
Copper nanoparticles induce zebrafish intestinal defects via endoplasmic reticulum and oxidative stress†
Guang
Zhao
,
Ting
Zhang
,
HaoJie
Sun
and
Jing-Xia
Liu
 *
*
College of Fisheries, Key Laboratory of Freshwater Animal Breeding, Ministry of Agriculture, Huazhong Agricultural University, Wuhan, 430070, China. E-mail: ichliu@mail.hzau.edu.cn; Fax: +86-27-87282113
First published on 28th November 2019
Abstract
As an essential trace element, copper plays key roles in the activation of multiple enzymes, neurotransmitter biosynthesis and denaturation, as well as the decomposition of superoxide and the synthesis of collagen. The intestines is the main organ for copper absorption and transfer, and intestinal copper accumulation is observed in some patients with gene mutations. However, a vertebrate model to link copper accumulation with intestinal diseases and defects is still lacking, and the data concerning the mechanisms underlying this link are still scarce. In this study, the effects of exogenous copper (CuNPs or their released Cu2+) on intestinal development in zebrafish embryos were investigated after their exposure to copper. The results showed that over 0.10 mg L−1 of CuNPs or Cu2+ damaged the zebrafish intestinal development, including thinned epithelial cells as well as few and shortened intestinal villi. Under CuNP stress, the expression was significantly (p < 0.05) reduced for the intestinal marker genes (slc15a1b, cyp3a65, cyp8b1, fabp2), but increased for the endoplasmic reticulum (ER) stress marker (bip) in the zebrafish intestines. Additionally, immunofluorescence analysis revealed that CuNPs or Cu2+ induced the production of ER stress (indicated by PDI) and oxidative stress (indicated by 4-HNE) in the intestinal cells. The expression of the aforementioned intestinal marker genes could be restored to normal by inhibiting the production of ER stress or oxidative stress with ER stress alleviator PBA (4-phenylbutyric acid) or ROS scavengers GSH (reduced Glutathione) or NAC (Nacetylcysteine) in CuNP or Cu2+ stressed embryos, suggesting that copper induces intestinal defects mainly by ER and oxidative stress. Moreover, obvious intestinal defects were observed in copper-stressed cox17−/− and atp7a−/− mutants, implying that blocking the transportation of copper to the mitochondria or trans-Golgi network by deleting cox17 or atp7a could not alleviate copper-induced intestinal developmental defects. This is probably the first report to reveal that copper nanoparticles (CuNPs) and their released ions (Cu2+) cause intestinal developmental defects via inducing ER and ROS stresses. It is also the first report on the intestinal developmental responses of cox17−/− or atp7a−/− mutants to copper stimulation.
Significance to metallomicsThis study unveiled that both CuNPs and their released Cu2+ damaged embryonic intestinal development in a dosage-dependent manner via inducing ROS and ER stresses in intestinal cells. Partially blocking the transportation of copper to mitochondria (cox17−/−) or the trans-Golgi network (atp7a−/−) could not alleviate copper-induced intestinal developmental defects. This study provides an integrated study system associating copper with its chaperones in intestinal developmental abnormalities and diseases. |
Introduction
Copper homeostasis in cells is important for the normal and healthy development of vertebrates, and either copper deficiency or excess in cells will lead to development abnormalities and diseases. Previous reports have unveiled that the serum copper level is significantly higher in children with inflammatory bowel disease (IBD).1 Additionally, excess copper induces intestinal immune imbalance2 and causes intestinal cell apoptosis in chickens.3 Copper uptake is mainly performed through the intestines, suggesting that the intestines may be the organ most susceptible to excess copper. However, we still lack a vertebrate model to link excess copper in the intestines with its resultant diseases or developmental abnormalities.Due to their antibacterial activities and rapid electrical conductivity, copper nanoparticles (CuNPs) are extensively applied in various fields, such as additives in lubricants, polymers/plastics, metallic coating and inks, etc.4–6 The increased consumption of CuNPs might increase their contamination in the environment, and their released ions, Cu2+, are believed to be one of the most important pollution-causing metals. Studies have unveiled that aquatic organisms may suffer from exposure to Cu concentration ten to fifty times higher than required.7
Excess copper is harmful to the healthy development of aquatic organisms. For instance, copper can cause gill injury and acute lethality in adult zebrafish.8–10 It can also induce fish embryonic developmental abnormalities, such as short body length, dysfunctional locomotor behavior,11,12 abnormal swim bladder development and delayed hatching.11,13 However, rare studies have examined the relationship of excess copper with abnormal intestinal development in vertebrates.
Previous studies have shown that CuNPs increased the production of reactive oxygen species (ROS) and reactive nitrogen species (RNS), altered the expression levels of oxidative stress related biomarkers, and activated mitochondria-induced apoptosis,14 with their toxic effects mainly exerted through the released ions, Cu2+.11 One of our recent studies has unveiled that CuNPs induced retinal defects by oxidative and endoplasmic reticulum (ER) stress (data under revision in another journal). Inspired by this, we set out to investigate whether CuNPs and their released Cu2+ damage intestinal development via ROS and ER stresses in this study.
The normal trafficking of copper to different organelles and the balanced homeostasis of copper in cells are pivotal for normal vertebrate development. In the cytoplasm, the absorbed copper will be delivered to the mitochondrion for metalation of cytochrome c oxidase (CcO) by copper chaperone Cox17 and enter the circulation by the pumping action of copper ATPase7a (ATP7a) which resides in the trans-Golgi network (TGN).15,16 In this study, cox17−/− and atp7a−/− mutants were used to detect the integral function of copper chaperones Cox17 and Atp7a in copper induced intestinal development abnormalities during fish embryogenesis.
The purpose of this study was to investigate the effects of copper (CuNPs and their released Cu2+) on intestinal development during zebrafish embryogenesis and the related potential molecular mechanisms. To this end, zebrafish embryos were treated with CuNPs and Cu2+ as reported previously.11,12 The morphological and histological characteristics of the intestinal cells in copper stressed embryos were tested first by transmission electron microscopy (TEM) and Hematoxylin and Eosin (H&E) staining. Then, the expressions of intestinal genes and genes associated with ROS and ER stress sensors in the intestinal cells were evaluated, followed by the examination of the neutralization effects of ROS scavengers reduced Glutathione (GSH) & N-acetylcysteine (NAC) and ER stress alleviator 4-phenylbutyric acid (PBA) on copper induced intestinal defects. Finally, cox17−/− and atp7a−/− mutants were used to verify the correlation between the deficiency of copper trafficking to a specific organelle and the occurrence of intestinal developmental defects in copper-stressed embryos.
Materials and methods
Fish lines, reagents, and antibodies
The AB wild type (WT), and cox17−/− and atp7a−/− adult zebrafish were cultured in a circulating filtration system (28 ± 0.5 °C, 14![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :10 h light:dark). Natural spawning eggs were obtained and maintained in a 28.5 °C incubator. The ages of the embryos and larvae were expressed by hours post-fertilization (hpf) or days post-fertilization (dpf).
:10 h light:dark). Natural spawning eggs were obtained and maintained in a 28.5 °C incubator. The ages of the embryos and larvae were expressed by hours post-fertilization (hpf) or days post-fertilization (dpf).
The following reagents and antibodies were used in this study: copper sulfate (Cu2+), copper nano-powders (CuNPs) and PBA (Sigma-Aldrich, USA); NAC, reduced GSH, and RIPA (Radio Immunoprecipitation Assay) lysis buffer (Beyotime, China), and TRIzol (Life Technology Co, USA). Antibodies: 4-HNE (4-Hydroxynonenal) (Alpha Diagnostic, USA).
Cox17 and atp7a homozygous mutants
Cox17 and atp7a homozygous mutants were constructed by CRISPR/Cas9 technology in our lab recently, detailed information and transcriptional characters for cox17−/− were reported recently,11,17 and detailed information for the atp7a−/− line was presented in another manuscript under review in another journal. In this study, we unveiled the respective roles of the copper transporting proteins of Cox17 and Atp7a in copper induced intestinal developmental defects by disrupting the transfer of copper to mitochondria (cox17−/−) or to the trans-Golgi network & its pumping to circulation (atp7a−/−).Embryo treatment and phenotype observation
Copper (CuNPs and Cu2+) was prepared according to our recently reported methods,11 and embryos were exposed to copper before the sphere stage (4 hpf) as reported in our previous work.18 0.25 mg L−1 copper, including CuNPs and Cu2+, was used for almost all copper stress experiments in this study as we reported recently.17,19 Additionally, different concentrations of copper (0.05 mg L−1, 0.10 mg L−1, 0.15 mg L−1, 0.20 mg L−1) were used in this study to unveil the dosage window of copper in inducing intestinal developmental defects. NAC (200 μM) and GSH (100 μM) were added 2 h before copper treatment. PBA (50 μM) was added at 24 hpf. The embryos were collected at the indicated stages. Embryos from the control and the treated groups were observed and photographed using a light microscope (Leica M205FA) to examine their embryonic morphology.Measure of the copper content in the treated embryos
Inductively coupled plasma mass spectrometry (ICP-MS) analysis was used to test the copper content in the 0.15 mg L−1 copper treated embryos at 24 hpf as we performed previously,12,18,19 and the chorinated18 but not the dechorinated embryos12,19 were used in this study. Copper ions and copper particles have been successfully separated by a modified cloud point extraction protocol (CPEP) in the embryos.19In this study, the samples for testing copper ions and copper particles in embryos stressed with 0.15 mg L−1 CuNPs were prepared using a modified CPEP and were then used for ICP-MS detection. Briefly, embryos at 24 hpf were lysed with 1 mL of ultrapure H2O (normal pH), followed by incubation in ice for 15 min. Thereafter, embryo homogenates were quickly added with 9 mL H2O (pH 3.4, supplemented with 0.2 mL of 0.5 M Na2S2O3 and 0.3 mL of Triton X-114) and were incubated for 10 min in ice, followed by incubation in a water bath for 30 min at 40 °C. These tubes then were centrifuged at 3000g for 5 min to facilitate phase separation, CuNPs were concentrated into a TX-114-rich organic phase, and the released Cu2+ in the embryos was left in the aqueous phase.
Intestinal gene analysis based on RNA-Seq data
Transcriptional profiles in copper (CuNPs or Cu2+) stressed embryos and the control embryos at 96 hpf were unveiled recently.11 Genes that were significantly altered as a result of the CuNP or Cu2+ treatments (adjusted P < 0.05) were defined as differentially expressed genes (DEGs). In this study, we focused on intestinal DEGs based on the transcriptional profiles to unveil the transcriptional characters underlying intestinal development defects in copper stressed embryos. Hierarchical Clustering was performed using TIGR Multiexperiment Viewer (Mev) to form different heatmaps showing the expression patterns of genes among different samples.TEM
The copper (CuNPs and Cu2+) stressed WT, cox17−/−, and atp7a−/−, together with their controls, were collected separately at 96 hpf and fixed with 2% glutaraldehyde at 4 °C for at least 4 h. After processing in a series of solutions as reported previously,20 the embryos were embedded in Quetol 812 (Nisshin EM Co., Ltd; Tokyo, Japan) for sectioning. Finally, the sections were observed under a TEM (Hitachi H-7650 TEM Japan) to analyze the structure of the mitochondria and ER in the intestinal cells of copper stressed embryos and their controls.Frozen sections
The embryos at 72 hpf and 96 hpf were fixed with 4% PFA overnight at 4 °C, and then were dehydrated with 30% sucrose PBS solution for 2 h at room temperature. Next, the permeated embryos were embedded in Tissue-Tek® O.C.T. compound (Sakura Finetek, USA) for cryosectioning at 6–8 μm thickness using frozen microtomy (Thermo scientific, USA). After drying at 4 °C, the sections were stored for H&E staining and immunofluorescence assays.Hematoxylin and Eosin (H&E) staining
The H&E staining was performed as reported previously,20 and the high-resolution images for the H&E staining sections were obtained under a microscope (ZEISS Axio Imager A2).Immunofluorescence
The immunofluorescence was performed with the primary antibodies against 4-HNE in a 1:200 dilution. Secondary antibody conjugated with Alexa Fluor 488 was used in a 1:500 dilution. 40,6-Diamidino-2-phenylendole (DAPI) was used to label nuclei. The immunofluorescence images were obtained with a confocal microscope (Olympus FV1000 Confocal Microscope, Japan).
RNA preparation and real-time PCR analysis
Total RNA was extracted with TRIzol Reagent, and the cDNA was synthesized by using an M-MLV Reverse-Transcript Kit (Applied Biological Materials Inc., BC, Canada). Real-time qPCR was performed using iQ™ SYBR® Green Super mix (Bio-Rad Laboratories, Hercules, CA, USA) in a CFX Connect™ Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA, USA). Differences were calculated by the ΔΔCt comparative quantization method using β-actin as an internal control, and the data were analyzed with one-way analysis of variance (ANOVA) and post hoc Tukey's test (**, P < 0.01; *, P < 0.05). The specificity of all primers was tested before use, and the sequences of these primers are listed in Table S1 (ESI†).Whole-mount in situ hybridization (WISH)
WISH was performed as described in our previous study.12 Gene-specific primers were designed based on the available information, and the PCR products were cloned into pGEM-T Easy vectors for anti-sense RNA probe synthesis. The primers are listed in Table S1 (ESI†). The anti-sense RNA probes labeled with digoxygenin were used to verify the expression of target genes in whole-mount embryos. The images were taken using a Leica microscope (Leica M205FA, Germany).Statistical analysis
The sample size for different experiments in each group was larger than 10 embryos (n > 10) with 3 biological replicates for each test. The in situ hybridization data were examined by hypergeometric distribution analysis using the software of R-console. The data of qPCR were analyzed by one-way analysis of variance (ANOVA) and post hoc Tukey's test on Statistic Package for Social Science (SPSS) 19.0 software. Statistically significant differences among groups were indicated by ** P < 0.01, and * P < 0.05.Results
Excess copper causes intestinal defects in zebrafish embryos
Histological staining showed that the copper-stressed zebrafish embryos had narrowed intestinal lumen and thinned intestinal cells (Fig. 1A). Additionally, the TEM analysis unveiled short and sparse zebrafish intestinal microvilli (indicated by red arrows) and atrophied intestinal epithelial cells (indicated by green dotted lines) in copper stressed embryos (Fig. 1B2 and B3) versus the control embryos (Fig. 1B1). RNA-seq analysis showed that copper down-regulated the expression of the intestinal related genes (Fig. S1, ESI†). qPCR revealed the down-regulated expression of four intestinal related genes (slc15a1b, cyp3a65, cyp8b1, and fabp2) in copper stressed embryos (Fig. 1C). WISH analysis further confirmed the down-regulated expression of slc15a1b and fabp2 in the whole copper stressed embryos at 96 hpf (Fig. 1D).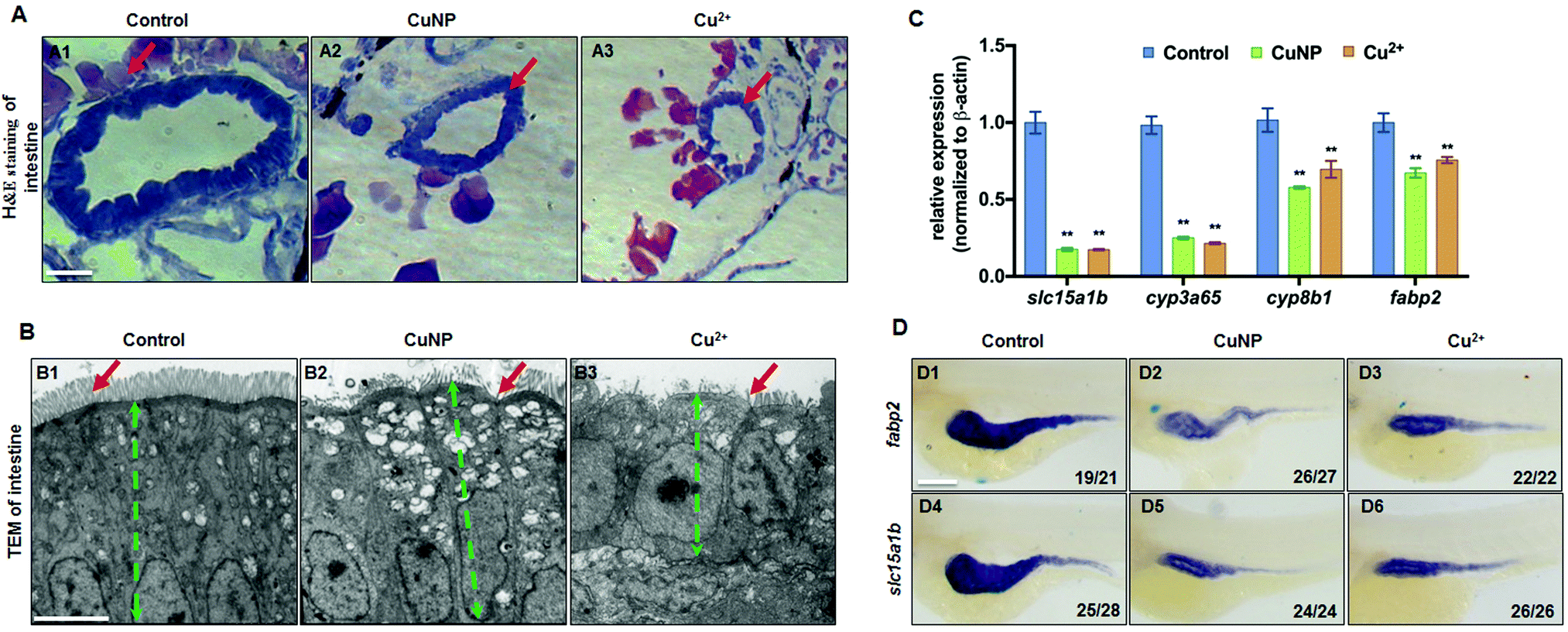 | ||
| Fig. 1 Intestine developmental defects in copper stressed embryos. (A) H&E staining analysis of the intestines (indicated by the red arrows) of embryos from control, Cu2+-stressed and CuNP-stressed groups at 96 hpf. (B) TEM analysis of the intestines of embryos from control (B1), Cu2+-stressed (B2), and CuNP-stressed groups (B3) at 96 hpf. Red arrows indicate the intestinal villi, and green dotted lines indicate the thickness of the intestinal epithelial cells. (C) Expression of intestinal marker genes slc15a1b, cyp3a65, cyp8b1, and fabp2 in embryos from control, Cu2+-stressed, and CuNP-stressed groups. (D) WISH data of fabp2 (D1–D3) and slc15a1b (D4–D6) in embryos from control, Cu2+-stressed, and CuNP-stressed groups, respectively. The number in the lower right corner in each panel indicates the number of embryos that exhibited reduced expression/the number of total tested embryos in each group, and the same for the indicated numbers in the following figures for WISH assays. (A) Cross sections at the intestine domain; (B) sagittal sections at the intestine domain; (D) lateral view, anterior to the left; scale bar: (A) 50 μm; (B) 5 μm; (D) 100 μm. ** P < 0.01. | ||
To verify the effective copper concentration window that causes intestinal defects, fertilized embryos were exposed to copper at different concentrations including 0.05 mg L−1, 0.10 mg L−1, 0.15 mg L−1 and 0.20 mg L−1. qPCR (genes fabp2 and slc15a1b) and WISH (gene fabp2) data indicated that the expression of intestinal markers was significantly reduced in embryos stressed with copper at and over 0.15 mg L−1 but not at 0.05 mg L−1 and 0.10 mg L−1 (Fig. 2A–D). Furthermore, the total copper content in 0.15 mg L−1 CuNPs- and Cu2+-stressed chlorinated embryos at 24 hpf was 10.31 μg g−1 and 11.8 μg g−1 respectively (Fig. 2E). Additionally, the content of copper particles and ions in 0.15 mg L−1 CuNPs-exposed embryos was 2.19 μg g−1 and 8.12 μg g−1 at 24 hpf, respectively (Fig. 2E).
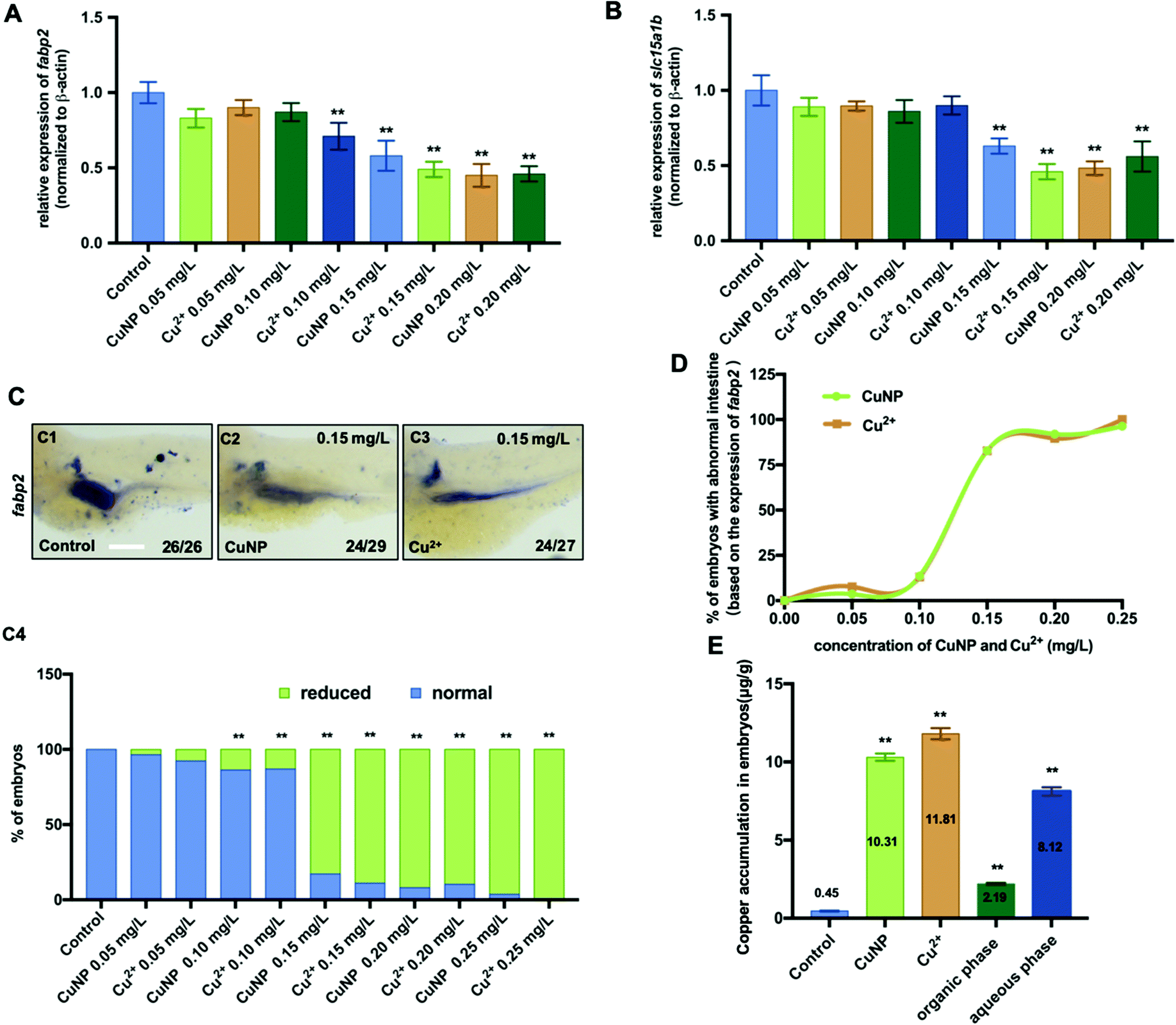 | ||
| Fig. 2 Effects of different concentrations of copper on the expression of intestinal marker genes and copper accumulation in embryos. (A) qPCR detection unveiled the expression of fabp2 in embryos treated with different concentrations of copper (CuNPs or Cu2+) from 0.05 mg L−1 to 0.20 mg L−1. (B) qPCR detection unveiled the expression of slc15a1b in embryos treated with different concentrations of copper (CuNPs or Cu2+) from 0.05 mg L−1 to 0.20 mg L−1. (C) WISH assays of fabp2 in embryos treated with different concentrations of copper (CuNPs or Cu2+) from 0.05 mg L−1 to 0.20 mg L−1, and the percentages of embryos that exhibited reduced expression of fabp2 in different groups were calculated in (C4). (D) Toxic concentration curve of copper (CuNPs or Cu2+) in embryos with intestine defects (based on the expression of fabp2). (E) Copper concentration in embryos exposed to 0.15 mg L−1 copper (CuNPs or Cu2+). (C) Lateral view, anterior to the left; scale bar: (C) 100 μm. ** P < 0.01. | ||
Copper induces ROS and ER stresses in embryonic cells
The mechanism by which copper damages the intestine cells was investigated by TEM analysis of the structure of mitochondria and the ER. Compared with WT, the ER formed a loose structure in copper stressed embryos (Fig. 3A, indicated by green arrows). Additionally, the mitochondrial inner membrane was found to be decreased and form large vacuoles in copper stressed embryonic intestinal cells (Fig. 3A, indicated by red arrows). Copper has been widely reported to be associated with ER stress and ROS production.14,21 Therefore, ER stress and ROS markers in intestinal cells were further investigated in this study. The WISH analysis revealed the increased expression of an ER stress marker (bip) in intestinal cells in copper-treated embryos (Fig. 3B). Moreover, immunofluorescence with a 4-HNE antibody (ROS marker) showed that copper induced an obvious increase of 4-HNE fluorescence intensity in zebrafish embryonic intestinal cells (Fig. 3C).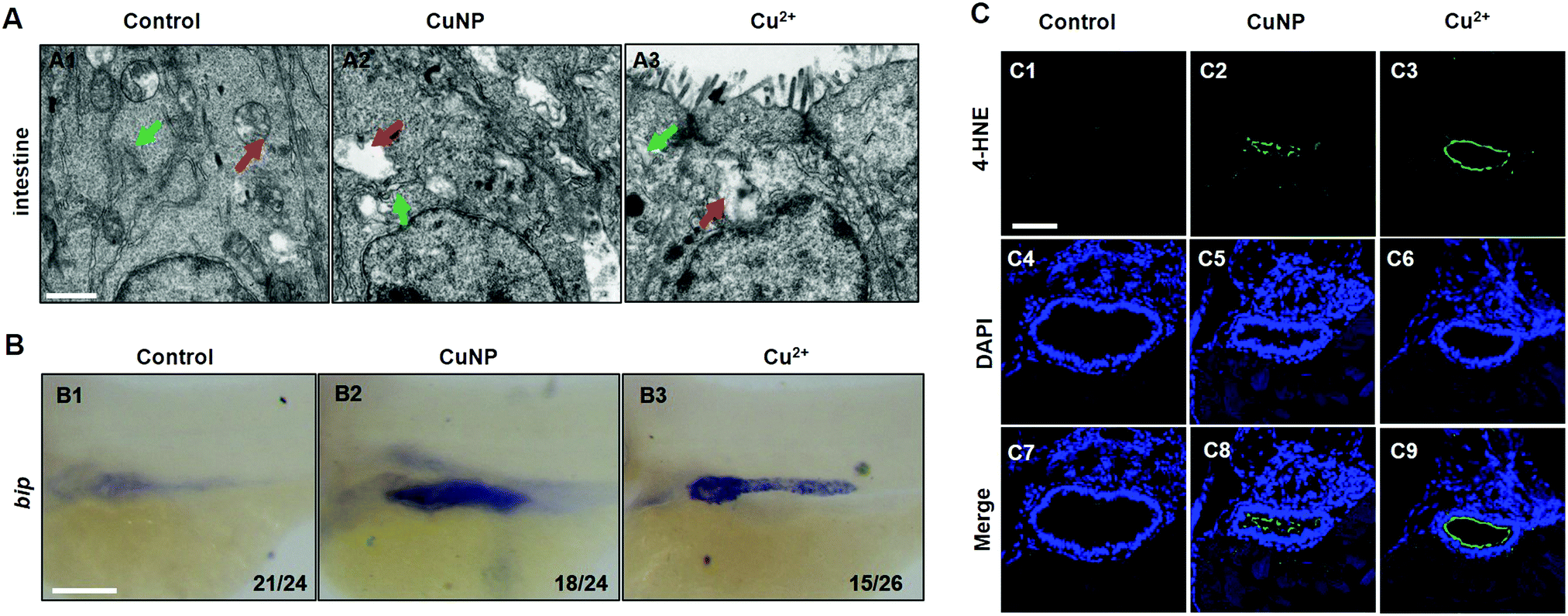 | ||
| Fig. 3 Mitochondrial and ER structure and stresses in copper-stressed embryos. (A) TEM analysis of the mitochondrial and ER structure in intestinal cells in control and copper-stressed embryos respectively at 96 hpf. Red arrow indicates mitochondria and green color indicates ER. (B) WISH assays of an ER stress marker gene (bip) from Control, Cu2+-stressed, and CuNP-stressed groups. (C) Immunofluorescence with an antibody of 4-HNE in intestinal sections at 96 hpf. (A) Sagittal sections at the intestine domain; (B) lateral view, anterior to the left; (C) cross sections at the intestine domain. Scale bar: (A) 0.5 μm; (B and C) 100 μm. | ||
Furthermore, two different ROS scavengers (NAC and reduced GSH) and an ER stress alleviator (PBA) were used to rescue the copper-caused intestinal tract defects in this study. Quantitative PCR revealed that all copper-induced down-regulated expression of intestinal marker genes could be rescued by the addition of either ROS scavengers NAC&GSH or ER stress alleviator PBA (Fig. 4A). WISH data showed that the expression of slc15a1b was restored to nearly normal levels by GSH supplementation in copper-treated embryos (Fig. 4B).
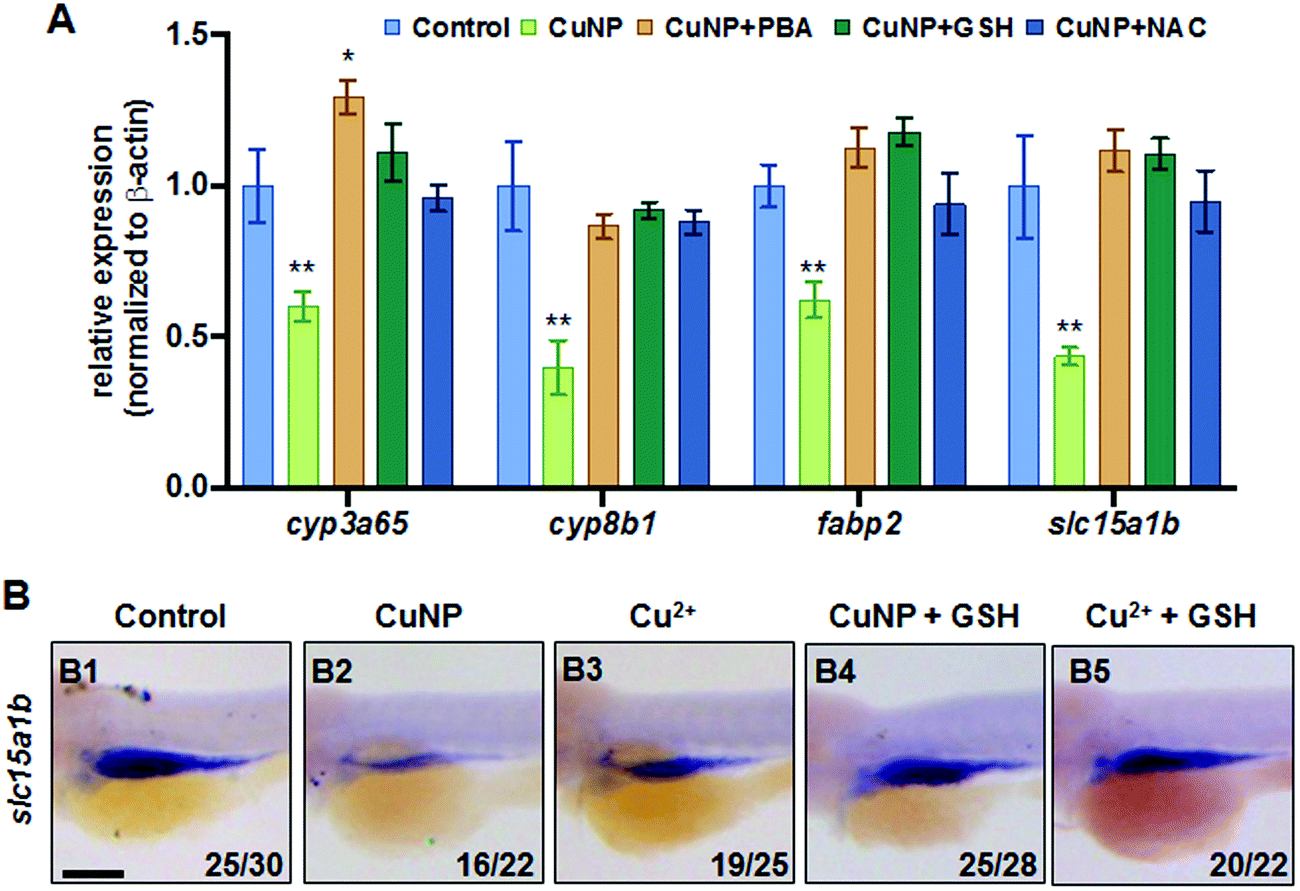 | ||
| Fig. 4 Rescue of intestinal defects with the addition of ROS scavenger or ER stress scavenger. (A) ROS scavengers GSH & NAC and ER scavenger PBA recovered the down-regulated expression of intestinal markers (slc15a1b, cyp3a65, cyp8b1, and fabp2) induced by copper stresses. (B) WISH data of slc15a1b in copper stressed and copper plus GSH co-treated embryos. (B) Lateral view, anterior to the left; scale bar: 100 μm. ** P < 0.01. | ||
Intestinal development in copper stressed cox17−/− and atp7a−/− mutants
The cox17 mutant was used to test the failure of copper trafficking to mitochondria in intestinal defects under copper treatment. In the cox17−/− mutant, the loose ER and slightly swollen mitochondria were observed in intestinal cells (Fig. 5A2). After copper treatment, more seriously swollen mitochondria (Fig. 5A3 and A4, indicated by red arrows) and loose ER (Fig. 5A3 and A4, indicated by green arrows) were observed in the intestinal cells.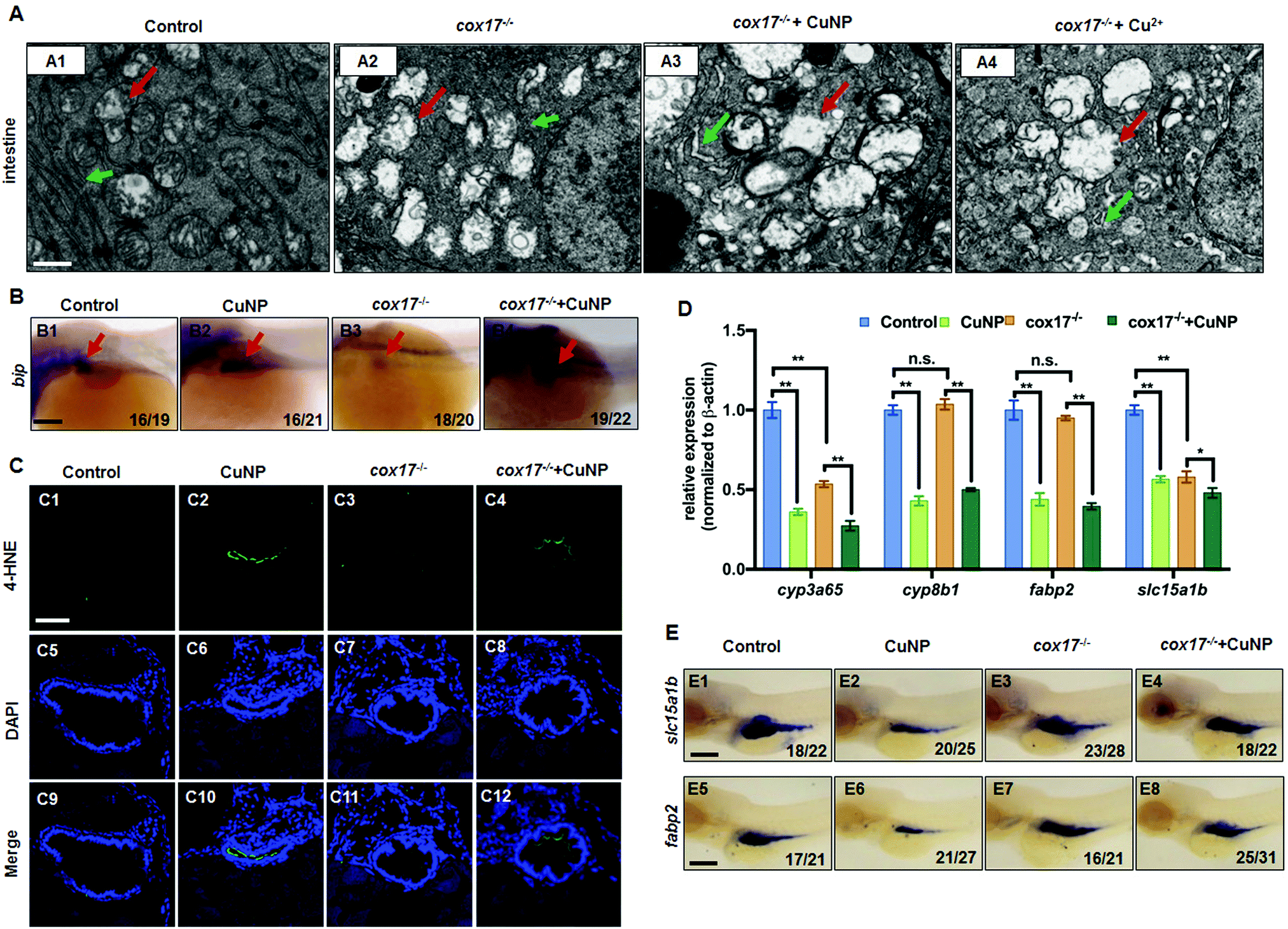 | ||
| Fig. 5 Intestinal development in the cox17−/− mutant and copper stressed cox17−/− mutant. (A) TEM analysis of intestinal cells in zebrafish embryos (Control and cox17−/− mutant) at 96 hpf. (A1–A4) Sagittal slides of the intestines; red color indicates mitochondria and green color indicates ER. (B) WISH analysis of the ER stress marker bip in cox17−/− mutants. (C) Immunofluorescence with the antibody of 4-HNE in the intestinal section of cox17−/− mutants. (D) Quantitative PCR of intestinal markers (cyp3a65, cyp8b1, fabp2, and slc15a1b). (E) WISH analysis of slc15a1b and fabp2. (A) Sagittal sections at the intestine domain; (B and E) lateral view, anterior to the left; (C) cross sections at the intestine domain. Scale bar: (A) 1 μm; (B and E) 100 μm; (C) 100 μm. ** P < 0.01, * P < 0.05. | ||
The ER stress marker bip exhibited increased expression in intestinal cells in the copper-stressed cox17−/− mutant (Fig. 5B), despite its obviously lower expression levels in cox17−/− mutants (Fig. 5B3) than in the WT control (Fig. 5B1). Additionally, the ROS marker 4-HNE exhibited no positive signals in the intestines of cox17−/− mutants (Fig. 5C3 and C11) and slightly positive signals in copper-stressed cox17−/− mutants (Fig. 5C4 and C12), which were obviously lower than those in copper-stressed WT intestinal cells (Fig. 5C2 and C10).
Moreover, qPCR detection showed that, among the four detected intestinal marker genes (cyp3a65, cyp8b1, fabp2, and slc15a1b), cyp3a65 and slc15a1b exhibited significantly reduced expression in cox17−/− mutants, and the expression of the four maker genes was more significantly down-regulated after copper treatment, similar to their expression in copper-stressed WT embryos (Fig. 5D). Additionally, the WISH analysis unveiled slightly reduced expression of intestinal markers slc5a1b (Fig. 5E3) and fabp2 (Fig. 5E7) in cox17−/− mutants, while their expression was further reduced after copper treatment (Fig. 5E4 and E8).
TEM analysis unveiled slightly swollen mitochondria and loose ER in intestinal cells in atp7a−/− mutants. After the copper treatment, more seriously swollen mitochondria (Fig. 6A3 and A4, indicated by red arrows) and no structural ER (Fig. 6A3 and A4, indicated by green arrows) were observed in atp7a−/− intestinal cells.
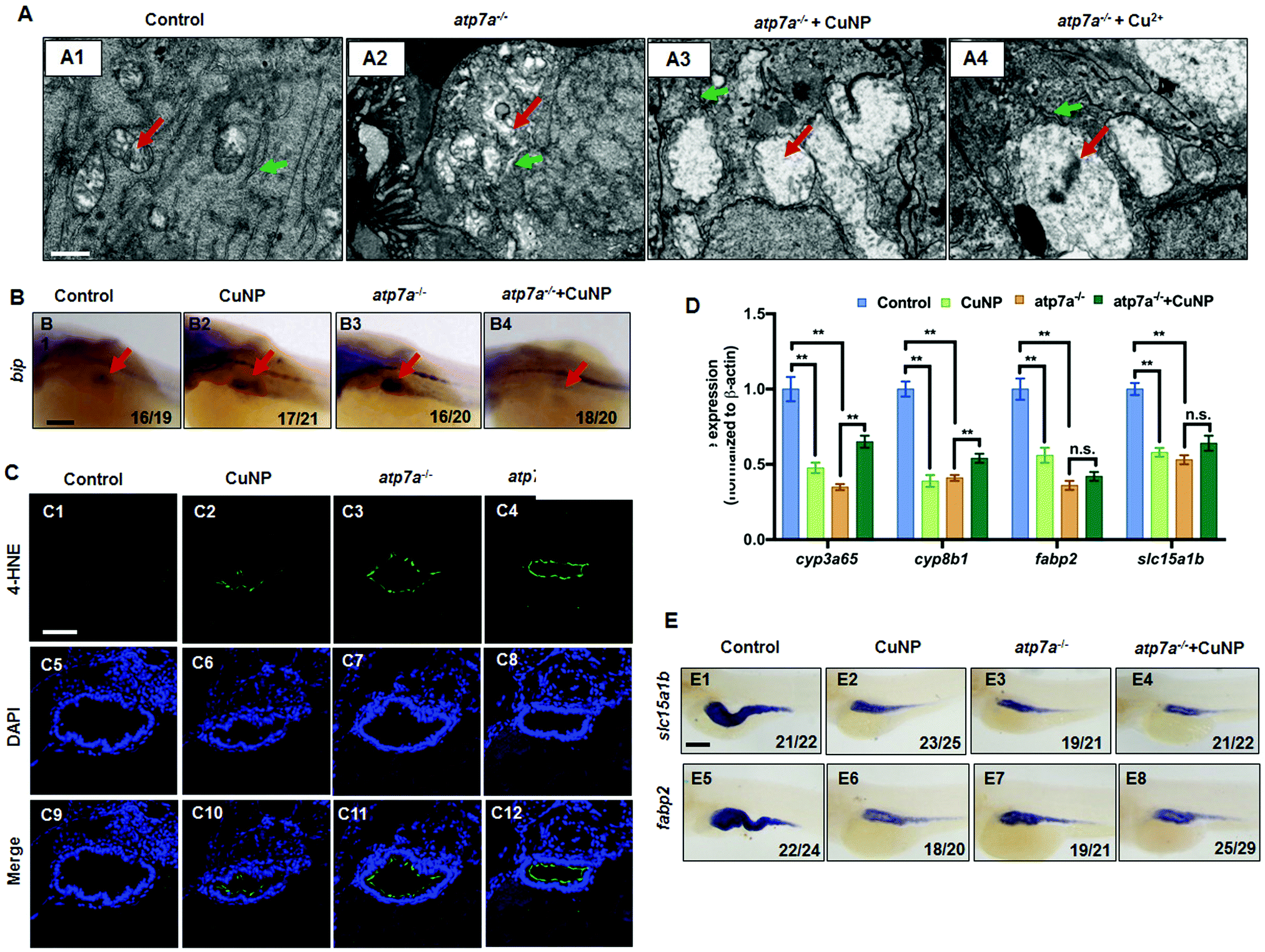 | ||
| Fig. 6 Intestinal development in the atp7a−/− mutant and copper stressed atp7a−/− mutant. (A) TEM analysis of intestinal cells in zebrafish embryos (control and atp7a−/− mutant) at 96 hpf. (A1–A4) Sagittal slides of the intestines; red color indicates mitochondria and green color indicates ER. (B) WISH analysis of ER stress marker bip in atp7a−/− mutants. (C) Immunofluorescence with antibody of 4-HNE in intestinal sections in atp7a−/− mutants. (D) Quantitative PCR of intestinal markers (cyp3a65, cyp8b1, fabp2, and slc15a1b). (E) WISH analysis of slc15a1b and fabp2. (A) Sagittal sections at the intestine domain; (B and E) lateral view, anterior to the left; (C) cross sections at the intestine domain. Scale bar: (A) 1 μm; (B and E) 100 μm; (C) 0.5 μm. ** P < 0.01, * P < 0.05. | ||
WISH detection also revealed an obviously increased expression of the ER gene bip in the intestinal domain in atp7a−/− mutants (Fig. 6B3, indicated by red arrows) versus WT embryos (Fig. 6B1), in contrast to its notably reduced expression (Fig. 6B4) in the copper-treated mutants versus the untreated mutants (Fig. 6B3). Furthermore, the ROS marker 4-HNE showed a positive fluorescence increase in the intestines of atp7a−/− mutants (Fig. 6C3 and C11), while no fluorescence was observed in the intestines of the WT embryos (Fig. 6C1 and C9). Meanwhile, a positive fluoresce increase was observed in the intestines of copper stressed WT (Fig. 6C2 and C10) or atp7a−/− embryos (Fig. 6C4 and C12) versus their respective control.
Significantly reduced expression was unveiled in the intestinal cell markers (slc15a1b, cyp3a65, cyp8b1, and fabp2) in untreated atp7a−/− mutants by both qPCR (Fig. 6D) and WISH analyses (Fig. 6E). After copper treatment, their expressions were not further reduced in atp7a−/− mutants, with the expression of cyp3a65 and cyp8b1 being even increased in the mutants (Fig. 6D).
Discussion
CuNPs are widely used in various fields, and produce obvious toxic effects on organisms. Almost all of the copper is absorbed into the body through the intestines and is pumped into the portal circulation through the basolateral membrane of intestinal epithelial cells via P-type ATPase protein ATP7A.22 Copper overload has been reported in the intestines and liver of ATP7A patients23 and in zebrafish atp7a−/− mutants.24 While the mechanisms underlying hepatic damage under the copper overload condition have been extensively studied,25 there is still a lack of data on the molecular mechanisms of intestinal damage induced by copper overload. In this study, WT, cox17−/− or atp7a−/− embryos stressed with copper (CuNPs or their released Cu2+) were used to detect the effect of excess copper on the zebrafish intestinal development, with the purpose of establishing a vertebrate model for mimicking the initiation and process of intestinal damage in the copper overload condition. This study unveiled that copper damaged the ER and mitochondrial structure in intestinal cells, and induced intestinal defects via oxidative and ER stress. The data in this study might provide some hints for therapy targeted research on copper overload and the resultant intestinal defects and diseases.In this study, H&E staining unveiled a thinned intestinal cell layer and narrowed intestinal lumen in copper-stressed zebrafish embryos, and the TEM analysis unveiled short and sparse intestinal villi in the intestines of copper (CuNPs or their released Cu2+) stressed zebrafish embryos, suggesting that copper damages the intestinal structure and development in vertebrates. The RNA-seq data unveiled a significantly down-regulated expression of intestinal related genes in 4 dpf zebrafish embryos. Additionally, both WISH and qPCR analyses detected the reduced expression of intestinal genes (slc15a1b, cyp3a, cyp8b1, fabp2) in copper stressed embryos, and the reduced protein level of FABP2 was also observed in the intestinal cells of copper stressed embryos. Slc15a1b (also named as PEPT1), a peptide transporter of zebrafish, is highly expressed in the intestines.26 Both cyp3a65 and cyp8b1 genes are localized to the intestines and are closely related to its metabolic function.27,28Fabp2 is mainly involved in the absorption and transport of fat by the body. Recent studies have shown that fabp2 is closely related to metabolic diseases and inflammatory bowel injury.29,30 The down-regulated expression of the aforementioned genes in copper stressed embryos suggests that copper might disrupt the normal metabolism homeostasis of the intestinal cells and cause developmental defects of the intestines in zebrafish.
In order to unveil the dosage window of copper in intestinal development during embryogenesis, the embryos were exposed to a gradient of copper, and the expression of intestinal marker genes was used as a damage indicator in this study. It has been unveiled that significantly reduced expression of intestinal marker genes (slc15a1b, and fabp2) was observed in copper stressed embryos at concentrations ≥0.15 mg L−1. Meanwhile, the CuNPs and their released Cu2+ concentration in 0.15 mg L−1 copper exposed embryos was evaluated by ICP-MS, which revealed that nearly 80% of the total copper is copper ions. This suggests that CuNPs may cause intestinal defects through their released Cu2+ in this study.
The quantification limits for copper detections using ICP-MS were 0.17 μg L−1 to 0.5 μg L−1 respectively. It has been unveiled that over 30 μg g−1 copper was unveiled in 0.25 mg L−1 copper stressed chlorinated embryos at 24 hpf,18 and nearly 10 μg g−1 copper was unveiled in 0.15 mg L−1 copper stressed embryos in this study, and the percentage of embryos that exhibited intestinal defects increased with increased copper concentrations, suggesting the dosage-dependent effects of copper on intestinal development during embryogenesis.
CuNPs and their released Cu2+ have been unveiled to induce ROS in embryos,11,12 and one of our recent studies unveils that CuNPs cause retinal developmental defects via inducing oxidative stress and ER stress (manuscript under review). In this study, TEM assays unveiled expanded vacuolar mitochondria and cracked ER in intestinal cells in copper (CuNPs or Cu2+) stressed embryos. Additionally, 4-NHE (an indicator for oxidative stress) and PDI (an indicator for ER stress) exhibited increased expression in intestinal cells in copper stressed embryos. All these results demonstrate that CuNPs or Cu2+ caused oxidative stress and ER stress in zebrafish intestinal cells and the sustained stresses damaged the structure of the ER and mitochondria. ER and oxidative stresses have been reported to produce cell damage and induce cell apoptosis,31,32 and have a synergistic effect on tissue damage in some cases.33 In this study, increased ER and ROS stresses were observed in intestinal cells in copper stressed embryos, and ROS scavengers and ER stress antagonists could effectively restore the copper induced intestinal developmental defects, further confirming that CuNPs and their released Cu2+ induced intestinal defects through both oxidative and ER stresses. The point raised in this study is consistent with a previous report showing that copper induced the production of reactive oxygen species in grass carp, thereby increasing protein oxidation and lipid peroxidation in enterocytes.7
In this study, the association between ROS & ER stress and intestinal damage was further verified by using cox17−/− and atp7a mutants. Specifically, under an excess copper condition, the cox17−/−mutant with a 4-bp deletion was assumed to have lost the capacity to traffic copper to mitochondria in cells, and the atp7a−/− mutant with a 5-bp deletion was assumed to have lost the capacity to traffic copper to the TGN and pump it into the portal circulation through the basolateral membrane of intestinal epithelial cells. Both the mutants were used in this study with two aims in mind: (i) to test the failure of copper transfer to different organelles in copper induced intestinal developmental defects, and (ii) to identify the roles of copper transporter proteins Cox17 and Atp7a in copper induced intestinal defects.
In this study, the cox17−/− mutant was used to study the association of copper with oxidative stress and intestinal developmental defects. In the cox17−/− mutant, a slight mitochondrial swelling, but a normal ER structure was observed in intestinal cells. Under copper stress, the ER and mitochondria in cox17−/− mutant intestinal cells were significantly damaged, coupled with the down-regulated expression of intestinal-related genes. Our recent studies have revealed that the knockout of the zebrafish cox17 gene can effectively inhibit copper induced ROS in embryos by DCFH-DA (2′,7′-dichlorodihydrofluorescein-diacetate) staining and expression assays of oxidation reduction genes.31 However, mitochondrial swelling was still observed in intestinal cells in the cox17−/− mutant before and after copper treatment, which we suspect might result from the damage to another function of Cox17 in the cox17−/− mutant. It has been unveiled that besides its role in copper metalation of cytochrome c oxidase (CcO) for mitochondrial respiratory energy production, functional Cox17 is also required for modulating the MICOS complex integrity and involved in maintaining the architecture of the MICOS complex.34 However, we still need more solid evidence to confirm the association of the mitochondrial structure damage and ROS production in cox17−/− mutants.
The ER stress and ER structural damage were still observed in the intestinal cells in the cox17−/− mutants by the WISH and TEM analysis, and ER sensors were up-regulated in the intestinal cells of cox17−/− mutants after copper treatment. These observations not only support the assumption that copper-mitochondrial trafficking and ROS production are blocked in cox17−/− mutants but not ER stress, but also suggest that ER stress might contribute most to the further down-regulated expression of intestinal markers in cox17−/− mutants after copper treatment.
Additionally, swollen mitochondria and cracked ER in intestinal cells accompanied by significant down-regulated expression of intestinal markers were observed in the atp7a−/− mutant in this study. Previous studies have revealed copper accumulation in intestinal cells in the zebrafish atp7a−/− mutants.24 Taken together, we demonstrated that copper accumulation in intestinal cells induced mitochondrial and ER structural damage and stress, leading to intestinal developmental defects in the zebrafish atp7a−/− mutants. However, more severely swollen mitochondria and relatively normal ER were observed in intestinal cells in the atp7a−/− mutant after copper treatment, coupled with significantly down-regulated expression of ER sensors and no significantly reduced expression of intestinal marker genes. One possible explanation for this observation is the moderately sustained unfolded protein response (UPR) which has been unveiled to maintain a normal intracellular environment,35 and this protective mechanism might be activated in atp7a−/− mutants. Another possible explanation is that an excessive overload of copper in intestinal cells might induce the collapse of the ER and ROS response systems in the atp7a−/− mutants after copper treatment, because of the failure to pump the copper from intestinal cells to circulation and the accumulation of more copper in the intestinal cells in atp7a−/− mutants after copper treatment.
This study made the first attempt to reveal the detailed molecular characteristics of copper-induced intestinal tract defects. Specifically, copper induces ROS and ER stresses, giving rise to embryonic intestinal developmental defects. The integral function of copper transporters Cox17 and Atp7a in intestinal developmental defects was explored using cox17−/− and atp7a−/− mutants in this study. A simple schema was provided to illustrate the transport of copper in the intestinal cells and the functions of cox17 & atp7a in zebrafish embryos after copper treatment (Fig. 7).
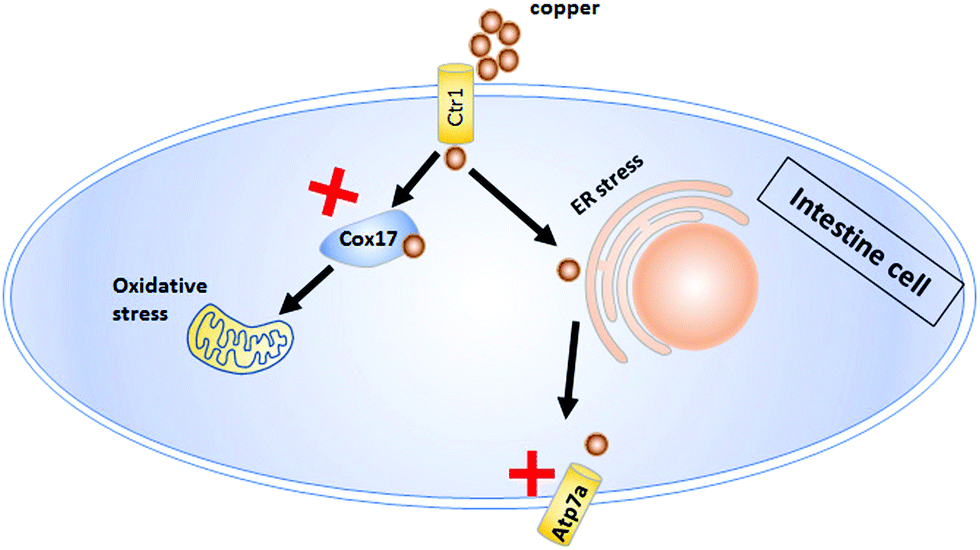 | ||
| Fig. 7 Model for the mechanism of copper induced intestinal developmental defects in WT, cox17−/−, and atp7a−/− embryos. Copper (Cu2+) is mainly absorbed by intestinal cells in larvae, then pumped to the circulation and transferred to different cells by Atp7a. The atp7a−/− mutant fails to pump copper to the circulation and causes copper overload in intestinal cells, leading to intestinal defects in the atp7a−/− mutant. Cox17 is a key carrier protein for copper transport to mitochondria, and the failure of the cox17 mutant to transport copper to the mitochondria causes no notable oxidative stress in the cox17 mutant and thus no significant intestinal defects. | ||
Funding
This work was supported by National Key R&D Program of China (2018YFD0900101), and by the project 2662018JC024 of the Fundamental Research Funds for the Central University (to J.-X. L.).Conflicts of interest
The authors declare that they have no competing interests.References
- A. Ojuawo and L. Keith, The serum concentrations of zinc, copper and selenium in children with inflammatory bowel disease, Cent. Afr. J. Med., 2002, 48, 116–119 CAS.
- Y. Wang, H. J. Zhao, J. J. Liu, Y. Z. Shao, J. L. Li, L. Y. Luo and M. W. Xing, Copper and arsenic-induced oxidative stress and immune imbalance are associated with activation of heat shock proteins in chicken intestines, Int. Immunopharmacol., 2018, 60, 64–75 CrossRef CAS PubMed.
- H. J. Zhao, Y. Wang, Y. Z. Shao, J. J. Liu, Y. H. Liu and M. W. Xing, Deciphering the ionic homeostasis, oxidative stress, apoptosis, and autophagy in chicken intestine under copper(II) stress, Environ. Sci. Pollut. Res., 2018, 25, 33172–33182 CrossRef CAS.
- A. S. Adeleye, E. A. Oranu, M. Tao and A. A. Keller, Release and detection of nanosized copper from a commercial antifouling paint, Water Res., 2016, 102, 374–382 CrossRef CAS.
- R. Ganesh, J. Smeraldi, T. Hosseini, L. Khatib, B. H. Olson and D. Rosso, Evaluation of nanocopper removal and toxicity in municipal wastewaters, Environ. Sci. Technol., 2010, 44, 7808–7813 CrossRef CAS.
- A. Kahru and K. Savolainen, Potential hazard of nanoparticles: From properties to biological and environmental effects, Toxicology, 2010, 269, 89–91 CrossRef CAS.
- J. Jiang, X. Y. Wu, X. Q. Zhou, L. Feng, Y. Liu, W. D. Jiang, P. Wu and Y. Zhao, Glutamate ameliorates copper-induced oxidative injury by regulating antioxidant defences in fish intestine, Br. J. Nutr., 2016, 116, 70–79 CrossRef CAS PubMed.
- B. M. Prabhu, S. F. Ali, R. C. Murdock, S. M. Hussain and M. Srivatsan, Copper nanoparticles exert size and concentration dependent toxicity on somatosensory neurons of rat, Nanotoxicology, 2010, 4, 150–160 CrossRef CAS PubMed.
- R. J. Griffitt, R. Weil, K. A. Hyndman, N. D. Denslow, K. Powers, D. Taylor and D. S. Barber, Exposure to copper nanoparticles causes gill injury and acute lethality in zebrafish (Danio rerio), Environ. Sci. Technol., 2007, 41, 8178–8186 CrossRef CAS PubMed.
- R. J. Griffitt, K. Hyndman, N. D. Denslow and D. S. Barber, Comparison of molecular and histological changes in zebrafish gills exposed to metallic nanoparticles, Toxicol. Sci., 2008, 107, 404–415 CrossRef PubMed.
- Y. J. Zhang, R. T. Zhang, H. J. Sun, Q. Chen, X. D. Yu, T. Zhang, M. Yi and J. X. Liu, Copper inhibits hatching of fish embryos via inducing reactive oxygen species and down-regulating Wnt signaling, Aquat. Toxicol., 2018, 205, 156–164 CrossRef CAS.
- X.-Y. Zhou, T. Zhang, L. Ren, J.-J. Wu, W. Wang and J.-X. Liu, Copper elevated embryonic hemoglobin through reactive oxygen species during zebrafish erythrogenesis, Aquat. Toxicol., 2016, 175, 1–11 CrossRef CAS.
- J. P. Xu, R. T. Zhang, T. Zhang, G. Zhao, Y. Huang, H. L. Wang and J. X. Liu, Copper impairs zebrafish swimbladder development by down-regulating Wnt signaling, Aquat. Toxicol., 2017, 192, 155–164 CrossRef CAS.
- A. Sarkar, J. Das, P. Manna and P. C. Sil, Nano-copper induces oxidative stress and apoptosis in kidney via both extrinsic and intrinsic pathways, Toxicology, 2011, 290, 208–217 CrossRef.
- K. M. Davies, J. F. Mercer, N. Chen and K. L. Double, Copper dyshomoeostasis in Parkinson's disease: implications for pathogenesis and indications for novel therapeutics, Clin. Sci., 2016, 130, 565–574 CrossRef PubMed.
- P. de Bie, P. Muller, C. Wijmenga and L. W. Klomp, Molecular pathogenesis of Wilson and Menkes disease: correlation of mutations with molecular defects and disease phenotypes, J. Med. Genet., 2007, 44, 673–688 CrossRef CAS.
- H. Sun, M. Chen, Z. Wang, G. Zhao and J. X. Liu, Transcriptional profiles and copper stress responses in zebrafish cox17 mutants, Environ. Pollut., 2019, 113364, DOI:10.1016/j.envpol.2019.113364.
- T. Zhang, L. Xu, J. J. Wu, W. M. Wang, J. Mei, X. F. Ma and J. X. Liu, Transcriptional Responses and Mechanisms of Copper-Induced Dysfunctional Locomotor Behavior in Zebrafish Embryos, Toxicol. Sci., 2015, 148, 299–310 CrossRef CAS.
- Y. Zhang, Z. Ding, G. Zhao, T. Zhang, Q. Xu, B. Cui and J. X. Liu, Transcriptional responses and mechanisms of copper nanoparticle toxicology on zebrafish embryos, J. Hazard. Mater., 2018, 344, 1057–1068 CrossRef CAS PubMed.
- X. Niu, J. Hong, X. Zheng, D. B. Melville, E. W. Knapik, A. Meng and J. Peng, The nuclear pore complex function of Sec13 protein is required for cell survival during retinal development, J. Biol. Chem., 2014, 289, 11971–11985 CrossRef CAS.
- S. Oe, K. Miyagawa, Y. Honma and M. Harada, Copper induces hepatocyte injury due to the endoplasmic reticulum stress in cultured cells and patients with Wilson disease, Exp. Cell Res., 2016, 347, 192–200 CrossRef CAS.
- V. Lalioti, G. Muruais, Y. Tsuchiya, D. Pulido and I. V. Sandoval, Molecular mechanisms of copper homeostasis, Front. Biosci., Landmark Ed., 2009, 14, 4878–4903 CrossRef CAS PubMed.
- J. Camakaris, L. Ackland and D. M. Danks, Abnormal copper metabolism in cultured fibroblasts from patients with Wilson's disease, J. Inherited Metab. Dis., 1980, 3, 155–157 CrossRef CAS.
- C. M. Ackerman, P. K. Weber, T. Xiao, B. Thai, T. J. Kuo, E. Zhang, J. Pett-Ridge and C. J. Chang, Multimodal LA-ICP-MS and nanoSIMS imaging enables copper mapping within photoreceptor megamitochondria in a zebrafish model of Menkes disease, Metallomics, 2018, 10, 474–485 RSC.
- P. Manna, M. Ghosh, J. Ghosh, J. Das and P. C. Sil, Contribution of nano-copper particles to in vivo liver dysfunction and cellular damage: Role of I kappa B alpha/NF-kappa B, MAPKs and mitochondrial signal, Nanotoxicology, 2012, 6, 1–21 CrossRef CAS.
- T. Verri, G. Kottra, A. Romano, N. Tiso, M. Peric, M. Maffia, M. Boll, F. Argenton, H. Daniel and C. Storelli, Molecular and functional characterisation of the zebrafish (Danio rerio) PEPT1-type peptide transporter, FEBS Lett., 2003, 549, 115–122 CrossRef CAS.
- H. P. Tseng, T. H. Hseu, D. R. Buhler, W. D. Wang and C. H. Hu, Constitutive and xenobiotics-induced expression of a novel CYP3A gene from zebrafish larva, Toxicol. Appl. Pharmacol., 2005, 205, 247–258 CrossRef CAS.
- I. Bjorkhem and G. Eggertsen, Genes involved in initial steps of bile acid synthesis, Curr. Opin. Lipidol., 2001, 12, 97–103 CrossRef CAS.
- C. Albala, J. L. Santos, M. Cifuentes, A. C. Villarroel, L. Lera, C. Liberman, B. Angel and F. Perez-Bravo, Intestinal FABP2 A54T polymorphism: Association with insulin resistance and obesity in women, Obes. Res., 2004, 12, 340–345 CrossRef CAS.
- S. Heimerl, C. Moehle, A. Zahn, A. Boettcher, W. Stremmel, T. Langmann and G. Schmitz, Alterations in intestinal fatty acid metabolism in inflammatory bowel disease, Biochim. Biophys. Acta, 2006, 1762, 341–350 CrossRef CAS.
- R. Sano and J. C. Reed, ER stress-induced cell death mechanisms, Biochim. Biophys. Acta, 2013, 1833, 3460–3470 CrossRef CAS.
- H.-U. Simon, A. Haj-Yehia and F. Levi-Schaffer, Role of reactive oxygen species (ROS) in apoptosis induction, Apoptosis, 2000, 5, 415–418 CrossRef CAS.
- X. Xue, J.-H. Piao, A. Nakajima, S. Sakon-Komazawa, Y. Kojima, K. Mori, H. Yagita, K. Okumura, H. Harding and H. Nakano, Tumor necrosis factor α (TNFα) induces the unfolded protein response (UPR) in a reactive oxygen species (ROS)-dependent fashion, and the UPR counteracts ROS accumulation by TNFα, J. Biol. Chem., 2005, 280, 33917–33925 CrossRef CAS.
- M. Chojnacka, A. Gornicka, S. Oeljeklaus, B. Warscheid and A. Chacinska, Cox17 Protein Is an Auxiliary Factor Involved in the Control of the Mitochondrial Contact Site and Cristae Organizing System, J. Biol. Chem., 2015, 290, 15304–15312 CrossRef CAS.
- P. Walter and D. Ron, The Unfolded Protein Response: From Stress Pathway to Homeostatic Regulation, Science, 2011, 334, 1081–1086 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c9mt00210c |
| This journal is © The Royal Society of Chemistry 2020 |