
 Open Access Article
Open Access Article This Open Access Article is licensed under a
This Open Access Article is licensed under a Creative Commons Attribution 3.0 Unported Licence
Valorization of byproducts from meat and dairy industries through fermentation to produce peptides
Chih-Chun
Kuo
*a,
Da
Chen
b,
Rafael
Jiménez-Flores
a,
Macdonald
Wick
c and
Osvaldo
Campanella
 *a
*a
aDepartment of Food Science and Technology, The Ohio State University, Columbus, 43210, OH, USA. E-mail: campanella.20@osu.edu
bDepartment of Food Science, Purdue University, West Lafayette, 47907, IN, USA
cDepartment of Animal Science, The Ohio State University, Columbus, 43210, OH, USA
First published on 5th July 2024
Abstract
The escalating global issue of waste streams, particularly within the food industry, necessitates a sustainable approach to valorizing food wastes and incorporating these valorized compounds into new products. This study addresses the limitations of existing protein extraction methods by proposing an innovative bioprocessing technology to effectively recover them from waste streams. The primary objective is to regulate protein hydrolysis through a fermentation procedure applied to waste streams from the meat and dairy industries. Sodium-citrated whole blood from cattle and pre-sterilized acid whey from cottage cheese production were blended, followed by the addition of Lactobacillus rhamnosus (OSU-PECh-69) due to its high proteolytic activity. The fermentation process, conducted at 37 °C for 5 days, revealed that L. rhamnosus maintained viability at ∼9 log CFU g−1, while coliforms remained below the detection limit of 250 CFU g−1. The acidity in the acid whey favored the growth of lactic acid bacteria over other pathogens, resulting in a decline in pH, which limited coliform growth. The fermentation mixture with the addition of L. rhamnosus achieved a degree of hydrolysis of 6%. SDS-PAGE analysis confirmed the modification of proteins into smaller fragments during fermentation. This biotechnological process demonstrates the potential to valorize nutrient-dense byproducts through fermentative hydrolysis, offering a promising avenue for creating economically viable and sustainable processing solutions to make better use of the food industry byproducts.
Sustainable spotlightDirect disposal of byproducts from the food industries is no longer sustainable and poses risks. By utilizing a fermentation process using bacteria, we aim to regulate protein hydrolysis in two waste streams. This approach not only may transform nutrient-dense byproducts into high-value ingredients but aligns with the UN's Sustainable Development Goals. It directly contributes to Goal 12 (Responsible Consumption and Production) by overcoming protein extraction limitations and encouraging sustainable processing solutions. Additionally, our approach supports Goal 9, associated to Industry, Innovation and Infrastructure by promoting innovation in waste management and sustainable practices. Overall, our work contributes to responsible food processing, promotes a circular economy in the food industry, and provides a long-term sustainable solution to food waste management challenges. |
Introduction
Waste management has been a major challenge for the food industry seeking a balance between economic value and environmental impact.1,2 Due to the increasing awareness of food security and sustainable food production, recovery of valuable components from food byproducts is highly recommended. As traditional extraction methods require the use of organic solvents and a high amount of water or heat, integrated bioprocessing technologies, often referred to as green extraction processes, are preferable as they are more sustainable.3 According to literature, ideal green solvents are expected to have the following properties: (1) low toxicity to humans and other organisms; (2) be easily biodegradable; (3) occur naturally; (4) be derived from renewable sources; (5) be produced as byproducts (6) have low vapor pressure, and (7) eliminate the need for traditional evaporation steps post-extraction.4 By utilizing byproducts from the food industry as a source of biomass, we can manage residual streams effectively without compromising food and feed availability. Consequently, the adoption of such green extraction techniques is crucial for sustainable bioprocessing, enabling the valorization of food industry byproducts in a manner that supports environmental and economic goals. Furthermore, no solvents are used in the current research, and extraction of important compounds can be obtained with the help of the fermentation process.The surge in global meat consumption, with approximately 40% of protein intake being of animal origin, poses significant waste management challenges to the meat industry.5 Approximately 50% of animal byproducts are deemed unsuitable for conventional consumption due to their physical and chemical properties. Moreover, animal byproducts are hardly consumed in the U.S. due to cultural reasons and public safety concerns. These byproducts generate tremendous disposal problems, and byproduct management to promote environmental protection and sustainability.6 Despite being byproducts, they are a great source of macro and micronutrients (e.g., protein, fat, iron, folic acid, zinc, vitamin A, B6, and B12).7 For example, blood, as an edible byproduct of the meat industry, possesses key characteristics that make it valuable. It is rich in essential nutrients, including proteins, vitamins, minerals, and amino acids, providing significant nutritional value.8 A common approach to recover their macro- and micronutrients is through the incorporation of animal byproducts into animal feed.
Acid whey is a byproduct of soft cheese and Greek yogurt production from the dairy industry. The market for such products has been growing rapidly in recent years. Because due to practical reasons, acid whey is underutilized and inexpensive, finding uses in other applications offers a practical and economic opportunity. Acid whey contains protein, calcium, and lactose and has a high concentration of Lactoccus (from the starter culture of the cheese-making process), which can degrade complex compounds (mainly proteins and lipids) and synthesize molecules such as exopolysaccharides.9 Venegas-Ortega et al. reported that lactic acid bacteria could be a feasible alternative to valorize food waste.10 Moreover, previous research has shown promising results in valorizing nutrient-rich byproducts from the dairy and fish industries through fermentations. This valorization has led to an increase in peptide concentrations as well as microbial diversity, offering a viable alternative for addressing the disposal issue of acid whey and fish waste.11
Valorization of byproducts by recovering key nutrients and incorporating them into new products has been considered a sustainable approach to overcome the world's growing human population and the massive production of unused byproducts. However, due to the limitations of the protein extraction methods, a new, innovative bioprocessing technology needs to be developed to efficiently extract these components. Several bioprocessing technologies have shown the effectiveness of recovering the nutritive value and the additional bioactivity from animal byproducts. Fermentation is one of the most common processing tools to increase the nutrition and functionality of animal byproducts.12 Research has shown that by fermentation, proteins can be modified to expose more polar groups on the surface for increased water solubility.11,13 The type of the bacteria culture strain starter can also affect the physicochemical characteristics and the functionality of proteins after fermentation. In addition, previous studies have pointed out that the lactic acid bacteria present in acid whey may preserve the same fermentation ability but not accelerate the proteolysis of meat byproducts.14,15 The competition of the natural microbiota within acid whey will strongly depend on the available nutrients and the type of byproduct source. In that matter, the low acidity of acid whey is favorable for the growth of lactic acid bacteria over other bacteria communities.
Recent advancements in bioprocessing technologies have further emphasized the potential of valorizing byproducts from food processes into high-added value compounds. As discussed previously, fermentation is a technology that can help recover valuable nutrients from various food-process waste streams. For example, different types of research have been done to comprehensively valorize by-products from seafood processing, enabling the recovery of valuable nutrients and enhancing their utilization.16 Lactic acid bacteria are commonly used for fermentation of fish products where their proteolytic activity aids the breakdown of proteins, enhancing their solubility and functional properties of the resulting products. Furthermore, lactobacillus fermentation ensures the safety and stability of the end products by pH reduction.17 The quality of antioxidant compounds, oil, and protein hydrolysates was shown to improve after LAB fermentation of fish byproducts.18
This study proposes a solution for waste management by leveraging fermentation to control protein hydrolysis in byproducts from the meat and dairy industries. We hypothesized that the fermentation of blood-acid whey with the addition of molasses and the starter culture L. rhamnosus would provide an optimal fermentation performance with higher LAB viable cell counts and degree of hydrolysis. The objective of this study was to define parameters for suitable fermentation parameters and characterize the fermentation kinetics of reactions of interest (microbial count, pH, degree of hydrolysis).
Materials and methods
Materials
Acid whey was donated from Superior Dairy; it is a byproduct of cottage cheese production. Cattle whole blood was provided by the USDA-inspected meat processing plant in the Department of Animal Sciences at The Ohio State University supplemented with sodium citrate solution (1% v/v) to avoid coagulation after and during collection. Blood was obtained from the animal during the exsanguination step in the slaughtering process. The animal was unconscious at the time of exsanguination and felt no pain. This process was done by trained staff in a USDA-FSIS inspected meat processing facility following all USDA-FSIS meat safety regulations and humane handling guidelines. Before fermentation, acid whey and blood were stored at −20 °C.Whole blood and pre-sterilized acid whey were mixed in a ratio of 1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :3 (w/w). Acid whey and blood were thawed and blended with unsulfured blackstrap molasses at a concentration of 2.5% w/w (Golden Barrel, Honey Brook, PA) as a carbohydrate source for supplemental purpose. The strain of Lactobacillus rhamnosus (OSU-PECh-69), Lr., was selected as the lactic acid bacteria (LAB) starter culture inoculated into the fermentation system due to its high proteolytic ability. This strain was originally isolated from provolone cheese and has shown to produce antimicrobial agents against both Gram-positive and Gram-negative bacteria.19,20
:3 (w/w). Acid whey and blood were thawed and blended with unsulfured blackstrap molasses at a concentration of 2.5% w/w (Golden Barrel, Honey Brook, PA) as a carbohydrate source for supplemental purpose. The strain of Lactobacillus rhamnosus (OSU-PECh-69), Lr., was selected as the lactic acid bacteria (LAB) starter culture inoculated into the fermentation system due to its high proteolytic ability. This strain was originally isolated from provolone cheese and has shown to produce antimicrobial agents against both Gram-positive and Gram-negative bacteria.19,20
Methodology of fermentation
The fermentation process was based on previous studies on acid whey fermentation of fish waste.11 Acid whey, whole blood, and molasses were mixed thoroughly. Four treatments were considered for this study: (1) whole blood + acid whey, (2) whole blood + acid whey + molasses, (3) whole blood + acid whey + Lr., and (4) whole blood + acid whey + molasses + Lr. (Table 1). The fermentation was conducted in a 1 liter fermenter with an S-shape air stopper to ensure ventilation and maintain the microaerobic condition for the growth of LAB for five days at 37 °C. Samples were collected from day 0 to day 5 and stored at −80 °C until analysis.| Treatment | Abbreviation | Composition | Blood (%) | Acid whey (%) | Molasses (%) | Lr. (CFU ml−1) |
|---|---|---|---|---|---|---|
| 1 | BW | Blood + whey | 25 | 75 | — | — |
| 2 | BWM | Blood + whey + molasses | 25 | 72.5 | 2.5 | — |
| 3 | BWL | Blood + whey + Lr. | 25 | 75 | — | 1 × 108 |
| 4 | BWML | Blood + whey + molasses + Lr. | 25 | 72.5 | 2.5 | 1 × 108 |
Inoculation of the starter culture
One loop of the starter culture of Lactobacillus rhamnosus (OSU-PECh-69) in a −80 °C glycerol stock was activated by inoculating it into 10 mL sterilized MRS broth medium (BD Difco™, Frankin Lakes, NJ) and then incubated aerobically for 16 h at 37 °C. One hundred (100) μl of the overnight culture was transferred to 10 mL of new sterilized MRS broth medium to reactivate the bacteria and incubated aerobically for 16 h at 37 °C. Cells were harvested by centrifugation (3500 × g, 20 min) and washed twice with 0.85% (w/v) NaCl. Afterward, the supernatant was removed, and cell pellets were collected and inoculated into the fish-whey mixture. The total amount of starter culture inoculation was 108 CFU mL−1.Monitoring of fermentation
The pH value of the samples was measured with a pH meter, whereas LAB microbial growth was monitored by the pour plate technique on DeMan, Rogosa, and Sharpe (MRS) agar with bromocresol green as a pH indicator. Total coliforms were monitored by plating the samples on eosin methylene blue agar. Plates were incubated at 37 °C for 48 h (LAB) and 24 h (total coliform) under aerobic conditions. The viability of the microorganisms was expressed as colony-forming units per gram of sample (CFU g−1).Molecular weight analysis: sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE)
The fermented samples were diluted to a protein concentration of ∼2 mg mL−1. An aliquot of diluents was added to the reduced-form Laemmli Buffer (with β-mercaptoethanol) at a ratio of 1:1 and heated for 5 min at 95 °C. A 10× Tris/glycine/SDS buffer was used as the electrophoresis gel buffer. 15 μL of samples and 10 μL of protein standard (10–250 kDa, Precision Plus Protein™) was loaded into a 12% hand-casted gel (TGX Stain-Free Fastcast Acrylamide Kit, 12%) and ran at 200 V for 30 min. The gel was then fixed with polypeptide fixative solution containing 40% methanol, 10% acetic acid, and 50% water (v/v) for 30 min, washed twice, stained in Coomassie Brilliant Blue G-250 for 1 h, destined with 10% acetic acid three times prior to being imaged using the GelDoc Go Imaging System (BioRad, Hercules, CA).
Degree of hydrolysis (DH)
DH was determined by the modified O-phthalaldehyde (OPA) method described by Nielsen et al.21 The total amount of amino acid in the fermentation mixture was measured by complete hydrolysis with 6 N HCl and heated at 120 °C for 24 h prior to analysis. Before the measurement of absorbance at 340 nm, the fermented samples or complete hydrolysate was mixed with OPA reagents at room temperature for 2 min. The DH was calculated from the following formula:
ODsample, ODtotal and ODblank are the absorbance of sample, complete hydrolysates, and water control, respectively.
Statistical analysis
All treatments were conducted in triplicate. Statistical analysis was performed by analysis of variance (ANOVA) with the post hoc Tukey's HSD significant difference test (α = 0.05) using the JMP Pro 16.0 software (SAS, Cary, NC).Results and discussion
Monitoring of fermentation
Fig. 1 illustrates the viable cell counts for LAB in the BW treatment on day 0 and the BWM treatment on Day 0 and 1 were below the detection limit means that the viability was less than 10 CFU g−1. During the BW treatment, the viability of LAB increased significantly after the first day of fermentation. Although the viability of LAB continued to increase in BW, it was found not to be significant from Day 2 to Day 5 (p > 0.05). A similar trend was found in BWM. In contrast, treatments with starter culture had a viability of LAB that maintained at ∼9 log CFU g−1 throughout the 5 day fermentation period (BWL and BWML), while no significant differences (p = 0.998) were found between BWL and BWML across the whole fermentation process.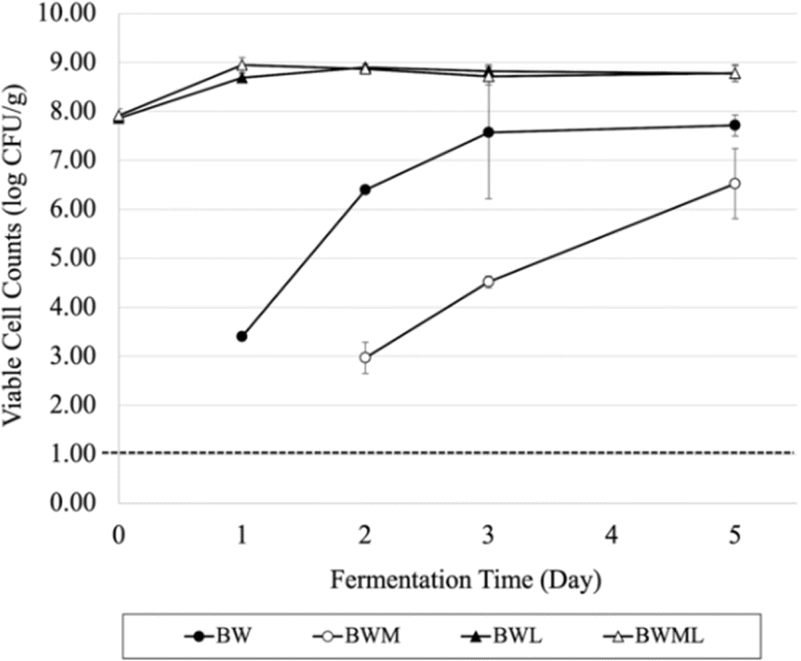 | ||
| Fig. 1 Viable cell counts of lactic acid bacteria in blood-whey (BW), blood-whey-molasses (BWM), blood-whey-Lr. (BWL), and blood-whey-molasses-Lr. (BWML) during the fermentation. Note: Dotted line represents the detection limit of 250 CFU g−1. | ||
When comparing within the same day across the four treatments, LAB viability was significantly the highest (p < 0.05) in the BWL and BWML treatments, followed by BW and BWM in Day 2 and 3. However, on day 5 of fermentation, LAB viability in BWL and BWML treatments were significantly higher than BWM (p < 0.01), but no significant difference was found between them. In conclusion, LAB counts in treatments inoculated with starter culture remained constant until the last day of fermentation. Even though an increase in LAB counts in BW and BWM was observed, the viability was still below those observed on samples with inoculation. Furthermore, the addition of molasses in treatments with starter culture did not make a significant difference in terms of LAB counts. Coliforms remained below the detection limit of 250 CFU g−1 during the five-day fermentation period for all treatments (not shown in Fig. 1).
Acid whey from cottage cheese has a pH of 4.09 ± 0.12 (mean ± SE) and increased to 4.14 ± 0.09 after the addition of molasses. Cattle blood, on the other hand, has a pH of 7.49 ± 0.11. The pH values for the four treatments across the 5 day fermentation period are presented in Fig. 2. The pH of treatments with starter culture (BWL and BWML) dropped significantly after the first day of fermentation (p < 0.05) and gradually decreased through the end of fermentation. The lowest pH was found to be 3.54 ± 0.16 in the BWL treatment and 3.65 ± 0.09 in the BWML treatment in day 5. However, no significant differences were found in these two groups at the end of the fermentation period, indicating that the addition of molasses did not have a significant difference. In contrast, the pH of the treatments without starter culture (BW and BWM) was maintained between 4.5 and 5 along the 5 day fermentation, and no significant difference was found between the BW and BWM treatment during the whole fermentation period. The low acidity of acid whey is favorable for the growth of lactic acid bacteria over pathogens. As the pH level continued to drop with fermentation time, the growth of coliforms was limited (below the detection limit of 250 CFU g−1).
 | ||
| Fig. 2 pH value of blood-whey (BW), blood-whey-molasses (BWM), blood-whey-Lr. (BWL), and blood-whey-molasses-lactic acid bacteria Lr. (BWML) during the fermentation period. | ||
The measured pH values of our fermentation materials, cattle blood and acid whey, are similar to those previously reported.22,23 The low pH in acid whey is primarily attributed to the presence of lactic acid.24 This low pH plays a key role in the microbial ecological dynamics during fermentation, which either foster or inhibit the growth of specific microorganisms. For lactic acid bacteria for example, the low pH environment acts as a facilitator for the growth of lactic acid bacteria (LAB) while concurrently serving as a deterrent for pathogenic organisms.25 As previously reported, LAB are known for thriving in acidic conditions.26,27 Its acidophilic nature allows LAB to dominate the microbial community during the fermentation period, contributing to the substantial pH drop noted in our study. The continued decline in pH throughout the fermentation period further supports the notion of LAB dominance, underscoring the effectiveness of low pH conditions in inhibiting the growth of potential pathogens, such as E. coli, that are sensitive to acidic environments.
Bovine blood is 80.9% water and 17.3% protein, with carbohydrates typically constituting less than 1%.28 Therefore, molasses was added into the formulations as an external source of sugar to facilitate the glycolytic metabolism of lactic acid bacteria in the fermentation systems. However, we did not observe any significant difference on the LAB viable cell counts and pH changes with the addition of molasses between the treatments with starter culture (BWL and BWML treatments). These results are in contrast to results reporting that the use of carbohydrate sources has a significant impact on the fermentation of fish waste with lactic acid bacteria.11,29,30 Although the carbohydrate concentration is low in blood, its main composition includes glucose which could be utilized directly as a source for hexose fermentation (glycolysis) in lactic acid bacteria.31 Our results show that the carbohydrate source present in blood and acid whey is sufficient to positively affect the LAB metabolism. Thus, the addition of molasses may be less important.
Molecular weight distribution: SDS-PAGE
SDS-PAGE results during the fermentation period are shown in Fig. 3. The low molecular weight bands (bracketed red arrow, Fig. 3d and e) that begins to show on the SDS-PAGE during fermentation could be either the results of hydrolysis or de novo synthesis (synthesis of complex compounds from simple molecules) caused by LAB. It should be noted that, both effects are possible. Peptides can either be generated from protein hydrolysis with the cleavage of peptide bonds or synthesis by two or more amino acids/peptides to form other peptides.32Fig. 3a shows the protein molecular weight distribution of acid whey (lane 2) and blood (lane 3) alone. The main proteins in acid whey are β-lactoglobulin and α-lactalbumin.33 Whereas albumin and hemoglobin are the two main proteins in whole blood.34,35 | ||
| Fig. 3 SDS-PAGE profile of (a) acid whey and blood alone, (b) blood-whey (BW), (c) blood-whey-molasses (BWM), (d) blood-whey-Lr. (BWL), and (e) blood-whey-molasses-Lr. (BWML) during the fermentation period. | ||
In Fig. 3d and e, the bands of large-molecule proteins (above 100 kDa and 66.5 kDa) hydrolyzed (proteolyzed) during fermentation. In addition, there was a large accumulation of smaller molecular weight bands from day 1 to day 5 in both treatments. Bands with molecular weights greater than 100 kDa are likely immunoglobulins (∼150 kDa) presented in both acid whey and blood; whereas bands around 66.5 kDa are likely albumins from the two sources.36,37 The lower molecular weight bands (below 15 kDa) consistently increase throughout the fermentation, while the large molecular weight bands (>100 kDa) disappear gradually, suggesting the occurrence of protein hydrolysis. However, the accumulation of lower molecular weight bands is less obvious in the fermentation conducted with no starter culture (Fig. 3b and c). Moreover, larger molecular weight bands (>100 kDa) remain unhydrolyzed during the fermentation period. This result suggests that the accumulation of lower molecular weight bands (<15 kDa), may not be the effect of protein hydrolysis but de novo synthesis, where low molecular weight bands form through peptides and amino acids synthesis.
For peptide synthesis, amino acids or small peptides are required as the material to synthesize peptides and this is why or how protein hydrolysis and peptide synthesis could happen at the same time via unidentified enzyme activity. Furthermore, amino acids and peptides generated from protein hydrolysis could serve as building material for peptide synthesis. As the proteins are breaking into small peptides or free amino acids due to hydrolysis, the band intensity on the same molecular weight should appear in a progressive way on the gel profile (intensity of bands with large molecular weight disappearing and intensity of small molecular weight bands accumulating). But if peptide synthesis is happening simultaneously during the hydrolysis, the bands of lower molecular weight (below 10 kDa) on the same row may not build up in a gradual way.
Therefore, both protein hydrolysis and peptide synthesis coexist during the fermentation process. As intact proteins are being hydrolyzed, new peptides may also be synthesized. The key to distinguishing between these processes on an SDS-PAGE gel is to monitor the time-dependent changes in band patterns, intensities, and the appearance of new bands.
Degree of hydrolysis
A degree of hydrolysis (6%) was achieved in the blood-whey fermentation treatment (BWL, BWML) with the addition of Lr. (Fig. 4). No significant difference was found between the BWML and BWL treatments (with and without the addition of molasses, respectively) in the ones inoculated with the starter culture. On the other hand, the degree of hydrolysis of the treatments with no starter culture (BW and BWM) remained below 2%. | ||
| Fig. 4 Degree of hydrolysis in blood-whey (BW), blood-whey-molasses (BWM), blood-whey-Lr. (BWL), and blood-whey-molasses-Lr. (BWML) during the fermentation period. | ||
Conclusions
This study introduces an innovative bioprocessing technology for the efficient extraction of components from waste streams in the meat and dairy industries. The fermentation process, utilizing Lactobacillus rhamnosus as a starter culture, demonstrated an effective regulation of protein hydrolysis in waste streams from the meat and dairy industries. The addition of molasses did not make a difference in the fermentation process. The low pH environment of acid whey favored the growth of lactic acid bacteria, limiting the proliferation of coliforms. SDS-PAGE analysis confirmed the hydrolysis of large-molecule proteins into smaller molecules during fermentation.This approach of mixed and complementary substrate fermentation holds significant promise in valorizing nutrient-dense byproducts through fermentation, offering sustainable and economically viable processing solutions for the food industry. This work could be applied to other biological systems to produce suitable and economically viable products in the food and feed industries, as well as create long-term sustainable processing solutions. Moreover, these peptides may have bioactivities such as antioxidant, antimicrobial, ACE (define)-inhibitory activities that could be incorporated into food and animal feed as an additive, which will bring out health-promoting benefits beyond basic nutrition. Our positive experience with the capabilities of the byproduct acid whey to ferment other organic matter, as well as the development of novel approaches to obtain valuable compounds, will be expanded to produce animal feed utilizing innovative and sustainable extrusion technology.
Data availability
Data can be obtained by request to the authors.Author contributions
Chih-Chun Kuo (data curation, investigation, writing – original draft), Da Chen (methodology, conceptualization, writing – review and editing), Rafael Jiménez-Flores (conceptualization, writing – review and editing), Macdonald Wick (conceptualization, writing – review and editing), Osvaldo Campanella (conceptualization, project administration, supervision, funding acquisition, writing – review and editing).Conflicts of interest
The authors declare no conflicts of interest related to this research.Acknowledgements
The acknowledgements come at the end of an article after the conclusions and before the notes and references. We gratefully acknowledge the support provided by the Carl E. Haas Endowment in Food Industries and the Department of Food Science and Technology at The Ohio State University, which played a pivotal role in the successful completion of this research. Special thanks are extended to Ethan Scheffler, the Meat Laboratory Manager, for their invaluable assistance in procuring cattle blood from the Department of Animal Sciences. Additionally, we express our sincere appreciation to Dr Rafael Jiménez-Flores and his laboratory team in the Department of Food Science and Technology for their generous provision of resources, contributing significantly to the advancement of this study. The collaborative efforts and support from these groups have been key in realizing our research objectives.References
- Z. Dou, C. Cochran, S. Finn, D. Galligan and N. Goldstein, Issue Paper-Council for Agricultural Science and Technology, 2018, vol. 62 Search PubMed.
- A. Seberini, in SHS Web of Conferences EDP Sciences, 2020, vol. 7403010 Search PubMed.
- F. J. Barba, E. R. Soto, M. Brncic and J. M. Rodriquez, Green Extraction and Valorization of By-Products from Food Processing, CRC Press, Boca Raton, 2019 Search PubMed.
- M. Herrero and E. Ibañez, J. Supercrit. Fluids, 2018, 134, 252–259 CrossRef CAS.
- P. Sans and P. Combris, Meat Sci., 2015, 109, 106–111 CrossRef CAS PubMed.
- W. Russ and R. Meyer-Pittroff, Crit. Rev. Food Sci. Nutr., 2004, 44, 57–62 CrossRef PubMed.
- H. K. Biesalski, Meat Sci., 2005, 70, 509–524 CrossRef PubMed.
- D. L. Marti, R. J. Johnson and Jr K. H. Mathews, J. Curr. Issues Glob., 2012, 5, 397 Search PubMed.
- L. D. Lasrado and A. K. Rai, Biosynthetic Technology and Environmental Challenges, 2018, pp. 185–203 Search PubMed.
- M. G. Venegas-Ortega, A. C. Flores-Gallegos, J. L. Martínez-Hernández, C. N. Aguilar and G. V. Nevárez-Moorillón, Compr. Rev. Food Sci. Food Saf., 2019, 18, 1039–1051 CrossRef CAS PubMed.
- A. C. Mayta-Apaza, I. García-Cano, K. Dabrowski and R. Jiménez-Flores, Microorganisms, 2021, 9, 100 CrossRef CAS PubMed.
- P. K. Sadh, S. Kumar, P. Chawla and J. S. Duhan, Molecules, 2018, 23, 2560 CrossRef PubMed.
- Y. Xiao, G. Xing, X. Rui, W. Li, X. Chen, M. Jiang and M. Dong, LWT--Food Sci. Technol., 2015, 63, 1317–1324 CrossRef CAS.
- F. Yang, W. S. Xia, X. W. Zhang, Y. S. Xu and Q. X. Jiang, Food Chem., 2016, 207, 86–92 CrossRef CAS PubMed.
- J. Kameník, Fermented Meat Products: Health Aspects, 2017, pp. 95–126 Search PubMed.
- S. A. Siddiqui, D. Lakshmikanth, C. Pradhan, Z. Farajinejad, R. Castro-Muñoz and A. Sasidharan, Crit. Rev. Food Sci. Nutr., 2023, 1–28 CrossRef PubMed.
- V. Venugopal, Front. Sustain. Food Syst., 2021, 5, 611835 CrossRef.
- F. J. Marti-Quijal, F. Remize, G. Meca, E. Ferrer, M. J. Ruiz and F. J. Barba, Curr. Opin. Food Sci., 2020, 31, 9–16 CrossRef.
- I. García-Cano, D. Rocha-Mendoza, J. Ortega-Anaya, K. Wang, E. Kosmerl and R. Jiménez-Flores, Appl. Microbiol. Biotechnol., 2019, 103, 5243–5257 CrossRef PubMed.
- I. García-Cano, W. E. Hussein, D. Rocha-Mendoza, A. E. Yousef and R. Jiménez-Flores, Microbiol. Resour. Announc., 2020, 9, 10–128 Search PubMed.
- P. M. Nielsen, D. Petersen and C. J. Dambmann, J. Food Sci., 2001, 66, 642–646 CrossRef CAS.
- B. Pawliński, M. Gołębiewski, M. Trela and O. Witkowska-Piłaszewicz, Sci. Rep., 2023, 13, 1414 CrossRef PubMed.
- L. Skudra, A. Blija, E. Sturmović, L. Dukaļska, A. Áboltiņš and D. Ḱ rkliņa, Acta Biotechnol., 1998, 18, 277–286 CrossRef CAS.
- J. Chandrapala, M. C. Duke, S. R. Gray, M. Weeks, M. Palmer and T. Vasiljevic, Separ. Purif. Technol., 2017, 172, 489–497 CrossRef CAS.
- T. Cai, O. C. Pancorbo, H. M. Barnhart, J. E. Sander and W. C. Merka, J. Appl. Poultry Res., 1994, 3, 49–60 CrossRef.
- L. Favaro and S. D. Todorov, Probiotics Antimicrob. Proteins, 2017, 9, 444–458 CrossRef CAS PubMed.
- A. Samaddar and A. Kaviraj, Int. J. Recycl. Org. Waste Agric., 2014, 3, 1–8 CrossRef.
- F. A. Alencar, thesis dissertation, Faculdade de Engenharia de Alimentos, Universidade Estadual de Campinas, 1983 Search PubMed.
- C. Bower, K. Hietala and T. Delaca, J. Food Process. Preserv., 2011, 35, 822–831 CrossRef CAS.
- A. Shabani, F. Boldaji, B. Dastar, T. Ghoorchi and S. Zerehdaran, J. Sci. Food Agric., 2018, 98, 4097–4103 CrossRef CAS PubMed.
- O. Kandler, Antonie Leeuwenhoek, 1983, 49, 209–224 CrossRef CAS PubMed.
- E. Oshimura and K. Sakamoto, Cosmet. Sci. Technol. Theor. Princ. Appl., 2017, 3, 285–303 Search PubMed.
- E. A. Pena-Ramos and Y. L. Xiong, J. Dairy Sci., 2001, 84, 2577–2583 CrossRef CAS PubMed.
- G. Sun and A. F. Palmer, J. Chromatogr. B, 2008, 86, 1–7 Search PubMed.
- S. Lee, S. Lee and K. B. Song, Food Chem., 2003, 82, 521–526 CrossRef CAS.
- G. Biosa, M. F. Addis, A. Tanca, S. Pisanu, T. Roggio, S. Uzzau and D. Pagnozzi, J. Proteonomics, 2011, 75, 93–99 CrossRef CAS PubMed.
- P. Lievore, D. R. Simões, K. M. Silva, N. L. Drunkler, A. C. Barana, A. Nogueira and I. M. Demiate, J. Food Sci. Technol., 2015, 52, 2083–2092 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2024 |