
 Open Access Article
Open Access Article This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported Licence
This Open Access Article is licensed under a Creative Commons Attribution-Non Commercial 3.0 Unported LicencePhotosystem ratio imbalance promotes direct sustainable H2 production in Chlamydomonas reinhardtii†
Pilla Sankara
Krishna‡
,
Stenbjörn
Styring
and
Fikret
Mamedov
 *
*
Molecular Biomimetics, Department of Chemistry – Ångström Laboratory, Uppsala University, 751 20 Uppsala, Sweden. E-mail: fikret.mamedov@kemi.uu.se; Tel: +46 18 471 65 81
First published on 31st July 2019
Abstract
The green alga Chlamydomonas reinhardtii can photoproduce H2 gas for only a few minutes under anaerobic conditions due to the inhibition of hydrogenase by O2 produced by Photosystem II (PSII). A few days of sustained H2 production can only be achieved when O2 and H2 production are temporally separated under two-stage processes such as sulfur deprivation. Under sulfur deprivation, H2 production is initiated after the over-reduction of the plastoquinone pool and decreased PSII activity in the thylakoid membrane. As a result, activated hydrogenase consumes the excess of electrons produced by PSII [Volgusheva et al., Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 7223]. Here, we report that similar conditions can be achieved by simply altering the ratio between photosystem I (PSI) and PSII. In the C3 mutant of C. reinhardtii, we found a lower PSI/PSII ratio than in the wild type, 0.33 vs. 0.85, respectively. This imbalance of photosystems resulted in the over-reduced state of the plastoquinone pool and activation of hydrogenase in the C3 mutant that allowed the photoproduction of H2 continuously for 42 days. This is an unprecedented duration of H2 production in green algae under standard growth conditions without any nutrient limitation. Photosynthetic electron flow from PSII to hydrogenase was closely regulated during this long-term H2 production. The amount of PSII was decreased and the amount of PSI was increased reaching a PSI/PSII ratio of more than 5 as shown by EPR and fluorescence spectroscopy. This fine-tuning of photosystems allows to sustain the long-term production of H2 in C. reinhardtii by a direct photosynthetic pathway.
1. Introduction
Solar energy is a renewable energy source that is sufficient to meet future energy demands, provided it can be efficiently harnessed and stored.1 Conserving solar energy in the form of fuel is the most effective way of storage for multiple uses. Therefore, so-called solar fuels are recognized as likely to become an important component of the future energy landscape2 on both the local and global scales.Molecular H2 is a unique energy source which becomes indispensable if can be produced as a solar fuel. It has the highest energy density (ca. 3 times higher than diesel in the liquid form) and, if produced by the direct photolysis of water, is not involved in any CO2 release or CO2 regeneration cycles.3,4 This makes it an environmentally friendly potential energy source and there already exists an appreciable industry making and using H2. Thus, many technical aspects concerning safety, storage and transportation have been established already. Conventional methods of H2 production are either based on fossil fuels (natural gas) or dependent on the electrolysis of water.5 There are, as yet, few technological processes available making H2 from solar energy.
Photobiological H2 production by green algae and cyanobacteria is an area of biotechnology that could become important to fill this gap.2,3 The H2 producing ability of some unicellular green algae has been known for more than seven decades.6 However, there are some shortfalls that prevent its use on the industrial scale. Green alga C. reinhardtii possess both a highly active water oxidation photocatalyst, photosystem II (PSII),7 and a productive H2 producing catalyst, Fe–Fe hydrogenase (HydA).8 This seems to be an ideal system, but in nature, the photoproduction of H2 by microalgae is limited to short bursts that occur only when dark adapted cells in pre-established anaerobic conditions are exposed to light.9 Long-term H2 production is not possible due to the high O2 sensitivity of the HydA enzyme activity and maturation, and the fact that O2 is a by-product of the water oxidation reaction of PSII during its normal turnover.10
This problem can be partially solved by the temporal separation of the PSII activity and H2 production in the two-stage process that occurs during sulfur (S) deprivation in C. reinhardtii.11 Such separation generally leads to several (5–7) days of H2 production in the second stage and has been widely used to study the biochemistry and physiology of this phenomenon.12–15 S-deprivation (the most studied) and other similar two-stage processes (P, N, and Mg-deprivation) commonly lead to an inhibition of the de novo PSII synthesis.16–18 A significant decrease in the PSII amount in the thylakoid membrane eventually makes the cellular O2 consumption due to various respiratory processes overcome O2 evolution in PSII. This situation creates local anaerobiosis in the cell which is a suitable condition for HydA activation and H2 production. It takes 24–48 h of S-deprivation before HydA is activated. During this time, starch is accumulated in the cell.11,19 Both starch and the remaining PSII are electron sources for H+ reduction by HydA.20–22 The reduction of HydA is light-driven by photosystem I (PSI) and wires the plastoquinone (PQ) pool with a stroma soluble ferredoxin (Fd) protein that is the terminal electron carrier to HydA.4 Anaerobiosis is an immediate precondition for HydA expression and function,10,23 although the remaining PSII centers are still active and can supply as much as 80% of electrons that finally reach HydA.12
Significant efforts have been made to increase the efficiency and durability of the H2 production by S-deprived cells of C. reinhardtii. Two major strategies were employed to achieve this goal. The first one employs changing the culture conditions such as light intensity,24 cell immobilization25 and addition of fresh cells growing in the S-deprived medium or dosed sulfate addition.26–28 The second strategy uses a wide range of mutant strains which have limited antenna properties,29–31 decreased PSII activity,32,33 increased respiration,12,34 inhibited cyclic electron flow14,15 or controlled CO2 fixation.35
Several studies have reported a higher H2 yield using S-deprivation than in the original study.11 Scoma et al.32 reported a mutant with double substitution in the PSII D1 protein (L159I-N230Y) with the light to H2 conversion efficiency reaching 3.3%, which is nearly 2 times higher than that in the corresponding WT cells. The stm6 mutant with a high respiration rate showed a 4 times higher final H2 formation than in the WT,12,34 with the total yield reaching more than 13 mmol L−1 after 130 h of S-deprivation. Similarly, in a proton gradient regulation pgr5 mutant, a 10-fold higher H2 yield was shown,15 while in another proton gradient regulation mutant hpm91, a 30-fold increase was obtained.14
The long-term durability and stability of the H2 production is crucial for any technical application. Again, several strategies have been employed. Immobilization of S-deprived cells of C reinhardtii increased continuous H2 photoproduction for at least 90 days under a constant flow of the Tris Acetate Phosphate (TAP) medium containing μM sulfate concentrations. The longest reported period of H2 production so far was achieved by using a two-stage sulfate limited chemostat system with the inflow of fresh culture for 167 days;28 see also ref. 26 and 27. However, all these approaches still have significant limitations such as application of a two-stage process, complex reactor designs and high operation costs.
From our previous studies it became clear that independent of the method used to initiate the H2 production in C. reinhardtii, the starting conditions of this process are (i) decrease in the PSII amount and/or activity that lead to anaerobic conditions in the cell (shown in ref. 12 and 36) and (ii) reduction of the PQ-pool. It is our hypothesis that both these conditions could be achieved by simply altering the balance between two photosystems, PSII and PSI which both consecutively drive the electron transfer in the thylakoid membrane. We tested this hypothesis and now report that, in the C3 mutant strain of C. reinhardtii,37 with an abnormally low PSI to PSII ratio, we indeed observed H2 production for a record period of 42 days in the standard growth media without any modifications.
2. Materials and methods
The C3 mutant strain was obtained from the Chlamydomonas Resource Center (http://www.chlamycollection.org). The C3 mutant and the corresponding WT-137C strain were grown photoheterotropically in 1 L Erlenmeyer flasks containing 300 mL TAP medium at pH 7.0, 25 °C (ref. 49) under continuous white light illumination of 30 μmol photons per m2 per s and constant shaking at 100 rpm for 6 days. After the culture reached a concentration of 12 μg Chl per mL, the cells were collected by centrifugation at 3000g for 2 min. The cell pellet was gently re-suspended in fresh TAP and placed in sealed bioreactors (air tight flasks, 300 ml culture + 20 ml air headspace) at a concentration of 6.0 μg Chl per mL for the long-term incubation experiments that were performed at the same light and temperature conditions. Samples were anaerobically withdrawn for analysis from the sealed bioreactors at the indicated time points. H2 and O2 gas quantifications from the head space were done as described previously.12After every measurement from 24 h and beyond, the H2 amount in the air space of the bioreactor was made zero by flushing with argon. For the experiments in the presence of 20 μM DCMU, 25 mL aliquots were sealed anaerobically in incubation tubes and the H2 yield was determined after 24 h of light incubation. Chl content, flash-induced fluorescence and thermoluminescence measurements were performed according to the published methods.12,36 EPR quantifications of YD• and P700+ to determine the amounts of PSII and PSI, respectively, were performed with a Bruker BioSpin EMX-micro spectrometer equipped with an EMX-Premium bridge and an ER4119HS resonator using a quartz flat cell as described previously.12,38
All data shown are average of at least triplicate data sets. For EPR, fluorescence and thermoluminescence results, typical spectra or traces are shown.
3. Results
The C3 mutant has a lower PSI content and an over-reduced PQ-pool
The C3 mutant cells grow at a similar rate as the WT cells in our standard conditions. The relative amounts of PSI and PSII, present in the thylakoid membrane were determined by EPR spectroscopy.38 Two characteristic EPR signals are observed from the photosystems in photosynthetically active cells at room temperature: the stable YD• radical from PSII and the light or chemically oxidizable P700+ chlorophyll (Chl) radical from the PSI reaction center. Both radicals, when present, correspond to 1 spin per respective photosystem and serve as a robust quantitative probe.12,38Fig. 1A and B show the EPR spectra from fully oxidized YD•, present in the dark adapted cells (black spectra) and spectra from the same cells in the presence of 10 mM K4[Fe(CN)6], which in addition to YD•, already present in the dark, also contain fully oxidized P700 (red spectra). The difference spectra show the deconvoluted, pure EPR spectra from the maximally achievable P700+ (blue spectra). Quantification of YD• and P700+ in the WT cells showed a PSI/PSII ratio of 0.85, which is normal for C. reinhardtii cells (Fig. 1A). In the C3 mutant cells, this ratio was much lower and found to be 0.33. This is almost 3 times lower than that in the WT (Fig. 1B). Thus, there is only one PSI center for every 3 PSII centers present in the C3 mutant. | ||
| Fig. 1 Photosynthetic properties of the WT and C3 mutant of C. reinhardtii. EPR based quantification of PSI and PSII in WT (A) and C3 mutant (B) cells. The spectra shown are from dark adapted cells (YD•, black), the same cells in the presence of 10 mM K4[Fe(CN)6] and following oxidation for 5 min (red) and difference spectra (red spectrum minus black spectrum) respectively (P700+, blue). The EPR quantification of respective photosystems was done according to ref. 38. EPR conditions: microwave frequency 9.76 GHz, microwave power 8 mW, modulation amplitude 5 G, and room temperature. Normalized flash-induced fluorescence decay kinetics (C) and thermoluminescence curves (D) from the control WT (black) and C3 mutant (red) cells. Thermoluminescence was measured in the absence (solid) and presence of 20 μM DCMU (dotted lines). | ||
We also studied the reduction level of the PQ pool in the WT and C3 mutant by flash-induced variable fluorescence decay and thermoluminescence measurements. Fluorescence decay kinetics report on the electron transfer from the primary quinone electron acceptor in PSII (QA−) after a single flash in PSII.39–41Fig. 1C shows the fluorescence decay traces in the control WT (black) and the C3 mutant cells (red). The WT cells show a fast fluorescence decay of more than 80% of the total variable fluorescence. This is normal, and the fast decay reflects efficient forward electron transfer from QA− to QB in PSII centers with an occupied QB-site with QB bound to its pocket in PSII (t1/2 = 232 μs, 58% of the total amplitude) or an empty QB-site (which first requires QB binding, t1/2 = 3.5 ms, 25%).39–41 In the C3 mutant cells, the fluorescence decay was very different: the μsec phase was smaller (38%) and 2 times slower (t1/2 = 462 μsec) indicating that less QB is present in its site. The rest of the decay is dominating by a decay phase (t1/2 = 119 ms, 44%) representing a very slow QB binding. A severe delay of the msec decay phase indicates that the QB binding was significantly perturbed in the C3 mutant. Together, these data show that forward electron transport from QA− is severely slowed down or even inhibited in the C3 mutant.
The situation is further clarified by using thermoluminescence measurements which are a useful complement to the analysis of the fluorescence kinetics.36,42,43Fig. 1D shows the thermoluminescence glow curves from the WT (black) and the C3 mutant cells (red). The WT cells exhibited the characteristic so called the B1-band with the dominating peak at 38 °C. This peak reflects the recombination between QB− and the S2 state of the oxygen evolving complex (OEC) (Fig. 1D, solid black curve, see also Fig. 2S, B†). In contrast, in the C3 mutant cells, the main fraction of the thermoluminescence band, measured under the same conditions, was shifted to 19 °C (Fig. 1D, solid red curve). This clearly indicates that a large proportion of the charge recombination in PSII instead occurs between QA− and the S2 state (the so-called Q-band). The presence of a significant amount of QA− is additional evidence of the mostly empty QB-site in the C3 mutant, again pointing to the reduced state of the PQ-pool favoring QA− S2 recombination instead of QB− S2 recombination.
Upon addition of 20 μM 3-(3,4-dichlorophenyl)-L,L-dimethylurea (DCMU), which completely blocks the QB-site, the clean Q-band could be observed at 3 °C in both WT and C3 mutant (Fig. 1D, dotted curves, WT (black) and C3 (red)).
Long-term H2 production by the C3 mutant
The reduced state of the PQ pool, the low PSI content and the correspondingly low PSI/PSII ratio in the C3 mutant are indicative of severe limitations in the photosynthetic electron transport and will result in a slow PSII turnover and low O2 formation. This could lead to a situation where local anaerobic conditions are established in the cell that could then activate HydA expression.Indeed, when the active C3 mutant culture was placed in gas-tight bioreactors, the O2 concentration quickly decreased and after 17 h, no O2 was detected in the bioreactor's air space. It is important that this occurred without any further modifications of the growth medium or nutrient limitations (Fig. 2, open blue circles). In addition, immediately after the disappearance of O2, H2 production commenced in the C3 mutant culture (Fig. 2, closed blue circles).
 | ||
| Fig. 2 Long-term H2 production in the C3 mutant (filled blue circles) and its absence in the WT (filled black circles) cells of C. reinhardtii in sealed bioreactors under illumination of 30 μmol photons per m2 per s in the complete TAP medium. The arrows show the time points for the fluorescence and EPR measurements (Fig. 3). The inset shows the effect of DCMU addition on the H2 production in the C3 mutant. Blue bars – H2 production in the absence and red bars in the presence of 20 μM DCMU at different time points. The numbers at the top indicate the percentage of inhibition. Decrease in oxygen evolution at the beginning of incubation is shown by open circles for the WT (black) and in the C3 mutant (blue). | ||
The H2 production for the first 42 days is shown in Fig. 2. The amount of H2 released from the C3 cells was increasing during the initial 9 days to more than 3 mL of H2 per L of culture per day. It then decreased somewhat for the next 6 days (15 days in total) to 1 mL of H2 per L of culture per day, and from there on, the cells produced H2 at a relatively stable rate of 0.5–0.7 mL of H2 per L of culture per day for the rest of the incubation (Fig. 2, Table 1). Remarkably, the H2 production by the C3 cells continued for at least 6 weeks (42 days) without any modification of the culture or change in the incubation conditions.
| Culture | Chl (a + b) | Chl a/b | F V/FM | PSII (%) | PSI (%) | PSI/PSII | H2 mL L−1 d−1 |
|---|---|---|---|---|---|---|---|
| 0 h (control) | 5.8 ± 0.10 | 2.00 ± 0.06 | 0.47 ± 0.02 | 100 | 100 | 0.33 | 0 |
| 1 day | 6.4 ± 0.07 | 2.56 ± 0.05 | 0.36 ± 0.03 | 95 | 112 | 0.39 | 1.10 ± 0.12 |
| 2 days | 7.0 ± 0.53 | 2.44 ± 0.33 | 0.22 ± 0.05 | 103 | 166 | 0.53 | 1.25 ± 0.33 |
| 6 days | 8.0 ± 0.11 | 2.65 ± 0.11 | 0.23 ± 0.02 | 58 | 233 | 1.31 | 3.11 ± 0.54 |
| 12 days | 3.5 ± 0.06 | 3.16 ± 0.18 | 0.11 ± 0.06 | 38 | 287 | 2.51 | 1.05 ± 0.47 |
| 20 days | 3.4 ± 0.08 | 3.47 ± 0.27 | 0.20 ± 0.06 | 18 | 236 | 4.36 | 0.51 ± 0.14 |
| 30 days | 3.2 ± 0.06 | 3.35 ± 0.21 | 0.19 ± 0.04 | 14 | 218 | 5.15 | 0.76 ± 0.13 |
It should be mentioned, that when the WT cells were placed in the bioreactors under similar conditions, no H2 production was detected during 30 days of incubation (the first 7 days are shown in Fig. 2, black circles).
We also estimated the contribution of the PSII produced electrons to the H2 production in the C3 mutant. Aliquots of 25 mL of the C3 mutant culture were taken after 1, 6 and 20 days of H2 production and incubated for 24 h in the absence or presence of 20 μM DCMU, a well-known inhibitor of PSII activity. The results are shown in the inset to Fig. 2. After 1 day, only 36% of H2 production was inhibited by the DCMU addition. After 6 days, the inhibition was more than 2 times stronger – 77% and after 20 days, it reached almost 90% (Fig. 2, inset). These results indicate that although PSII is an important supplier of electrons to HydA in the beginning of the H2 production, it becomes indispensable in this capacity during the long-term H2 production in the C3 mutant.
Photosynthetic changes during H2 production in the C3 mutant: Chl content
During the long-term H2 production, the cells of the C3 mutant of C. reinhardtii underwent changes in the Chl content, antenna composition and photosynthetic electron transfer properties. The total amount of Chl in the culture increased during the first 6 days of H2 production from 6 to 8 μg mL−1 and then decreased to 3.5 μg mL−1 during the next 6 days (12 days in total). After this, the total Chl concentration decreased very slowly (Table 1). The Chl composition also changed: the Chl a/b ratio steadily increased for the first 20 days from 2.00 to 3.47 and then remained on the same level over the next 10 days (Table 1). The total number of cells did not change significantly during these 30 days of H2 production. Therefore, these changes are most likely associated with a decrease of Chl b containing the light harvesting complex II (LHCII) antenna associated with both PSII and PSI centers. This conclusion is corroborated by the 77 K fluorescence emission spectra obtained during the course of H2 production (see the ESI and Fig. 1S†).Photosynthetic changes during H2 production in the C3 mutant: relative content of the photosystems
The relative amounts of PSII and PSI during the long-term H2 production in the C3 mutant were determined by electron paramagnetic resonance (EPR) spectroscopy (Fig. 3A and B). In the control sample, there was an imbalance in the photosystems ratio, where 3 PSII centers were found for each PSI center (see above, Fig. 1A and B). During the first 2 days of incubation in the closed bioreactors, the amount of PSII did not change, while the amount of PSI increased by 40% (Fig. 3A, B, 4 and Table 1). Correspondingly, the PSI/PSII ratio had increased to 0.5 (2 PSII centers per 1 PSI center (Fig. 4 and Table 1)).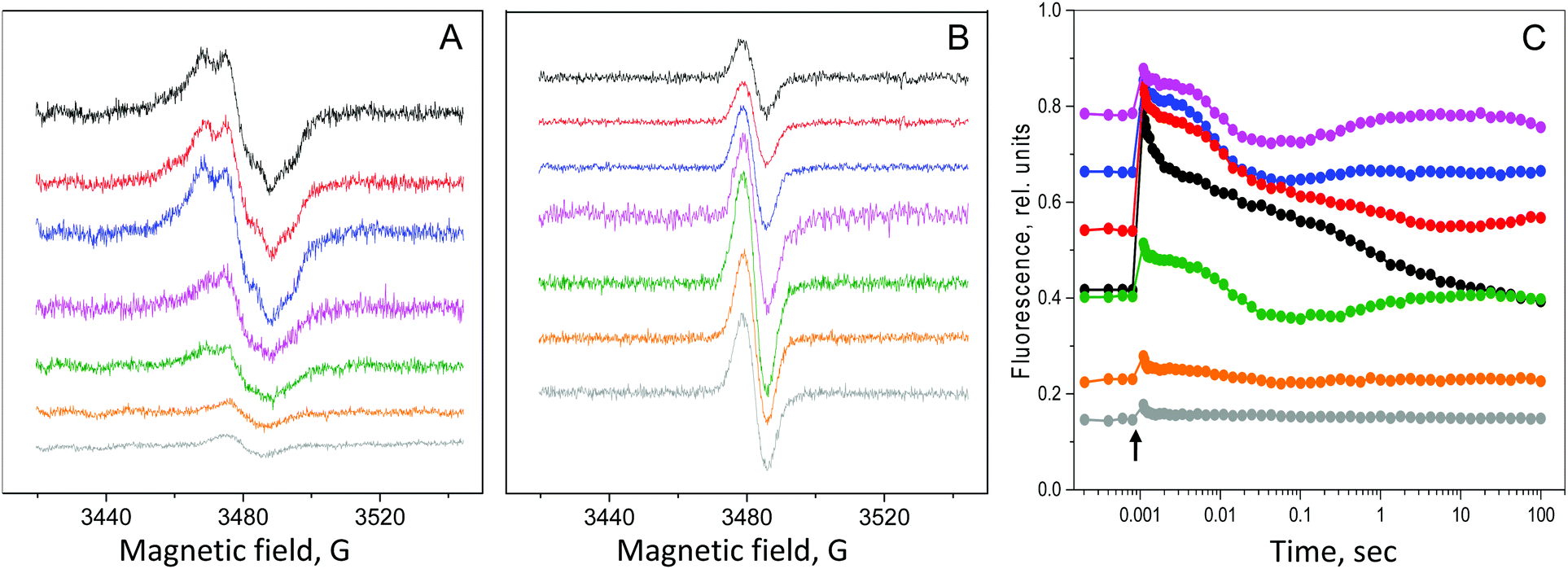 | ||
| Fig. 3 Changes in photosynthetic activity in the cells of the C3 mutant of C. reinhardtii during long-term H2 production in sealed bioreactors. The EPR signal from YD• in PSII (A) and the EPR signal from P700+ in PSI (B) measured by EPR spectroscopy. The amount of respective radicals in the control sample was taken as 100%. EPR conditions as in Fig. 1A and B. The spectra in A and B are normalized to the same Chl concentration. (C) Flash-induced fluorescence decay kinetics of the C3 mutant during the long-term H2 production in sealed bioreactors. The samples from cells of 0 days (control, black), 1 day (red), 2 days (blue), 6 days (pink), 12 days (green), 20 days (orange) and 30 days (grey) of incubation are shown. | ||
 | ||
| Fig. 4 Changes in the amount of PSII (blue), PSI (red) and the PSI/PSII ratio (black) during the first 30 days of H2 photoproduction in the C3 mutant of C. reinhardtii. | ||
During the next 4 days (6 days in total), the amount of PSII decreased to 58% of the amount in the control. Thereafter, the PSII concentration continued to decrease to 14% of the control amount after 30 days (Fig. 3A, 4 and Table 1). The fate of PSI was very different – during the next 10 days (12 days in total), the PSI amount increased almost 3 times to 287% (reaching the amount of PSII in the control sample), and then during the next 18 days (30 days in total), decreased to 218% (Fig. 3B, 4 and Table 1).
As a result, the PSI/PSII ratio underwent significant changes associated with the H2 production in the C3 mutant. It steadily increased from a ratio of 0.33 and reached a ratio of more than 5 (Fig. 4 and Table 1). Thus, while at the beginning of H2 production there were ca. 3 PSII centers per 1 PSI center, this imbalance between the two photosystems was completely reversed during the next 30 days and resulted in a ratio of 5![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :1 PSI to PSII centers in the thylakoid membrane of the C3 mutant.
:1 PSI to PSII centers in the thylakoid membrane of the C3 mutant.
Photosynthetic changes during H2 production in the C3 mutant: PSII and the PQ pool
The changes in the PSI/PSII ratio that occurred during the H2 production stage in the C3 mutant had a marked effect on the electron transfer between PSII and PSI. This effect was studied by flash-induced variable fluorescence decay kinetics and thermoluminescence measurements and the results are shown in Table 1, Fig. 3C and ESI Fig. S2.†The F0 level of the fluorescence increased almost two times during the first 6 days of H2 production from ca. 0.4 to 0.8, Fig. 3C, and then started to decrease to reach 0.15 which is a very low level during the next 24 days (30 days in total, Fig. 3C). The decrease in the F0 level occurred concomitantly with a decrease in the FM level which decreased ca. 5-fold during the same time (Fig. 3C).
The increase in the F0 level reflects the rise in the reduced QA− in PSII which is a consequence of the over-reduced PQ-pool and increasingly reduced conditions in the thylakoid membrane.12,36 While the membrane stays over-reduced during H2 production, the overall decrease in the PSII amount was reflected in the decrease in both F0 and FM and correspondingly in the FV/FM ratio (from 0.47 in the control cells to 0.19 after 30 days of H2 production, see Table 1). It follows the decrease in the amount of PSII centers which was directly quantified by EPR spectroscopy (Table 1 and Fig. 3A), although these two parameters are not directly comparable, since the FV/FM ratio also represents the photochemical efficiency of PSII.44,45
Very informative results were also derived from the analysis of the fluorescence decay kinetics. In the control C3 mutant cells, the flash induced fluorescence decay was slower than in the WT cells as was discussed above (Fig. 1C; also shown in Fig. 3C, black trace). The kinetics were then slowed down further with the establishment of anaerobic conditions and beginning of the H2 production after 1 day in the sealed bioreactors (Fig. 3C, red trace). The μsec decay phase (reflecting forward electron transfer from QA− to bound QB), completely disappeared and was replaced by the slower decay phases in the msec time regime. These slow decay kinetics became even slower after 2 days of incubation (Fig. 3C, blue trace).
During prolonged incubation, 6 to 30 days, concomitantly with a sharp decrease in the FM and F0 parameters (Table 1), the single kinetics with half times of 200–300 ms dominated the fluorescence decay (Fig. 3C, rest of the traces). This phase represents a mixture of very slow QB binding and recombination to the donor side of PSII. In addition, a small rise of variable fluorescence developed after the initial decay which is similar to the kinetics observed in the WT control. This decay pattern indicates that the electron transfer from PSII, although very slow and inefficient, still exist in a strongly reduced environment in the C3 mutant during the whole 30 days of H2 production in the C3 mutant (Fig. 3C). These conclusions from the fluorescence decay data were also supported by the thermoluminescence measurements obtained during H2 production (see the ESI and Fig. S2†).
To conclude, our analysis reveals that PSII in the C3 mutant cells is functional and provides significant electron pressure at the level of the PQ pool even at the over-reduced conditions in the thylakoid membrane. This electron pressure exists if cells are producing H2.
4. Discussion
PSII is both good and bad for long-term H2 production
Long-term direct and sustainable H2 production by photosynthetic microorganisms is an ideal approach to produce renewable fuel from solar energy. There are scientific and technological challenges in the implementation of this approach, the most crucial of which is the O2 sensitivity of HydA. Since photosynthesis is an ultimate electron and proton source for HydA, O2 released by PSII is an unavoidable companion of any long-term H2 production.Two main strategies were undertaken to eliminate this problem. The first one was to increase the O2 tolerance of HydA by genetic manipulation. Although this partially worked for the Ni–Fe type hydrogenases, the results for the algal Fe-Fe HydA have not been encouraging so-far.46,47 The second strategy is to separate O2 evolution and H2 production in time by the so-called two-stage processes namely S-deprivation.11,16,17 However, this approach is not suitable for sustainable long-term H2 production.
In this study, we suggest a new strategy to circumvent the O2 sensitivity of HydA without implementation of any two-stage processes, which we see as very promising for long-term H2 production. The strategy is to suppress the O2 concentration in the algal cell to the level where it does not inhibit the HydA activity. This should be achieved under the standard growth conditions, i.e. without application of any two-stage processes. The way to achieve this is to create a specific balance between PSII and HydA activity in such a way that will allow continuous supply of electrons from PSII and at the same time to have a low intracellular (stromal) O2 concentration which will not inhibit the HydA activity. This means that the PSII activity must be confined to the certain level. The PSII activity is indispensable for long-term cell survival, but it is also important for H2 production since large amounts of electrons for H2 production will be provided by the water splitting activity of PSII (Fig. 2, inset).12 Therefore, we can postulate that PSII is necessary but harmful, i.e. both good and bad for H2 production in green algae. We should also mention that starch reserves as an alternative electron source for HydA will be obsolete in the absence of the two-stage process.
Thus, a novel strategy for long-term H2 production in C. reinhardtti should be to find a right balance between PSII and HydA activities under normal growth conditions. The PSII activity must be high enough to sustain algal cells and, at the same time, low enough in order not to inhibit the HydA activity. Seemingly, the most optimal way to approach this is to limit the PSII activity, for example by mutagenesis.
There are many mutants in PSII (including C. reinhardtii) with a wide range of inhibition of O2 evolution to an almost desirable extent. However, the diminished PSII activity in these mutants will most likely result in the oxidized state of the PQ-pool, and due to the large prevalence of the PSI mediated electron transfer, increase the PSI/PSII ratio. From previous studies, we know that one of the important pre-conditions for the initiation of H2 production in C. reinhardtii is the over-reduced state of the PQ pool which is prerequisite to cellular anaerobiosis.12,16 While inhibition of the PSII activity by mutagenesis could lead to the anaerobic conditions in stroma where HydA is located, it is not the optimal way to achieve the high reduction state of the thylakoid membrane. Under illumination, unaffected PSI activity will mainly keep the PQ pool in the oxidized state.
Alternatively, imposing a limitation on the PSI activity is a more promising approach and we believe it could lead to conditions where the PQ-pool will stay in the reduced state all the time. It will also limit the PSII activity since the electron transfer chain beyond PSII will be log-jammed. In this case, HydA activation and sustainable H2 production could be achieved under the standard growth conditions. This hypothesis was successfully tested in the C3 mutant of C. reinhardtti.
C3 mutant has an optimal ratio between two photosystems for long-term H2 production
The C3 mutant cells have a low PSI/PSII ratio and possesses only 1 PSI center for 3 PSII centers as was shown by our EPR measurements (Fig. 1B). However, most of the antenna was associated with the PSI reaction centers (Fig. 1S, A†) to rebalance the excitation energy distribution towards PSI. Still, the electron transfer on the acceptor side of PSII in the C3 mutant was significantly slowed down as was shown by our flash-induced fluorescence and thermoluminescence measurements (Fig. 1C and D). The slow electron transfer from QA− to QB and the dominating QA− S2 state recombination band are both indicative of the limited electron transfer beyond the PSII complex in the thylakoid membrane of the C3 mutant. Thus, a low PSI content resulted in the reduced state of the PQ-pool at our light growth conditions (30 μmol photons per m2 per s1) in the C3 mutant.When the culture of the C3 mutant in the standard TAP medium was placed in the sealed bioreactors, anaerobic conditions were quickly established. The lower PSI content limited the PSII activity allowing respiration to overtake O2 evolution within 17 h (Fig. 2, open circles). This altered ratio of the two photosystems to 1:3 for PSI:PSII, providing favorable initial conditions for H2 production by the C3 mutant.
Tuning of photosynthesis during long-term H2 production
After anaerobiosis was established in the C3 mutant culture, H2 evolution commenced after 17 h (Fig. 2). Due to the specific starting conditions in the thylakoid membrane of the C3 mutant, this process took place in the complete TAP media without any deprivation procedure in a one-stage direct process. The remarkable result was that this H2 production continued for a long period of time (Fig. 2). Our experiments routinely showed that the H2 production continues for more than 40 days (6 weeks). To our knowledge, this is the longest direct H2 production by green algae under the standard cultivation conditions (light, growth medium, and absence of any nutrient manipulations) known so far.The rate of H2 production in the C3 mutant was not constant during these 40 days. During the first 6 days of H2 production, it increased 3-fold, and then during the next 6 days (12 days in total), it decreased to the initial value and stayed constant for the rest of the experiment (Table 1). These unique H2 production properties are based on changes in the amount and activity of both photosystems and the retuning of the overall electron transport from PSII to HydA.
Although the amount of PSII did not change during the first 2 days of H2 production, these PSII centers become more active as was shown by the complete reduction of the PQ pool (Fig. 3C and Fig. 2S, C†). In addition, more LHCII was re-associated with these PSII centers during this time in order to help its activation (Fig. 1S, B†). This boost in the PSII activity was, most probably, initiated by an increase in the amount of PSI which was occurring steadily during the first 12 days of H2 production (Table 1 and Fig. 3B, 4). This allowed relief of the reductive electron pressure produced by PSII and opened the electron flow from PSII to HydA as was observed by fluorescence and thermoluminescence measurements (Fig. 3C and Fig. 2S, D–F†). Thus, the initial increase in the PSI amount is responsible for the high rates of H2 production during the first 12 days (Fig. 2).
In the next 18 days (30 days in total), the burden of the long-term anaerobic incubation started to be seen as the internal cell supplies became exhausted. The amount of PSI started to decrease consequently with the amount of PSII which also had significantly diminished to less than 15% (Table 1 and Fig. 3A). It was accompanied by the decrease in the associated antenna and loss of the total Chl (Table 1 and Fig. 1S†). Taken together, these resulted in the decrease in the H2 production rates (Table 1 and Fig. 2).
Our data show that fine tuning of the PSI/PII ratio is needed not only for initiation but also for prolongation of the direct H2 production in one-stage process. While in the beginning of H2 production, there was an excess of PSII over PSI, after 2–6 days, the amount of PSI had equalized with PSII. After 6 days, it had exceeded the amount of PSII, reaching a PSI/PSII ratio of more than 5 (Fig. 4 and Table 1). Thus, a small amount of PSI is required to quickly initiate H2 production in the C3 mutant and then an excess of PSI over PSII is needed at the later stages for long-term production.
Interestingly, the amount of PSI correlated with the rate of H2 production in the C3 mutant (Table 1). This is not surprising since PSI is indispensable for light-induced H2 production in C. reinhardtii.48 In addition to the important role of PSII in H2 production as a primary electron source, our experiments also show that by changing the PSI properties, H2 production could be initiated and supported for at least 42 days. This introduces a new approach to achieve sustainable, long-term H2 fuel production from solar energy in green algae.
Conflicts of interest
There are no conflicts to declare.Acknowledgements
The authors are grateful to the Carl Tryggers Foundation (grant number CTS14:314 and CTS15:319) for financial support and Prof. Tony Miller, John Innes Centre, UK for the critical reading of the manuscript.References
- V. Smil, Futurist, 2008, 42, 58–58 Search PubMed.
- A. Magnuson, M. Anderlund, O. Johansson, P. Lindblad, R. Lomoth, T. Polivka, S. Ott, K. Stensjo, S. Styring, V. Sundstrom and L. Hammarstrom, Acc. Chem. Res., 2009, 42, 1899–1909 CrossRef CAS PubMed.
- B. Hankamer, F. Lehr, J. Rupprecht, J. H. Mussgnug, C. Posten and O. Kruse, Physiol. Plant., 2007, 131, 10–21 CrossRef CAS PubMed.
- M. Oey, A. L. Sawyer, I. L. Ross and B. Hankamer, Plant Biotechnol. J., 2016, 14, 1487–1499 CrossRef CAS PubMed.
- N. Z. Muradov and T. N. Veziroglu, Int. J. Hydrogen Energy, 2005, 30, 225–237 CrossRef CAS.
- H. Gaffron and J. Rubin, J. Gen. Physiol., 1942, 26, 219–240 CrossRef CAS PubMed.
- D. J. Vinyard and G. W. Brudvig, in Annu. Rev. Phys. Chem., ed. M. A. Johnson and T. J. Martinez, 2017, vol. 68, pp. 101–116 Search PubMed.
- S. T. Stripp and T. Happe, Dalton Trans., 2009, 9960–9969, 10.1039/b916246a.
- T. Happe and A. Kaminski, Eur. J. Biochem., 2002, 269, 1022–1032 CrossRef CAS PubMed.
- M. L. Ghirardi, R. K. Togasaki and M. Seibert, Appl. Biochem. Biotechnol., 1997, 63–65, 141–151 CrossRef CAS PubMed.
- A. Melis, L. Zhang, M. Forestier, M. L. Ghirardi and M. Seibert, J. Plant Physiol., 2000, 122, 127–136 CrossRef CAS PubMed.
- A. Volgusheva, S. Styring and F. Mamedov, Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 7223–7228 CrossRef CAS PubMed.
- F. Jans, E. Mignolet, P. A. Houyoux, P. Cardol, B. Ghysels, S. Cuine, L. Cournac, G. Peltier, C. Remacle and F. Franck, Proc. Natl. Acad. Sci. U. S. A., 2008, 105, 20546–20551 CrossRef CAS PubMed.
- M. Chen, J. Zhang, L. Zhao, J. Xing, L. Peng, T. Kuang, J. D. Rochaix and F. Huang, J. Integr. Plant Biol., 2016, 58, 943–946 CrossRef CAS PubMed.
- J. Steinbeck, D. Nikolova, R. Weingarten, X. Johnson, P. Richaud, G. Peltier, M. Hermann, L. Magneschi and M. Hippler, Front. Plant Sci., 2015, 6, 892 Search PubMed.
- A. Volgusheva, G. Kukarskikh, T. Krendeleva, A. Rubin and F. Mamedov, RSC Adv., 2015, 5, 5633–5637 RSC.
- G. Philipps, T. Happe and A. Hemschemeier, Planta, 2012, 235, 729–745 CrossRef CAS PubMed.
- K. A. Batyrova, A. A. Tsygankov and S. N. Kosourov, Int. J. Hydrogen Energy, 2012, 37, 8834–8839 CrossRef CAS.
- L. Zhang, T. Happe and A. Melis, Planta, 2002, 214, 552–561 CrossRef CAS PubMed.
- T. K. Antal, A. A. Volgusheva, G. P. Kukarskih, T. E. Krendeleva and A. B. Rubin, Int. J. Hydrogen Energy, 2009, 34, 9087–9094 CrossRef CAS.
- M. L. Ghirardi, A. Dubini, J. Yu and P. C. Maness, Chem. Soc. Rev., 2009, 38, 52–61 RSC.
- V. Chochois, D. Dauvillee, A. Beyly, D. Tolleter, S. Cuine, H. Timpano, S. Ball, L. Cournac and G. Peltier, J. Plant Physiol., 2009, 151, 631–640 CrossRef CAS PubMed.
- M. Forestier, P. King, L. Zhang, M. Posewitz, S. Schwarzer, T. Happe, M. L. Ghirardi and M. Seibert, Eur. J. Biochem., 2003, 270, 2750–2758 CrossRef CAS PubMed.
- A. Scoma, L. Durante, L. Bertin and F. Fava, New Phytol., 2014, 204, 890–900 CrossRef CAS PubMed.
- T. V. Laurinavichene, S. N. Kosourov, M. L. Ghirardi, M. Seibert and A. A. Tsygankov, J. Biotechnol., 2008, 134, 275–277 CAS.
- S. Oncel and F. Vardar-Sukan, Int. J. Hydrogen Energy, 2009, 34, 7592–7602 CrossRef CAS.
- T. Yagi, K. Yamashita, N. Okada, T. Isono, D. Momose, S. Mineki and E. Tokunaga, J. Plant Res., 2016, 129, 771–779 CrossRef CAS PubMed.
- A. S. Fedorov, S. Kosourov, M. L. Ghirardi and M. Seibert, Appl. Biochem. Biotechnol., 2005, 121–124, 403–412 CrossRef CAS PubMed.
- M. Oey, I. L. Ross, E. Stephens, J. Steinbeck, J. Wolf, K. A. Radzun, J. Kugler, A. K. Ringsmuth, O. Kruse and B. Hankamer, PLoS One, 2013, 8, e61375 CrossRef CAS PubMed.
- S. Grewe, M. Ballottari, M. Alcocer, C. D'Andrea, O. Blifernez-Klassen, B. Hankamer, J. H. Mussgnug, R. Bassi and O. Kruse, Plant Cell, 2014, 26, 1598–1611 CrossRef CAS PubMed.
- H. Berberoglu, L. Pilon and A. Melis, Int. J. Hydrogen Energy, 2008, 33, 6467–6483 CrossRef CAS.
- A. Scoma, D. Krawietz, C. Faraloni, L. Giannelli, T. Happe and G. Torzillo, J. Biotechnol., 2012, 157, 613–619 CrossRef CAS PubMed.
- K. Batyrova and P. C. Hallenbeck, Int. J. Mol. Sci., 2017, 18, 647 CrossRef PubMed.
- O. Kruse, J. Rupprecht, K. P. Bader, S. Thomas-Hall, P. M. Schenk, G. Finazzi and B. Hankamer, J. Biol. Chem., 2005, 280, 34170–34177 CrossRef CAS PubMed.
- J. Marin-Navarro, M. G. Esquivel and J. Moreno, World J. Microbiol. Biotechnol., 2010, 26, 1785–1793 CrossRef CAS.
- A. Volgusheva, O. Kruse, S. Styring and F. Mamedov, Algal Res., 2016, 18, 296–304 CrossRef.
- J. Girard-Bascou, Curr. Genet., 1987, 12, 483–488 CrossRef CAS.
- R. Danielsson, P. A. Albertsson, F. Mamedov and S. Styring, Biochim. Biophys. Acta, Bioenerg., 2004, 1608, 53–61 CrossRef CAS PubMed.
- I. Vass, D. Kirilovsky and A. L. Etienne, Biochemistry, 1999, 38, 12786–12794 CrossRef CAS PubMed.
- F. Mamedov, H. Stefansson, P. A. Albertsson and S. Styring, Biochemistry, 2000, 39, 10478–10486 CrossRef CAS PubMed.
- J. L. Roose, L. K. Frankel and T. M. Bricker, Biochemistry, 2010, 49, 36–41 CrossRef CAS PubMed.
- I. Vass, Photosynth. Res., 2003, 76, 303–318 CrossRef CAS PubMed.
- J. M. Ducruet, V. Peeva and M. Havaux, Photosynth. Res., 2007, 93, 159–171 CrossRef CAS PubMed.
- A. R. Crofts and C. A. Wraight, Biochim. Biophys. Acta, Rev. Bioenerg., 1983, 726, 149–185 CrossRef CAS.
- K. Cser, Z. Deak, A. Telfer, J. Barber and I. Vass, Photosynth. Res., 2008, 98, 131–140 CrossRef CAS PubMed.
- S. Dementin, F. Leroux, L. Cournac, A. L. de Lacey, A. Volbeda, C. Leger, B. Burlat, N. Martinez, S. Champ, L. Martin, O. Sanganas, M. Haumann, V. M. Fernandez, B. Guigliarelli, J. C. Fontecilla-Camps and M. Rousset, J. Am. Chem. Soc., 2009, 131, 10156–10164 CrossRef CAS PubMed.
- M. T. Stiebritz and M. Reiher, Chem. Sci., 2012, 3, 1739–1751 RSC.
- K. Redding, L. Cournac, I. R. Vassiliev, J. H. Golbeck, G. Peltier and J. D. Rochaix, J. Biol. Chem., 1999, 274, 10466–10473 CrossRef CAS PubMed.
- D. S. Gorman and R. P. Levine, Proc. Natl. Acad. Sci. U. S. A., 1965, 54, 1665–1669 CrossRef CAS PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c9gc01416k |
| ‡ Present address: Department of Molecular Microbiology, John Innes Centre, Norwich, NR4 7UH, United Kingdom. |
| This journal is © The Royal Society of Chemistry 2019 |