
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceAging-related gastrointestinal conditions decrease the bioaccessibility of plant sterols in enriched wholemeal rye bread: in vitro static digestion
Diego
Miedes
 ,
Mussa
Makran
,
Antonio
Cilla
*,
Reyes
Barberá
,
Guadalupe
Garcia-Llatas
and
Amparo
Alegría
,
Mussa
Makran
,
Antonio
Cilla
*,
Reyes
Barberá
,
Guadalupe
Garcia-Llatas
and
Amparo
Alegría
Nutrition and Food Science Area, Faculty of Pharmacy, University of Valencia, Av. Vicent Andrés Estellés s/n, 46100 Burjassot, Valencia, Spain. E-mail: Antonio.cilla@uv.es
First published on 18th May 2023
Abstract
The prevention of cardiovascular disease using foods fortified with plant sterols (PS), with a hypocholesterolemic effect, is important for the elderly population. This study aimed at identifying the different PS present in PS-enriched wholemeal rye bread (WRB) and in the ingredient source of PS, to evaluate their bioaccessibility in WRB by simulated static digestion. The gastrointestinal conditions of the elderly were adapted, and the results were compared with the adult population. Nine PS were identified, and a total amount of 2.18 g/100 g WRB was determined. Bioaccessibility was reduced in the elderly model with gastrointestinal adaptation vs. the adult model (11.2 vs. 20.3%), but no differences were observed when adapting only the gastric phase. Even though there was lower bioaccessibility of PS in the elderly, they could benefit from the consumption of WRB as it has a good nutritional profile. Further investigation including in vivo assays is needed to strengthen the results.
1. Introduction
In 2019, the elderly population (people over 65 years old) was 703 million persons, but it is expected to double to 1.5 billion in 2050.1 This fact implies that a greater effort must be devoted to preventing chronic non-communicable diseases, which the elderly population has a higher risk of suffering from. Cardiovascular diseases stand out among these pathologies, being the world's biggest cause of death (up to 27% of total deaths in 2019).2 One of the most important risk factors for these diseases is hypercholesterolemia.The intake of 1.5 to 3 g day−1 of plant sterols (PS) has a hypocholesterolemic effect. It can reach between 7 and 12.5% reduction in plasma cholesterol in yellow fat spreads, dairy products, mayonnaise and salad dressings.3 Since it is very difficult to achieve this daily intake with the usual diet, fortification of foods with these compounds may be promising. Among the foods for which the legislation allows their addition is rye bread.4 Wholemeal rye bread stands out for being rich in fiber. Compared to wheat bread, it contains up to twice as much β-glucan, which has been shown to have a hypocholesterolemic effect that could complement that provided by PS. This effect seems to be independent of the origin of the β-glucan.5 Also, arabinoxylans, the main component of rye fiber, have shown a hypocholesterolemic effect due to their capacity to increase the viscosity of the food bolus. In fact, fibers with high viscosity, together with their bile salt sequestration activity, could have the same effect. Therefore, rye fiber as a whole product could have a greater hypocholesterolemic effect,6 being a beneficial matrix for the dietary prevention of cardiovascular diseases. For this reason, it could be an alternative in the diet to traditionally consumed white wheat bread.
Aging leads to distinct changes in the functioning of the gastrointestinal tract. There is a decrease in saliva secretion and a worsening of chewing force.7,8 An increased prevalence of hypochlorhydria due to atrophic gastritis is observed in elders. Furthermore, there is a reduction in gastric and intestinal enzyme and bile salt secretion and a general slowing down of digestion.8–11 These changes are expected to influence the digestibility of foods, their different nutrients, and bioactive compounds such as PS, so when adapting simulated digestion methods, they should be considered.
In vitro static digestion assays are economical in terms of time and money and simple to develop. They serve as a preliminary test for more expensive studies such as dynamic digestion trials or in vivo studies and therefore allow the screening of a huge number of matrices and parameters. The INFOGEST method was created in consensus to standardize in vitro static assays and allow comparison between them.12 The method has been subsequently updated,13 including the use of gastric lipase (GL).
To date, numerous in vitro studies have been carried out for determining the differences between the elderly and adult digestion conditions in various matrices. Most of them have evaluated proteolysis but the bioaccessibility of the other macronutrients and some micronutrients has also been studied.14
Cholesterol bioaccessibility has been evaluated under adult conditions with a multi-compartmental model in a fiber-enriched yogurt with egg yolk and sunflower oil15 and with an INFOGEST digestion assay in fiber-enriched pork patties.16 Later, Makran et al.17 introduced the joint use of GL and cholesterol esterase (CE) to make the digestion of PS more physiological. This study was taken as the basis for evaluating, for the first time, the bioaccessibility of PS by our research group in a PS-enriched milk-based fruit beverage under elderly adapted conditions.18 But the influence of a solid matrix in the bioaccessibility of PS is hitherto unknown and could be affected in a different way by the oral and gastrointestinal conditions of the elderly. GL is more effective in solid foods than in liquids,19 which could change the previously observed pattern of lipolysis and micellarization of sterols. Among the factors that stand out for being able to cause differences between this matrix and a liquid one are chewing and the presence of dietary fiber. The latter can interfere not only with the PS themselves,15,16 but also with the different enzymes and bile salts.
The main objective of this study is to evaluate the bioaccessibility of PS in 100% wholemeal rye bread under INFOGEST 2.0 adapted conditions to the elderly, comparing it with the standard conditions of adults, since PS-enriched foods constitute an adequate dietary treatment for moderate hypercholesterolemia. Additionally, determining the possible influence of a solid and fiber-rich matrix is an extra aim of the research.
2. Materials and methods
2.1 Reagents
5-Cholesten-3β-ol (cholesterol) (purity 99%), 5,22-cholestadien-24-ethyl-3β-ol (stigmasterol) (purity 97%), 24α-ethyl-5α-cholestan-3β-ol (sitostanol) (purity 97%) and the internal standard (IS) 5β-cholestan-3α-ol (epicoprostanol) (purity 99%) were purchased from Merck LifeScience S.L.U. (Madrid, Spain). 24α-Methyl-5-cholesten-3β-ol (campesterol) (purity 98%) and 5-cholesten-24β-ethyl-3β-ol (β-sitosterol) (purity 98%) were supplied from Chengdu Biopurify Phytochemicals Ltd (Sichuan, China).Trimethylchlorosilane (TMCS) (≥98%, v/v) and anhydrous pyridine (≥99.5%, v/v) obtained from Acros Organics (Geel, Belgium) and hexamethyldisilazane (HMDS) (≥99.9%, v/v) obtained from Merck LifeScience S.L.U. (Madrid, Spain) were used as derivatization reagents. α-Amylase from human saliva (E.C 3.2.1.1), ammonium carbonate, ammonium chloride, anhydrous sodium sulfate, bovine bile salts, butylhydroxytoluene, calcium chloride dihydrate, CE from porcine pancreas (E.C 3.1.1.13), hydrochloric acid (purity 37% w/w), magnesium chloride hexahydrate, pancreatin from porcine pancreas, pepsin from porcine gastric mucosa (E.C 3.4.23.1), potassium chloride, potassium dihydrogen phosphate, potassium hydroxide, sodium chloride and sodium hydroxide were purchased from Merck LifeScience S.L.U. (Madrid, Spain). Rabbit gastric extract (RGE15, 15 U lipase per mg powder) was acquired from Lipolytech (Marseille, France). Absolute ethanol (≥99.8%, v/v), sodium bicarbonate and sodium hydroxide were purchased from Panreac (Barcelona, Spain). Cyclohexane (≥99.9%, v/v), diethyl ether (≥99.5%, v/v) and n-hexane (≥96%, v/v) were obtained from Scharlau (Barcelona, Spain). Water was purified using a Milli-Q system (Millford, MA, United States).
2.2 Sample preparation
The sample used was 100% wholemeal rye bread enriched with PS (WRB). The bread was made with the following proportions (w/w) of ingredients in the bread dough: whole rye flour (57%), compressed yeast (1.4%), common salt (0.9%), ultrapure water (38.2%), ascorbic acid (0.006%) and microencapsulated free PS source powder ingredient from tall oil (2.5%) (Lypophytol® ME dispersable palm free, Lipofoods, Barcelona, Spain).The WRB manufacturing process began by mixing all the ingredients in a mixer with rotating blades. Immediately afterwards, the bread dough was left to rest for 10 minutes and divided into individual portions of 102.5 g. These were manually balled and left to rest for another 15 minutes. Subsequently, the dough was fermented at a temperature of 28 °C with a relative humidity of 85% for 45 minutes. Once fermented, the bread doughs were baked for 25 minutes at a temperature of 180 °C. Four individual loaves with an average weight of 81.32 ± 1.09 g were obtained from each baking process. With the aim to evaluate the influence of the manufacturing process in the PS content, experiments with WRB from different manufacturers (baking 1, baking 2 and baking 3) were carried out with the same batch of flours under identical baking conditions and at three different days. To obtain a more homogeneous and stable sample, fresh WRB was partially dehydrated at 24 °C overnight until the humidity was below 14% (w/w) and further homogenized with a sample homogenizer until a breadcrumb texture was obtained (this is the partially dried bread (PDB)).
The nutritional information (g/100 g WRB) was determined experimentally by official methods: water 32.7 ± 1.0; lipids 3.19 ± 0.1; carbohydrates 43.9 ± 0.4; soluble fiber, 3.4 ± 0.1; insoluble fiber, 10.3 ± 1.4; proteins, 5.16 ± 0.03; ash, 1.37 ± 0.04.
2.3 Simulated gastrointestinal digestion
Simulated salivary fluid (SSF), simulated gastric fluid (SGF) and simulated intestinal fluid (SIF) were prepared according to the INFOGEST protocol.12 Enzymatic activity and bile salt content were analyzed following the conditions of this protocol and collected in a previous study of our research group.17 CE is not contemplated in the INFOGEST method as there is no standardized methodology for determining its activity. Thus, the one given by the manufacturer for the batch (34.8 U mg−1 powder) was used.The healthy adult control conditions were performed following the INFOGEST 2.0 standardized methodology proposed by Brodkorb et al.13 with the inclusion of CE17 to make it more physiological. Changes in the elderly conditions E1 and E2 have been selected considering a recent work by our group,18 based on the pathophysiological changes that occur with aging and in the previous elderly adapted static in vitro digestion trials that also analyzed lipolysis and lipophilic compounds20 since, to date, there is no standardization to apply the conditions of the elderly in simulated digestion. The simulated digestion process and the differences between the conditions of the adult and the elderly are detailed in Fig. 1. For the in vitro oral phase, 3.7 g of PDB were mixed with water until reaching 26.2% humidity (from fresh bread) and 3.5 mL of SSF. The mixture was shaken for one min in a sample homogenizer (Masticator Basic 400, IUL, Spain). As a blank of digestion, the same amount of water (5 g) was used to substitute the fresh WRB sample. In Elderly 1 conditions (E1), the gastric phase was adapted by reducing the activity of pepsin and lipase to 75% and 15%, respectively, reducing agitation and increasing the pH and the duration of this phase (see conditions in Fig. 1). In Elderly 2 conditions (E2), E1 changes in the gastric phase were maintained and some additional changes were made in the intestinal phase: reduction of both pancreatin activity and bile salts concentration by 50%, reduction of agitation and increase in the duration (see conditions in Fig. 1). Preliminary digestion tests were carried out by applying an in vivo oral phase to fresh WRB with human mastication. A healthy adult volunteer (41 years, amylase activity: 245.3 ± 16.9 U mL−1 saliva) was used and mastication cycles were reduced with respect to the adult model (20 vs. 40).21
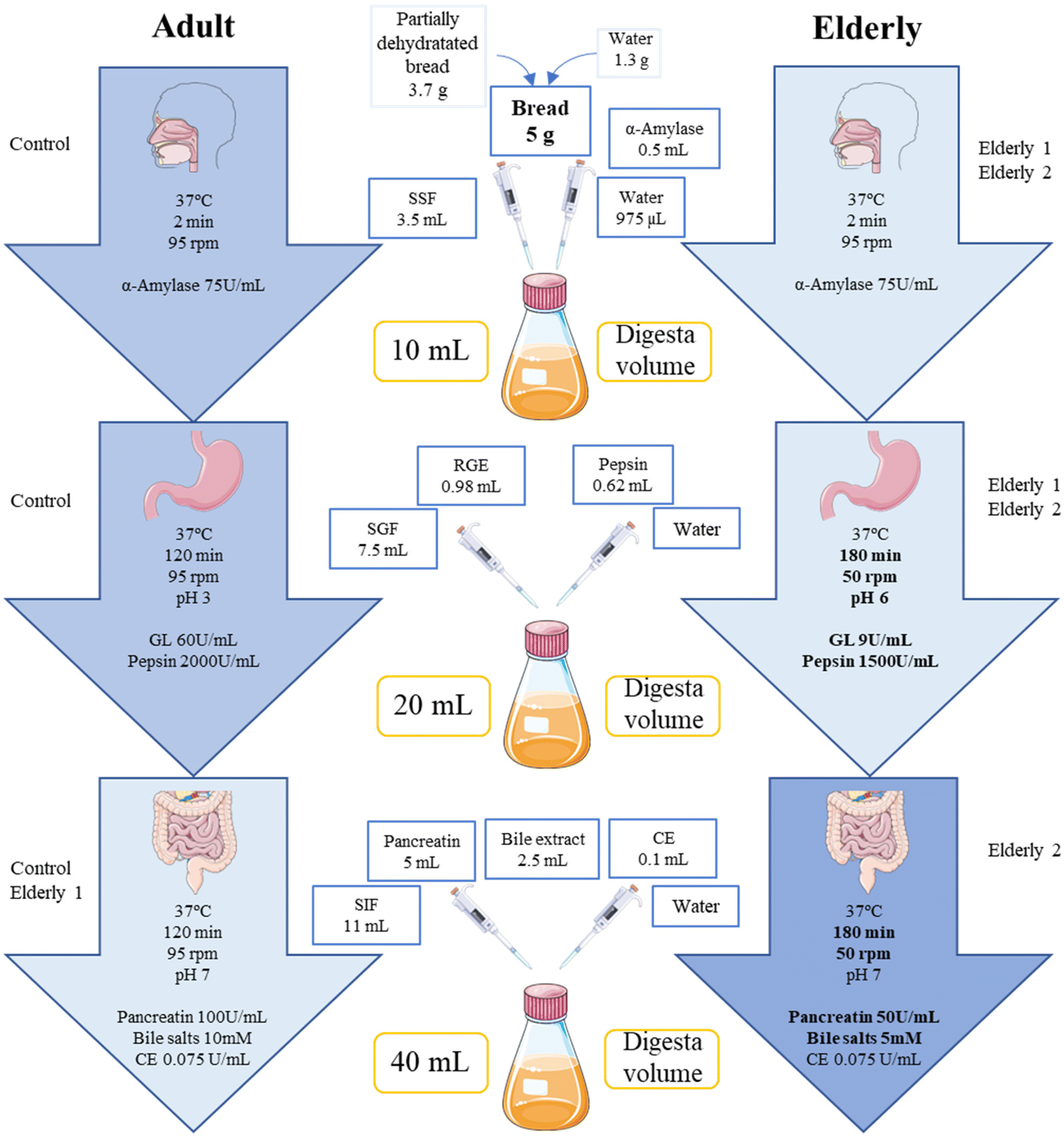 | ||
| Fig. 1 Summary of the simulated gastrointestinal digestion process on partially dried plant sterol-enriched wholemeal rye bread under specific conditions adapted for adults (extracted from ref. 17) and the elderly (extracted from ref. 18). Graphics also shows conditions used for each test (control, elderly 1 and elderly 2). SSF: simulated salivary fluid; SGF: simulated gastric fluid; SIF: simulated intestinal fluid; GL: gastric lipase; RGE: rabbit gastric extract; CE: cholesterol esterase. | ||
The digesta obtained (from control, E1 or E2) was centrifuged for 90 min at 4 °C and 3100 g (Centrifuge 5810 R, Eppendorf AG, Germany) and the supernatant corresponding to the bioaccessible fraction (BF) was collected. The formula [(sterol content in BF/sterol content in WRB) × 100] was used to calculate the bioaccessibility of total and individual sterols. In addition, the contents of the digestion blank were subtracted from the samples to obtain the actual PS content provided by the WRB.
2.4 Sterol determination
Acid hydrolysis and saponification were carried out following the method proposed by Piironen, Toivo & Lampi.22 The IS (epicoprostanol) (1 mg), and 1 mL of ethanol were added to partially dried WRB (0.35 g), or PS-ingredient sample (20 mg), both weighed in a 30 mL test tube at the beginning of the determination. For acid hydrolysis, 5 mL of 6M HCl were added, and the tube contents were mixed for 30 s with a test tube mixer (Vortex MS2, Janke and Kunkel, Ika Labortechnik, Germany), before the tube was placed into a shaking water bath (SBS40, Cole-Parmer, United Kingdom) (80 °C) for 60 min. After the tube was cooled, lipids were extracted with 20 mL of an n-hexane and diethyl ether (1![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1) solvent mixture by shaking for 10 min at 400 rpm using a sample tube rocker (KS 260 Basic, Ika, Germany). Then, it was centrifuged for 10 minutes at 500 rpm and the organic layer was transferred to a round-bottom flask and evaporated to dryness using a rotary evaporator (B490, BÜCHI Labortechnik, Switzerland) at 50 °C, and then resuspended in 8 mL of absolute ethanol.
:1) solvent mixture by shaking for 10 min at 400 rpm using a sample tube rocker (KS 260 Basic, Ika, Germany). Then, it was centrifuged for 10 minutes at 500 rpm and the organic layer was transferred to a round-bottom flask and evaporated to dryness using a rotary evaporator (B490, BÜCHI Labortechnik, Switzerland) at 50 °C, and then resuspended in 8 mL of absolute ethanol.
The determination of BF, since acid hydrolysis was not carried out, was started directly by adding 2 g of BF, (weighed into a round-bottom flask), and then 0.2 mg of IS and 6 mL of absolute ethanol were added and transferred to a 50 mL test tube. Both BF and the previously hydrolyzed samples (PS ingredient and WRB) were saponified with 0.5 mL of saturated aqueous KOH solution. The tube contents were mixed (10 s) using the test tube mixer and they were placed in a shaking water bath (80 °C) for 30 min (100 rpm) and then cooled. For extraction of the unsaponifiable fraction, 12 mL of water and 20 mL of cyclohexane were added, followed by shaking at 400 rpm for 10 min using the sample tube rocker. The organic layer was transferred to a round-bottom flask and evaporated to dryness in a rotary evaporator at 50 °C. The residue was dissolved in 1 mL of hexane. A silica cartridge (Finisterre SPE tube Si 500 mg/6 mL, Teknokroma, Spain) was activated by 5 mL of hexane. After filtering and applying the sample as a hexane extract, the cartridge was washed with 5 mL of hexane and 5 mL of a hexane and diethyl ether (90:10) solvent. The sterol fraction was eluted with 5 mL of a hexane and diethyl ether (50:50) solvent and evaporated to dryness. In WRB and sterol ingredient, the sample was transferred to a 5 mL volumetric flask, from which 1 mL is taken for derivatization. In the BF, the sample was not diluted, and the derivatization proceeded directly.
The derivatization was made following Alvarez-Sala et al.23 The trimethylsilylether derivatives, prepared from 600 μL of pyridine:HMDS:TMCS (5:2:1, v/v/v) (25 min, 40 °C), were dissolved in hexane after the evaporation of the reagent, filtered through a 0.45 μm filter (syringe driven Millex FH with filter 1 mL Millipore, Milford, MA, United States) and dissolved in 100 μL of n-hexane. 1 μL of the above solution was injected in a GC-FID (YL Instrument 6500 GC System, Gyeonggi-do, Korea). A capillary column CP-Sil 8 low bleed/MS (50 m × 0.25 mm × 0.25 μm film thickness) (Chrompack-Varian, Middelburg, The Netherlands) and hydrogen as the carrier gas (2 mL min−1) were used. The oven temperature was increased from the initial 280 °C (held 20 min) to 290 °C (held 5 min) at a rate of 0.7 °C min−1 and then increased to 320 °C (5 min) °C at a rate of 30 °C min−1. The temperature of both the injector port and the detector was 325 °C, and a split ratio of 1:20 was used.
Sterols were identified by comparing their relative retention times with those of the pure standards. Sterols without commercialized standard (campestanol, Δ5-avenasterol, Δ5,24(25)-stigmastadienol, Δ7-stigmastenol and Δ7-avenasterol) have been identified by mass spectrometry, comparing the main ions with the ones provided by the bibliography22,24,25 and mass spectra library,26 and a single sterol was assigned for each relative retention time. Calibration curves containing 200 μg of IS and increasing amounts of sterol standards were used for quantification. Therefore, due to the mentioned the lack of standard for them, the β-sitosterol calibration curve was used for their determination.
2.5 Statistical analysis
Sterol characterization in WRB and PS-ingredient was carried out in triplicate. All assays of sterol bioaccessibility were determined in two independent experiments with three replicates for each experiment (n = 6 replicates). Shapiro–Wilk and levene tests were used to evaluate normality and homoscedasticity, respectively. Then, to determine statistically significant differences (p < 0.05) between different baking or gastrointestinal conditions for the total and individual PS contents in WRB, PS-ingredient, BF or bioaccessibility, one-way analysis of variance (ANOVA), followed by Tukey's post hoc test, was used. The program used for statistical analysis was Graphpad Prism 8.0.2 (GraphPad Software Inc., San Diego, CA, USA).3. Results and discussion
3.1 PS in PS-enriched wholemeal rye bread
PS identified in the ingredient, WRB and their bioaccessible fractions (BF) are shown in Table 1, which includes the retention time, relative retention time and characteristic ions of each sterol identified by GC/MS. In a total analysis time of 30 min, seven phytosterols (campesterol, stigmasterol, β-sitosterol, Δ5-avenasterol, Δ5,24(25)-stigmastadienol, Δ7-stigmastenol and Δ7-avenasterol) and two phytostanols (campestanol and sitostanol) were identified (Fig. 2A–C). After IS (epicoprostanol), cholesterol and cholestanol were detected only in BF. This fact is explained by the contributions of these sterols by porcine pancreatin, bovine bile and gastric lipase used during digestion. This was estimated in a previous work, without significant contribution of PS.17 Besides, other peaks eluted (Fig. 2) corresponding to compounds with steroidal structures, which have not been identified as PS. | ||
| Fig. 2 Chromatograms of sterols identified in plant sterol (PS)-enriched wholemeal rye bread and its bioaccessible fractions. (A) PS ingredient; (B) PS-enriched 100% wholemeal rye bread; (C) bioaccessible fraction from elderly gastrointestinal adapted in vitro digestion. 1. Epicoprostanol (internal standard); 2. campesterol; 3. campestanol; 4. stigmasterol; 5. β-sitosterol; 6. sitostanol; 7. Δ5-avenasterol; 8. Δ5,24-stigmastadienol; 9. Δ7-stigmastenol; 10. Δ7-avenasterol; a. cholesterol; b. cholestanol; *. steroid structures. | ||
| Sterols | Rt (min) | RRt (min) | Characteristic ions | Ref. | |
|---|---|---|---|---|---|
| Rt and RRt values were expressed as media ± standard deviation of 4 independent assays by triplicate (n = 12) and 2 independent assays by triplicate (n = 6) in wholemeal rye bread and their bioaccessible fractions, respectively. IS: internal standard. | |||||
| 1 | Epicoprostanol (IS) | 15.454 ± 0.487 | 445.4, 370.4, 355.3, 316.3, 257.2, 215.2,161.1 | Mass spectra of sterol standard and Mass spectra library.25 | |
| 2 | Campesterol | 22.442 ± 0.700 | 1.452 ± 0.001 | 472.4, 430.2, 382.4, 343.3, 289.2, 255.2 | |
| 3 | Campestanol | 22.882 ± 0.707 | 1.481 ± 0.002 | 474.9, 459.4, 417.3, 373.6, 369.3, 343.3, 306.2, 215.2 | |
| 4 | Stigmasterol | 23.689 ± 0.712 | 1.533 ± 0.003 | 484.5, 469.7, 394.3, 379.6, 355, 295.5, 255.2, 213.4 | |
| 5 | β-Sitosterol | 26.484 ± 0.834 | 1.714 ± 0.004 | 486.4, 471.4, 396.4, 357.4, 275.3, 255.2, 213.2 | |
| 6 | Sitostanol | 26.849 ± 0.806 | 1.738 ± 0.004 | 488.3, 473.4, 431.4, 383.4, 305.2, 215.2 | |
| 7 | Δ5-Avenasterol | 27.023 ± 0.785 | 1.749 ± 0.005 | 484.4, 473.4, 431.4, 386, 296.0, 357.3, 55.0 | 21, 23, 24 and 37 |
| 8 | Δ5,24(25)-Stigmastadienol | 28.206 ± 0.785 | 1.825 ± 0.007 | 484.4, 469.3, 386.2, 345, 296.3, 257.0, 55.0 | 23, 24 and 37 |
| 9 | Δ7-Stigmastenol | 28.987 ± 0.799 | 1.876 ± 0.008 | 486.4, 471.4, 396, 381.3, 303.2, 255.2, 213.2 | 23, 24 and 37 |
| 10 | Δ7-Avenasterol | 29.758 ± 0.793 | 1.92± 0.010 | The literature indicates that it elutes after Δ7-stigmastenol | 21, 23 and 24 |
PS contents in PS-ingredient and WRB prepared under different baking days are tabulated in Table 2. The total PS content ranged between 2.17 and 2.23 g/100 g of fresh rye bread under various baking conditions (mean 2.18 ± 0.09), including the sum of free and covalently bound sterols (steryl esters with fatty acids, or phenolic acids, steryl glucosides, and acylated steryl glycosides) due to acid hydrolysis applied in the analysis that breaks all bonds of conjugated sterols.27 This content is significantly higher than that indicated by other authors for rye bread containing wheat flour (80.3 mg/100 g) and rye bread (90.2 mg/100 g).22 This was expected, given that the bread has been enriched with an ingredient source of PS with an experimentally determined purity of 74.65%.
| PS | Baking 1 | Baking 2 | Baking 3 | Mean | PS-ingredient |
|---|---|---|---|---|---|
| Contents were expressed as media ± standard deviation (n = 3). No statistically significant differences were observed in the individual nor in the total PS content in different baking. | |||||
| Campesterol | 156.08 ± 7.53 (7.18) | 160.95 ± 6.12 (7.21) | 151.58 ± 6.86 (7.09) | 156.20 ± 7.20 (7.16) | 5.55 ± 0.39 (7.43) |
| Campestanol | 21.23 ± 2.03 (0.98) | 24.32 ± 0.16 (1.09) | 24.45 ± 1.78 (1.14) | 23.34 ± 2.08 (1.1) | 1.02 ± 0.06 (1.37) |
| Stigmasterol | 8.79 ± 0.51 (0.40) | 9.70 ± 0.21 (0.43) | 9.38 ± 0.66 (0.44) | 9.29 ± 0.59 (0.43) | 0.39 ± 0.02 (0.52) |
| β-Sitosterol | 1720.54 ± 9.88 (79.18) | 1762.84 ± 56.18 (79.01) | 1685.23 ± 79.68 (78.86) | 1722.87 ± 72.91 (79.02) | 58.82 ± 3.84 (78.79) |
| Sitostanol | 225.65 ± 9.88 (10.38) | 232.44 ± 8.82 (10.42) | 223.75 ± 11.02 (10.47) | 227.28 ± 9.48 (10.43) | 7.62 ± 0.41 (10.21) |
| Δ5-Avenasterol | 15.00 ± 0.86 (0.69) | 16.23 ± 0.99 (0.73) | 14.57 ± 0.94 (0.68) | 15.26 ± 1.10 (0.70) | 0.52 ± 0.04 (0.7) |
| Δ5, 24-Stigmastadienol | 4.25 ± 0.04 (0.23) | 5.13 ± 0.01 (0.20) | 5.67 ± 0.66 (0.27) | 5.11 ± 0.74 (0.23) | 0.12 ± 0.04 (0.16) |
| Δ7-Stigmastenol | 13.56 ± 1.85 (0.62) | 13.34 ± 0.96 (0.60) | 14.12 ± 0.70 (0.66) | 13.68 ± 1.15 (0.63) | 0.42 ± 0.04 (0.56) |
| Δ7-Avenasterol | 8.22 ± 1.01 (0.33) | 7.03 ± 1.18 (0.32) | 8.12 ± 0.37 (0.38) | 7.64 ± 1.00 (0.35) | 0.31 ± 0.04 (0.42) |
| Total PS | 2173.01 ± 101.26 | 2231.25 ± 73.12 | 2136.86 ± 101.51 | 2180.37 ± 90.43 | 74.65 ± 5.15 |
The order of individual PS relative percentage from the PS-ingredient, determined from the greatest to least abundance, is as follows: β-sitosterol (78.79%), sitostanol (10.21%), campesterol (7.43%), campestanol (1.37%), Δ5-avenasterol (0.7%), Δ7-stigmastenol (0.56%), stigmasterol (0.52%), Δ7-avenasterol (0.42%) and Δ5,24(25)-stigmastadienol (0.16%). These results comply with the European regulations that regulate the enrichment with PS of rye bread and states that the relative percentages of different PS must be: <80% β-sitosterol < 15% β-sitostanol < 40% campesterol < 5% campestanol < 30% stigmasterol < 3% brassicasterol < 3% other sterols/stanols.4 These data are comparable with previous determinations with different ingredients but of the same origin, showing stability in this type of PS-source ingredient, not only in their total content (70.9 vs. 74.7 g PS/100 g), but also in the relative percentages of different individual PS: β-sitosterol (78.86%), sitostanol (11.95%), campesterol (7.13), campestanol (1.20), stigmasterol (0.82), and brassicasterol (0.04).28 These authors identified brassicasterol (0.04), not detected in our ingredient. The phytosterols and phytostanols identified in WRB represent an average relative percentage of 88.5% and 11.5%, respectively. In this sense, phytostanols, mainly campestanol, predominated in the steryl ferulates of grain milling fractions (including rye), whereas their proportion of the total sterols was 14–30%,28 representing about 19.3% in rye bread.22
The order of individual PS contents in WRB from the highest to lowest is as follows: β-sitosterol, sitostanol, campesterol, campestanol, Δ5-avenasterol, Δ7-stigmastenol, stigmasterol, Δ7-avenasterol and 24(25)-stigmastadienol. This result is consistent with the fact that cereal products are one of the most important natural sources of PS, 4-desmethylsterols (sitosterol, campesterol, stigmasterol, Δ5-avenasterol, Δ7-avenasterol) being predominant, specially β-sitosterol (representing in our rye bread 79% of the total PS). The abovementioned order is similar to that indicated in rye bread by other authors,22 except that campesterol is higher than sitostanol and stigmasterol of the same order as Δ5-avenasterol. These changes are explained by the contribution of the PS-ingredient, whose huge PS content causes the order of these sterols to vary. The content of sitostanol added with the PS-ingredient was higher than that of campesterol (10.21 vs. 7.43% of total ingredient PS) and the content of Δ5-avenasterol added with the PS-ingredient was also higher than that of stigmasterol (0.7 vs. 0.52% of total ingredient PS).
No statistically significant differences (p > 0.05) were observed in the individual and total PS contents under different baking days (see Table 2). It is confirmed that the desired PS-enrichment is maintained under different baking days if they are carried out with the same batch of flour and if baking conditions remain identical between them. It is known that the content of PS in grains of different rye cultivars from the same geographical area and in the same harvest does not differ significantly. The most important determining factor of the PS content is the composition of ingredients from different milling products (wholemeal flour, bran and germ) or flours with different degrees of extraction (% ash).22,29 The results obtained allows comparing PS bioaccessibility assays in rye bread, carried out under different digestion conditions without influencing individual and total PS contents.
3.2 Adaptation of the gastrointestinal conditions to the elderly in PDB
Previous experiments of gastrointestinal digestion adapted to elderly with in vivo mastication were first carried out on fresh WRB with the conditions of gastric and intestinal phases indicated in Fig. 1. The results showed a great variability (27–64%) in individual and total PS content of BF and bioaccessibility between the different digestion replicates (data not shown).Mastication with only 20 cycles does not allow an adequate disintegration of the bread, which could cause variability in the action of the enzymes due to mechanical reasons and hinder the release of the matrix, and the micellarization of PS. That hypothesis could be explained with the results of a trial comparing matrix disintegration in rye bread and wheat bread after mastication and in vitro gastric digestion. It was observed that in sourdough wholemeal rye bread a greater number of large-sized particles (>2 mm) and a greater aggregation of these were maintained compared to wheat bread.30
Since the data obtained with human mastication adapted to older people could not be used in this work to accurately assess the bioaccessibility of PS, tests with partially dehydrated WRB were reproduced to achieve more homogeneous digestion. In addition, due to economic costs (it will be needed to increase the concentration of α-amylase), together with the fact that the oral phase is not a determinant in the bioaccessibility of lipophilic compounds such as plant sterols, lead us to maintain adult oral phase conditions. The PS contents determined in the PDB and the BF of both three conditions (adult, E1 and E2) assessed are shown in Table 3. The bioaccessibility of each individual PS and total PS compared between the conditions studied can be seen in Fig. 3.
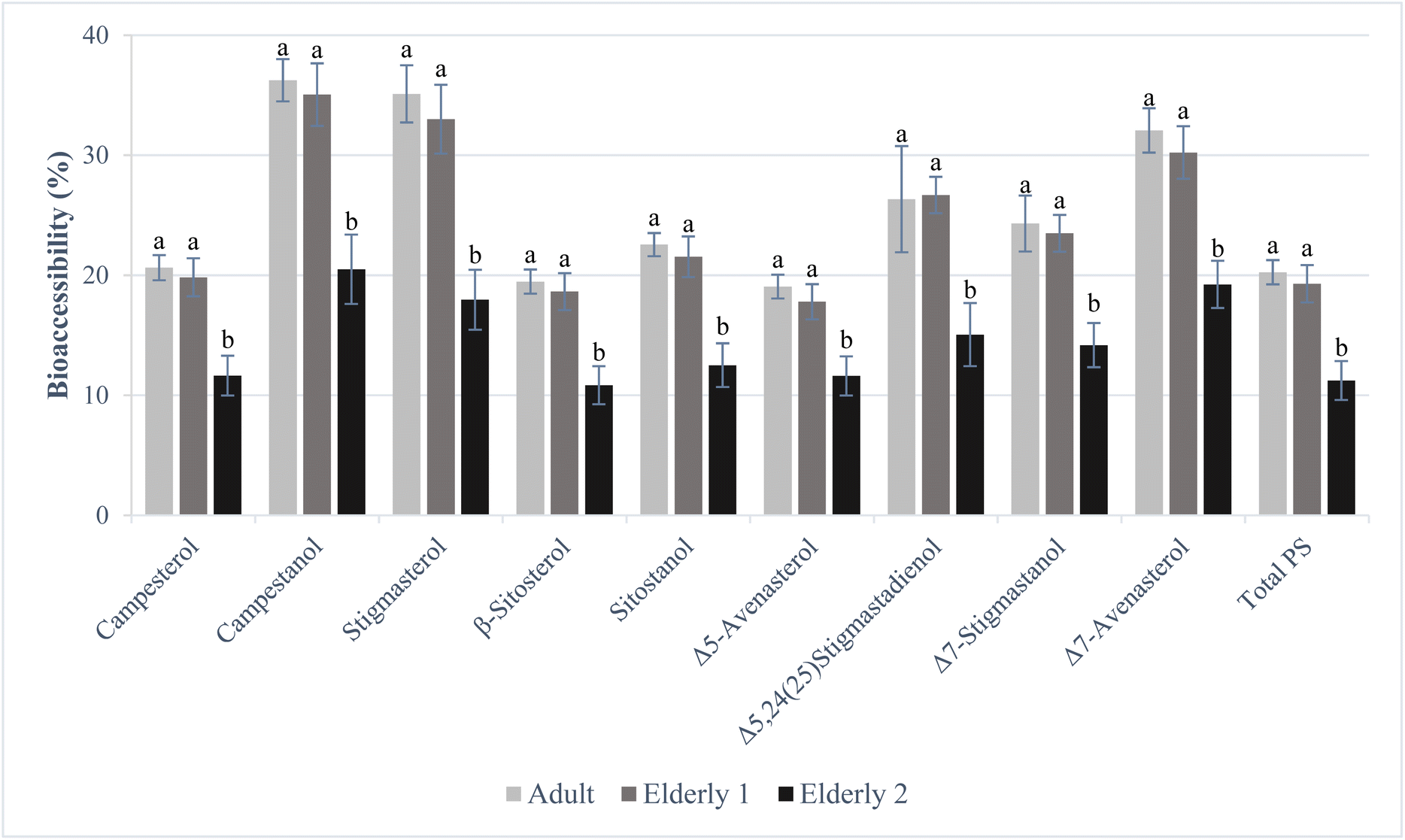 | ||
| Fig. 3 Bioaccessibility (%) of plant sterols (PS) assessed under adult, elderly 1 and elderly 2 conditions. Results were expressed as mean ± standard deviation (n = 6). Different lowercase letters (a and b) in each PS indicate statistically significant differences (p < 0.05) in PS bioaccessibility between conditions. | ||
| Plant sterol | Rye bread | Control (C) BF | Elderly 1 BF | Elderly 2 BF |
|---|---|---|---|---|
| Campesterol | 159.47 ± 6.68 | 32.90 ± 1.66a | 31.63 ± 2.53a | 18.57 ± 2.65b |
| Results were expressed as mean ± standard deviation (n = 6 replicate). Different lowercase letters (a and b) in each sterol indicate statistically significant differences (p < 0.05) between the FB content under each condition. | ||||
| Campestanol | 17.61 ± 2.51 | 6.38 ± 0.31a | 6.17 ± 0.46a | 3.61 ± 0.51b |
| Stigmasterol | 8.76 ± 0.80 | 3.08 ± 0.21a | 2.89 ± 0.25a | 1.57 ± 0.22b |
| β-Sitosterol | 1815.75 ± 116.78 | 353.68 ± 18.33a | 338.56 ± 27.92a | 196.93 ± 28.86b |
| Sitostanol | 225.09 ± 30.95 | 50.78 ± 2.17a | 48.51 ± 3.82a | 28.17 ± 4.09b |
| Δ5-Avenasterol | 20.56 ± 2.64 | 3.92 ± 0.20a | 3.66 ± 0.30a | 2.39 ± 0.33b |
| Δ5,24-Stigmastadienol | 4.65 ± 1.09 | 1.23 ± 0.21a | 1.24 ± 0.07a | 0.70 ± 0.12b |
| Δ7-Stigmastenol | 15.58 ± 0.91 | 3.79 ± 0.36a | 3.66 ± 0.24a | 2.21 ± 0.29b |
| Δ7-Avenasterol | 10.31 ± 1.58 | 3.30 ± 0.19a | 3.12 ± 0.23a | 1.98 ± 0.20b |
| Total PS | 2277.79 ± 158.62 | 459.06 ± 22.98a | 439.44 ± 35.62a | 256.14 ± 36.93b |
No statistically significant differences (p > 0.05) were observed in the individual and the total PS contents of the BF or their respective bioaccessibility under gastric elderly conditions (E1 modifications) (individual PS range of 17.8–35.1%, total PS 19.3%) with those obtained under healthy adult conditions (individual PS range of 19.1–36.3%, total PS 20.3%). However, when these same two conditions were compared with the gastrointestinal elderly conditions (E2 modifications), significant reductions (p < 0.05) were observed (individual PS range of 10.9–20.5%, total PS 11.2%). The relative magnitude of the reduction was similar in all nine PS determined: Δ5-avenasterol (39%), Δ7-avenasterol (40%), Δ7-stigmastenol (42%), Δ5,24-stigmastadienol (43%), campestanol (43%), campesterol (44%), β-sitosterol (44%), sitostanol (45%) and stigmasterol (49%).
The changes made in the gastric phase (modification E1) did not produce a significant change in the bioaccessibility vs. adult, in contrast with another previous study of adaptation of digestion to the elderly conditions that evaluated sterol bioaccessibility in a PS-enriched beverage. The same modification of the gastric phase produced a significant increase (73%) in the bioaccessibility of total PS.18 The differences between both food matrices (solid vs. liquid) could partially explain this observation. In fact, it has been observed that the GL contributes to a greater extent to the digestion of TAG in liquid foods (25%) than in solid foods (10%).20 In this context, GL has difficult access to its substrate (TAG), and its action could be greatly reduced since WRB is a very complex matrix rich in fiber with a solid structure. Besides, this could explain why, even though it has been observed that a decrease in the activity of the GL noted in the adaptation to the elderly translates into a higher bioaccessibility of PS in a liquid matrix by not being able to perform its action correctly,17 in a solid matrix, neither in the control nor in the gastric elderly model of the present study did these expected differences occurred.
The lack of the effect of GL could be attributed to the influence of the soluble fiber present in the WRB, that contains up to twice as much β-glucan and fructan than wheat bran, while it contains up to half the cellulose.31 In this sense, the addition of different soluble fibers (guar gum, pectins or Arabic gum) at increasing concentrations (0.3 and 2% w/v) to a lipid mixture (triolein/phosphatidylcholine/cholesterol) produced larger lipid droplets compared to those without the presence of fiber. This made it difficult for GL to access triacylglycerols. The authors attributed this to the higher viscosity in the case of the presence of soluble fiber. In addition, this effect seems to be dose dependent. The presence of Arabic gum and high viscosity guar gum decreases the degree of lipolysis of triacylglycerols through a decrease in the effectiveness of GL.32 This may explain why a food as rich in fiber as WRB (and in soluble fiber in particular), has its lipolysis at the gastric level greatly diminished.
The reduction of PS bioaccessibility observed (Fig. 3) under gastric and intestinal modified elderly conditions (E2) in WRB could be justified by the fiber effect. Fiber might cause lower lipid and cholesterol absorption. Lower bioaccessibility (up to 24% reduction in fat bioaccessibility and up to 22% reduction in cholesterol bioaccessibility) was observed after digestion in a multi-compartmental gastrointestinal model,15 when a partially hydrolyzed guar gum at 3 and 6% concentration (cholesterol:fiber ratio 0.04 and 0.02 respectively) was added to yogurt enriched with egg yolk (4%) and sunflower oil (3%). Similar findings were obtained from an INFOGEST simulated digestion trial with pork patties with and without the addition of different fiber extracts (from lemon, pomegranate, lemon albedo, grapefruit and tiger nut). Lower cholesterol solubility (ranging from 10 to 31% reduction depending on the fiber extract used) from fiber-added patties, when compared with fiber-less patties, was observed.16 However, in another INFOGEST simulated digestion study in a PS-enriched milk-based fruit beverage, the addition of 2.5 and 5 g of galactooligosaccharides per 250 mL of beverage did not cause any change in the bioaccessibility of sterols compared to the beverage without added galactooligosaccharides. The authors attributed this fact to the low content of fiber in relation with sterols (ratio of sterol:fiber 0.98–0.6), which could have prevented the fiber from having an effect on reducing the bioaccessibility of sterols.33 This is consistent with the results obtained in our work, since the PS:fiber ratio in this case is 0.17, more similar to that obtained by Minekus et al.15 It could explain that, in order to observe the effect of reducing the bioaccessibility of sterols due to fiber, not only the total fiber content must be considered, but also its ratio with respect to sterols.
The elderly conditions that varied in the intestinal phase with respect to the adult ones could be responsible for the decrease in bioaccessibility, highlighting the reduction of enzymes and bile salts and agitation during digestion. The reduction of pancreatin was 50%, which translates into lower activity. The pancreatic lipase, in this case responsible for the complete lipolysis (since in the previous phase the GL could not exert its effect correctly), will produce partial lipolysis. This hypothesis is supported by the results obtained in olive oil by adding different concentrations of high molecular weight soluble fiber extracted from Lentinula edodes. An inhibition of the activity of pancreatic lipase in the sample was observed, and this inhibition was correlated with the fiber concentration (up to 81.9% inhibition with 10 mg mL−1 of fiber).34
In addition to the activity of the enzymes themselves, for the correct micellarization of sterols, greater emulsification is necessary. Bile salts increase the bioaccessibility of lipophilic compounds such as sterols since they support correct emulsification of lipids. Fiber is one of the compounds that binds to bile acids, as it occurs when joining methylcellulose and bile salt sodium deoxycholate,35 preventing bile acids from emulsifying sterols. When they are bound to the fiber, bile acids are eliminated in the feces, and they are not available to emulsify sterols.36 As far as we know, there are no studies that have evaluated the impact of fibers on the action of bile salts and sterol micellarization, all combined. However, it has been observed that compounds such as polyphenols bind with bile salts due to their physicochemical properties, preventing the correct micellarization of sterols. This aspect was observed in a trial combining epigallocatechin gallate and taurocholic acid, where such a combination caused lower solubility of cholesterol and phosphatidylcholine in the micelles.37 This effect is consistent with the need for a greater amount of bile salts (as is the case under adult gastrointestinal conditions) to emulsify the same amount of sterols in the presence of fiber, which could explain the decrease of bioaccessibility in the elderly due to the halving of bile salts. This explanation could also be supported by the decrease in agitation during intestinal digestion. To achieve the emulsion, it is necessary for the fat drops to be mixed correctly, and the least frequency of agitation could possibly prevent this process, supported by the high viscosity of the mixture due to the presence of soluble fiber.
The dietary fiber content also produces a slower release of carbohydrates, which can also affect fat breakdown and micellarization.38 This would support the hypothesis of reduced intestinal absorption of sterols and hence the change in bioaccessibility by reducing bile salts and pancreatin.
4. Conclusions
The results of this study suggest that the elderly gastrointestinal conditions can reduce PS bioaccessibility in WRB compared with adult conditions. This fact could be attributed to both the solid nature of the food and the interactions of matrix components (ratio of PS/fiber) and bile salts, as well as to the reduction of other gastric and intestinal enzymes under elderly conditions. Even though this could be a problem for the elderly population, this food matrix may be a good dietary option for them, especially if its consumption replaces the consumption of white wheat bread. WRB has a good nutritional profile due to its high dietary fiber content, which can add to the hypocholesterolemic effect provided by the PS and act as a good dietary treatment for moderate hypercholesterolemia. The main limitation of this study is the difficulty of correctly simulating the conditions of the elderly, either due to the lack of standardization, the impossibility of static studies to generate a pH gradient, so the chosen pH may not represent the entire elderly population, or the fact of not having been able to perform in vivo adapted mastication. Additional dynamic digestion and in vivo assays are needed to assess the bioactivity of PS in this population and ultimately be able to tailor dietary recommendations specifically to the elderly, and also to evaluate the possible health benefits from the consumption of WRB in this population group.Author contributions
Diego Miedes and Mussa Makran: investigation, formal analysis, and writing original draft; Guadalupe Garcia-Llatas, Amparo Alegría, Reyes Barberá and Antonio Cilla: conceptualization, funding acquisition, project administration, supervision, writing original draft, and review & editing.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This publication is part of the project PID2019-104167RB-I00 funded by MCIN/AEI/10.13039/501100011033. Mussa Makran holds an FPU19/00156 grant from the Ministry of Universities (Spain). Diego Miedes holds an CPI-22-450 contract from the Investigo Program (Generalitat Valenciana, Spain).The authors are grateful to Dra. Claudia Monika Haros from the Institute of Agrochemistry and Food Technology (IATA-CSIC) for contribution to the wholemeal rye bread production.
References
- United Nations, Department of International Economic and Social Affairs, Population Division, World Population Ageing 2019, https://www.un.org/en/development/desa/population/publications/pdf/ageing/WorldPopulationAgeing2019-Highlights.pdf, Accessed 2022-04-12.
- World Health Organization, The top 10 causes of death, 2020, https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death, Accessed 2022-05-16.
- Commission Regulation (EU) No 686/2014 of 20 June 2014 amending Regulations (EC) No 983/2009 and (EU) No 384/2010 as regards the conditions of use of certain health claims related to the lowering effect of plant sterols and plant stanols on blood LDL-cholesterol, Off. J. Eur. Union, 2014, L182, 27–30 Search PubMed.
- Commission Decision 2006/59/EC of 24 January 2006 authorizing the placing on the market of rye bread with added phytosterols/phytostanols as novel foods or novel food ingredients under Regulation (EC) No 258/97 of the European Parliament and of the Council, Off. J. Eur. Union, 2006, L31, 21–23 Search PubMed.
- J. Wouk, R. F. H. Dekker, E. A. I. F. Queiroz and A. M. Barbosa-Dekker, β-Glucans as a panacea for a healthy heart? Their roles in preventing and treating cardiovascular diseases, Int. J. Biol. Macromol., 2021, 177, 176–203 CrossRef CAS PubMed.
- I. M. V. Silva, F. Machado, M. J. Moreno, C. Nunes, M. A. Coimbra and F. Coreta-Gomes, Polysaccharide structures and their hypocholesterolemic potential, Molecules, 2021, 26, 4559 CrossRef CAS PubMed.
- R. M. Nagler and O. Hershkovich, Relationships between age, drugs, oral sensorial complaints and salivary profile, Arch. Oral Biol., 2005, 50, 7–16 CrossRef PubMed.
- D. Rémond, D. R. Shahar, D. Gille, P. Pinto, J. Kachal, M.-A. Peyron, C. N. Dos Santos, B. Walther, A. Bordoni, D. Dupont, L. Tomás-Cobos and G. Vergères, Understanding the gastrointestinal tract of the elderly to develop dietary solutions that prevent malnutrition, Oncotarget, 2015, 6, 13858–13898 CrossRef PubMed.
- H. Moreau, R. Lautier, Y. Gargouri, F. Ferrato and R. Verger, Human preduodenal lipase is entirely of gastric fundic origin, Gastroenterology, 1988, 95, 1221–1226 CrossRef CAS PubMed.
- R. Laugier, J. P. Bernard, P. Berthezene and P. Dupuy, Changes in pancreatic exocrine secretion with age: Pancreatic exocrine secretion does decrease in the elderly, Digestion, 1991, 50, 202–211 CAS.
- N. Salles, Basic mechanisms of the aging gastrointestinal tract, Dig. Dis., 2007, 25, 112–117 CrossRef CAS PubMed.
- M. Minekus, M. Alminger, P. Alvito, S. Ballance, T. Bohn, C. Bourlieu, F. Carrière, R. Boutrou, M. Corredig, D. Dupont, C. Dufour, L. Egger, M. Golding, S. Karakaya, B. Kirkhus, S. Le Feunteun, U. Lesmes, A. Macierzanka, A. Mackie, S. Marze, D. J. McClements, O. Ménard, I. Recio, C. N. Santos, R. P. Singh, G. E. Vegarud, M. S. J. Wickham, W. Weitschies and A. Brodkorb, A standardised static in vitro digestion method suitable for food—An international consensus, Food Funct., 2014, 5, 1113–1124 RSC.
- A. Brodkorb, L. Egger, M. Alminger, P. Alvito, R. Assunção, S. Ballance, T. Bohn, C. Bourlieu-Lacanal, R. Boutrou, F. Carrière, A. Clemente, M. Corredig, D. Dupont, C. Dufour, C. Edwards, M. Golding, S. Karakaya, B. Kirkhus, S. Le Feunteun, U. Lesmes, A. Macierzanka, A. R. Mackie, C. Martins, S. Marze, D. J. McClements, O. Ménard, M. Minekus, R. Portmann, C. N. Santos, I. Souchon, R. P. Singh, G. E. Vegarud, M. S. J. Wickham, W. Weitschies and I. Recio, INFOGEST static in vitro simulation of gastrointestinal food digestion, Nat. Protoc., 2019, 14, 991–1014 CrossRef CAS PubMed.
- M. Makran, D. Miedes, A. Cilla, R. Barberá, G. Garcia-Llatas and A. Alegría, Understanding the influence of simulated elderly gastrointestinal conditions on nutrient digestibility and functional properties, Trends Food Sci. Technol., 2022, 129, 283–295 CrossRef CAS.
- M. Minekus, M. Jelier, J. Z. Xiao, S. Kondo, K. Iwatsuki, S. Kokubo, M. Bos, B. Dunnewind and R. Havenaar, Effect of partially hydrolyzed guar gum (PHGG) on the bioaccessibility of fat and cholesterol, Biosci. Biotechnol. Biochem., 2005, 69, 932–938 CrossRef CAS PubMed.
- M. C. López-Marcos, C. Bailina, M. Viuda-Martos, J. A. Pérez-Álvarez and J. Fernández-López, Effects of various fibre-rich extracts on cholesterol binding capacity during in vitro digestion of pork patties, Food Funct., 2015, 6, 3473–3478 RSC.
- M. Makran, N. Faubel, G. López-García, A. Cilla, R. Barberá, A. Alegría and G. García-Llatas, Sterol bioaccessibility in a plant sterol-enriched beverage using the INFOGEST digestion method: Influence of gastric lipase, bile salts and cholesterol esterase, Food Chem., 2022, 382, 132305 CrossRef CAS PubMed.
- D. Miedes, M. Makran, R. Barberá, A. Cilla, A. Alegría and G. Garcia-Llatas, Elderly gastrointestinal conditions increase sterol bioaccessibility in a plant sterol-enriched beverage: adaptation of the INFOGEST method, Food Funct., 2022, 13, 4478–4485 RSC.
- S. Amara, C. Bourlieu, L. Humbert, D. Rainteau and F. Carrière, Variations in gastrointestinal lipases, pH and bile acid levels with food intake, age and diseases: Possible impact on oral lipid-based drug delivery systems, Adv. Drug Delivery Rev., 2019, 142, 3–15 CrossRef CAS PubMed.
- E. Hernández-Olivas, S. Muñoz-Pina, A. Andrés and A. Heredia, Impact of cooking preparation on in vitro digestion of eggs simulating some gastrointestinal alterations in elders, J. Agric. Food Chem., 2021, 69, 4402–4411 CrossRef PubMed.
- N. Faubel, M. Makran, A. Cilla, A. Alegría, R. Barberá and G. Garcia-Llatas, Bioaccessibility of plant sterols in wholemeal rye bread using the INFOGEST protocol: influence of oral phase and enzymes of lipid metabolism, J. Agric. Food Chem., 2022, 70, 13223–13232 CrossRef CAS PubMed.
- V. Piironen, J. Toivo and A. M. Lampi, Plant sterols in cereals and cereal products, Cereal Chem., 2002, 79, 148–154 CrossRef CAS.
- A. Alvarez-Sala, G. Garcia-Llatas, A. Cilla, R. Barberá, L. M. Sánchez-Siles and M. J. Lagarda, Impact of lipid components and emulsifiers on plant sterols bioaccessibility from milk-based fruit beverages, J. Agric. Food Chem., 2016, 64, 5686–5691 CrossRef CAS PubMed.
- A. Yorulmaz, Y. S. Velioglu, A. Tekin, A. Simsek, J. C. Drover and J. Ates, Phytosterols in 17 Turkish hazelnut (Corylus avellana L.) cultivars, Eur. J. Lipid Sci. Technol., 2009, 111, 402–408 CrossRef CAS.
- M. Drira, H. Jabeur, F. Marrakchi and M. Bouaziz, Delta-7-stigmastenol: quantification and isomeric formation during chemical refining of olive pomace oil and optimization of the neutralization step, Eur. Food Res. Technol., 2018, 244, 2231–2241 CrossRef CAS.
- NIST Database of organic chemistry compounds: chemistry WebBook, 2015, https://webbook.nist.gov/chemistry, Accessed 2022-01-20.
- G. Garcia-Llatas, A. Alegría, R. Barberá and A. Cilla, Current methodologies for phytosterol analysis in foods, Microchem. J., 2021, 168, 106377 CrossRef CAS.
- M. González-Larena, G. García-Llatas, M. C. Vidal, L. M. Sánchez-Siles, R. Barberá and M. J. Lagarda, Stability of plant sterols in ingredients used in functional foods, J. Agric. Food Chem., 2011, 59, 3624–3631 CrossRef PubMed.
- P. Hakala, A. M. Lampi, V. Ollilainen, U. Werner, M. Murkovic, K. Wähälä, S. Karkola and V. Piironen, Steryl phenolic acid esters in cereals and their milling fractions, J. Agric. Food Chem., 2002, 50, 5300–5307 CrossRef CAS PubMed.
- E. Nordlund, K. Katina, H. Mykkänen and K. Poutanen, Distinct characteristics of rye and wheat breads impact on their in vitro gastric disintegration and in vivo glucose and insulin responses, Foods, 2016, 5, 24 CrossRef PubMed.
- A. Kamal-Eldin, H. N. Lærke, K. E. B. Knudsen, A. M. Lampi, V. Piironen, H. Adlercreutz, K. Katina, K. Poutanen and P. Aman, Physical, microscopic and chemical characterisation of industrial rye and wheat brans from the Nordic countries, Food Nutr. Res., 2009, 53, 1912 CrossRef PubMed.
- B. Pasquier, M. Armand, C. Castelain, F. Guillon, P. Borel, H. Lafont and D. Lairon, Emulsification and lipolysis of triacylglycerols are altered by viscous soluble dietary fibres in acidic gastric medium in vitro, Biochem. J., 1996, 314, 269–275 CrossRef CAS PubMed.
- V. Blanco-Morales, G. López-García, A. Cilla, G. Garcia-Llatas, R. Barberá, M. J. Lagarda, L. M. Sánchez-Siles and A. Alegría, The impact of galactooligosaccharides on the bioaccessibility of sterols in a plant sterol-enriched beverage: adaptation of the harmonized INFOGEST digestion method, Food Funct., 2018, 9, 2080–2089 RSC.
- Z. Xue, X. Gao, Y. Jia, Y. Wang, Y. Lu, M. Zhang, P. Panichayupakaranant and H. Chen, Structure characterization of high molecular weight soluble dietary fiber from mushroom Lentinula edodes (Berk.) Pegler and its interaction mechanism with pancreatic lipase and bile salts, Int. J. Biol. Macromol., 2020, 153, 1281–1290 CrossRef CAS PubMed.
- S. K. Behera, S. Mishra and M. Mohapatra, Physicochemical study of dietary fiber methylcellulose and human intestinal bile salt micellar aggregates, Colloid Interface Sci. Commun., 2021, 44, 100493 CrossRef CAS.
- J. Singh, R. Metrani, S. R. Shivanagoudra, G. K. Jayaprakasha and B. S. Patil, Review on bile acids: Effects of the gut microbiome, interactions with dietary fiber, and alterations in the bioaccessibility of bioactive compounds, J. Agric. Food Chem., 2019, 67, 9124–9138 CrossRef CAS PubMed.
- K. Ogawa, S. Hirose, S. Nagaoka and E. Yanase, Interaction between tea polyphenols and bile acid inhibits micellar cholesterol solubility, J. Agric. Food Chem., 2015, 64, 204–209 CrossRef PubMed.
- E. C. Shin, R. B. Pegg, D. Phillips and R. R. Eitenmiller, Commercial peanut (Arachis hypogaea L.) cultivars in the United States: phytosterol composition, J. Agric. Food Chem., 2010, 58, 9137–9146 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2023 |