
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicenceTryptophan promoted β-defensin-2 expression via the mTOR pathway and its metabolites: kynurenine banding to aryl hydrocarbon receptor in rat intestine
Zhiru Tanga,
Baoshi Shi†
a,
Weizhong Sun†a,
Yulong Yinb,
Qingju Chena,
Taha Mohameda,
Changwen Lua and
Zhihong Sun *a
*a
aLaboratory for Bio-feed and Molecular Nutrition, College of Animal Science and Technology, Southwest University, Chongqing 400715, China. E-mail: sunzh2002cn@aliyun.com
bInstitute of Subtropical Agriculture, The Chinese Academy of Sciences, Changsha 410125, P. R. China
First published on 21st January 2020
Abstract
In this study, we investigated the signalling pathways mediating tryptophan (Trp)-promoted β-defensin-2 (BD-2) expression in rat intestinal mucosa. Sprague Dawley rats were administered with L-Trp and treated with rapamycin (RAPA), 1-methyltryptophan (1-MT), or para-chlorophenyl-amine (PCPA) to inhibit mammalian target of rapamycin (mTOR), indoleamine-2,3-dioxygenase (IDO), or tryptophan hydroxylase (TPH), respectively. The mRNA and protein levels of BD-2 in the jejunal and ileal mucosa of rats increased with administration of L-Trp. Intraperitoneal injection of RAPA significantly decreased the mRNA level of BD-2 and the concentrations of p-mTORC1 and BD-2 in the jejunal and ileal mucosa of rats with administration of L-Trp (P < 0.05). Oral administration of 1-MT decreased the IDO activity and the mRNA and protein levels of BD-2, and increased the concentrations of tumour necrosis factor (TNF-α), interleukin (IL)-17, and IL-22 in the jejunal and ileal mucosa of rats with administration of L-Trp (P < 0.05). Intraperitoneal injection of PCPA decreased the TPH activity and increased the mRNA and protein levels of BD-2, but did not change the concentrations of TNF-α, IL-17, or IL-22 in the jejunal and ileal mucosa of rats with administration of L-Trp. The results indicate the Trp-promoted BD-2 expression in the jejunum and ileum via the mTOR pathway and its metabolites: kynurenine banding to aryl hydrocarbon receptor in rat intestine.
Introduction
Beta-defensin-2 (BD-2) plays a key role in the prevention of intestinal pathogenic microbial infection. The BD-2 expression is induced by infective stimuli, including Gram-negative and Gram-positive bacteria and their components or pro-inflammatory mediators, for example, tumour necrosis factor (TNF-α) and interleukin (IL)-1β.1–3Amino acids (AAs) have been shown to play important roles in immune responses by regulating (1) the production of cytokines, antibodies and other cytotoxic substances;4,5 (2) gene expression, cellular redox state and lymphocyte proliferation; and (3) the activation of B and T lymphocytes, natural killer cells and macrophages. Studies have shown that dietary specific AA supplementation enhances the immune status of animals with malnutrition and infectious diseases, thereby reducing morbidity and mortality.6,7 Notably, tryptophan (Trp) is associated with many important immune functions. Dietary Trp supplementation improves the growth performance of piglets challenged by pathogenic Escherichia coli.8,9 Other studies have shown that Trp is useful in decreasing intestinal permeability and improving the intestinal barrier function.10 Hashimoto et al.11 reported that, Trp controls expression of small intestinal antimicrobial peptides (AMPs).
L-Trp plays important roles in protein synthesis and as a precursor of various bioactive compounds, such as kynurenine (Kyn) and 5-hydroxytryptamine (5-HT).12,13 Thus, L-Trp might promote the expression of BD-2 in the epithelial cells through the following pathways: (1) activating the mTOR pathway via its intracellular receptors thereby promoting the expression of BD-2; (2) promoting the expression of BD via Kyn, which is as a ligand of aryl hydrocarbon receptor (AhR) can promote intestinal epithelial cell secretion of IL-22, IL-17, and other cytokines that enhance the expression of BD-2 in the intestinal epithelial cells via the NF-κB pathway;14–16 or (3) promoting the expression of BD via 5-HT. In this study, Sprague Dawley rats were used to investigate the signalling pathways mediating Trp-promoted BD-2 expression in the rat intestinal mucosa.
Materials and methods
Animal use and care
Specific pathogen-free SD rats were obtained from Chongqing Academy of Chinese Materia Madica (Chongqing, China) at 5 weeks of age. All experimental procedures were approved by the License of Experimental Animals (SYXK 2014-0002) of the Animal Experimentation Ethics Committee of Southwest University, Chongqing, China.Experimental diets and design
The basal diet (Table 1) was formulated according to the AIN-93G requirements reported by the American Institute of Nutrition. The rats were kept individually in pens (0.5 m × 0.5 m × 0.5 m) in a temperature-controlled (25 ± 1.0 °C) and mechanically ventilated room. Feed and water were provided ad libitum.| Ingredients | (%) |
|---|---|
a Providing the following per kg diet: vitamin A, 14![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000 IU; vitamin D, 1500 IU; vitamin E, 120 IU; vitamin K, 5 mg; vitamin B1, 13 mg; vitamin B2, 12 mg; vitamin B6, 12 mg; vitamin B12, 0.022 mg; nicotinic acid, 60 mg; biotin, 0.2 mg; pantothenic acid, 24 mg; folic acid, 6 mg.b Providing the following per kg diet: Mg, 2000 mg; K, 5000 mg; Fe, 120 mg; Zn, 30 mg; Cu, 10 mg; I, 0.5 mg; Mn, 75 mg. 000 IU; vitamin D, 1500 IU; vitamin E, 120 IU; vitamin K, 5 mg; vitamin B1, 13 mg; vitamin B2, 12 mg; vitamin B6, 12 mg; vitamin B12, 0.022 mg; nicotinic acid, 60 mg; biotin, 0.2 mg; pantothenic acid, 24 mg; folic acid, 6 mg.b Providing the following per kg diet: Mg, 2000 mg; K, 5000 mg; Fe, 120 mg; Zn, 30 mg; Cu, 10 mg; I, 0.5 mg; Mn, 75 mg. |
|
| Corn starch | 74.36 |
| Soy-bean oil | 6.00 |
| Sucrose | 13.67 |
| Ferric citrate | 1.00 |
| Calcium hydrophosphate | 2.00 |
| Vitamin premixa | 0.30 |
| Mineral mixtureb | 2.74 |
| Salt | 0.03 |
| Total | 100.00 |
| Composition (measured values) | |
| Digestible energy (MJ kg−1) | 14.68 |
| Crude protein | 0.00 |
:1:1. After anaesthetization, the rats were exsanguinated by severing the jugular vein and carotid artery. The abdominal cavity was then opened, 1–2 g of jejunal and ileal mucosa were scraped with a glass slide, immediately placed in liquid nitrogen and stored at −80 °C.000 rpm for 2 min at 4 °C. The supernatant was filtered using 0.22 nm filter and the filtrates were harvested for Trp and Kyn analysis. The rats were then exsanguinated and mucosal samples from the jejunum and ileum were collected following the procedures used in experiment 1.Chemical analysis
Trp, Kyn, and 5-HT were measured using liquid chromatography (Agilent Technologies, Santa Clara, CA, USA) on a biphenyl column (100 mm × 2.1 mm, Kinetex 2.6 μm, Biphenyl, 100 Å, Phenomenex, CA, USA). The mobile phase comprised 0.2 M zinc acetate, 8.3 mM acetic acid and 2.8% acetonitrile; elution was performed at a flow rate of 1.5 mL min−1, using an elution gradient. The excitation and emission wave lengths for Trp, Kyn, and 5-HT were 254 nm and 404 nm, 365 nm and 480 nm, and 285 nm 480 nm, respectively. The column temperature was 25 °C and the analysis period of each sample was 60 min. Results were expressed as ng of Trp and Kyn and μg of 5-HT per mL.For ELISA, the mucosa were placed in chilled lysis buffer (1.5% Triton X-100, 1× Tris buffered saline, 0.5% deoxycholic acid sodium salt, 0.1% SDS, 1 mM PMSF, and protease inhibitor cocktail). The homogenate was placed on ice for 30 min, and then centrifuged at 10000 × g at 4 °C for 30 min. The supernatant was harvested for ELISA. The concentrations of TPH (PPS039, R&D systems, MN, USA), TNF-α (ab100785; Abcam, Cambridge, MA, USA), IL-17 (ab214028; Abcam), and IL-22 (ab223857; Abcam) in the jejunal and ileal mucosa were determined by rat-specific ELISA kits.
For the western blotting analysis, 100 mg of the jejunal and ileal mucosa was homogenized in 1 mL RIPA buffer (50 mM Tris-base, 1.0 mM EDTA, 150 mM NaCl, 0.1% SDS, 1% Triton X-100, 1% sodium deoxycholate, 1 mM PMSF) and separated by SDS-polyacrylamide gel electrophoresis (SDS-PAGE). The proteins were transferred to a PVDF membrane by the semi-dry transfer method. The PVDF membranes were blocked in a blocking buffer overnight at 4 °C, incubated in blocking buffer with rabbit-anti rat BD-2 (ab178728, 1:1000, Abcam, Cambridge, MA, USA), β-actin (5125S, 1:1000, CST, Danvers, USA), mTORC1 (ab32028; 1:1000, Abcam) or phospho S2448-mTORC1 (ab109268, 1:1000, Abcam), and incubated in blocking buffer with F(ab)2 of goat-anti rabbit Ig (1:2500) labelled with horseradish peroxidase diluted in PBSS. The PVDF membrane was soaked in a chemiluminescent liquid (Millipore). Pictures were taken using a Chemiluminescence Imaging System (Bio-Rad).
To extract total mRNA, 10 mL TRIzol reagent (Invitrogen, Carlsbad, CA) was added to 0.05 g jejunal and ileal mucosa tissue samples and then homogenized on ice. A Revert Aid First Strand cDNA Synthesis Kit (Fermentas Inc., Burlington, Ontario, Canada) was used for the reverse transcription of the extracted mRNA to cDNA. The cDNA was used for Real-time PCR in an ABI 7900 Sequence Detection System (Applied Biosystems, Foster, CA, USA) with the following thermal cycling parameters: 95 °C for 30 s, followed by 38 cycles at 95 °C for 5 s, 55 °C for 20 s, and 72 °C for 20 s. The primers used in the real-time PCR are shown in Table 2. The raw data were analysed using the ‘fit point’ option of the LightCycler® software (version 3.5; Idaho Technology Inc., Salt Lake, UT, USA). The mRNA expression levels of the target gene were normalized to that of glyceraldehyde 3-phosphate dehydrogenase (GAPDH; a housekeeping gene). The relative expression of the target gene was calculated using the 2−ΔΔCt method.25 Analyses were performed in triplicate and mean values were calculated.
| Gene | Product length | Primer sequences (5′ → 3′) | Tm |
|---|---|---|---|
| a Abbreviations: GAPDH: glyceraldehyde-3-phosphate dehydrogenase; BD-2: β-defensin. | |||
| GAPDH | 149 bp | F: GAAGGTCGGAGTGAACGGAT | 65 °C |
| R: CATGGGTAGAATCATACTGGAACA | |||
| BD-2 | 150 bp | F: ACATTGGGCTCCTGCTCA | 61 °C |
| R: CCTGGCTGCTCCTCAGTATT | |||
Statistical analysis
All statistical analyses were conducted using the general linear model procedure in SAS statistical software (SAS Institute Inc., Cary, NC, USA). Because the experiments were performed in a 2 × 2 factorial design, a two-way ANOVA was used to test the effects of Trp concentration, MT, RAPA or PCPA concentration, and Trp × MT, Trp × RAPA and Trp × PCPA concentration interactions. Duncan's multiple range test was performed to identify differences among groups. The significance was set at P < 0.05. All data are presented as means ± SEM.Results
Oral administration of L-Trp increased the serum Trp and Kyn concentrations, the IDO activity (assessed as the kynurenine to tryptophan ratio) and the concentrations of IL-17, IL-22, and TNF-α in the jejunal and ileal mucosa (Table 3; P < 0.05). Oral administration of 1-MT increased the serum L-Trp concentrations, while decrease the serum Kyn and Kyn/Trp concentration, the IDO activity and the jejunal and ileal mucosa of IL-17, IL-22, and TNF-α concentrations (Table 3; P < 0.05). There were significant interactions in the serum Kyn concentration and the IL-17 concentration in the jejunal and ileal mucosa of the rats between the MT and TRP treatments (Table 3; P < 0.05).| MT0 | MT1 | SEM | P-value | |||||
|---|---|---|---|---|---|---|---|---|
| Trp0 | Trp1 | Trp0 | Trp1 | MT | Trp | MT × Trp | ||
| a Data are presented as mean ± SEM (n = 6). Abbreviations: MT0 or MT1, rats were orally administered with 0.5 mL saline containing or no containing 1-methyltryptophan (50 mg kg−1) daily; Trp0 or Trp1, rats were orally administered with 0.5 mL saline containing or no containing L-Trp (260 mg kg−1) daily; Kyn, kynurenine; IDO, indoleamine-2,3-dioxygenase; IL-17, interleukin 17; TNF-α, tumour necrosis factor. | ||||||||
| Serum | ||||||||
| Trp (μg mL−1) | 6.27 | 17.8 | 7.20 | 18.6 | 0.13 | 0.007 | <0.001 | 0.239 |
| Kyn (μg mL−1) | 0.12 | 0.24 | 0.07 | 0.10 | 0.01 | <0.001 | <0.001 | <0.001 |
| Kyn/Trp (%) | 1.79 | 1.29 | 1.24 | 0.54 | 0.04 | <0.001 | <0.001 | <0.001 |
|
||||||||
| Jejunum mucosa (ng g−1 protein) | ||||||||
| IDO | 42.3 | 60.1 | 25.3 | 39.2 | 0.87 | <0.001 | <0.001 | <0.144 |
| IL-17 | 328 | 881 | 316 | 469 | 17.5 | <0.001 | <0.001 | <0.001 |
| IL-22 | 253 | 377 | 182 | 320 | 5.32 | <0.001 | <0.001 | 0.347 |
| TNF-α | 259 | 461 | 147 | 358 | 13.5 | <0.001 | <0.001 | 0.813 |
|
||||||||
| Ileum mucosa (ng g−1 protein) | ||||||||
| IDO | 41.3 | 74.6 | 30.6 | 60.2 | 2.70 | <0.001 | <0.005 | 0.622 |
| IL-17 | 701 | 1362 | 628 | 807 | 31.0 | <0.001 | <0.001 | <0.001 |
| IL-22 | 315 | 356 | 234 | 247 | 5.22 | 0.002 | <0.001 | 0.084 |
| TNF-α | 242 | 432 | 139 | 323 | 8.87 | <0.001 | <0.001 | 0.802 |
As shown in Table 4, oral administration of L-Trp increased the serum Trp and 5-HT concentrations and the TPH activity and the concentrations of IL-17, IL-22, and TNF-α in the jejunal and ileal mucosa (P < 0.05). Intraperitoneal injection of PCPA increased the serum Trp concentration and the IL-17, IL-22, and TNF-α concentration in the jejunal and ileal mucosa, while decreased the serum 5-HT concentration and the of TPH activity in the jejunal and ileal mucosa (Table 4; P < 0.05). The interactions of the serum 5-HT concentration and the TPH activity in the jejunal and ileal mucosa of rats between PCPA and TRP treatments were observed (Table 4; P < 0.05).
| PCPA0 | PCPA1 | SEM | P-value | |||||
|---|---|---|---|---|---|---|---|---|
| Trp0 | Trp1 | Trp0 | Trp1 | PCPA | Trp | PCPA × Trp | ||
| a Data are presented as mean ± SEM (n = 6). Abbreviations: PCPA0 or PCPA1: rats were intraperitoneally injected with 0.5 mL saline containing or no containing para-chlorophenyl-amine (200 mg kg−1) daily; Trp0 or Trp1, rats were orally administered with 0.5 mL saline containing or no containing L-Trp (260 mg kg−1) daily; 5-HT, 5-hydroxytryptamine; IL-17, interleukin 17; TNF-α, tumour necrosis factor; TPH, tryptophan hydroxylase. | ||||||||
| Serum | ||||||||
| Trp (μg mL−1) | 6.28 | 17.4 | 6.94 | 19.1 | 0.19 | <0.001 | <0.001 | 0.312 |
| 5-HT (ng mL−1) | 32.8 | 57.0 | 23.9 | 31.1 | 1.65 | <0.001 | <0.001 | <0.001 |
|
||||||||
| Jejunum mucosa (ng g−1 protein) | ||||||||
| TPH | 48.8 | 96.3 | 40.9 | 68.2 | 1.27 | <0.001 | <0.001 | <0.001 |
| IL-17 | 298 | 842 | 313 | 844 | 19.72 | 0.255 | <0.001 | 0.879 |
| IL-22 | 245 | 372 | 254 | 380 | 8.56 | 0.267 | <0.001 | 0.777 |
| TNF-α | 266 | 468 | 287 | 472 | 15.52 | 0.428 | <0.001 | 0.865 |
|
||||||||
| Ileum mucosa (ng g−1 protein) | ||||||||
| TPH | 39.4 | 59.9 | 32.6 | 45.1 | 1.38 | <0.001 | <0.001 | 0.011 |
| IL-17 | 706 | 1367 | 717 | 1368 | 37.92 | 0.275 | <0.001 | 0.805 |
| IL-22 | 255 | 386 | 264 | 397 | 5.85 | 0.076 | <0.001 | 0.750 |
| TNF-α | 332 | 453 | 339 | 463 | 7.85 | 0.225 | <0.001 | 0.754 |
Oral administration of L-Trp increased the mRNA level of BD-2 in the jejunal and ileal mucosa of the rats (Table 5; P < 0.05). Oral administration of 1-MT decreased the mRNA level of BD-2 in the jejunal and ileal mucosa (Table 5; P < 0.05). Intraperitoneal injection of RAPA decreased the mRNA level of BD-2 in the jejunal and ileal mucosa of rats (Table 5; P < 0.05). Intraperitoneal injection of PCPA did not change the mRNA level of BD-2 in the jejunal and ileal mucosa (Table 5; P < 0.05). Significant interactions were observed in the mRNA level of BD-2 in the jejunal mucosa between the MT and Trp treatments and between the PAPA and Trp treatments (Table 5; P < 0.05).
| Jejunal mucosa | Ileal mucosa | ||
|---|---|---|---|
| a Data are presented as mean ± SEM (n = 6). Abbreviations: BD-2, β-defensin 2; MT0 or MT1, rats were orally administered with 0.5 mL saline containing or no containing 1-methyltryptophan (50 mg kg−1) daily; Trp0 or Trp1, rats were orally administered with 0.5 mL saline containing or no containing L-Trp (260 mg kg−1) daily; RAPA0 or RAPA1: rats were intraperitoneally injected with 0.5 mL saline containing or no containing rapamycin (1.5 mg kg−1) daily; PCPA0 or PCPA1: rats were intraperitoneally injected with 0.5 mL saline containing or no containing para-chlorophenyl-amine (200 mg kg−1) daily. | |||
| MT0 | Trp0 | 0.62 | 1.68 |
| Trp1 | 3.95 | 5.18 | |
| MT1 | Trp0 | 0.30 | 0.85 |
| Trp1 | 1.43 | 3.34 | |
| Pooled SEM | 0.20 | 0.26 | |
| P-value | MT | <0.001 | <0.001 |
| Trp | <0.001 | <0.001 | |
| MT × Trp | <0.001 | 0.067 | |
| RAPA0 | Trp0 | 3.12 | 4.03 |
| Trp1 | 34.3 | 19.9 | |
| RAPA1 | Trp0 | 1.90 | 2.86 |
| Trp1 | 7.19 | 6.51 | |
| Pooled SEM | 1.35 | 0.87 | |
| P-value | RAPA | <0.001 | <0.001 |
| Trp | <0.001 | <0.001 | |
| RAPA × Trp | <0.001 | <0.001 | |
| PCPA0 | Trp0 | 4.53 | 5.08 |
| Trp1 | 20.4 | 20.2 | |
| PCPA1 | Trp0 | 4.35 | 4.97 |
| Trp1 | 18.0 | 19.7 | |
| Pooled SEM | 0.88 | 0.80 | |
| P-value | PCPA | 0.160 | 0.714 |
| Trp | <0.001 | <0.001 | |
| PCPA × Trp | 0.225 | 0.821 | |
Intraperitoneal injection of RAPA decreased the concentrations of BD-2 and p-mTORC1 in the jejunal and ileal mucosa of the rats (Fig. 1; P < 0.05). Oral administration of L-Trp increased the concentrations of BD-2 and p-mTORC1 in the jejunal and ileal mucosa (Fig. 1; P < 0.05). Intraperitoneal injection of RAPA or oral administration of L-Trp did not change the concentrations of total mTORC1 in the jejunal and ileal mucosa (Fig. 1; P > 0.05). There were interactions in the concentrations of BD-2 and p-mTORC1 in the jejunal and ileal mucosa between RAPA and TRP treatments (Fig. 1; P > 0.05). Oral administration of 1-MT decreased the concentration of BD-2 in the jejunal and ileal mucosa (Fig. 2; P < 0.05) and oral administration of L-Trp increased the concentration of BD-2 in the jejunal and ileal mucosa (Fig. 2; P < 0.05). There were interactions in the concentration of BD-2 in the jejunal and ileal mucosa between 1-MT and TRP treatments (Fig. 2; P < 0.05). Intraperitoneal injection of PCPA increased the concentration of BD-2 in the jejunal and ileal mucosa of the rats administered with L-Trp (Fig. 3; P < 0.05). Oral administration of L-Trp increased the concentration of BD-2 in the jejunal and ileal mucosa (Fig. 3; P < 0.05) and there were no significant interaction in the concentration of BD-2 in the jejunal and ileal mucosa between 1-MT and TRP treatments (Fig. 3; P > 0.05).
 | ||
| Fig. 1 Oral L-Trp affects the concentrations of BD-2, p-mTOR, and mTOR in the jejunal and ileal mucosa of the rats. Data are presented as mean ± SEM (n = 6). a,b,cValues with different letter superscripts within the same index mean significant difference (P < 0.05). Abbreviations: CON, rats were orally administered with 0.5 mL saline and intraperitoneally injected with 0.5 mL saline daily; RAPA, rats were orally administered with 0.5 mL saline and intraperitoneally injected 0.5 mL saline containing rapamycin (1.5 mg kg−1) daily; TRP, rats were orally administered with 0.5 mL saline containing L-Trp (260 mg kg−1) and intraperitoneally injected with 0.5 mL saline daily; RAPA + TRP, rats were orally administered with 0.5 mL saline containing L-Trp (260 mg kg−1) and intraperitoneally injected with 0.5 mL saline containing rapamycin (1.5 mg kg−1) daily; BD-2, β-defensin-2; p-mTOR, phosphorylated mammalian target of rapamycin. | ||
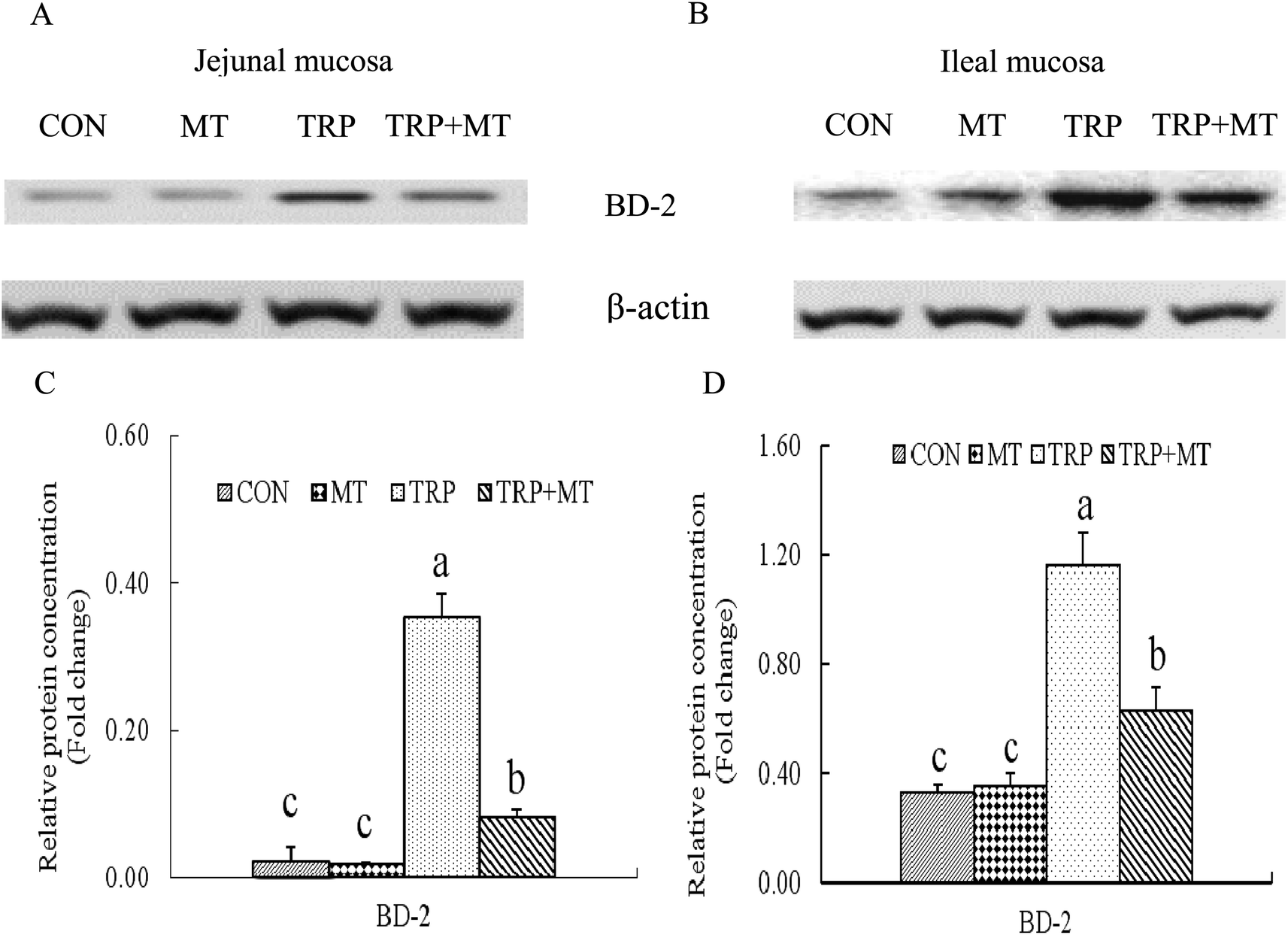 | ||
| Fig. 2 Oral L-Trp affects the concentrations of BD-2 in the jejunal and ileal mucosa of rats. Data are presented as mean ± SEM (n = 6). a,b,cValues with different letter superscripts within the same index mean significant difference (P < 0.05). Abbreviations: CON, rats were orally administered with 0.5 mL saline daily; MT, rats were orally administered with 0.5 mL saline containing 1-MT (50 mg kg−1) daily; TRP, rats were orally administered with 0.5 mL saline containing L-Trp (260 mg kg−1) daily; TRP + MT, rats were orally administered with 0.5 mL saline containing 1-MT (50 mg kg−1 day−1) and L-Trp (260 mg kg−1) daily; BD-2, β-defensin-2; 1-MT, 1-methyltryptophan. | ||
 | ||
| Fig. 3 Oral L-Trp affects the concentrations of BD-2 in the jejunum and ileum mucosa of rats. Data are presented as mean ± SD (n = 6). a,b,cValues with different letter superscripts within the same index mean significant difference (P < 0.05). Abbreviations: CON: rats were orally administered with 0.5 mL saline and intraperitoneally injected with 0.5 mL saline daily; PCPA: rats were orally administered with 0.5 mL saline and intraperitoneally injected with 0.5 mL saline containing PCPA (200 mg kg−1) daily; TRP, rats were orally administered with 0.5 mL saline containing L-Trp (260 mg kg−1) and intraperitoneally injected with 0.5 mL saline daily; PCPA + TRP: rats were orally administered with 0.5 mL saline containing L-Trp (260 mg kg−1) and intraperitoneally injected with 0.5 mL saline containing PCPA (200 mg kg−1) daily; BD-2, β-defensin-2; PCPA, para-chlorophenyl-amine. | ||
Discussion
BD-2 is an inducible AMP that is present in various epithelia. AAs play important roles in epithelial BD expression.26,27 Dietary Trp controls the expression of small intestinal AMPs.11 However, information about the effects of Trp on the expression of BD-2 and its relative mechanisms is limited. In the present study, intraperitoneal injection with RAPA decreased the concentration of p-mTORC1 and the mRNA and protein level of BD-2 in the jejunum and ileum of rats. The results indicate that Trp-promoted BD-2 expression in the intestinal mucosa of rats is related to the mTOR signal pathway. Although AAs are the most potent activator of mTOR,28 the mechanism by which mTOR senses AA signalling is not well understood. However, it is clear that the Rag guanosine triphosphatases (GTPases) and the Ras-homolog enriched in the brain have necessary but distinct roles.29–32 L-Trp is the precursor in two important metabolic pathways: 5-HT synthesis and Kyn synthesis. 5-HT is synthesized from L-Trp by TPH and aromatic AA decarboxylase. TPH exists in the gastrointestinal tract,33 and aromatic AA decarboxylase exists in the small intestine,34 appendix,34 and liver.35 Kyn is synthesized by IDO from L-Trp, which accounts for 90% of Trp catabolism.36 Theoretically, Trp exerts its effects on intestinal AMPs by Kyn or through 5-HT. The possible pathways in which it acts need to be explored.This study demonstrated that the rats administered with L-Trp have a higher serum Kyn and Kyn/Trp ratio, which reflects IDO activity.37,38 The metabolites of Trp, mainly Kyn and kynurenic acid, all have AhR–ligand activity.39 The process that produces these metabolites is initiated and controlled by IDO.38 AhR in combination with these ligands can regulate expression of downstream target genes.16 In this study, with the increase of dietary Trp, serum Kyn concentration, the mRNA and protein level of BD-2, the concentrations of IL-17, IL-22, and TNF-α in the jejunal and ileal mucosa were increased. IL-17, IL-22, and TNF-α are important pro-inflammatory factors. Pro-inflammatory cytokines, such as IL-17 and TNF-α, can induce the expression of BD-2 in epithelial cells.2,14 IL-17 mediates its effect on BD-2 mostly primarily through a Janus kinase and nuclear factor kappa-B signalling events.14 We expected there to be two-way regulation of inflammatory cytokines and BD-2. On one hand, the increase of inflammatory factor levels stimulates BD-2 expression. On the other hand, the increase in BD-2 expression reduces the release of inflammatory cytokines. This provides a reasonable explanation for the decrease in the concentrations of IL-17, IL-22, and TNF-α in the jejunal and ileal mucosa of rats administered with L-Trp with increasing BD-2. Oral administration of 1-MT decreased the IDO activity in the jejunal and ileal mucosa, and serum Kyn concentration and consequently reduced the mRNA and protein level of BD-2 and decreased the concentrations of IL-17, IL-22, and TNF-α in the jejunal and ileal mucosa of the rats. The results indicate that Kyn mediates the signalling pathways of Trp-promoted BD-2 expression in the intestinal mucosa of rats.
Intraperitoneal injection with PCPA decreased the serum 5-HT concentration and the TPH activity in the mucosa of jejunum and ileum. 5-HT is synthesized in enterochromaffin cells by the rate-limiting enzyme TPH1 and in the brainstem and myenteric plexus neurons by TPH-2.40–43 The decrease in concentration of 5-HT in the serum and the TPH activity suggests that intraperitoneal injection with PCPA was successful. In this study, intraperitoneal injection with PCPA increases the concentration of BD-2 in the jejunal and ileal mucosa of rats fed diets supplemented with L-Trp. In addition, intraperitoneal injection with PCPA did not change the concentrations of IL-17, IL-22 and TNF-α in the mucosa of the jejunum and ileum. These results indicate that Trp-promoted BD-2 expression in the intestinal mucosa of rats is not related to 5-HT.
In conclusion, rats administered L-Trp showed a higher concentration of BD-2 in the mucosa of the jejunum and ileum. Intraperitoneal injection of PAPA decreased the concentrations of p-mTORC1 and BD-2 in the jejunum and ileum mucosa; oral administration of 1-MT decreased the mRNA and protein level of BD-2 in mucosa of the jejunum and ileum; and intraperitoneal injection of PCPA increased the mRNA and protein level of BD-2 in the mucosa of the jejunum and ileum of the rats. Our study indicates the Trp-promoted BD-2 expression in the jejunum and ileum via the mTOR pathway and its metabolites: kynurenine banding to aryl hydrocarbon receptor in rat intestine.
Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors gratefully acknowledge the financial support from the National Natural Science Foundation of China (31772610, 31872370), the Fundamental Research Funds for the Central Universities (XDJK2019B014), the Chongqing Key Innovation Project for Overseas Students (cx2017024), and the China Scholarship Council (201508505170).References
- L. X. Deng, G. X. Wu, Y. Cao, B. Fan, X. Gao and X. H. Tang, The chromosomal protein HMGN2 mediates the LPS-induced expression of b-defensins in mice, Inflammation, 2012, 35, 456–473 CrossRef CAS PubMed.
- J. Harder, U. Meyer Hoffert, L. M. Teran, L. Schwichtenberg, J. Bartels and S. Maune, et al., Mucoid Pseudomonas aeruginosa, TNF-alpha, and IL-1 beta, but not IL-6, induce human beta defensin-2 in respiratory epithelia, Am. J. Respir. Cell Mol. Biol., 2000, 22, 714–721 CrossRef CAS PubMed.
- P. K. Singh, H. P. Jia, K. Wiles, J. Hesselberth, L. Liu and B. A. Conway, et al., J Production of beta-defensins by human airway epithelia, Proc. Natl. Acad. Sci. U. S. A., 1998, 95, 14961–14966 CrossRef CAS PubMed.
- P. Li, Y. L. Yin, D. F. Li, S. W. Kim and G. Y. Wu, Amino acids and immune function, Br. J. Nutr., 2007, 98, 237–252 CrossRef CAS PubMed.
- S. W. Kim, R. D. Mateo, Y. L. Yin and G. Y. Wu, Functional amino acids and fatty acids for enhancing production performance of sows and piglets, Asian-Australas. J. Anim. Sci., 2007, 20, 295–306 CrossRef CAS.
- I. R. Johnsona, R. O. Ball, V. E. Baracos and C. J. Field, Glutamine supplementation influences immune development in the newly weaned piglet, Dev. Comp. Immunol., 2006, 30, 1191–1202 CrossRef PubMed.
- J. B. Ewaschuk, G. K. Murdoch, I. R. Johnson, K. L. Madsen, P. Bosi and C. J. Field, Glutamine supplementation improves intestinal barrier function in a weaned piglet model of Escherichia coli infection, Br. J. Nutr., 2011, 106, 870–877 CrossRef CAS PubMed.
- P. Trevisi, E. Corrent, S. Messori, L. Casini and P. Bosi, Healthy newly weaned pigs require more tryptophan to maximize feed intake if they are susceptible to Escherichia coli K88, Livest. Sci., 2010, 134, 236–238 CrossRef.
- M. Stefano, T. Paolo, S. Aude, P. Davide and B. Paolo, Effect of susceptibility to enterotoxigenic Escherichia coli F4 and of dietary tryptophan on gut microbiota diversity observed in healthy young pigs, Vet. Microbiol., 2013, 162, 173–179 CrossRef PubMed.
- W. Liu, S. Mi, Z. Ruan, J. Li, X. Shu and K. Yao, et al., Dietary tryptophan enhanced the expression of tight junction protein ZO-1 in intestine: Trp enhanced tight junction protein ZO-1 in intestine, J. Food Sci., 2017, 82, 562–567 CrossRef CAS PubMed.
- T. Hashimoto, T. Perlot, A. Rehman, J. Trichereau, H. Ishiguro and M. Paolino, et al., ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation, Nature, 2012, 487, 477–481 CrossRef CAS PubMed.
- N. L. Floc’H and B. Seve, Biological roles of tryptophan and its metabolism: potential implications for pig feeding, Livest. Sci., 2007, 112, 23–32 CrossRef.
- P. J. Kennedy, J. F. Cryan, T. G. Dinan and G. Clarke, Kynurenine pathway metabolism and the microbiota-gut-brain axis, Neuropharmacology, 2017, 112, 399–412 CrossRef CAS PubMed.
- C. Y. Kao, Y. Chen, P. Thai, S. Wachi and F. Huang, et al., IL-17 markedly up-regulates β-defensin-2 expression in human airway epithelium via JAK and NF-κB signaling pathways, J. Immunol., 2004, 173, 3482–3491 CrossRef CAS.
- N. Steubesand, K. Kiehne, G. Brunke, R. Pahl, K. Reiss and K. H. Herzig, et al., The expression of the beta-defensins hBD-2 and hBD-3 is differentially regulated by NF-kappaB and MAPK/AP-1 pathways in an in vitro model of Candida esophagitis, BMC Immunol., 2009, 10, 36 CrossRef PubMed.
- S. J. MacEachern, C. M. Keenan, E. Papakonstantinou, K. A. Sharkey and B. A. Patel, Alterations in melatonin and serotonin signaling in the colonic mucosa of mice with dextran-sodium sulfate induced colitis, Br. J. Pharmacol., 2018, 175, 1535–1547 CrossRef CAS PubMed.
- M. S. Marta, A. George, K. Buranee, B. Michael and M. Michael, Comorbidity between depression and inflammatory bowel disease explained by immune-inflammatory, oxidative, and nitrosative stress; tryptophan catabolite and gut-brain pathways, CNS Spectr., 2016, 21, 184–198 CrossRef PubMed.
- A. Jzcue and F. Powrie, Malnutrition promotes rogue bacteria, Nature, 2012, 487, 437–439 CrossRef PubMed.
- R. Metz, S. Rust, J. B. DuHadaway, M. R. Mautino, D. H. Munn and N. N. Vahanian, et al., IDO inhibits a tryptophan sufficiency signal that stimulates mTOR, Oncoimmunology, 2012, 1, 1460–1468 CrossRef PubMed.
- T. Ganz, Antimicrobial polypeptides in host defense of the respiratory tract, J. Clin. Invest., 2002, 109, 693–697 CrossRef CAS PubMed.
- T. Ganz, Defensins: antimicrobial peptides of innate immunity, Nat. Rev. Immunol., 2003, 3, 710–720 CrossRef CAS PubMed.
- K. Ogawa, T. Hara, M. Shimizu, S. Ninomiya, J. Nagano and H. Sakai, et al., Suppression of azoxymethane- induced colonic preneoplastic lesions in rats by 1-methyltryptophan, an inhibitor of indoleamine 2, 3-dioxygenase, Cancer Sci., 2012, 5, 951–958 CrossRef PubMed.
- P. Zhang, T. Shan, X. Liang, C. Deng and S. Kuang, Mammalian target of rapamycin is essential for cardiomyocyte survival and heart development in mice, Biochem. Biophys. Res. Commun., 2014, 452, 53–59 CrossRef CAS PubMed.
- J. W. Kang, H. I. Cho and S. M. Lee, Melatonin inhibits mTOR-dependent autophagy during liver ischemia/reperfusion, Cell. Physiol. Biochem., 2014, 33, 23–36 CrossRef CAS.
- K. J. Livak and T. W. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method, Methods, 2001, 25, 402–408 CrossRef CAS.
- P. Fehlbaum, M. Rao, M. Zasloff and G. M. Anderson, An essential amino acid induces epithelial beta-defensin expression, Proc. Natl. Acad. Sci. U. S. A., 2000, 97, 12723–12728 CrossRef CAS PubMed.
- X. Mao, S. Qi, B. Yu, Z. Q. Huang, H. Chen and Q. Mao, et al., Dietary l-arginine supplementation enhances porcine β-defensins gene expression in some tissues of weaned pigs, Livest. Sci., 2012, 148, 103–108 CrossRef.
- H. Wang, Y. Ji, G. Y. Wu, K. J. Sun, Y. L. Sun and W. Li, et al., L-tryptophan activates mammalian target of rapamycin and enhances expression of tight junction proteins in intestinal porcine epithelial cells, J. Nutr., 2015, 145, 1156–1162 CrossRef CAS PubMed.
- D. H. Kim, D. D. Sarbassov, S. M. Ali, J. E. King, R. R. Latek and H. Erdjument-Bromage, et al., mTOR interacts with raptor to form a nutrient-sensitive complex that signals to the cell growth machinery, Cell, 2002, 110, 163–175 CrossRef CAS.
- M. Roccio, J. L. Bos and F. J. Zwartkruis, Regulation of the small GTPase Rheb by amino acids, Oncogene, 2006, 25, 657–664 CrossRef CAS.
- E. M. Smith, S. G. Finn, A. R. Tee, G. J. Browne and C. G. Proud, The tuberous sclerosis protein TSC2 is not required for the regulation of the mammalian target of rapamycin by amino acids and certain cellular stresses, J. Biol. Chem., 2005, 280, 18717–18727 CrossRef CAS PubMed.
- S. Menon, C. C. Dibble, G. Talbott, G. Hoxhaj, A. J. Valvezan and H. Takahashi, et al., Spatial control of the TSC complex integrates insulin and nutrient regulation of mTORC1 at the lysosome, Cell, 2014, 156, 771–785 CrossRef CAS.
- P. L. Yu, M. Fujimura, K. Okumiya, M. Kinoshita, H. Hasegawa and M. Fujimiya, Immunohistochemical localization of tryptophan hydroxylase in the human and rat gastrointestinal tracts, J. Comp. Neurol., 1999, 411, 654–665 CrossRef CAS.
- P. Facer, J. M. Polak, B. M. Jaffe and A. G. Pearse, Immunocytochemical demonstration of 5-hydroxytryptamine in gastrointestinal endocrine cells, Histochem. J., 1979, 11, 117–121 CrossRef CAS PubMed.
- L. Kubovcakova, O. Krizanova and R. Kvetnansky, Identification of the aromatic L-amino acid decarboxylase gene expression in various mice tissues and its modulation by immobilization stress in stellate ganglia, Neuroscience, 2004, 126, 375–380 CrossRef CAS PubMed.
- D. M. Richard, M. A. Dawes, C. W. Mathias, A. Acheson and D. M. Dougherty, L-tryptophan: basic metabolic functions, behavioral research and therapeutic indications, Int. J. Tryptophan Res., 2009, 2, 45–60 CAS.
- A. Laich, G. Neurauter, B. Widner and D. Fuchs, More rapid method for simultaneous measurement of tryptophan and kynurenine by HPLC, Clin. Chem., 2002, 48, 579–581 CAS.
- S. Nikolaus, B. Schulte, N. Al-Massad, F. Thieme, D. M. Schulte and J. Bethge, et al., Increased tryptophan metabolism is associated with activity of inflammatory bowel diseases, Gastroenterology, 2017, 153, 1504–1516 CrossRef CAS PubMed.
- W. Wang, L. Huang, J. Y. Jin, S. Jolly, Y. Zang and H. Wu, et al., IDO immune status after chemoradiation may predict survival in lung cancer patients, Cancer Res., 2018, 78, 809–816 CrossRef CAS PubMed.
- F. Cote, E. Thevenot, C. Fligny, Y. Fromes, M. Darmon and M. A. Ripoche, et al., Disruption of the nonneuronal tph1 gene demonstrates the importance of peripheral serotonin in cardiac function, Proc. Natl. Acad. Sci. U. S. A., 2003, 100, 13525–13530 CrossRef CAS PubMed.
- Z. Li, A. Chalazonitis, Y. Y. Huang, J. J. Mann, K. G. Margolis and Q. M. Yang, et al., Essential roles of enteric neuronal serotonin in gastrointestinal motility and the development/survival of enteric dopaminergic neurons, J. Neurosci., 2011, 31, 8998–9009 CrossRef CAS PubMed.
- K. B. Neal, L. J. Parry and J. C. Bornstein, Stain-specific genetics, anatomy and function of enteric neural serotonergic pathways in inbred mice, J. Physiol., 2009, 587, 567–586 CrossRef CAS PubMed.
- D. J. Walther, J. U. Peter, S. Bashammakh, H. Hortnagl, M. Voits and H. Fink, et al., Synthesis of serotonin by a second tryptophan hydroxylase isoform (Abstract), Science, 2003, 299, 76 CrossRef CAS PubMed.
Footnote |
| † Equally to this work. |
| This journal is © The Royal Society of Chemistry 2020 |