlis, J. V. Olsen and M. Mann, In-gel digestion for mass spectrometric characterization of proteins and proteomes, Nat. Protoc., 2006, 1, 2856–2860 CrossRef CAS PubMed.
- W. Morelle and J.-C. Michalski, Analysis of protein glycosylation by mass spectrometry, Nat. Protoc., 2007, 2, 1585–1602 CrossRef CAS PubMed.
- E. Lisowska, M. Duk and A. M. Wu, Preparation of Biotinylated Lectins and Application in Microtiter Plate Assays and Western Blotting. in: A Laboratory Guide to Biotin-Labeling in Biomolecule Analysis, BioMethods. Birkhäuser, Basel, 1996, 115–129 Search PubMed.
- M. Miguel, I. Recio, J. A. Gómez-Ruiz, M. Ramos and R. López-Fandiño, Angiotensin I-converting enzyme inhibitory activity of peptides derived from egg white proteins by enzymatic hydrolysis, J. Food Prot., 2004, 67, 1914–1920 CrossRef CAS PubMed.
- Z. Yu, Y. Yin, W. Zhao, Y. Yu, B. Liu, J. Liu and F. Chen, Novel peptides derived from egg white protein inhibiting alpha-glucosidase, Food Chem., 2011, 129, 1376–1382 CrossRef CAS.
- I. M. E. Lacroix and E. C. Y. Li-Chan, Inhibition of dipeptidyl peptidase (DPP)-IV and α-glucosidase activities by pepsin-treated whey proteins, J. Agric. Food Chem., 2013, 61, 7500–7506 CrossRef CAS PubMed.
- R. Re, N. Pellegrini, A. Proteggente, A. Pannala, M. Yang and C. Rice-Evans, Antioxidant activity applying an improved ABTS radical cation decolorization assay, Free Radicals Biol. Med., 1999, 26, 1231–1237 CrossRef CAS PubMed.
- J. A. J. Housmans, G. Wu, J. Schymkowitz and F. Rousseau, A guide to studying protein aggregation, FEBS J., 2023, 290, 554–583 CrossRef CAS PubMed.
- D. Chaudhary and F. Waris, Nutraceuticals and its health benefits: a review, MIT International Journal of Pharmaceutical Sciences, 2020, 6, 51–59 Search PubMed.
- M. Janusz and A. Zabłocka, Colostral proline-rich polypeptides – immunoregulatory properties and prospects of therapeutic use in Alzheimer's disease, Curr. Alzheimer Res., 2010, 7, 323–333 CrossRef CAS PubMed.
- P. Edman, A method for the determination of amino acid sequence in peptides, Arch. Biochem., 1949, 22, 475 CAS.
- A. L. Rockwood, M. M. Kushnir and N. J. Clarke, in Principles and Applications of Clinical Mass Spectrometry, ed. N. Rifai, A. R. Horvath and C. T. Wittwer, Elsevier, 2018, pp. 33–65 Search PubMed.
- A. L. Hughes, Life-history evolution at the molecular level: adaptive amino acid composition of avian vitellogenins, Proc. Biol. Sci., 2015, 282, 20151105 Search PubMed.
- R. J. Solá and K. Griebenow, Effects of Glycosylation on the Stability of Protein Pharmaceuticals, J. Pharm. Sci., 2009, 98, 1223–1245 CrossRef PubMed.
- Q. Zhou and H. Qiu, The Mechanistic Impact of N-Glycosylation on Stability, Pharmacokinetics, and Immunogenicity of Therapeutic Proteins, J. Pharm. Sci., 2019, 108, 1366–1377 CrossRef CAS PubMed.
- G. J. Cavallero, M. Landoni and A. S. Couto, In depth N-glycoproteomics shows glyco-features of chicken egg white, Food Biosci., 2020, 35, 100590 CrossRef CAS.
- J. M. M. Diaz, A. B. Moran, S. R. Peel, J. L. Hendel and D. I. R. Spencer, Egg yolk sialylglycopeptide: purification, isolation and characterization of N -glycans from minor glycopeptide species, Org. Biomol. Chem., 2022, 20, 4905–4914 RSC.
- R. G. Elkin, M. B. Freed, S. A. Danetz and C. A. Bidwell, Proteolysis of Japanese quail and chicken plasma apolipoprotein B and vitellogenin by cathepsin D: similarity of the resulting protein fragments with egg yolk polypeptides, Comp. Biochem. Physiol., Part B: Biochem. Mol. Biol., 1995, 112, 191–196 CrossRef CAS PubMed.
- A. Szmyt, A. Zabłocka, J. Macała, J. Chrzanowska and A. Dąbrowska, C-Terminal Fragment of Vitellogenin II, a Potential Yolkin Polypeptide Complex Precursor Protein-Heterologous Expression, Purification, and Immunoregulatory Activity, Int. J. Mol. Sci., 2021, 22, 7223 CrossRef CAS PubMed.
- L. Sheng, Z. He, J. Chen, Y. Liu, M. Ma and Z. Cai, The impact of N-glycosylation on conformation and stability of immunoglobulin Y from egg yolk, Int. J. Biol. Macromol., 2016, 96, 129–136 CrossRef PubMed.
- A. Helenius and M. Aebi, Intracellular Functions of N-Linked Glycans, Science, 2001, 291, 2364–2369 CrossRef CAS PubMed.
- R. J. Simpson, Stabilization of Proteins for Storage, Cold Spring Harbor Protoc., 2010, 2010, pdb.top79 CrossRef PubMed.
- Q. A. Al-Maqtari, J. K. Mohammed, A. A. Mahdi, W. Al-Ansi, M. Zhang, A. Al-Adeeb, M. Wei, H. M. Phyo and W. Yao, Physicochemical properties, microstructure, and storage stability of Pulicaria jaubertii extract microencapsulated with different protein biopolymers and gum arabic as wall materials, Int. J. Biol. Macromol., 2021, 187, 939–954 CrossRef CAS PubMed.
- C. Ramírez-Santana, F. J. Pérez-Cano, C. Audí, M. Castell, M. G. Moretones, M. C. López-Sabater, C. Castellote and A. Franch, Effects of cooling and freezing storage on the stability of bioactive factors in human colostrum, J. Dairy Sci., 2012, 95, 2319–2325 CrossRef PubMed.
- M. Rahban, F. Ahmad, M. A. Piatyszek, T. Haertlé, L. Saso and A. A. Saboury, Stabilization challenges and aggregation in protein-based therapeutics in the pharmaceutical industry, RSC Adv., 2023, 13, 35947–35963 RSC.
- M. Manno, E. F. Craparo, A. Podestà, D. Bulone, R. Carrotta, V. Martorana, G. Tiana and P. L. San Biagio, Kinetics of Different Processes in Human Insulin Amyloid Formation, J. Mol. Biol., 2007, 366, 258–274 CrossRef CAS PubMed.
- A. Cao, D. Hu and L. Lai, Formation of amyloid fibrils from fully reduced hen egg white lysozyme, Protein Sci., 2004, 13, 319–324 CrossRef CAS PubMed.
- Z. Wei, S. Wu, J. Xia, P. Shao, P. Sun and N. Xiang, Enhanced Antibacterial Activity of Hen Egg-White Lysozyme against Staphylococcus aureus and Escherichia coli due to Protein Fibrillation, Biomacromolecules, 2021, 22, 890–897 CrossRef CAS PubMed.
- N. Kummer, T. Wu, K. J. De France, F. Zuber, Q. Ren, P. Fischer, S. Campioni and G. Nyström, Self-Assembly Pathways and Antimicrobial Properties of Lysozyme in Different Aggregation States, Biomacromolecules, 2021, 22, 4327–4336 CrossRef CAS PubMed.
- A. D'Souza, J. D. Theis, J. A. Vrana and A. Dogan, Pharmaceutical amyloidosis associated with subcutaneous insulin and enfuvirtide administration, Amyloid, 2014, 21, 71–75 CrossRef PubMed.
- A. Hawe, M. Sutter and W. Jiskoot, Extrinsic Fluorescent Dyes as Tools for Protein Characterization, Pharm. Res., 2008, 25, 1487–1499 CrossRef CAS PubMed.
- H. L. Svilenov, T. Menzen, K. Richter and G. Winter, Modulated Scanning Fluorimetry Can Quickly Assess Thermal Protein Unfolding Reversibility in Microvolume Samples, Mol. Pharmaceutics, 2020, 17, 2638–2647 CrossRef CAS PubMed.
- S. H. Kim, H. J. Yoo, E. J. Park and D. H. Na, Nano Differential Scanning Fluorimetry-Based Thermal Stability Screening and Optimal Buffer Selection for Immunoglobulin G, Pharmaceuticals, 2021, 15, 29 CrossRef PubMed.
- H. Yao, E. Wynendaele and B. De Spiegeleer, Thermal sensitivity as a quality control attribute for biotherapeutics: The L-asparaginase case, Drug Test. Anal., 2020, 12, 67–77 CrossRef CAS PubMed.
- A. O. Magnusson, A. Szekrenyi, H.-J. Joosten, J. Finnigan, S. Charnock and W.-D. Fessner, nanoDSF as screening tool for enzyme libraries and biotechnology development, FEBS J., 2019, 286, 184–204 CrossRef CAS PubMed.
- A. Zambrowicz, M. Pokora, B. Setner, A. Dąbrowska, M. Szołtysik, K. Babij, Z. Szewczuk, T. Trziszka, G. Lubec and J. Chrzanowska, Multifunctional peptides derived from an egg yolk protein hydrolysate: isolation and characterization, Amino Acids, 2015, 47, 369–380 CrossRef CAS PubMed.
- I. Marcet, J. Delgado, N. Díaz, M. Rendueles and M. Díaz, Peptides recovery from egg yolk lipovitellins by ultrafiltration and their in silico bioactivity analysis, Food Chem., 2022, 379, 132145 CrossRef CAS PubMed.
- W. Liu, M. Zhao, S. Li, D. U. Ahn, N. Chen and X. Huang, Advances in preparation and bioactivity of phosvitin phosphopeptides, J. Future Foods, 2022, 2, 213–222 CrossRef.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d4fo03023k |
| This journal is © The Royal Society of Chemistry 2024 |