
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceIn vitro comparison of whey protein isolate and hydrolysate for their effect on glucose homeostasis markers
Mathie
Tenenbaum
*a,
Camille
Dugardin
a,
Joanna
Moro
b,
Julie
Auger
b,
Alain
Baniel
b,
Audrey
Boulier
b,
Rozenn
Ravallec
a and
Benoit
Cudennec
 *a
*a
aUMR Transfrontalière BioEcoAgro No 1158, Univ. Lille, INRAE, Univ. Liège, UPJV, YNCREA, Univ. Artois, Univ. Littoral Côte d'Opale, ICV – Institut Charles Viollette, F-59000 Lille, France. E-mail: mathie.tenenbaum@univ-lille.fr; benoit.cudennec@univ-lille.fr
bIngredia S.A., 51 Av. Lobbedez, CS 60946, 62033 Arras Cedex, France
First published on 29th March 2023
Abstract
Research on new strategies to regulate glucose homeostasis to prevent or manage type 2 diabetes is a critical challenge. Several studies have shown that protein-rich diets could improve glucose homeostasis. Whey protein hydrolysis allows the release of amino acids and bioactive peptides, which exert numerous well-documented bioactivities. This study evaluates and compares the hypoglycemic potential of a whey protein hydrolysate and a whey protein isolate after static in vitro simulated gastrointestinal digestion (SGID) using the INFOGEST protocol. The peptide molecular mass distributions of the digested samples were evaluated by size exclusion chromatography and show that after digestion, the whey hydrolysate is significantly more hydrolyzed. After SGID, the whey protein hydrolysate induces a significative greater secretion of GLP-1 after two hours of contact with the enteroendocrine STC-1 cell line than the whey protein after isolation. In addition, the digested whey hydrolysate increases preproglucagon (GCG) and pro-convertase-1 (PCSK1) expression. The digested hydrolysate also inhibits the DPP-IV activity after an intestinal barrier passage challenge using a Caco-2/HT29-MTX mixed-cell model. Our results highlight that the prehydrolysis of whey proteins modify the intestinal peptidome, leading to a potentially greater hypoglycemic effect. This study confirms the previously observed in vitro hypoglycemic effect of this hydrolysate and evidences the beneficial impact of the industrial hydrolysis process.
1. Introduction
Glucose homeostasis allows the maintenance of an adequate glucose concentration in the blood throughout the day and according to energy intake. Many organs, including the intestine, take part in glucose homeostasis regulation. Its dysregulation leads to various pathologies, including diabetes. Although there are different types of diabetes, type 2 diabetes (T2D) is the most widespread form, representing 90% of diagnosed forms of diabetes.1 The etiology of the disease is complex, involving both genetic and environmental causes. In addition, some factors favor the appearance of T2D, such as obesity or ageing.2 The disease results from insufficient insulin production to compensate for an increased demand caused by insulin resistance of the target tissues.3 Diabetes may lead to many complications such as ketoacidosis, vascular complications, retinopathy, nephropathy, heart disease, and neuropathy.1 In addition to physical exercise and dietary measures, there are various antidiabetic treatments, including metformin, alpha-glucosidase inhibitors, glucagon-like peptide 1 (GLP-1) mimetics, and dipeptidyl peptidase-4 (DPP-IV) inhibitors.4At the intestinal level, glucose homeostasis is regulated mainly by intestinal hormones, including incretins such as GLP-1, which is known to not only increase insulin secretion and decrease glucagon secretion in the pancreas but also to delay gastric emptying and increase satiety.5 GLP-1 is produced by the enzymatic cleavage of the pro-glucagon protein, which is derived from the preproglucagon gene (GCG). The enzyme responsible for its cleavage is pro-convertase-1 (PCSK-1). GLP-1 secretion is stimulated by glucose, fatty acids, dietary peptides, amino acids, acetylcholine, and interleukin-6. Its receptor, GLP-1-R, is present on the surface of various cell types, including key organs involved in glucose homeostasis regulation.6 However, the half-life of GLP-1 is short due to the action of DPP-IV, which is a peptidase of the serine protease family. DPP-IV exists in transmembrane and circulatory forms and is widely expressed and present in the blood compartment and enterocytes.7 This enzyme is involved in numerous cellular and molecular functions. Regarding glucose homeostasis, DPP-IV inactivates the incretins, glucose-dependent insulinotropic polypeptide (GIP) and GLP-1.8
Cow milk contains an average of 35 g L−1 of proteins, which are caseins and whey proteins, representing 80% and 20% of the protein content, respectively. Whey proteins are mainly β-lactoglobulin, α-lactalbumin, and other proteins such as immunoglobulin and enzymes. Whey could be used in obesity and T2D prevention since its action on insulin secretion, postprandial glycemia, and weight loss have been demonstrated via the incretin system.9–12
During gastrointestinal digestion, dietary proteins are broken down into peptides by enzymes in the stomach and the intestine and then digested by peptidases in the brush border into free amino acids and di- and tri-peptides, which are absorbed. Numerous resistant peptides interact especially with receptors of enteroendocrine cells (EECs), leading to intestinal hormone secretion, such as GLP-1.13,14In vitro, it was evidenced that digested dietary proteins stimulate the secretion of intestinal hormones, including GLP-1, and concomitantly inhibit the action of DPP-IV.15–18
The regulation of blood glucose at the intestinal level may also involve the increase of the proglucagon gene expression. In the intestine, several peptides are released after the proteolytic cleavage of proglucagon by the enzyme pro-convertase 1 (PCSK1), including GLP-1. These different peptides (GLP-1, GLP-2, Oxyntomodulin, Glicentin, and IP-2) have a major role in glucose homeostasis, particularly in maintaining the circulating GLP-1 levels.19,20 Dietary proteins also increase proglucagon and PCSK1 gene expression in enteroendocrine STC-1 cells.21,22
Pep2dia® is a whey protein hydrolysate containing AP dipeptides, which are known to inhibit alpha-glucosidase, a digestive enzyme that hydrolyzes polysaccharides into glucose. A previous clinical study evidenced that when it was taken before a meal by prediabetic individuals, the postprandial glycemia was reduced significantly.23
In this study, we measure the impact of static in vitro gastrointestinal digestion (SGID) on the peptide molecular mass distribution of a digested whey protein hydrolysate (dWPH) and a digested whey protein isolate (dWPI) and investigate and compare their effects on GLP-1 secretion and DPP-IV activity. The ability of dWPH to modulate GCG and PCSK1 gene expressions and to inhibit the DPP-IV activity after intestinal barrier (IB) passage was also assayed.
2. Experimental
2.1. Materials
The whey protein isolate (WPI), Promilk852FB1, was produced from native whey extracted by filtration according to Boutrou et al.24 Pep2Dia® is a whey protein hydrolysate (WPH) obtained by protease hydrolysis of the WPI and containing a bioactive arginine-proline (AP) dipeptide (between 0.15% and 0.4%) with alpha-glucosidase inhibiting properties.20 The two protein preparations were prepared by Ingredia S.A. (Arras CEDEX, France). The total nitrogen content of the powders was evaluated by the Kjeldahl method and was converted to the protein content with a conversion factor of 6.3. The protein contents of WPI and WPH were 80.1% and 73.0%, respectively.2.2. Simulated gastrointestinal digestion (SGID)
WPI and WPH (2 grams of protein) were digested in vitro according to the harmonized static INFOGEST protocol,25 which was adapted for the protein alone.26 The simulated fluid composition and the different digestion steps were described previously.27 Briefly, the samples previously solubilized in water (8 mL) were added to the simulated salivary fluid (8 mL) for 5 minutes at pH 7.0. In the second step, simulated gastric fluid containing pepsin (SIGMA, P6887, 6500 U mL−1) was added (volume to volume) to the incubator, and the whole mixture was incubated for 2 hours at pH 3.0. In the last step, simulated intestinal fluid containing pancreatin (SIGMA, P1750, trypsin activity 45 U mL−1) was added (volume to volume) to the mix and incubated for 2 hours at pH 7.0. All steps were performed at 37 °C under agitation. The intestinal phase was collected, heated at 95 °C for 5 minutes for enzyme denaturation, centrifuged at 13![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 400g for 5 minutes, and the supernatant was frozen at −20 °C until the different experiments. The control used for the various experiments is a blank SGID (BLK): SGID performed without any sample but with gastrointestinal enzymes and fluids.
400g for 5 minutes, and the supernatant was frozen at −20 °C until the different experiments. The control used for the various experiments is a blank SGID (BLK): SGID performed without any sample but with gastrointestinal enzymes and fluids.
2.3. Size exclusion chromatography by fast protein liquid chromatography (SEC-FPLC)
Digested samples (50 μL) at a concentration of 31.25 mg mL−1 were separated on a Superdex peptide 10/300 GL column (Cytiva, Velizy Villacoublay, France) under isocratic elution with a flow rate of 0.5 mL min−1 in a solvent: 69.9% ultrapure water; 30% acetonitrile; 0.1% TFA. The separation was performed for 1 hour. The column was previously calibrated with the following molecular mass standards: Bovine Serum Albumin (60 kDa); Cytochrome C (12384 Da); Vitamin B12 (1355 Da); Glutathione (612.6 Da) from Sigma-Aldrich (St Louis, MO, USA), a decapeptide (1266.7 Da) and a tri-decapeptide (3416 Da) chemically synthesized (Genecust, Boynes, France). The absorbance was measured at 214 nm using an ÂKTA purifier protein purification system (Cytiva).
2.4. Cell line and culture
In this study, three intestinal cell lines were used: Caco-2 cells, human colorectal carcinoma cell line (Sigma-Aldrich, St Louis, MO, USA); HT29-MTX cells, human mucus-secreting adenocarcinoma cell line (Sigma-Aldrich); and STC-1 cells, murine enteroendocrine cell line (Sigma-Aldrich). Cell lines were cultured in a complete medium: Dulbecco's Modified Eagle's Medium (DMEM) (PAN Biotech, Aidencach, Germany), supplemented with 10% (v/v) Fetal Bovine Serum (FBS), with 100 U mL−1 of antibiotics (penicillin and streptomycin) and 2 mM L-glutamine. The cells were maintained in 75 cm2 flasks (Sarstedt, Nümbrecht, Germany) at 37 °C under 5% CO2.2.5. GLP-1 secretion
STC-1 cells were seeded in 24-well plates at a concentration of 20000 cells per cm2 and were grown at 37 °C under 5% CO2 until confluence. On the day of the experiment, cells were washed in PBS and incubated with different digested samples diluted to the chosen concentration in HEPES buffer (4.5 mM KCl, 1.2 mM CaCl2, 140 mM NaCl, and 20 mM Hepes, pH 7.4) for 2 hours. Supernatants were recovered and centrifuged at 1200 rpm for 5 minutes to eliminate cellular debris. Then, the supernatants were frozen at −80 °C until the day of analysis. Secreted GLP-1 was measured using the Glucagon-Like Peptide-17–36 Amide EIA kit, EK-028-11CE (Phoenix Pharmaceuticals, Burlingame, USA). The viability of the cells (>95%) was verified for each condition using the cell counting kit-8 (CCK-8, Tebu-Bio, Le Perray en Yvelines, France).
2.6. DPP-IV activity determination
To measure the intestinal DPP-IV activity before the IB passage, the method using Caco-2 cells described by Caron et al. was used with minor modifications.28 Concisely, cells were seeded in black 96-well plates at 25000 cells per cm2. On the day of the experiment, Caco-2 cells were washed with pH 7.4 PBS. To measure the inhibition of epithelial cell DPP-IV activity, cells were incubated with 125 μL of digested samples diluted in PBS at four different concentrations and 50 μL of Gly-Pro-AMC fluorescent substrate at a final concentration of 1 mM. The viability of the cells (>95%) was verified for each condition using the CCK-8 assay (Tebu-Bio, France).
To measure the DPP-IV activity without cells, human recombinant DPP-IV was used (Sigma-Aldrich). Digested samples (100 μL) diluted in PBS at four different concentrations were incubated with 25 μL of recombinant human DPP-IV enzyme at 0.018 U mL−1 and 50 μL of fluorescent DPP-IV substrate, Gly-Pro-AMC at a final concentration of 1 mM.
To measure the DPP-IV activity of dWPH after IB passage, 100 μL of the recovered basal medium were incubated with the recombinant enzyme and the fluorescent substrate under the same conditions as mentioned before.
In each protocol, positive control was performed containing only enzyme and substrate in the absence of samples. The fluorescence (360 nm excitation, 438 nm emission) was recorded for 1 hour at 37 °C on a Xenius-XC spectrofluorometer (Xenius Safas, Monaco). Slopes were calculated and used to determine the IC50 of the digested protein samples.
2.7. Intestinal barrier (IB) passage experiment
The model described by Fleury et al. with minor adaptations was used for the analysis of the IB passage,29 a recognized IB model using cocultured intestinal epithelial absorptive (Caco-2) and goblet (HT29-MTX) differentiated cells.30,31In brief, Caco-2 and HT29-MTX cells were seeded at a ratio of 90/10 and a density of 20000 cells per cm2 on inserts (microporous PET membrane, 3 μm pore size, Corning, Glendale, CA, USA) and grown for 21 days to ensure good cell differentiation. On the day of the experiment, the differentiated cell monolayer was washed with PBS, and 500 μL of the digested samples were diluted in DMEM without FBS, at 10 and 31.25 mg mL−1, and placed in an apical compartment, while 1.5 mL of DMEM without FBS was added in the basolateral compartment. After two hours of incubation at 37 °C under 5% CO2, the basolateral media was recovered and frozen at −20 °C until analysis. The viability of the cells (>95%) was verified for each condition using the CCK-8 assay (Tebu-Bio, France). The IB integrity was evaluated on the day of the experiment at 0 and 120 minutes by measuring the transepithelial electrical resistance (TEER) with a Voltmeter Ohmmeter MilliCell Electrical Resistance System (Merck-Millipore Burlington, MA, USA).
The apparent paracellular permeability was measured using Lucifer Yellow (LY) just after the experiment. LY (1 mL) diluted at 100 μM in HBSS was added in the apical compartment, whereas 2 mL Hank's buffer containing HEPES were added in the basolateral compartment. Caco-2/HT29-MTX monolayer was then incubated for 90 minutes and 100 μL of the basolateral media were collected at 15, 30, 45, 60, and 90 minutes. The luminescence was read with a Xenius-XC spectrophotometer (Safas) at 530 nm (excitation) and 585 nm (emission). The apparent permeability coefficient (Papp) was calculated according to the following equation.
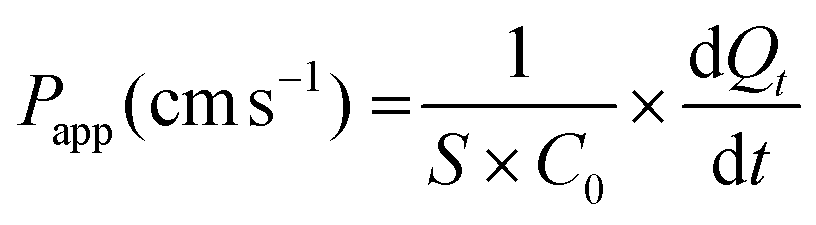
2.8. GCG and PCSK1 gene expression
STC-1 cells were seeded at a density of 20000 cells per cm2 in 24-well plates and grown to confluence in complete DMEM at 37 °C under 5% CO2. On the day of the experiment, cells were washed in PBS and incubated with digested samples at 10 mg mL−1 for 4 and 24 hours in DMEM medium without FBS. The viability of the cells (>95%) was verified for each condition using the CCK-8 assay (Tebu-Bio, France). At the end of the incubation, the supernatant was removed, and the RNAs were extracted using the modified NucleoZOL protocol (Macherey-Nagel, Düren, Germany). The concentration and purity of the RNAs were determined using a Nanodrop lite spectrophotometer (ThermoFisher Scientific, Waltham, MA, USA). RNAs were retrotranscribed into cDNAs using the RevertAid H Minus First Strand cDNA Synthesis kit (ThermoFisher Scientific). Quantitative PCR experiments were performed with the Takyon™ No Rox SYBR® MasterMix dTTP Blue kit (Eurogentec, Seraing, Belgium) on the CFX connect Real-Time PCR detection system (Biorad, Hercules, Ca, USA). The specific primers (Eurogentec) of the different genes studied in this study are presented in Table 1.
| Species | Genes | Forward primer | Reverse primer |
|---|---|---|---|
| Mus musculus | PCSK1 | GCCGAAGAACTGGGGTATGA | CCCACGTCACACGATCATCA |
| GCG | AGAGACATGCTGAAGGGACC | CTTTCACCAGCCACGCAATG | |
| HPRT1 | CAGCGTCGTGATTAGCGATGA | CGAGCAAGTCTTTCAGTCCTGT |
2.9. Statistical analysis
Data were expressed as mean values with their standard deviations. Statistical analyses were carried out using GraphPad Prism. The experiments including two groups were analyzed with a t-test or a Mann–Whitney test. The experiments including more than two group were analyzed with a one-way ANOVA, followed by a Turkey or a Dunnett test when the values follow a normal distribution. A Kruskal–Wallis test with a Dunn test was performed when the values did not follow a normal distribution.3. Results
3.1. dWPH and dWPI peptide molecular mass profiles are different after SGID
Whey protein isolate and hydrolysate before and after SGID were subjected to size exclusion chromatography to compare their peptide molecular mass distribution (Fig. 1A and B).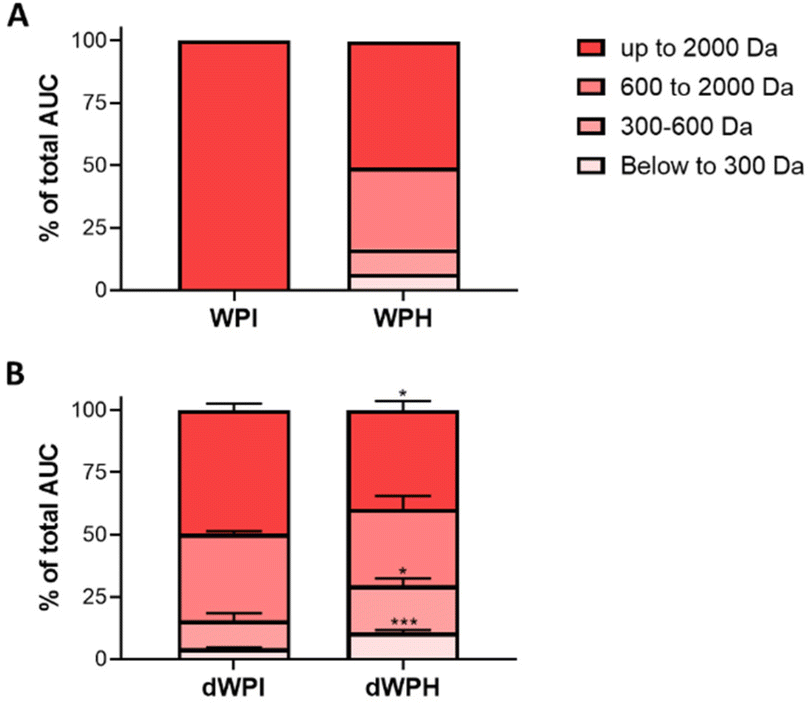 | ||
| Fig. 1 WPI and WPH peptide molecular mass distribution before and after SGID. The peptide molecular mass profiles of the whey protein isolate (WPI) and the digested whey protein hydrolysate (WPH) before (A) and after (B) digestion (dWPI and dWPH) were obtained by size exclusion chromatography-fast protein liquid chromatography. The apparent molecular weight distribution, expressed as a percentage of the total area under the curve (AUC), was calculated from the linear regression relationship, which correlates the log of standard molecular mass molecules and their elution volume. The SGID was performed in three replicates (N = 3) and the peptide molecular mass dispersion of dWPI and dWPH was compared in each class of molecular mass by an unpaired T-Test (*p < 0.05, **p < 0.001, ***p < 0.0001). | ||
The molecular mass distribution before digestion confirms the presence of unhydrolyzed proteins in the WPI (data not shown) conversely to WPH, which presented a half of the peptide with an estimated molecular mass lower than 2000 Da.
The molecular profiles of the dWPH and the dWPI were significantly different after SGID. Indeed, the digested whey hydrolysate was significantly more hydrolyzed after SGID as it contained more than a 2-fold higher proportion of low molecular mass peptides (<600 Da) and more than a 1.5-fold lower proportion of high molecular mass peptides (>2000 Da).
3.2. dWPH stimulates GLP-1 secretion dose-dependently and is more potent than dWPI
STC-1 enteroendocrine cells were first incubated with increasing concentrations of dWPH at 2, 5, and 10 mg mL−1. A dose-dependent secretion of GLP-1 was observed (Fig. 2A). | ||
| Fig. 2 dWPH and dWPI effects on GLP-1 secretion in STC-1 cells. STC-1 enteroendocrine cells were incubated for 2 hours with increasing concentrations of dWPH (A). STC-1 cells were incubated for 2 hours with dWPI and dWPH at 10 mg mL−1 (B). The amount of GLP-1 was determined using an EIA kit and results are expressed as a fold of the Blk SGID mean, which is set at 1. Results are the mean ± SD of 3 independent experiments (N = 3). After a normality and log normality test, a Kruskal–Wallis followed by a Dunn's test (A) and a one-way ANOVA followed by a Tukey's test (B) were performed. (**p < 0.01, ****p < 0.0001). | ||
Thus, at 2 mg mL−1, the induction of GLP-1 secretion was low and not significant, whereas GLP-1 secretion increased significantly at 5 and 10 mg mL−1 and was respectively 2.77 and 8.79 fold higher than the one obtained with the Blk SGID. We subsequently compared the effect of digested whey protein isolate to the digested whey protein hydrolysate on GLP-1 secretion at 10 mg mL−1 (Fig. 2B). The GLP-1 amount recovered in the cell supernatants after 2 hours of contact with dWPH was significantly 2.1- and 3.8-fold higher than with dWPI and blK SGID, respectively.
3.3. dWPI and dWPH inhibit DPP-IV activity in vitro and in situ in intestinal epithelial cells
The ability of dWPI and dWPH to inhibit DPP-IV activity was compared in vitro using human recombinant DPP-IV and in situ in Caco-2 live cells. The digested samples were tested at four different concentrations to characterize the relationship between the DPP-IV activity inhibition (% of control value) and the digested sample concentration, permitting the calculation of the concentration for which the enzyme activity was reduced by 50% when compared to the control (IC50).No significant differences were observed between the IC50 values obtained with dWPI and dWPH both in vitro and in situ (Table 2).
| DPP-IV origin | dWPI – IC50 (mg mL−1) | dWPH – IC50 (mg mL−1) |
|---|---|---|
| Inhibitory concentration inducing 50% DPP-IV activity inhibition (IC50) of the dWPI and dWPH digests. Values are reported as the mean from triplicate determinations with SD. | ||
| Caco-2 | 1.36 ± 0.55 | 1.49 ± 0.46 |
| Recombinant | 1.31 ± 0.49 | 1.28 ± 0.38 |
3.4. Digested whey protein hydrolysate inhibits DPP-IV activity after intestinal barrier passage
To know if the pool of peptides recovered after the IB passage was able to inhibit the DPP-IV activity, we used a recognized IB model using cocultured of Caco-2 and HT29-MTX. The ability of the basolateral media recovered after the permeation of dWPH at 10 and 31.25 mg mL−1 and Blk SGID at the same dilutions was assessed. When cells were incubated with 10 mg mL−1 dWPH in the apical compartment, the basolateral media tend to inhibit DPP-IV activity but not significantly (p = 0.0745) when compared to the blk SGID. When tested at 31.25 mg mL−1, the recovered basolateral media of dWPH significantly decreased (p = 0.0007) the DPP-IV activity by 3.8% when compared to the blk SGID basolateral media (Fig. 3A). | ||
| Fig. 3 DPP-IV inhibition activity of dWPH after in vitro IB passage. A co-culture of Caco-2/HT29-MTX cells was grown on inserts for 3 weeks. Cell monolayers were incubated with dWPH or blk SGID at 10 and 31.25 mg mL−1 in the apical compartment for 2 hours. DPP-IV activity inhibition of the basolateral media was then measured in vitro using recombinant DPP-IV. Results are the mean ± SD of 3 independent experiments (N = 3) (A). ΔTEER is expressed as the percentage of variation of the TEER (Ω cm−2) after 2 hours-incubation with dWPH and blank SGID (B). The Papp (cm s−1) of the cell monolayer was determined using lucifer yellow (C). After a normality and log normality test, a Kruskal–Wallis followed by a Dunn's multiple comparison test (B and C) and a one-way ANOVA followed by a Dunnett's multiple comparison test (A) were performed (***p < 0.001, ****p < 0.0001). | ||
Concomitantly, the effects of dWPH and blk SGID on IB integrity and permeability were evaluated by measuring the TEER and the apparent permeability (Papp), respectively. At 10 mg mL−1, no significant changes were observed compared to the control condition (blank SGID) regarding apparent TEER (Fig. 3B) and permeability (Fig. 3C). At 31.25 mg mL−1, TEER was significantly decreased (Fig. 3B) without a significant impact on the Papp (Fig. 3C).
3.5. dWPH increases the preproglucagon and PC-1 mRNA levels
The effect of dWPH on GCG and PCSK1 gene expression was measured in STC-1 cells at 4 and 24 h. dWPH significantly increased the preproglucagon and PC-1 mRNA levels in enteroendocrine cells at 4 and 24 hours compared to blank SGID (Fig. 4).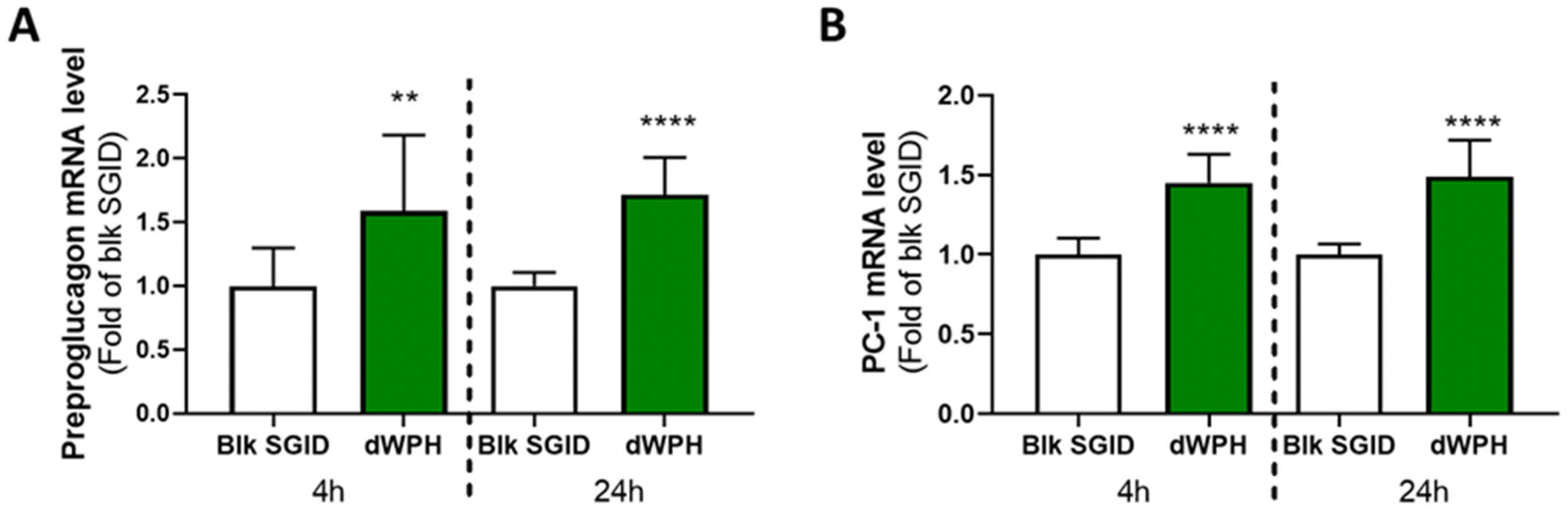 | ||
| Fig. 4 Digested whey protein hydrolysate increased GCG and PCSK1 gene expression. mRNA from STC-1 cells incubated for 4 and 24 h with 10 mg mL−1 of digested dWPH was extracted, retro-transcribed, and quantified by quantitative PCR. Preproglucagon (A) and PC-1 (B) mRNA relative levels were normalized to the housekeeping gene HPRT1. Results are expressed as relative amounts compared to the blank SGID (Blk SGID) condition, which is set at 1. Mean ± SD of 3 independent experiments (N = 3). After a normality and lognormality test, a Mann–Whitney test was performed (**p < 0.001, ****p < 0.0001). | ||
4. Discussion
The key to fighting diabetes and metabolic syndrome is a change in diet, including a reduction in saturated fat and cholesterol and an increase in the consumption of low-fat dairy products, vegetables, and cereals. A high-protein diet has already shown positive results on the satiety and production of hypoglycemic hormones such as insulin.32–34 In addition, a high-protein diet may yield better results than an isoenergetic diet of fat and carbohydrates.35Dairy products are known to be rich in proteins and thus could potentially exert antidiabetic effects. Whey protein preload administration was evidenced to increase GLP-1 response and decrease plasma glucose in women with polycystic ovarian syndrome, which is associated with a higher risk to develop T2D.36 The bioactive effect of dairy proteins is also attributed to the peptides resulting from their hydrolysis. Whey proteins are a good source of peptides and amino acids. After in vitro digestion, a comparative study of the potential antidiabetic effect of whey protein isolate (Promilk 852FB1®) and whey protein hydrolysate (Pep2Dia®) was performed here. Then, a part of the mechanisms by which digested WPH regulates glycemia at the intestinal level was investigated.
The enzymatic hydrolysis used to obtain the whey protein hydrolysate modifies the peptide profile of whey proteins after gastrointestinal digestion (Fig. 1). Indeed, the SGID of whey protein hydrolysate generates a higher proportion of small peptides than that of the whey protein isolate, potentially leading to different bioactivities. It has been shown that the protein source and thus the size and sequence of released peptides and amino acids during digestion were related to their effect on glycemic regulation, particularly on incretin secretion. Indeed, better response in plasma GLP-1 and insulin concentrations was observed in healthy subjects 3 hours after the ingestion of standardized breakfasts containing whey proteins than those containing soy proteins and caseins.37 Another study showed that the digestion of different casein matrices (micellar casein or sodium caseinate) produced different peptide profiles, impacting GLP-1 secretion stimulation in GLUTag cells.38 In humans, it has been shown that hydrolyzed whey increased insulin secretion compared to non-hydrolyzed protein.39 Amino acids, GLP-1, cholecystokinin, and glucose insulinotropic peptide plasma levels were found to be higher over 3 hours after preloaded ingestion of whey proteins compared to caseins.40 Due to the structural and physicochemical properties, caseins and whey proteins are differently digested and respectively considered as “slow” and “fast” proteins as caseins were digested in more than 6 hours and whey proteins in 2 hours in humans. These differences in digestive behavior can lead to different metabolic roles, particularly in the stimulation of protein synthesis. Indeed, the faster release in a higher proportion of amino acids during the digestion of whey proteins affect the appetite and coincide with a greater intestinal hormonal secretion and a stronger insulin response.41–43 Our results are in line with the aforementioned observations as the dWPH was able to induce GLP-1 secretion (Fig. 2A) and enhanced GLP-1 secretion by STC-1 cells compared to dWPI (Fig. 2B). Prehydrolysis of whey proteins increases and potentiates their effect on GLP-1 secretion after SGID. The different molecular mass distributions of dWPH and dWPI confirm the importance of the peptidome nature in intestinal peptide sensing, leading to GLP-1 secretion, as previously described.15,17
Recently, we showed that the origin of the proteins led to different gastrointestinal-resistant peptidomes notably as a result of their native sequence, digestibility, and conformation. These differences impacted the inhibitory potential of the DPP-IV activity in vitro and in situ at the intestinal level but also in vivo in rat plasma.29 In contrast, in the present study, the prehydrolysis of whey proteins did not modify the DPP-IV activity inhibitory effect in vitro and in situ at the intestinal level before the passage of the intestinal barrier. This is in keeping with the results obtained by Nongonierma et al., which evidenced the positive impact of the SGID on the DPP-IV inhibitory activity of a whey protein hydrolysate, for which the IC50 measured in vitro after SGID was closed (1.02 ± 0.05 mg mL−1) of the value obtained here for dWPH (1.28 ± 0.38 mg mL−1).44 The IC50 values obtained here for the digested whey protein isolate (1.36 ± 0.55 mg mL−1) with the intestinal cell-based in situ method are comparable with those we obtained (1.64 ± 0.11 mg mL−1) for dWPI in a precedent study and slightly higher than those obtained by Santos-Hernández et al. (1.17 ± 0.09 mg mL−1) for a digested whey protein concentrate.45
In the present work, the DPP-IV inhibitory activity of the dWPH was recovered after the in vitro IB passage (Fig. 3A). We previously observed in similar conditions that it was not the case for in vitro digested whey protein isolate inversely that for caseins, hemoglobin, and ovalbumin.26 Compared to dWPI, dWPH may contain free amino acids and low MW peptides that can facilitate passage through the intestinal barrier. The small peptides in dWPH, such as AP dipeptides, can pass the intestinal barrier via intracellular junctions, unlike larger peptides that can interact with membrane receptors of epithelial cells or transporters.46–48 Thus, whey protein prehydrolysis, followed by SGID, would allow the production of peptides with a better chance of being found in the blood compartment and thus increase their bioactive roles. This has been shown, for example, in insulin secretion: the insulinotropic effect of hydrolysates is correlated to a better absorption rate of amino acids and small peptides at the intestinal level.49
In addition, TEER and the apparent permeability of the co-culture monolayer were measured to investigate the effect of dWPH on intestinal barrier permeability and integrity. dWPH decreases TEER and increases apparent permeability after 2 hours of incubation at a concentration of 31.25 mg mL−1 but has no effect at 10 mg mL−1 (Fig. 3B and C). This is in agreement with our already published result, which showed that a digested whey protein isolate decreased the TEER of Caco-2/HT-29MTX monolayer.29 The apparent permeability coefficient calculated with 31.25 mg mL−1 of dWPH remained acceptable (9.2 × 10−7 ± 6.2 × 10−7 cm s−1). In the literature, the accepted apparent permeability coefficient for a monolayer of Caco-2 cells is 1 × 10−6 cm s−1.50 In our study, a co-culture involving HT29-MTX cells was used and could therefore explain the small decrease in this coefficient. Digested whey and infant milk formula containing digested whey proteins can affect the expression of tight junction proteins, which are involved in passive diffusion.29,51 Taken together, results on the peptide size and dWPH effect on membrane permeability suggest that peptides cross the epithelial membrane through passive diffusion, without excluding the possible involvement of peptide transporters also known to be involved in peptide uptake.46
Furthermore, in addition to GLP-1 secretion stimulation and DPP-IV activity inhibition, dWPH increased in STC-1 cells, the expression of gene coding for preproglucagon and PC-1 (Fig. 4). It has already been shown that the regulation of glucose homeostasis by the peptides generated during digestion could also involve gene expression upregulation52,53 but only for GCG. The expression of GLP-1 is due to posttranslational modifications involving tissue-specific enzymes, i.e., PC-1 in the intestine.54 In our study, we show that the induction of GLP-1 level could also be mediated by the increase in PC-1 expression, contrary to what had been shown with peptides derived from the hemoglobin-intestinal digest.52
5. Conclusion
This study evidenced in vitro that the prehydrolysis of whey proteins, followed by gastrointestinal digestion, allows the release of amino acids and small peptides involved in regulating glycemia at different levels. Indeed, the secretion of GLP-1 and the expression of GCG and PCSK1 genes in enteroendocrine cells were increased. The digested WPH exerted a good DPP-iV inhibitory activity at the intestinal level and after intestinal barrier passage. These results taken together with those obtained with the same WPH, which evidenced the reduction of postprandial glycemia in prediabetic individuals, confirm the good potential of this whey protein hydrolysate in preventing T2D.Abbreviations
| DPP-IV | Dipeptidyl peptidase-4 |
| dWPH | Digested whey protein hydrolysate |
| dWPI | Digested whey protein isolate |
| EECs | Enteroendocrine cells |
| GCG | Glucagon |
| GLP-1 | Glucagon-like peptide 1 |
| IB | Intestinal barrier |
| SGID | Static in vitro gastrointestinal digestion |
| T2D | Type 2 diabetes |
Author contributions
MT, CD, JM, JA, AlB, AuB, RR, and BC contributed to the study design and interpretation of the data. MT, performed the experiments. MT, CD, JM, BC wrote and edited the manuscript. All authors read and approved the submission of the manuscript.Conflicts of interest
The authors declare no competing financial interest.Acknowledgements
This research was funded in the framework of the joint laboratory project (Allinpep) between the UMRT BioEcoAgro and Ingredia funded by the Hauts-de-France region (FEDER) and in the Alibiotech research program, which is financed by the European Union, the French State, and the French Region of Hauts-de-France.References
- R. A. DeFronzo, E. Ferrannini, L. Groop, R. R. Henry, W. H. Herman and J. J. Holst, et al., Type 2 diabetes mellitus, Nat. Rev. Dis. Primer, 2015, 1(1), 1–22 CrossRef PubMed.
- S. Yuan and S. C. Larsson, An atlas on risk factors for type 2 diabetes: a wide-angled Mendelian randomisation study, Diabetologia, 2020, 63(11), 2359–2371 CrossRef CAS PubMed.
- B. J. Goldstein, Insulin resistance as the core defect in type 2 diabetes mellitus, Am. J. Cardiol., 2002, 90(5 Supplement 1), 3–10 CrossRef PubMed.
- J. J. Marín-Peñalver, I. Martín-Timón, C. Sevillano-Collantes and F. J. del Cañizo-Gómez, Update on the treatment of type 2 diabetes mellitus, World J. Diabetes, 2016, 7(17), 354–395 CrossRef PubMed.
- T. D. Müller, B. Finan, S. R. Bloom, D. D'Alessio, D. J. Drucker and P. R. Flatt, et al., Glucagon-like peptide 1 (GLP-1), Mol. Metab., 2019, 30, 72–130 CrossRef PubMed.
- B. A. McLean, C. K. Wong, J. E. Campbell, D. J. Hodson, S. Trapp and D. J. Drucker, Revisiting the Complexity of GLP-1 Action from Sites of Synthesis to Receptor Activation, Endocr. Rev., 2021, 42(2), 101–132 CrossRef PubMed.
- T. Nargis and P. Chakrabarti, Significance of circulatory DPP4 activity in metabolic diseases, IUBMB Life, 2018, 70(2), 112–119 CrossRef CAS PubMed.
- J. J. Holst and C. F. Deacon, Glucagon-like peptide-1 mediates the therapeutic actions of DPP-IV inhibitors, Diabetologia, 2005, 48(4), 612–615 CrossRef CAS PubMed.
- A. H. Frid, M. Nilsson, J. J. Holst and I. M. E. Björck, Effect of whey on blood glucose and insulin responses to composite breakfast and lunch meals in type 2 diabetic subjects, Am. J. Clin. Nutr., 2005, 82(1), 69–75 CrossRef CAS PubMed.
- D. Jakubowicz, J. Wainstein, Z. Landau, B. Ahren, M. Barnea and Y. Bar-Dayan, et al., High-energy breakfast based on whey protein reduces body weight, postprandial glycemia and HbA1C in Type 2 diabetes, J. Nutr. Biochem., 2017, 49, 1–7 CrossRef CAS PubMed.
- M. Nilsson, J. J. Holst and I. M. Björck, Metabolic effects of amino acid mixtures and whey protein in healthy subjects: studies using glucose-equivalent drinks, Am. J. Clin. Nutr., 2007, 85(4), 996–1004 CrossRef CAS PubMed.
- B. L. Petersen, L. S. Ward, E. D. Bastian, A. L. Jenkins, J. Campbell and V. Vuksan, A whey protein supplement decreases post-prandial glycemia, Nutr. J., 2009, 8(1), 47 CrossRef PubMed.
- J. Caron, D. Domenger, P. Dhulster, R. Ravallec and B. Cudennec, Protein Digestion-Derived Peptides and the Peripheral Regulation of Food Intake, Front. Endocrinol., 2017, 8, 85 CrossRef PubMed.
- M. Santos-Hernández, L. Amigo and I. Recio, Induction of CCK and GLP-1 release in enteroendocrine cells by egg white peptides generated during gastrointestinal digestion, Food Chem., 2020, 329, 127188 CrossRef PubMed.
- J. Caron, B. Cudennec, D. Domenger, Y. Belguesmia, C. Flahaut and M. Kouach, et al., Simulated GI digestion of dietary protein: Release of new bioactive peptides involved in gut hormone secretion, Food Res. Int., 2016, 89, 382–390 CrossRef CAS PubMed.
- B. Cudennec, R. Balti, R. Ravallec, J. Caron, A. Bougatef and P. Dhulster, et al., In vitro evidence for gut hormone stimulation release and dipeptidyl-peptidase IV inhibitory activity of protein hydrolysate obtained from cuttlefish (Sepia officinalis) viscera, Food Res. Int., 2015, 78, 238–245 CrossRef CAS PubMed.
- C. Dugardin, B. Cudennec, M. Tourret, J. Caron, L. Guérin-Deremaux and J. Behra-Miellet, et al., Explorative Screening of Bioactivities Generated by Plant-Based Proteins after In Vitro Static Gastrointestinal Digestion, Nutrients, 2020, 12(12), 3746 CrossRef CAS PubMed.
- S. Theysgeur, B. Cudennec, B. Deracinois, C. Perrin, I. Guiller and A. Lepoudère, et al. New Bioactive Peptides Identified from a Tilapia Byproduct Hydrolysate Exerting Effects on DPP-IV Activity and Intestinal Hormones Regulation after Canine Gastrointestinal Simulated Digestion, Molecules, 2020, 26(1), 136 CrossRef PubMed.
- Y. Song, J. A. Koehler, L. L. Baggio, A. C. Powers, D. A. Sandoval and D. J. Drucker, Gut-Proglucagon-Derived Peptides Are Essential for Regulating Glucose Homeostasis in Mice, Cell Metab., 2019, 30(5), 976–986 CrossRef CAS PubMed.
- D. A. Sandoval and D. A. D'Alessio, Physiology of proglucagon peptides: role of glucagon and GLP-1 in health and disease, Physiol. Rev., 2015, 95(2), 513–548 CrossRef CAS PubMed.
- J. Caron, D. Domenger, Y. Belguesmia, M. Kouach, J. Lesage and J. F. Goossens, et al., Protein digestion and energy homeostasis: How generated peptides may impact intestinal hormones?, Food Res. Int., 2016, 88, 310–318 CrossRef CAS.
- M. Cordier-Bussat, C. Bernard, F. Levenez, N. Klages, B. Laser-Ritz and J. Philippe, et al. Peptones stimulate both the secretion of the incretin hormone glucagon-like peptide 1 and the transcription of the proglucagon gene, Diabetes, 1998, 47(7), 1038–1045 CrossRef CAS PubMed.
- T. Sartorius, A. Weidner, T. Dharsono, A. Boulier, M. Wilhelm and C. Schön, Postprandial Effects of a Proprietary Milk Protein Hydrolysate Containing Bioactive Peptides in Prediabetic Subjects, Nutrients, 2019, 11(7), 1700 CrossRef CAS PubMed.
- R. Boutrou, C. Gaudichon, D. Dupont, J. Jardin, G. Airinei and A. Marsset-Baglieri, et al., Sequential release of milk protein-derived bioactive peptides in the jejunum in healthy humans, Am. J. Clin. Nutr., 2013, 97(6), 1314–1323 CrossRef CAS PubMed.
- A. Brodkorb, L. Egger, M. Alminger, P. Alvito, R. Assunção and S. Ballance, et al., INFOGEST static in vitro simulation of gastrointestinal food digestion, Nat. Protoc., 2019, 14(4), 991–1014 CrossRef CAS PubMed.
- N. Atallah, B. Deracinois, A. Boulier, A. Baniel, D. Jouan-Rimbaud Bouveresse and R. Ravallec, et al. In Vitro Assessment of the Impact of Industrial Processes on the Gastrointestinal Digestion of Milk Protein Matrices Using the INFOGEST Protocol, Foods, 2020, 9(11), E1580 CrossRef PubMed.
- C. Dugardin, L. Fleury, V. Touche, F. Ahdach, J. Lesage and M. Tenenbaum, et al., An Exploratory Study of the Role of Dietary Proteins in the Regulation of Intestinal Glucose Absorption, Front. Nutr., 2021, 8, 769773 CrossRef PubMed.
- J. Caron, D. Domenger, P. Dhulster, R. Ravallec and B. Cudennec, Using Caco-2 cells as novel identification tool for food-derived DPP-IV inhibitors, Food Res. Int. Ott. Ont., 2017, 92, 113–118 CrossRef CAS PubMed.
- L. Fleury, B. Deracinois, C. Dugardin, A. B. Nongonierma, R. J. FitzGerald and C. Flahaut, et al. In Vivo and In Vitro Comparison of the DPP-IV Inhibitory Potential of Food Proteins from Different Origins after Gastrointestinal Digestion, Int. J. Mol. Sci., 2022, 23(15), 8365 CrossRef CAS PubMed.
- A. Wikman-Larhed and P. Artursson, Co-cultures of human intestinal goblet (HT29-H) and absorptive (Caco-2) cells for studies of drug and peptide absorption, Eur. J. Pharm. Sci., 1995, 3(3), 171–183 CrossRef CAS.
- A. Ferraretto, M. Bottani, P. De Luca, L. Cornaghi, F. Arnaboldi and M. Maggioni, et al., Morphofunctional properties of a differentiated Caco2/HT-29 co-culture as an in vitro model of human intestinal epithelium, Biosci. Rep., 2018, 38(2), BSR20171497 CrossRef CAS PubMed.
- D. Jakubowicz, O. Froy, J. Wainstein and M. Boaz, Meal timing and composition influence ghrelin levels, appetite scores and weight loss maintenance in overweight and obese adults, Steroids, 2012, 77(4), 323–331 CrossRef CAS PubMed.
- F. B. Stentz, A. Mikhael, O. Kineish, J. Christman and C. Sands, High protein diet leads to prediabetes remission and positive changes in incretins and cardiovascular risk factors, Nutr. Metab. Cardiovasc. Dis., 2021, 31(4), 1227–1237 CrossRef CAS PubMed.
- T. P. Wycherley, M. Noakes, P. M. Clifton, X. Cleanthous, J. B. Keogh and G. D. Brinkworth, A High-Protein Diet With Resistance Exercise Training Improves Weight Loss and Body Composition in Overweight and Obese Patients With Type 2 Diabetes, Diabetes Care, 2010, 33(5), 969–976 CrossRef CAS PubMed.
- D. K. Layman, E. M. Evans, D. Erickson, J. Seyler, J. Weber and D. Bagshaw, et al. A moderate-protein diet produces sustained weight loss and long-term changes in body composition and blood lipids in obese adults, J. Nutr., 2009, 139(3), 514–521 CrossRef CAS PubMed.
- M. Rao, E. L. Zumbro, K. S. Broughton and M. J. LeMieux, Whey protein preload enhances the active GLP-1 response and reduces circulating glucose in women with polycystic ovarian syndrome, Nutr. Res., 2021, 92, 84–98 CrossRef CAS PubMed.
- M. A. B. Veldhorst, A. G. Nieuwenhuizen, A. Hochstenbach-Waelen, A. J. A. H. van Vught, K. R. Westerterp and M. P. K. J. Engelen, et al., Dose-dependent satiating effect of whey relative to casein or soy, Physiol. Behav., 2009, 96(4–5), 675–682 CrossRef CAS PubMed.
- Y. Komatsu, Y. Wada, H. Izumi, T. Shimizu, Y. Takeda and T. Hira, et al., Casein materials show different digestion patterns using an in vitro gastrointestinal model and different release of glucagon-like peptide-1 by enteroendocrine GLUTag cells, Food Chem., 2019, 277, 423–431 CrossRef CAS PubMed.
- M. Morifuji, M. Ishizaka, S. Baba, K. Fukuda, H. Matsumoto and J. Koga, et al. Comparison of Different Sources and Degrees of Hydrolysis of Dietary Protein: Effect on Plasma Amino Acids, Dipeptides, and Insulin Responses in Human Subjects, J. Agric. Food Chem., 2010, 58(15), 8788–8797 CrossRef CAS PubMed.
- W. L. Hall, D. J. Millward, S. J. Long and L. M. Morgan, Casein and whey exert different effects on plasma amino acid profiles, gastrointestinal hormone secretion and appetite, Br. J. Nutr., 2003, 89(2), 239–248 CrossRef CAS PubMed.
- Y. Boirie, M. Dangin, P. Gachon, M. P. Vasson, J. L. Maubois and B. Beaufrère, Slow and fast dietary proteins differently modulate postprandial protein accretion, Proc. Natl. Acad. Sci. U. S. A., 1997, 94(26), 14930–14935 CrossRef CAS PubMed.
- S. Mahé, N. Roos, R. Benamouzig, L. Davin, C. Luengo and L. Gagnon, et al., Gastrojejunal kinetics and the digestion of [15N]beta-lactoglobulin and casein in humans: the influence of the nature and quantity of the protein, Am. J. Clin. Nutr., 1996, 63(4), 546–552 CrossRef PubMed.
- A. I. Mulet-Cabero, A. Torcello-Gómez, S. Saha, A. R. Mackie, P. J. Wilde and A. Brodkorb, Impact of caseins and whey proteins ratio and lipid content on in vitro digestion and ex vivo absorption, Food Chem., 2020, 319, 126514 CrossRef CAS PubMed.
- A. B. Nongonierma and R. J. FitzGerald, Dipeptidyl peptidase IV inhibitory properties of a whey protein hydrolysate: Influence of fractionation, stability to simulated gastrointestinal digestion and food–drug interaction, Int. Dairy J., 2013, 32(1), 33–39 CrossRef CAS.
- M. Santos-Hernández, M. Cermeño, I. Recio and R. J. FitzGerald, In vitro dipeptidyl peptidase IV inhibitory activity and in situ insulinotropic activity of milk and egg white protein digests, Food Funct., 2021, 12(24), 12372–12380 RSC.
- L. Ozorio, C. Mellinger-Silva, L. M. C. Cabral, J. Jardin, G. Boudry and D. Dupont, The Influence of Peptidases in Intestinal Brush Border Membranes on the Absorption of Oligopeptides from Whey Protein Hydrolysate: An Ex Vivo Study Using an Ussing Chamber, Foods, 2020, 9(10), 1415 CrossRef CAS PubMed.
- M. Sánchez-Navarro, J. Garcia, E. Giralt and M. Teixidó, Using peptides to increase transport across the intestinal barrier, Adv. Drug Delivery Rev., 2016, 106, 355–366 CrossRef PubMed.
- M. Satake, M. Enjoh, Y. Nakamura, T. Takano, Y. Kawamura and S. Arai, et al. Transepithelial transport of the bioactive tripeptide, Val-Pro-Pro, in human intestinal Caco-2 cell monolayers, Biosci., Biotechnol., Biochem., 2002, 66(2), 378–384 CrossRef CAS PubMed.
- B. F. Geerts, M. G. J. van Dongen, B. Flameling, M. M. Moerland, M. L. de Kam and A. F. Cohen, et al., Hydrolyzed casein decreases postprandial glucose concentrations in T2DM patients irrespective of leucine content, J. Diet Suppl., 2011, 8(3), 280–292 CrossRef CAS PubMed.
- A. Béduneau, C. Tempesta, S. Fimbel, Y. Pellequer, V. Jannin and F. Demarne, et al., A tunable Caco-2/HT29-MTX co-culture model mimicking variable permeabilities of the human intestine obtained by an original seeding procedure, Eur. J. Pharm. Biopharm., 2014, 87(2), 290–298 CrossRef PubMed.
- S. L. Bavaro, G. Mamone, G. Picariello, M. J. Callanan, Y. Chen and A. Brodkorb, et al., Thermal or membrane processing for Infant Milk Formula: Effects on protein digestion and integrity of the intestinal barrier, Food Chem., 2021, 347, 129019 CrossRef CAS PubMed.
- J. Caron, D. Domenger, Y. Belguesmia, M. Kouach, J. Lesage and J. F. Goossens, et al., Protein digestion and energy homeostasis: How generated peptides may impact intestinal hormones?, Food Res. Int., 2016, 88, 310–318 CrossRef CAS.
- M. Cordier-Bussat, C. Bernard, F. Levenez, N. Klages, B. Laser-Ritz and J. Philippe, et al., Peptones stimulate both the secretion of the incretin hormone glucagon-like peptide 1 and the transcription of the proglucagon gene, Diabetes, 1998, 47(7), 1038–1045 CrossRef CAS PubMed.
- R. Ugleholdt, X. Zhu, C. F. Deacon, C. Ørskov, D. F. Steiner and J. J. Holst, Impaired intestinal proglucagon processing in mice lacking prohormone convertase 1, Endocrinology, 2004, 145(3), 1349–1355 CrossRef CAS PubMed.
| This journal is © The Royal Society of Chemistry 2023 |