
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceDietary quercetin intake is associated with lower ulcerative colitis risk but not Crohn's disease in a prospective cohort study and in vivo experiments†
Shi-Yuan
Lu‡
*a,
Lintao
Dan‡
 bc,
Sishen
Sun
d,
Tian
Fu
b and
Jie
Chen
*bc
bc,
Sishen
Sun
d,
Tian
Fu
b and
Jie
Chen
*bc
aDepartment of Gastroenterology, The Second Affiliated Hospital, Zhejiang University School of, Medicine, 88 Jiefang Road, Hangzhou, 310009, China. E-mail: shiyuan_lu@zju.edu.cn
bDepartment of Gastroenterology, The Third Xiangya Hospital, Central South University, 138 Tongzibo Road, Changsha, China. E-mail: med_chenjie@csu.edu.cn
cCentre for Global Health, Zhejiang University School of Medicine, 866 Yuhangtang Road, Hangzhou 310058, China. E-mail: med_chenjie@zju.edu.cn
dDepartment of Gastroenterology, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, China
First published on 11th May 2024
Abstract
Objectives: Previous preclinical evidence indicates a protective role of quercetin against inflammatory bowel disease (IBD). However, there is no evidence from human populations, resulting in knowledge gaps regarding the role of quercetin in the IBD development. We aimed to prospectively evaluate the associations between dietary quercetin intake and IBD in humans and in vivo animal models. Methods: We included 187![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 709 IBD-free participants from the UK Biobank. Dietary information was collected using validated 24-hour dietary recalls and the quercetin intake was estimated based on national nutrient databases. Incident IBD was ascertained via inpatient and primary care data. Cox proportional hazard models were used to estimate the multi-variable adjusted hazard ratios (aHRs) and 95% confidence intervals (CIs). Experiments were conducted in two chemical-induced (dextran sulfate sodium salt and trinitro-benzene-sulfonic acid) mouse models orally pretreated with quercetin (CAS number: 117-39-5) solution to evaluate the effects of quercetin at physiological levels. Results: After a mean follow-up of 9.7 years, we documented 863 incident IBD. Compared to participants with the lowest quintile intake of quercetin, those in the highest quintiles were associated with a lower risk of IBD (aHR 0.76, 95% CI 0.60–0.95; P-trend = 0.004) and ulcerative colitis (aHR 0.69, 95% CI 0.53–0.91; P-trend = 0.001), but not Crohn's disease (aHR 0.95, 95% CI 0.62–1.45; P-trend = 0.765). Mouse models showed that pretreatment with quercetin could attenuate the chemically induced colitis. Conclusions: Higher quercetin intake was associated with a lower risk of IBD, especially UC. The protective role of quercetin is promising in humans and warrants further investigation into downstream mechanisms.
709 IBD-free participants from the UK Biobank. Dietary information was collected using validated 24-hour dietary recalls and the quercetin intake was estimated based on national nutrient databases. Incident IBD was ascertained via inpatient and primary care data. Cox proportional hazard models were used to estimate the multi-variable adjusted hazard ratios (aHRs) and 95% confidence intervals (CIs). Experiments were conducted in two chemical-induced (dextran sulfate sodium salt and trinitro-benzene-sulfonic acid) mouse models orally pretreated with quercetin (CAS number: 117-39-5) solution to evaluate the effects of quercetin at physiological levels. Results: After a mean follow-up of 9.7 years, we documented 863 incident IBD. Compared to participants with the lowest quintile intake of quercetin, those in the highest quintiles were associated with a lower risk of IBD (aHR 0.76, 95% CI 0.60–0.95; P-trend = 0.004) and ulcerative colitis (aHR 0.69, 95% CI 0.53–0.91; P-trend = 0.001), but not Crohn's disease (aHR 0.95, 95% CI 0.62–1.45; P-trend = 0.765). Mouse models showed that pretreatment with quercetin could attenuate the chemically induced colitis. Conclusions: Higher quercetin intake was associated with a lower risk of IBD, especially UC. The protective role of quercetin is promising in humans and warrants further investigation into downstream mechanisms.
1. Introduction
Inflammatory bowel disease (IBD), including Crohn's disease (CD) and ulcerative colitis (UC), is an immune-related chronic gastrointestinal disease with a rapidly increasing global incidence.1 As a crucial modifiable behavioral risk factor, the diet has received widespread attention in the aetiopathogenesis of IBD.1 Among phytonutrients, there is emerging evidence of various flavonoids to play an important role in the development of IBD. They may be useful in alleviating acute or chronic intestinal inflammation through different mechanisms, including protecting against oxidative stress, enhancing intestinal epithelial barrier function, and modulating the immune response in the gut.2Quercetin is a flavonoid commonly enriched in many vegetables and fruits with various pharmacological functions.3,4 A meta-analysis of 11 preclinical experiments concluded a protective effect of quercetin against IBD in animal models via multiple mechanisms.5 However, there is no available evidence in humans. Previous randomized clinical trials reported that quercetin supplementation could significantly improve disease activity and reduce serum tumor necrosis factor (TNF)-α levels in patients with rheumatoid arthritis compared with the placebo group.6 In addition, as top sources of dietary quercetin, tea,7 vegetables,8 and fruits8 have been linked to a lower risk of IBD. These pieces of evidence make it rational for the possible benefits of quercetin in IBD prevention, but there are still knowledge gaps in further understanding the role of quercetin in the development of IBD in humans.
Therefore, in a prospective cohort of 187709 participants free of IBD, we aimed to examine the association between dietary quercetin intake and the subsequent development of IBD and of CD and UC separately. Secondly, we aimed to examine this association in different participant subgroups to identify populations that may benefit more from dietary quercetin intake. Animal experiments in mouse models were conducted to evaluate the effects of quercetin at physiological levels in vivo.
2. Methods
2.1 Study population
This study leveraged data from the UK Biobank, which is an ongoing national prospective cohort project that enrolled over half a million volunteers from 22 assessment centers in the UK between 2006 and 2010.9,10 All participants have signed an electronic consent and received a touchscreen questionnaire, a verbal interview, a physical examination, and biological sample collection. The details are described in detail elsewhere.9 The North West–Haydock Research Ethics Committee granted ethical approval to use the UK Biobank database (REC reference: 16/NW/0274).A web-based 24-hour dietary recall questionnaire (Oxford WebQ) was administered for five rounds in April 2009–September 2010, February 2011–April 2011, June 2011–August 2011, October 2011–December 2011, and April 2012–June 2012. In the present study (Fig. 1), a subset of UK Biobank participants has available 24-hour dietary data (n = 210948). Then, we excluded: (1) participants reporting implausible energy intake (defined as <800 or >4200 kcal per day for males, <600 or >3500 kcal per day for females)11 or a non-typical diet (n = 19883);12 (2) participants with a dietary quercetin intake outside the normal distribution (n = 978) based on Rosner's generalized extreme studentized deviate test;13 (3) participants with diagnosis of IBD before the baseline (i.e., the completion date of the last round of recalls in this study, n = 2378). This study followed the strengthening the reporting of observational studies in epidemiology (STROBE) guidelines.
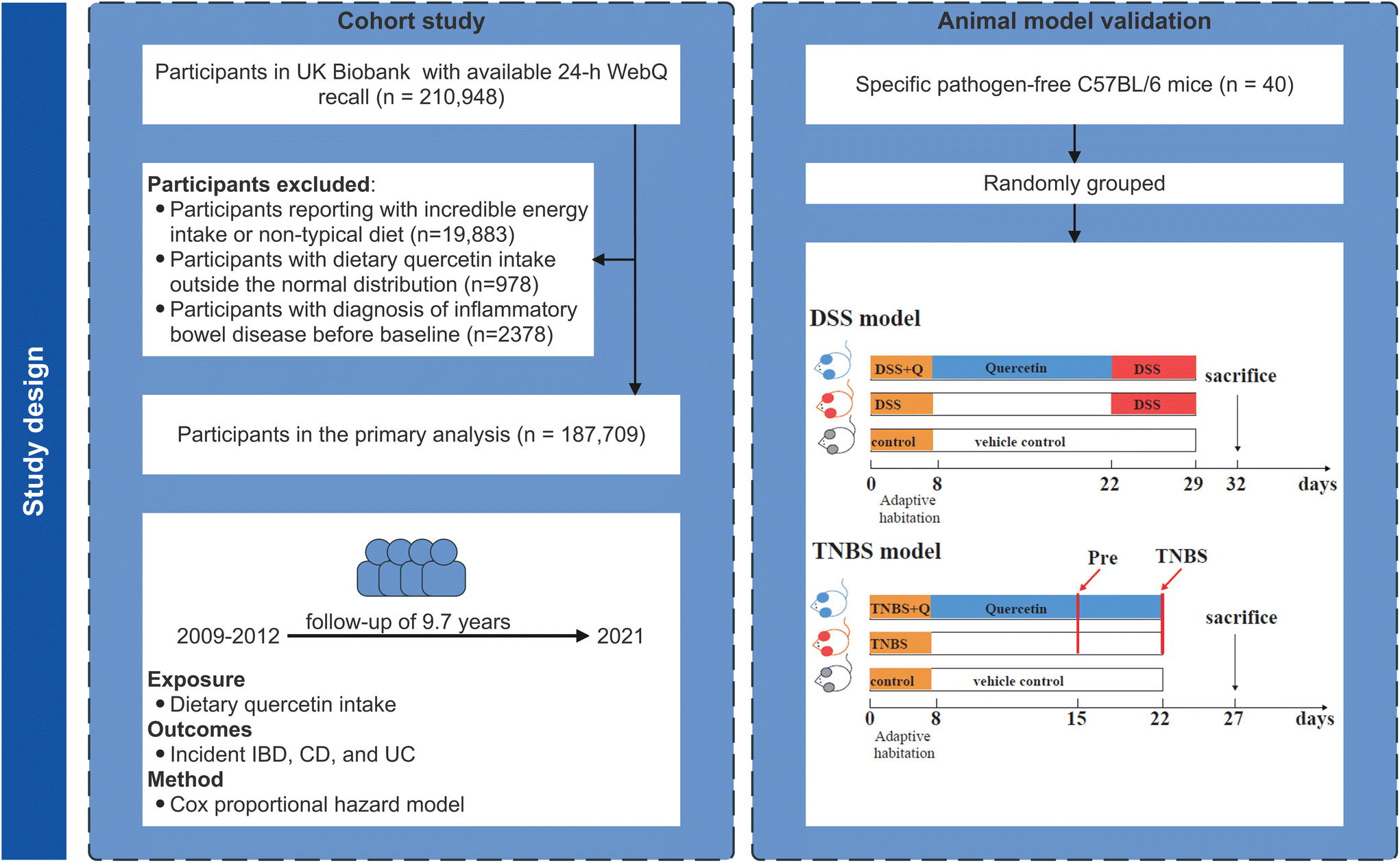 | ||
| Fig. 1 Flow chart of the current study. | ||
2.2 Assessment of quercetin intake
The dietary information was collected using a 24 h WebQ. Participants were presented with a list of up to 206 foods and 32 beverages commonly consumed in the UK and selected the number of portions consumed from each food. The exposure was the daily intake of quercetin from the diet and was estimated using the sum of the portion size multiplied by the corresponding quercetin content of each food. The portion size recorded in grams of each food was obtained from the publication by the Food Standards Agency.14 The quercetin content (in mg per 100 g) was obtained from the United States Department of Agriculture (USDA) database on flavonoids15 and expanded with values from Phenol-Explorer,16 a French database on polyphenols. The two databases were previously applied to estimate dietary intakes of flavonoids in European and other populations and the results show that the intake estimates from both are in good agreement.17–19 These studies suggested the rationale for using these databases to estimate participants in the current cohort.The web-based 24-hour dietary assessment was compared with an interviewer-administered 24-hour recall completed on the same day, and Spearman correlation coefficients calculated from the web-based questionnaire ranged from 0.5 and 0.9 (mean 0.6) for most nutrients.20 In the subgroup of participants who completed all five rounds of WebQ (n = 2218), comparing dietary measures based on a single recall versus the mean of all completed questionnaires, the Pearson correlation coefficients of the quercetin intake between the two measures were 0.703–0.767. Moreover, 86%–93% of participants were classified into the same or adjacent category of quercetin intake between the measures from a single recall and the average of five recalls.
In addition, a single round of a 24-hour WebQ recall has also shown good agreement with long-term consumption and frequency of food groups collected by baseline food frequency questionnaires.21 Therefore, we included participants with at least one 24 h recall in the primary analysis, given the sample size and the incidence of outcomes.
2.3 Ascertainment of outcomes
The primary outcomes of interest were inflammatory bowel disease during the follow-up, obtained through hospital inpatient data (International Classification of Disease ninth and tenth editions [ICD-9 and ICD-10]) and coded primary care data. Incident IBD cases were defined as CD (ICD-9 codes 555, ICD-10 codes K50) or UC (ICD-9 codes 556, ICD-10 codes K51). We use a single diagnostic code in the primary analysis to maximize the statistical power. Previous studies in the UK had reported acceptable diagnostic accuracy using at least one diagnosis code in inpatient data (PPV 87%)22 and primary care data (sensitivity 94%).23 Moreover, the UK Audit Commission review of 2009 to 2010 concluded that a single diagnostic coding ICD-10 has an overall accuracy of 87%.24 Therefore, we used a single diagnostic code to identify IBD in the primary analysis. A stricter definition, using at least two diagnostic codes or one diagnostic code plus one prescription that was described in detail in a previous study,25 was applied in a sensitivity analysis.Participants were followed up from the completed date of the last available 24-h WebQ to the date of incidence of IBD, death, loss, or the end of the follow-up (latest updated time for health data, September 2021 for participants in England, July 2021 for participants in Scotland, and February 2018 for participants in Wales), whichever came first.
2.4 Assessment of covariates
In this study, we used predefined covariates. We included age, sex (female and male), race (white and non-white), education level (college and below college), smoking status (never, previous or current), physical activity level (adequate and inadequate),26 and body mass index (BMI). We controlled the socioeconomic status using the Townsend Deprivation Index (TDI).27 Baseline comorbidities were evaluated using the Charlson Comorbidity Index (CCI).28 Serum C-reactive protein (CRP) was measured by immunoturbidimetric-high sensitivity analysis on a Beckman Coulter AU5800, with full details regarding sample and data processing available on the UKB website.29 Medication information was recorded in interviews with trained nurses, including the use of non-steroidal anti-inflammatory drugs (NSAIDs), proton pump inhibitors, antibiotics, and oral contraceptive pills. Dietary variables were collected using the 24 h WebQ, including total energy, common nutrients, and food groups. The details of nutrient and food group calculation are published elsewhere.30 Details of the definition and process of the covariates are presented in Table S1.† The intake of ultra-processed food defined by NOVA classification was included in the sensitivity analysis, the calculation of which has been described previously.31 If covariate information was missing or recorded as “unknown”, we imputed the median values for continuous variables or applied a most frequently used category for categorical variables.2.5 Statistical analysis for cohort study
Baseline characteristics were displayed by the quintiles of dietary quercetin intake. Characteristics were summarized in number (percentage) for categorical variables and in mean (standard deviation (SD)) for continuous variables. The associations between quercetin intake in quintiles and subsequent IBD, CD, and UC were first presented as a cumulative incidence plot. The Cox model was applied to estimate the hazard ratios (HRs) and 95% confidence intervals (CIs). Four multivariable models were constructed: model 1 (minimally adjusted model) adjusted for age, sex, and ethnic background; model 2 further adjusted for physical activity, TDI, smoking status, alcoholic drink (excluding wine) consumption, and the BMI based on model 1; model 3 further adjusted for total energy based on model 2; model 4 (fully adjusted model) further adjusted for the intake of cereal, meat, fish, and sugar-sweetened beverages based on model 3. The usage of oral contraceptive pills was included in the fully adjusted model when investigating associations in females. Proportional hazard assumptions were confirmed using a weighted residual method (P > 0.79).To visualize the association between quercetin and IBD, we also used restricted cubic splines (RCS) with three knots at the 10th, 50th, and 90th percentile based on the fully adjusted mode. We set the zero intake as the reference level for analysis using the RCS method. In a secondary analysis, we evaluated associations of energy-adjusted quercetin intake and quercetin intake by body weight with IBD. The energy-adjusted quercetin was calculated using the residual method, which can mitigate the effects of measurement error in data collected using self-reported dietary assessment instruments.32
Subgroup analyses were conducted and multiplication interactions were tested in population by age (<60, ≥60), sex, smoking status, alcohol consumption (none, moderate, heavy), physical activity, and BMI (<30, ≥30 kg m−2). The categorization for physical activity and the BMI in the subgroup analysis was based on the same definition found to be associated with an elevated risk of IBD in previous studies.33 We also conducted several sensitivity analyses: (1) lagged analysis excluding IBD within the first 2 and 4 years of follow-up; (2) restricting the analysis to individuals at 2 rounds of 24 h recalls; (3) further adjusting for CCI; (4) further adjusting for IBD-related medication; (5) further adjusting for CRP and conducting mediation analysis to quantify the contribution of CRP in quercetin-IBD/UC associations using R packages “mediation” (version 4.5.0) with 1000 bootstrap samples to estimate the bias-corrected bootstrap CI; (6) additionally adjusting for ultra-processed food and n-3 polyunsaturated fatty acid consumption; (7) identifying individuals with IBD using at least two medical records. All analyses were performed using R (version 4.2.1; R Development Core Team), and all statistical tests were two-sided, with a P value < 0.05 considered as statistically significant.
2.6 Validation in animal models
Animal experiments were conducted to verify the inverse associations between the quercetin intake and subsequent IBD. The objective of the animal experiments was to supplementally validate the credibility of observational studies in a framework and the quercetin dose matched our cohort study, not solely to pursue novel mechanistic discoveries. C57BL/6 male mice were purchased from and raised at Shanghai Model Organisms Center, Co., Ltd (Shanghai, China). All animals were randomly grouped and hosted under standardized specific pathogen-free (SPF) conditions with ad libitum access to water and chow. Quercetin (Solarbio, SQ8030; CAS number: 117-39-5) was dissolved in 2%-Tween 80 (Sigma, P4780)-containing ddH2O. Referring to the dosage of previous studies, we chose the dose of 15 mg kg−1 daily for quercetin treatment in mice, which was approximately equal to 84 mg d−1 in humans. After a one-week adaptive growth period, mice were gavaged with quercetin solution daily (15 mg kg−1) for 14 days. Then, we took the classical Dextran Sulfate Sodium Salt (DSS)-induced and trinitro-benzene-sulfonic acid (TNBS)-induced mouse models to establish the mouse models of UC (Fig. 1). We evaluated the disease activity, histological presentation, and specific mRNA expression among different groups. The detailed reagents, experiments, and analysis are provided in the ESI.† All mouse experiments were conducted as per the National Institutes of Health Guidelines for the Care and Use of Laboratory Animals. All of the study procedures were approved by the Institutional Animal Care and Use Committee of The Second Affiliated Hospital, School of Medicine, Zhejiang University (iACUC No.2023-130).3. Results
3.1 Characteristics of the study sample
In this study, 187709 individuals were included and followed up for a mean (SD) of 9.7 (1.6) years. Baseline characteristics are summarized in Table 1. Among the included study samples, the mean (SD) age was 59.0 (8.0) years and 103241 (55.0%) were female. The mean (SD) intake of quercetin was 22.61 (18.52) mg d−1. Compared with participants in the lowest quintiles of quercetin intake, those in the higher categories are more likely to be non-smokers, with a higher educational level, a lower BMI, and more physical activity.
| Characteristics | Total (n = 187709) |
Dietary quercetin intake in quintiles (lowest to highest, mg d−1) | ||||
|---|---|---|---|---|---|---|
| Q1 (n = 37542, 0–<8.9) |
Q2 (n = 37542, 8.9–<15.0) |
Q3 (n = 37541, 15.0–<21.6) |
Q4 (n = 37542, 21.6–<31.9) |
Q5 (n = 37542, ≥31.9) |
||
| BMI, body mass index; SD, standard deviation.a Mean (SD) values and percentages are reported for continuous and categorical variables, respectively.b Including intake of beer/cider, spirits, and other alcoholic drinks. | ||||||
| Age at the baseline (mean (SD)) | 59.04 (8.01) | 57.15 (8.19) | 58.83 (8.00) | 59.43 (7.89) | 59.84 (7.80) | 59.97 (7.81) |
| Sex (%) | ||||||
| Female | 103241 (55.0) |
17088 (45.6) |
19437 (51.8) |
20590 (54.8) |
22152 (59.0) |
23948 (63.8) |
| Male | 84468 (45.0) |
20390 (54.4) |
18105 (48.2) |
16951 (45.2) |
15390 (41.0) |
13594 (36.2) |
| Townsend deprivation index (mean (SD)) | −1.60 (2.86) | −1.26 (3.02) | −1.66 (2.82) | −1.71 (2.80) | −1.69 (2.80) | −1.66 (2.84) |
| Education (%) | ||||||
| Below college degree | 106976 (57.3) |
25584 (68.8) |
22146 (59.3) |
20387 (54.5) |
19077 (51.0) |
19735 (52.8) |
| College degree | 79823 (42.7) |
11621 (31.2) |
15202 (40.7) |
17001 (45.5) |
18322 (49.0) |
17662 (47.2) |
| Ethnicity (%) | ||||||
| White | 179449 (95.9) |
35229 (94.4) |
35859 (95.9) |
36178 (96.7) |
36192 (96.7) |
35939 (96.1) |
| Others | 7595 (4.1) | 2109 (5.6) | 1546 (4.1) | 1245 (3.3) | 1220 (3.3) | 1464 (3.9) |
| Smoking status (%) | ||||||
| Never smoked | 106358 (56.8) |
19731 (52.8) |
21070 (56.3) |
21419 (57.2) |
21568 (57.6) |
22525 (60.1) |
| Previous or current smokers | 80868 (43.2) |
17631 (47.2) |
16371 (43.7) |
16025 (42.8) |
15888 (42.4) |
14934 (39.9) |
| Physical activity (%) | ||||||
| Inadequate | 53357 (28.4) |
13192 (35.2) |
11491 (30.6) |
10378 (27.6) |
9358 (24.9) | 8915 (23.7) |
| Adequate | 134352 (71.6) |
24286 (64.8) |
26051 (69.4) |
27163 (72.4) |
28184 (75.1) |
28627 (76.3) |
| BMI, kg m −2 (mean (SD)) | 26.91 (4.63) | 27.96 (4.97) | 27.19 (4.66) | 26.77 (4.51) | 26.37 (4.34) | 26.24 (4.45) |
| Charlson comorbidity index (mean (SD)) | 0.24 (0.83) | 0.25 (0.85) | 0.23 (0.83) | 0.22 (0.78) | 0.22 (0.80) | 0.25 (0.88) |
| Non-steroidal anti-inflammatory drugs use (%) | 68419 (36.4) |
14843 (39.6) |
13853 (36.9) |
13366 (35.6) |
13216 (35.2) |
13118 (34.9) |
| Proton pump inhibitors use (%) | 15168 (8.1) |
3566 (9.5) | 3062 (8.2) | 2882 (7.7) | 2695 (7.2) | 2958 (7.9) |
| Antibiotic use (%) | 2124 (1.1) | 465 (1.2) | 469 (1.2) | 389 (1.0) | 372 (1.0) | 428 (1.1) |
| Alcohol, g d −1 (mean (SD)) | 15.22 (20.69) | 14.11 (21.52) | 15.49 (20.34) | 16.58 (20.95) | 16.71 (21.25) | 13.22 (19.10) |
| Total energy, KJ d −1 (mean (SD)) | 8464.46 (2265.29) | 8056.27 (2411.93) | 8298.58 (2217.72) | 8456.86 (2171.47) | 8635.49 (2177.10) | 8877.20 (2250.93) |
| Quercetin, mg d −1 (mean (SD)) | 22.61 (18.52) | 4.98 (2.53) | 11.98 (1.78) | 18.18 (1.89) | 26.10 (2.90) | 51.84 (19.58) |
| From vegetables | 4.13 (3.80) | 1.66 (1.84) | 3.33 (2.70) | 4.18 (3.25) | 5.32 (3.84) | 6.14 (4.91) |
| From fruits | 3.98 (4.21) | 1.15 (1.67) | 2.78 (2.62) | 3.91 (3.32) | 5.45 (4.14) | 6.60 (5.68) |
| From alcoholic beverages | 1.34 (2.63) | 0.40 (1.08) | 1.15 (2.17) | 1.63 (2.83) | 1.94 (3.22) | 1.59 (2.99) |
| From non-alcoholic beverages | 7.30 (15.21) | 0.87 (0.87) | 1.45 (1.84) | 2.27 (3.07) | 4.47 (5.56) | 27.46 (24.42) |
| Cereal intake, g d −1 (mean (SD)) | 52.72 (69.63) | 17.70 (22.37) | 27.75 (33.08) | 51.61 (63.65) | 77.63 (81.91) | 88.91 (90.04) |
| Meat intake, g d −1 (mean (SD)) | 86.68 (70.06) | 94.36 (76.53) | 91.31 (69.51) | 86.99 (67.76) | 82.82 (67.08) | 77.90 (67.73) |
| Fish intake, g d −1 (mean (SD)) | 26.07 (41.65) | 21.49 (42.19) | 24.12 (40.01) | 26.48 (41.04) | 28.38 (41.45) | 29.88 (42.97) |
| Ultra-processed food intake, serving per d (mean (SD)) | 8.48 (4.47) | 9.52 (4.92) | 8.72 (4.44) | 8.33 (4.32) | 7.91 (4.15) | 7.92 (4.28) |
| Sugar-sweetened beverage intake, serving per d (mean (SD)) | 0.28 (0.63) | 0.40 (0.78) | 0.30 (0.64) | 0.26 (0.59) | 0.23 (0.54) | 0.24 (0.57) |
| Alcoholic drink intake (excl. wine), g d −1 (mean (SD)) | 127.48 (369.62) | 225.62 (538.75) | 144.67 (381.85) | 113.65 (322.44) | 89.27 (272.98) | 64.18 (230.20) |
| Carbohydrate intake, g d −1 (mean (SD)) | 251.77 (75.02) | 233.59 (77.72) | 243.29 (71.97) | 249.59 (70.70) | 258.30 (71.97) | 274.07 (76.10) |
| Dietary fiber intake, g d −1 (mean (SD)) | 18.06 (6.57) | 13.49 (5.17) | 16.64 (5.21) | 18.31 (5.55) | 20.13 (6.04) | 21.75 (7.35) |
| Fat intake, g d −1 (mean (SD)) | 71.25 (27.02) | 70.89 (28.60) | 70.81 (26.85) | 70.87 (26.45) | 71.11 (26.26) | 72.57 (26.83) |
| Protein intake, g d −1 (mean (SD)) | 80.01 (23.26) | 75.62 (25.04) | 78.70 (22.69) | 80.08 (22.05) | 81.71 (22.18) | 83.93 (23.38) |
| n-3 polyunsaturated fatty acid, g d −1 (mean (SD)) | 1.96 (1.01) | 1.78 (0.97) | 1.89 (0.96) | 1.96 (0.98) | 2.04 (1.01) | 2.14 (1.08) |
| n-6 polyunsaturated fatty acid, g d −1 (mean (SD)) | 10.73 (5.04) | 10.14 (5.00) | 10.43 (4.87) | 10.65 (4.88) | 11.00 (5.02) | 11.43 (5.33) |
3.2 Primary and secondary analyses
Throughout the follow-up, we documented 863 IBD events (4.74 cases per 10000 person-years) including 256 CD (1.40 cases per 10000 person-years) and 607 UC (3.33 cases per 10000 person-years). As shown in Fig. 2, inverse associations were observed between quercetin and incident IBD and UC, but not CD. Based on the fully adjusted model, the dose–response associations of the dietary quercetin intake with IBD and UC seemed to reach a plateau at an intake amount of about 35 mg d−1 (both P-nonlinearity < 0.05). We observed that a higher intake of dietary quercetin was significantly associated with a lower risk of IBD and UC in different multi-variable adjusted models (Fig. S1, Table S2† and Table 2). Specifically, compared to individuals in the lowest quintiles of quercetin intake, the HRs (95% CIs) of the second to highest quintiles in the fully-adjusted model were 0.80 (0.65–0.98), 0.73 (0.60–0.91), 0.65 (0.52–0.82) and 0.76 (0.60–0.95) for IBD and 0.74 (0.59–0.94), 0.68 (0.53–0.87), 0.56 (0.43–0.74), 0.69 (0.53–0.91) for UC. We observed consistent inverse associations of energy-adjusted quercetin intake and quercetin intake by body weight with the risk of IBD and UC (all P-trend < 0.05, Table 3).
 | ||
| Fig. 2 Associations of dietary quercetin with the risk of IBD, CD, and UC using restricted cubic curves. The results were based on the fully adjusted model. The vertical axis represents the risk of IBD based on the fully adjusted model. The solid line in orange represents hazard ratios, and the dashed lines in orange represent 95% confidence intervals. Density plots of quercetin are presented in blue. CI, confidence interval; HR, hazard ratio. | ||
| Quercetin intake (mg d−1) | Case/person-years | Crude model | Minimally adjusted model (model 1)a | Fully adjusted model (model 4)b |
|---|---|---|---|---|
| HR (95% CI) | HR (95% CI) | HR (95% CI) | ||
| BMI, body mass index; CD, Crohn's disease; CI, confidence interval; HR, hazard ratio; IBD, inflammatory bowel disease; Ref, reference group; TDI, Townsend deprivation index; UC, ulcerative colitis.a Based on the minimally adjusted model adjusted for age, sex, and ethnic background.b Further adjusted for physical activity, BMI, TDI, smoking status, alcoholic drink consumption (exclude wine), intake of cereal, meat, fish, and sugar-sweetened beverages, and total energy based on model 1.c P < 0.05. Bold values indicate significant associations with P values < 0.05. | ||||
| Inflammatory bowel disease | ||||
| Q1 (0–<8.9) | 227/365342 |
Ref | Ref | Ref |
| Q2 (8.9–<15.0) | 174/358585 |
0.78 (0.64, 0.95) | 0.78 (0.64, 0.95) | 0.80 (0.65, 0.98) |
| Q3 (15.0–<21.6) | 159/358282 |
0.71 (0.58, 0.87) | 0.71 (0.58, 0.87) | 0.73 (0.60, 0.91) |
| Q4 (21.6–<31.9) | 141/357536 |
0.63 (0.51, 0.78) | 0.63 (0.51, 0.78) | 0.65 (0.52, 0.82) |
| Q5 (≥31.9) | 162/356681 |
0.73 (0.60, 0.89) | 0.73 (0.60, 0.90) | 0.76 (0.60, 0.95) |
| P-trend | <0.001 | <0.001 | 0.004 | |
| Crohn's disease | ||||
| Q1 (0–<8.9) | 58/369497 |
Ref | Ref | Ref |
| Q2 (8.9–<15.0) | 53/363798 |
0.93 (0.64, 1.36) | 0.94 (0.64, 1.36) | 0.96 (0.66, 1.41) |
| Q3 (15.0–<21.6) | 48/363588 |
0.85 (0.58, 1.24) | 0.85 (0.58, 1.25) | 0.90 (0.60, 1.33) |
| Q4 (21.6–<31.9) | 48/362847 |
0.85 (0.58, 1.24) | 0.86 (0.58, 1.26) | 0.92 (0.61, 1.39) |
| Q5 (≥31.9) | 49/362093 |
0.87 (0.59, 1.27) | 0.89 (0.60, 1.30) | 0.95 (0.62, 1.45) |
| P-trend | 0.377 | 0.447 | 0.765 | |
| Ulcerative colitis | ||||
| Q1 (0–<8.9) | 169/368980 |
Ref | Ref | Ref |
| Q2 (8.9–<15.0) | 121/363462 |
0.73 (0.58, 0.92) | 0.72 (0.57, 0.91) | 0.74 (0.59, 0.94) |
| Q3 (15.0–<21.6) | 111/363321 |
0.67 (0.53, 0.85) | 0.66 (0.52, 0.84) | 0.68 (0.53, 0.87) |
| Q4 (21.6–<31.9) | 93/362627 |
0.56 (0.44, 0.72) | 0.56 (0.43, 0.72) | 0.56 (0.43, 0.74) |
| Q5 (≥31.9) | 113/361764 |
0.68 (0.54, 0.87) | 0.68 (0.54, 0.87) | 0.69 (0.53, 0.91) |
| P-trend | <0.001 | <0.001 | 0.001 | |
| Daily quercetin intake | Inflammatory bowel disease | Crohn's disease | Ulcerative colitis | |||
|---|---|---|---|---|---|---|
| Case/person-years | HR (95% CI)a | Case/person-years | HR (95% CI) | Case/person-years | HR (95% CI) | |
| BMI, body mass index; CD, Crohn's disease; CI, confidence interval; HR, hazard ratio; IBD, inflammatory bowel disease; Ref, reference group; TDI, Townsend deprivation index; UC, ulcerative colitis.a Based on the fully adjusted model for age, sex, and ethnic background, adjusted for physical activity, BMI, TDI, smoking status, alcoholic drink consumption (excluding wine), intake of cereal, meat, fish, and sugar-sweetened beverages, and total energy.b The quercetin intake was adjusted by the energy intake using the residual method. Adjusted quercetin was the residual from a regression model in which the total energy intake is the independent variable and the absolute nutrient intake is the dependent variable.c P < 0.05.d Calculated using quercetin intake divided by participants’ weight. Bold values indicate significant associations with P values < 0.05. | ||||||
| Intake adjusted by energy (mg) | ||||||
| Q1 (<9.1) | 222/365568 |
Ref | 56/369760 |
Ref | 166/369244 |
Ref |
| Q2 (9.1–15.2) | 186/358614 |
0.90 (0.73, 1.09) | 56/363795 |
1.08 (0.74, 1.58) | 130/363422 |
0.83 (0.66, 1.05) |
| Q3 (15.2–<21.6) | 150/358425 |
0.73 (0.59, 0.91) | 45/363717 |
0.90 (0.60, 1.35) | 105/363494 |
0.68 (0.52, 0.87) |
| Q4 (21.6–<31.7) | 145/357816 |
0.72 (0.57, 0.90) | 48/363154 |
1.00 (0.66, 1.51) | 97/362898 |
0.62 (0.47, 0.82) |
| Q5 (≥31.7) | 160/356668 |
0.80 (0.64, 1.01) | 51/362067 |
1.08 (0.71, 1.63) | 109/361772 |
0.71 (0.54, 0.93) |
| P-trend | 0.011 | 0.989 | 0.002 | |||
| Intake according to weight (mg kg −1 ) | ||||||
| Q1 (<0.1) | 229/364320 |
Ref | 55/368557 |
Ref | 174/367999 |
Ref |
| Q2 (0.1–<0.2) | 184/358031 |
0.85 (0.70, 1.04) | 56/363080 |
1.10 (0.75, 1.61) | 128/362709 |
0.77 (0.61, 0.97) |
| Q3 (0.2–<0.3) | 161/357758 |
0.76 (0.61, 0.94) | 54/362913 |
1.11 (0.75, 1.64) | 107/362709 |
0.65 (0.50, 0.83) |
| Q4 (0.3–<0.45) | 143/356824 |
0.69 (0.55, 0.86) | 41/362330 |
0.88 (0.57, 1.36) | 102/362016 |
0.62 (0.48, 0.81) |
| Q5 (≥0.45) | 145/356113 |
0.71 (0.56, 0.89) | 50/361523 |
1.10 (0.71, 1.68) | 95/361306 |
0.59 (0.44, 0.78) |
| P-trend | 0.001 | 0.971 | <0.001 | |||
3.3 Subgroup and sensitivity analyses
In subgroup analyses, the inverse associations between quercetin and UC remained significant across age groups and sex (Table S3,† all P-trend < 0.05). When stratified by the smoking status, alcohol consumption, and physical activity level, not all subgroups showed a statistically significant inverse trend between a higher dietary quercetin intake and a lower risk of UC. However, we did not detect evidence of heterogeneity in the associations across these subgroups (Tables S4 and S5,† all P-interaction > 0.05). The direction of the associations was consistent with the primary analysis and the observed non-significant associations may be due to the inadequate statistical power. When comparing extreme quintile intake of dietary quercetin, the HR (95% CI) for UC risk was 0.59 (0.43–0.83) among individuals with BMI < 30 kg m−2 and 0.89 (0.52–1.54) among individuals with a higher BMI (Table S5†). Although the overall trend of the dietary quercetin–UC association remained inverse in individuals with obesity, it was observed that this association appeared to be statistically different from that in individuals with a lower BMI (P-interaction = 0.038).In the sensitivity analysis (Table S6†), we obtained similar results after excluding incident IBD in the first 2/4 years of follow-up, restricting to individuals with at least 2 rounds of 24 h recalls, further adjusting for CCI/IBD-related medication/CRP/ultra-processed food/n-6 polyunsaturated fatty acids, or when identifying individuals with IBD using at least two medical records only. Mediation analysis showed that CRP partially mediated the associations between quercetin and IBD and UC risk (Table S7,† mediating proportion = 7.15% [95% CI 0.54–11.31%] and 2.74% [95% CI 1.00–4.94%]).
3.4 Evaluation in an animal model
The flow frame of the experiment is shown in Fig. 1. We found that pretreatment of quercetin (15 mg kg−1, oral, daily) for 14 days before colitis induction could significantly reduce the DAI score (Fig. 3A and B) and the histological score (Fig. 3C and D). PAS-AB stain and immunohistochemistry stain of Claudin-1 (Fig. 3E and F) showed that quercetin pretreatment could protect the mouse mucosal barrier. | ||
| Fig. 3 Quercetin pretreatment could partially prevent ulcerative colitis in mouse models. (A and B) Disease activity index (DAI) score of each experimental group in DSS-induced (A) and TNBS-induced (B) mouse models. (C and D) Histological score of each experiment. (E and F) Representative image of Hematoxylin–Eosin (HE) stain, PAS-AB stain, and Claudin-1 stain in each experimental group. (G and H) Relative gene expression (referred to the colitis group) of inflammatory markers between the colitis group and the quercetin pretreated colitis group in DSS-induced (G) and TNBS-induced (H) mouse models. (I and J) Relative gene expression (referred to the colitis group) of Cxcl9 between the colitis group and the quercetin pretreated colitis group. | ||
Then, we conducted quantified real-time PCR of some common inflammatory markers in each group. We saw reduced expression of several reported inflammation-related genes (S100a8, S100a9, Lcn2) in colonic tissues (Fig. 3G and H), of which, S100a8 and S100a9 were components of calprotectin. Interestingly, we also found a decreased Cxcl9 mRNA expression level in colonic tissues of quercetin-pretreated mice (Fig. 3I and J), which was the potential target for UC treatment that we unearthed in a previous work.34 Therefore, we have demonstrated that pretreatment of quercetin can attenuate colitis in mouse models and contribute to intestinal barrier hemostasis.
4. Discussion
In the current study, we demonstrated that a higher quercetin intake was associated with 24% and 31% reduced risk of IBD and UC, respectively, when comparing extreme quintile intake. We did not observe significant associations between dietary quercetin intake and CD incidence. The associations were consistent in most subgroup and sensitivity analyses. In the in vivo mouse experiment, we found that oral pretreatment with quercetin can partially prevent chemical-induced inflammation in mouse models.The prevention and therapeutic potential of food or nutrients are receiving growing attention, such as quercetin. Previous preclinical experiments conclude a protective effect of quercetin against IBD in animal models via multiple mechanisms but lack the results in humans. Currently, there is no evidence directly linked to the quercetin intake and the development of IBD and UC. Previous umbrella reviews concluded an inverse association of tea consumption, vegetable consumption, and fruit consumption with UC,7 which were reported to be top sources of quercetin in UK.35 However, all results from these meta-analyses were mostly extracted from studies that comprehensively investigated food groups as dietary risk factors with a lack of separate estimates of specified food and nutrients. Therefore, our study provided a novel finding in humans that directly linked higher dietary quercetin to lower risk of IBD and UC.
We did not observe a significant association between dietary quercetin and CD. A possible explanation was the discrepancies in lesion sites between the two disease subtypes. The entire gastrointestinal tract can be affected in patients with CD while the lesion is localized to the colon and rectum in patients with UC. The pathways through which quercetin is absorbed, distributed, and metabolized in the digestive system and the bioactivities of different quercetin metabolites have been demonstrated before. The absorption of quercetin and related compounds partially (about 5 to 10%) is initiated from the small intestine, while most quercetin could be easily absorbed in the form of conjugates in the colon catabolized by the microflora and exert subsequent bioactivities.36,37,38–40 Accordingly, previous evidence showed quercetin could inhibit chemical-induced colorectal cancer but failed to reduce tumor incidence found in an APCmin/+ mouse model (a model with spontaneous tumor development mainly in the small intestine and also some in the colon).41
In subgroup analysis, we found that the association between quercetin and UC was statistically stronger in individuals with BMI < 30 kg m−2 than in those with a higher BMI. A previous cohort study in Danish found a similar weaker association between flavonoid intake and mortality among participants with BMI > 30 kg m−2.42 Microbiota was considered important to make quercetin and related metabolites bioactive.43 Reduction in the enrichment level of several probiotics (such as Bifidobacterium and Lactobacillus) was reported to decrease the bioactive transformation of quercetin.43 In fact, obesity will reshape the gut microbiota in humans,44 which also play driven roles in the inter-individual variability in absorption, metabolism, and excretion of polyphenols.45 However, there was no evidence to connect these clues and give biologically plausible explanations for the observed differences in the dietary quercetin intake–UC association across populations by BMI categories. The underlying mechanisms of the role of obesity in the associations still need more research.
In mediation analyses, we found that decreased CRP levels partially mediated (proportions of 7.15% and 2.74%) the protective association of dietary quercetin intake with IBD and UC. Subclinical inflammation is one of the most prominent features of the preclinical stage of IBD.46 Reversal of this pathologic process by decreasing the level of inflammation in the body is a reasonable assumption for the protective effect of quercetin against IBD. However, the low mediation proportion in our study may suggest an important role of other pathways in the effect of dietary quercetin, such as the contribution to intestinal barrier homeostasis found in our animal experiments. In addition, subsequent studies are needed to explore the mediating role of inflammation biomarkers other than CRP that are more closely associated with IBD, such as fecal calprotectin levels, in the quercetin–IBD association.
Previous studies have established mouse models and proven the protective role of quercetin in reducing colitis.5 These studies, however, varied much in quercetin dosage and feeding time. To match the prevention purpose, we only gave mice quercetin (generally matched to human protective dose) before colitis induction. Matched to the previous studies,5 we found prior quercetin feed was enough to significantly reduce the colitis severity, though it couldn't totally prevent the chemical-induced inflammation. Also, we found the significant downregulated colonic mRNA expression level of several recognized molecules that reflected gut inflammatory levels, in addition to serum cytokines that were reported in the previous studies.5 It is worth noticing that S100a8 and S100a9 are components of fecal calprotectin, a widely used clinical biomarker for evaluating gastro inflammation.47 Of note is that the expression of Cxcl9, which was found to be strongly related to the risk of a UC onset via our previous research,34 was also found to be induced upon colitis induction and reduced after quercetin treatment. As a detected potential target to prevent and ameliorate IBD, the inhibition of Cxcl9 expression could make sense to explain the potential mechanism of quercetin.34
We used 15 mg kg−1 quercetin in the in vivo study, equivalent to 84 mg d−1 (1.2 mg kg−1) for humans (weight 70 kg) based on the body surface area according to the US Food and Drug Administration.48 In the cohort study, we found that the protective association between quercetin and IBD risk remained significant when the quercetin intake peaked at around 80 mg (Fig. 2), which is comparable to the dosage in the animal study. Besides, though it was a little higher than the quercetin intake calculated in the human cohort, different quercetin forms (pure quercetin powder without conjugates was used in mouse models) and bioactivities in mice and humans should be considered.
To our knowledge, this is the first human study that investigated the association between quercetin intake and IBD risk. The strength of our study is the use of data from the large cohort UK Biobank, with a rich set of variables such as lifestyle, medication information, dietary intake, and a complete follow-up over time.
Nonetheless, our study also has several limitations. Firstly, although we included many potential confounders, we cannot rule out the possibility of residual confounding caused by unmeasured factors (e.g., use of prebiotics and probiotics) in an observational research design. Moreover, reverse causation that cannot be ruled out prevents us from proving the causality of our conclusions. However, we have obtained consistent results in analyses and experiments that demonstrate that our conclusions are not likely to be greatly affected by the above bias. For reverse causation, the lagged analysis excluding incident IBD cases in the first 2- and 4-year follow-up demonstrated that the delay in diagnosis of IBD was less likely to affect main conclusions. Results in animal experiments designed to match the design of our cohort study supported our primary findings. For potential confounding factors, we used multivariable adjustment and a propensity score to minimize the bias and obtain consistent results. Secondly, the quercetin we administered in the experiments was in the form of an aglycone while dietary quercetin is present mainly in O-glycosidic forms.49 This limitation should be noted when exploring the mechanisms of dietary quercetin effects on health in the future. Thirdly, although we used a validated 24 h WebQ, approximately 38% of the study sample had only 1 questionnaire data. We minimized the bias seen in self-reported diet information by excluding extreme energy intake and non-typical diets, as suggested by previous studies in the UK Biobank.12,50 Also, assessing the dietary quercetin intake using nutrient databases may introduce biases due to large variability in the polyphenol content of a given food and insufficient data on the composition of certain foods, though we have demonstrated the feasibility of the used databases in the European countries in the Methods section. In addition, the mean intake of dietary quercetin is 22 mg d−1 in our study, which is within the estimated intake (10–100 mg) by the previous review.51 In the Epic-Oxford study (∼30% vegetarians), the average intake of flavonoids was 54 mg d−1, of which quercetin accounted for 70% (i.e., 37.8 mg d−1).52 This may demonstrate the reliability of our estimate of the dietary quercetin intake to some extent. Finally, the vast majority of participants in our study were over 40 years of age and white, and caution is needed in generalizing our findings to other IBD populations.
In conclusion, higher quercetin intake was associated with a lower risk of IBD, especially UC. The protection role of quercetin against IBD was validated in two colitis mouse models. Together, our study showed the possibility of quercetin to prevent IBD in humans for the first time. These findings require further clinical interventional studies to validate and more in-depth experiments to elucidate the underlying mechanisms.
Author contributions
Shi-Yuan Lu: conceptualization (lead), methodology (equal), writing – original draft (lead), formal analysis (supporting), and validation (equal); Lintao Dan: formal analysis (lead) and writing – original draft (equal); Sishen Sun: methodology (supporting), validation (equal), and writing – review and editing (supporting); Tian Fu: methodology (supporting), writing – review and editing (supporting), and formal analysis (supporting); Jie Chen: conceptualization (equal), methodology (equal), writing – review and editing (lead), and project administration (lead). All authors have read and agreed to the published version of the manuscript.Ethical considerations
Ethical approval for the UK Biobank was granted by the North West-Haydock Research Ethics Committee (REC reference: 21/NW/0157). Written informed consent has been obtained from the patients to publish this study.Data availability
Researchers can require the data and approval from the UK Biobank (https://www.ukbiobank.ac.uk).Conflicts of interest
All authors declare that they have no competing interests.Acknowledgements
We acknowledge the National Undergraduate Training Program for Innovation and Entrepreneurship (grant number: 202310335170). This work was conducted using the UK Biobank Resource under application number 73595. We want to thank all UK Biobank participants and the management team for their participation and assistance. We also want to thank the researchers who contributed to Phenol-Explorer and the USDA database on flavonoids. All authors approved the final version of the article, including the authorship list.References
- G. G. Kaplan and J. W. Windsor, The four epidemiological stages in the global evolution of inflammatory bowel disease, Nat. Rev. Gastroenterol. Hepatol., 2021, 18, 56–66 CrossRef PubMed.
- T. Vezza, A. Rodríguez-Nogales, F. Algieri, M. P. Utrilla, M. E. Rodriguez-Cabezas and J. Galvez, Flavonoids in Inflammatory Bowel Disease: A Review, Nutrients, 2016, 8, 211 CrossRef PubMed.
- P. Shen, W. Lin, X. Deng, X. Ba, L. Han, Z. Chen, K. Qin, Y. Huang and S. Tu, Potential Implications of Quercetin in Autoimmune Diseases, Front. Immunol., 2021, 12, 689044 CrossRef CAS PubMed.
- F. Aghababaei and M. Hadidi, Recent Advances in Potential Health Benefits of Quercetin, Pharmaceuticals, 2023, 16(7), 1020 CrossRef CAS PubMed.
- S. Hu, M. Zhao, W. Li, P. Wei, Q. Liu, S. Chen, J. Zeng, X. Ma and J. Tang, Preclinical evidence for quercetin against inflammatory bowel disease: a meta-analysis and systematic review, Inflammopharmacology, 2022, 30, 2035–2050 CrossRef CAS PubMed.
- F. Javadi, A. Ahmadzadeh, S. Eghtesadi, N. Aryaeian, M. Zabihiyeganeh, A. Rahimi Foroushani and S. Jazayeri, The Effect of Quercetin on Inflammatory Factors and Clinical Symptoms in Women with Rheumatoid Arthritis: A Double-Blind, Randomized Controlled Trial, J. Am. Coll. Nutr., 2017, 36, 9–15 CrossRef CAS PubMed.
- D. Piovani, S. Danese, L. Peyrin-Biroulet, G. K. Nikolopoulos, T. Lytras and S. Bonovas, Environmental Risk Factors for Inflammatory Bowel Diseases: An Umbrella Review of Meta-analyses, Gastroenterology, 2019, 157, 647–659 CrossRef PubMed.
- A. Milajerdi, N. Ebrahimi-Daryani, L. A. Dieleman, B. Larijani and A. Esmaillzadeh, Association of Dietary Fiber, Fruit, and Vegetable Consumption with Risk of Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis, Adv. Nutr., 2021, 12, 735–743 CrossRef CAS PubMed.
- C. Sudlow, J. Gallacher, N. Allen, V. Beral, P. Burton, J. Danesh, P. Downey, P. Elliott, J. Green, M. Landray, B. Liu, P. Matthews, G. Ong, J. Pell, A. Silman, A. Young, T. Sprosen, T. Peakman and R. Collins, UK biobank: an open access resource for identifying the causes of a wide range of complex diseases of middle and old age, PLoS Med., 2015, 12, e1001779 CrossRef PubMed.
- L. J. Palmer, UK Biobank: bank on it, Lancet, 2007, 369, 1980–1982 CrossRef PubMed.
- W. Willett, Nutritional epidemiology, Oxford University Press, 2012 Search PubMed.
- H. Chen, T. Fu, L. Dan, X. Chen, Y. Sun, J. Chen, X. Wang and T. Hesketh, Meat consumption and all-cause mortality in 5763 patients with inflammatory bowel disease: A retrospective cohort study, EClinicalMedicine, 2022, 47, 101406 CrossRef PubMed.
- B. Rosner, Percentage Points for a Generalized ESD Many-Outlier Procedure, Technometrics, 1983, 25, 165–172 CrossRef.
- H. Crawley, Food portion sizes, 1988 Search PubMed.
- D. B. Haytowitz, X. Wu and S. Bhagwat, USDA database for the flavonoid content of selected foods, release 3.3, US Department of Agriculture, 2018, 173 Search PubMed.
- J. A. Rothwell, J. Perez-Jimenez, V. Neveu, A. Medina-Remon, N. M'hiri, P. García-Lobato, C. Manach, C. Knox, R. Eisner and D. S. Wishart, Phenol-Explorer 3.0: a major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content, Database, 2013, 2013, bat070 CrossRef PubMed.
- F. Lanuza, N. P. Bondonno, R. Zamora-Ros, A. L. Rostgaard-Hansen, A. Tjønneland, R. Landberg, J. Halkjær and C. Andres-Lacueva, Comparison of Flavonoid Intake Assessment Methods Using USDA and Phenol Explorer Databases: Subcohort Diet, Cancer and Health-Next Generations—MAX Study, Front. Nutr., 2022, 9, 873774 CrossRef PubMed.
- S. L. Anacleto, F. M. Lajolo and N. M. A. Hassimotto, Estimation of dietary flavonoid intake of the Brazilian population: A comparison between the USDA and Phenol-Explorer databases, J. Food Compos. Anal., 2019, 78, 1–8 CrossRef CAS.
- K. L. Ivey, K. Croft, R. L. Prince and J. M. Hodgson, Comparison of flavonoid intake assessment methods, Food Funct., 2016, 7, 3748–3759 RSC.
- B. Liu, H. Young, F. L. Crowe, V. S. Benson, E. A. Spencer, T. J. Key, P. N. Appleby and V. Beral, Development and evaluation of the Oxford WebQ, a low-cost, web-based method for assessment of previous 24 h dietary intakes in large-scale prospective studies, Public Health Nutr., 2011, 14, 1998–2005 CrossRef PubMed.
- K. E. Bradbury, H. J. Young, W. Guo and T. J. Key, Dietary assessment in UK Biobank: an evaluation of the performance of the touchscreen dietary questionnaire, J. Nutr. Sci., 2018, 7, e6 CrossRef PubMed.
- J. Barton, S. Gillon and A. Ferguson, Incidence of inflammatory bowel disease in Scottish children between 1968 and 1983; marginal fall in ulcerative colitis, three-fold rise in Crohn's disease, Gut, 1989, 30, 618–622 CrossRef CAS PubMed.
- J. D. Lewis, C. Brensinger, W. B. Bilker and B. L. Strom, Validity and completeness of the General Practice Research Database for studies of inflammatory bowel disease, Pharmacoepidemiol Drug Saf, 2002, 11, 211–218 CrossRef PubMed.
- Audit Commission for Local Authorities, the National Health Service in England and Wales, Improving data quality in the NHS: annual report on the PbR assurance programme, Audit Commission, 2010 Search PubMed , https://webarchive.nationalarchives.gov.uk/ukgwa/20150410163038/http://archive.audit-commission.gov.uk/auditcommission/aboutus/publications/pages/national-reports-and-studies-archive.aspx.html.
- M. Deng, L. Dan, S. Ye, X. Chen, T. Fu, X. Wang and J. Chen, Higher dietary fibre intake is associated with lower risk of inflammatory bowel disease: prospective cohort study, Aliment. Pharmacol. Ther., 2023, 58, 516–525 CrossRef CAS PubMed.
- D. M. Lloyd-Jones, Y. Hong, D. Labarthe, D. Mozaffarian, L. J. Appel, L. Van Horn, K. Greenlund, S. Daniels, G. Nichol, G. F. Tomaselli, D. K. Arnett, G. C. Fonarow, P. M. Ho, M. S. Lauer, F. A. Masoudi, R. M. Robertson, V. Roger, L. H. Schwamm, P. Sorlie, C. W. Yancy and W. D. Rosamond, Defining and setting national goals for cardiovascular health promotion and disease reduction: the American Heart Association's strategic Impact Goal through 2020 and beyond, Circulation, 2010, 121, 586–613 CrossRef PubMed.
- D. Blane, P. Townsend, P. Phillimore and A. Beattie, Health and Deprivation: Inequality and the North, Br. J. Sociol., 1987, 40, 344 CrossRef.
- H. Quan, V. Sundararajan, P. Halfon, A. Fong, B. Burnand, J. C. Luthi, L. D. Saunders, C. A. Beck, T. E. Feasby and W. A. Ghali, Coding algorithms for defining comorbidities in ICD-9-CM and ICD-10 administrative data, Med. Care, 2005, 43, 1130–1139 CrossRef PubMed.
- UK Biobank Biomarker Project-Companion Document to Accompany Serum Biomarker Data, 2019, https://biobank.ndph.ox.ac.uk/ukb/ukb/docs/serum_biochemistry.pdf, (accessed Mar 22, 2024).
- A. Perez-Cornago, Z. Pollard, H. Young, M. van Uden, C. Andrews, C. Piernas, T. J. Key, A. Mulligan and M. Lentjes, Description of the updated nutrition calculation of the Oxford WebQ questionnaire and comparison with the previous version among 207,144 participants in UK Biobank, Eur. J. Nutr., 2021, 60, 4019–4030 CrossRef CAS PubMed.
- J. Chen, J. Wellens, R. Kalla, T. Fu, M. Deng, H. Zhang, S. Yuan, X. Wang, E. Theodoratou, X. Li and J. Satsangi, Intake of Ultra-processed Foods Is Associated with an Increased Risk of Crohn's Disease: A Cross-sectional and Prospective Analysis of 187 154 Participants in the UK Biobank, J. Crohns Colitis, 2023, 17, 535–552 CrossRef PubMed.
- W. C. Willett, G. R. Howe and L. H. Kushi, Adjustment for total energy intake in epidemiologic studies, Am. J. Clin. Nutr., 1997, 65, S1220–S1228 CrossRef PubMed.
- Y. Sun, S. Yuan, X. Chen, J. Sun, R. Kalla, L. Yu, L. Wang, X. Zhou, X. Kong, T. Hesketh, G.-T. Ho, K. Ding, M. Dunlop, S. C. Larsson, J. Satsangi, J. Chen, X. Wang, X. Li, E. Theodoratou and E. L. Giovannucci, The Contribution of Genetic Risk and Lifestyle Factors in the Development of Adult-Onset Inflammatory Bowel Disease: A Prospective Cohort Study, Am. J. Gastroenterol., 2023, 118, 511–522 CrossRef CAS PubMed.
- J. Chen, Y. Zhou, Y. Sun, S. Yuan, R. Kalla, J. Sun, J. Zhao, L. Wang, X. Chen, X. Zhou, S. Dai, Y. Zhang, G. T. Ho, D. Xia, Q. Cao, Z. Liu, S. C. Larsson, X. Wang, K. Ding, J. Halfvarson, X. Li, E. Theodoratou and J. Satsangi, Bidirectional Mendelian Randomisation Analysis Provides Evidence for the Causal Involvement of Dysregulation of CXCL9, CCL11 and CASP8 in the Pathogenesis of Ulcerative Colitis, J. Crohns Colitis, 2023, 17, 777–785 CrossRef PubMed.
- R. Zamora-Ros, V. Knaze, L. Luján-Barroso, I. Romieu, A. Scalbert, N. Slimani, A. Hjartåker, D. Engeset, G. Skeie, K. Overvad, L. Bredsdorff, A. Tjønneland, J. Halkjær, T. J. Key, K. T. Khaw, A. A. Mulligan, A. Winkvist, I. Johansson, H. B. Bueno-de-Mesquita, P. H. Peeters, P. Wallström, U. Ericson, V. Pala, M. S. de Magistris, S. Polidoro, R. Tumino, A. Trichopoulou, V. Dilis, M. Katsoulis, J. M. Huerta, V. Martínez, M. J. Sánchez, E. Ardanaz, P. Amiano, B. Teucher, V. Grote, B. Bendinelli, H. Boeing, J. Förster, M. Touillaud, F. Perquier, G. Fagherazzi, V. Gallo, E. Riboli and C. A. González, Differences in dietary intakes, food sources and determinants of total flavonoids between Mediterranean and non-Mediterranean countries participating in the European Prospective Investigation into Cancer and Nutrition (EPIC) study, Br. J. Nutr., 2013, 109, 1498–1507 CrossRef CAS PubMed.
- D. Del Rio, A. Rodriguez-Mateos, J. P. Spencer, M. Tognolini, G. Borges and A. Crozier, Dietary (poly)phenolics in human health: structures, bioavailability, and evidence of protective effects against chronic diseases, Antioxid. Redox Signal., 2013, 18, 1818–1892 CrossRef CAS PubMed.
- A. Crozier, D. Del Rio and M. N. Clifford, Bioavailability of dietary flavonoids and phenolic compounds, Mol. Aspects Med., 2010, 31, 446–467 CrossRef CAS PubMed.
- V. A. Uyanga, F. K. Amevor, M. Liu, Z. Cui, X. Zhao and H. Lin, Potential Implications of Citrulline and Quercetin on Gut Functioning of Monogastric Animals and Humans: A Comprehensive Review, Nutrients, 2021, 13, 3782 CrossRef CAS PubMed.
- I. Erlund, T. Kosonen, G. Alfthan, J. Mäenpää, K. Perttunen, J. Kenraali, J. Parantainen and A. Aro, Pharmacokinetics of quercetin from quercetin aglycone and rutin in healthy volunteers, Eur. J. Clin. Pharmacol., 2000, 56, 545–553 CrossRef CAS PubMed.
- E. U. Graefe, H. Derendorf and M. Veit, Pharmacokinetics and bioavailability of the flavonol quercetin in humans, Int. J. Clin. Pharmacol. Ther., 1999, 37, 219–233 CAS.
- J. D. Lambert, J. Hong, G. Y. Yang, J. Liao and C. S. Yang, Inhibition of carcinogenesis by polyphenols: evidence from laboratory investigations, Am. J. Clin. Nutr., 2005, 81, 284s–291s CrossRef CAS PubMed.
- N. P. Bondonno, F. Dalgaard, C. Kyrø, K. Murray, C. P. Bondonno, J. R. Lewis, K. D. Croft, G. Gislason, A. Scalbert, A. Cassidy, A. Tjønneland, K. Overvad and J. M. Hodgson, Flavonoid intake is associated with lower mortality in the Danish Diet Cancer and Health Cohort, Nat. Commun., 2019, 10, 3651 CrossRef PubMed.
- U. Shabbir, M. Rubab, E. B. Daliri, R. Chelliah, A. Javed and D. H. Oh, Curcumin, Quercetin, Catechins and Metabolic Diseases: The Role of Gut Microbiota, Nutrients, 2021, 13, 206 CrossRef CAS PubMed.
- B. N. Liu, X. T. Liu, Z. H. Liang and J. H. Wang, Gut microbiota in obesity, World J. Gastroenterol., 2021, 27, 3837–3850 CrossRef CAS PubMed.
- R. P. Feliciano, C. E. Mills, G. Istas, C. Heiss and A. Rodriguez-Mateos, Absorption, Metabolism and Excretion of Cranberry (Poly)phenols in Humans: A Dose Response Study and Assessment of Inter-Individual Variability, Nutrients, 2017, 9, 268 CrossRef PubMed.
- J. J. Rudbaek, M. Agrawal, J. Torres, S. Mehandru, J.-F. Colombel and T. Jess, Deciphering the different phases of preclinical inflammatory bowel disease, Nat. Rev. Gastroenterol. Hepatol., 2024, 21, 86–100 CrossRef PubMed.
- N. Kapel, H. Ouni, N. A. Benahmed and L. Barbot-Trystram, Fecal Calprotectin for the Diagnosis and Management of Inflammatory Bowel Diseases, Clin. Transl. Gastroenterol., 2023, 14, e00617 CrossRef PubMed.
- M. Rockville, Guidance for Industry: Estimating the Maximum Safe Starting Dose in Adult Healthy Volunteer, USFDA, Silver Spring, MD, USA, 2005 Search PubMed.
- W. M. Dabeek and M. V. Marra, Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans, Nutrients, 2019, 11, 2288 CrossRef CAS PubMed.
- T. Fu, H. Chen, X. Chen, Y. Sun, Y. Xie, M. Deng, T. Hesketh, X. Wang and J. Chen, Sugar-sweetened beverages, artificially sweetened beverages and natural juices and risk of inflammatory bowel disease: a cohort study of 121,490 participants, Aliment. Pharmacol. Ther., 2022, 56, 1018–1029 CrossRef CAS PubMed.
- S. Egert and G. Rimbach, Which sources of flavonoids: complex diets or dietary supplements?, Adv. Nutr., 2011, 2, 8–14 CrossRef CAS PubMed.
- R. Zamora-Ros, V. Knaze, L. Luján-Barroso, N. Slimani, I. Romieu, V. Fedirko, M. S. De Magistris, U. Ericson, P. Amiano and A. Trichopoulou, Estimated dietary intakes of flavonols, flavanones and flavones in the European Prospective Investigation into Cancer and Nutrition (EPIC) 24 hours dietary recall cohort, Br. J. Nutr., 2011, 106, 1915–1925 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: https://doi.org/10.1039/d3fo05391a |
| ‡ These authors contributed equally. |
| This journal is © The Royal Society of Chemistry 2024 |