
Effects of dietary gamma-aminobutyric acid supplementation on the intestinal functions in weaning piglets†
Shuai
Chen
 ab,
Bie
Tan
*a,
Yaoyao
Xia
ab,
Simeng
Liao
ab,
Meiwei
Wang
c,
Jie
Yin
ab,
Jing
Wang
ab,
Hao
Xiao
ab,
Ming
Qi
ab,
Peng
Bin
ab,
Gang
Liu
a,
Wenkai
Ren
*d and
Yulong
Yin
acd
ab,
Bie
Tan
*a,
Yaoyao
Xia
ab,
Simeng
Liao
ab,
Meiwei
Wang
c,
Jie
Yin
ab,
Jing
Wang
ab,
Hao
Xiao
ab,
Ming
Qi
ab,
Peng
Bin
ab,
Gang
Liu
a,
Wenkai
Ren
*d and
Yulong
Yin
acd
aLaboratory of Animal Nutritional Physiology and Metabolic Process, Key Laboratory of Agro-ecological Processes in Subtropical Region, National Engineering Laboratory for Pollution Control and Waste Utilization in Livestock and Poultry Production, Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha, 410125, Hunan, China. E-mail: bietan@isa.ac.cn
bUniversity of Chinese Academy of Sciences, Beijing, China
cAnimal Nutrition and Human Health Laboratory, School of Life Sciences, Hunan Normal University, Changsha, 410081, Hunan, China
dGuangdong Provincial Key Laboratory of Animal Nutrition Control, Institute of Subtropical Animal Nutrition and Feed, College of Animal Science, South China Agricultural University, Guangzhou, China. E-mail: renwenkai19@126.com
First published on 14th December 2018
Abstract
This study aims to investigate the effects of dietary gamma-aminobutyric acid (GABA) supplementation on the growth performance, intestinal immunity, intestinal GABAergic system, amino acid profiles and gut microflora of the weaned piglets. Totally sixteen healthy piglets were randomly assigned into two groups to be fed with the basal diet (Con group) or the basal diet with GABA (20 mg kg−1) supplementation. Body weights and feed intakes were monitored weekly. Piglets were sacrificed after 3 weeks of GABA supplementation to collect the blood, ileum, ileal mucosa and luminal content. Immune-associated factors, GABAergic system, amino acid profiles, and microbiota in the ileum and serum amino acid profiles were explored. The results showed that GABA supplementation improved the growth performance and modulated the intestinal immunity with inhibiting the gene expressions of IL-22, proinflammatory cytokines (IL-1 and IL-18), and Muc1, but promoted the expressions of anti-inflammatory cytokines (IFN-γ, IL-4, and IL-10), TLR6 and MyD88. GABA regulated a few components of the intestinal GABAergic system, increased the levels of most amino acids in the ileal mucosa but reduced the serum amino acid profiles. GABA regulated the population and diversity of intestinal microbiota, such as the abundances of the dominant microbial populations, the community richness, and diversity of the ileal microbiota. In conclusion, GABA supplementation modulated the intestinal functions, including intestinal immunity, intestinal amino acid profiles and gut microbiota, and the results can be helpful for understanding the functions of GABA in the intestine.
Introduction
Piglet postnatal growth and development are very susceptible to multiple stressors (e.g., weaning), which have adverse effects on the intestinal health and growth performance, leading to great economic losses to the swine husbandry. Reduction in the nutritional intake during weaning is correlated with hyperplasia of the crypt, atrophy of the villi and reduction of the mucosal enzyme activity. Weaning reduces the intestinal integrity by increasing the intestinal permeability and disturbing the tight junctions, and increases the invasion of dietary and microbial antigen (e.g., lipopolysaccharide from Escherichia coli) resulting in inflammation and excess immune responses.1 Furthermore, weaning alters the intestinal microbiota, such as reducing the diversity of intestinal microbiota and the ratio of Lactobacilli to Enterobacteria.2 Thus, appropriate strategies should be adapted to prevent the weaning stress.Numerous studies have shown that an appropriate nutritional strategy is an effective way to prevent weaning stress. Amino acids, such as glutamine (Gln), glutamate (Glu) and arginine, are reported to promote growth and health in animals and humans.3,4 Gln is a crucial energy source for the gut, and acts as a modulator of the microbial community, intestinal immunity and metabolism of amino acids.5 For example, dietary supplementation of 1% to 4% Gln for 3 weeks increased the growth performance, jejunal villus height and levels of serum amino acids, while reduced the incidences of diarrhea.6 Glu, which could be generated from Gln by glutaminase, improves the intestinal integrity and function in weaning piglets, and the growth performance in growth-finishing pigs.7,8 Our previous studies have also shown that Gln or Glu improves the growth performance, gut function and microbiota during oxidative stress or infection in multiple animal models.9–11 Other amino acids, like gamma-aminobutyric acid (GABA), also show regulatory roles in the intestinal health and immunity.12 Thus, dietary amino acid supplementation could be an effective way to prevent weaning stress.
GABA is synthesized through the decarboxylation of Glu, catalyzed by glutamic acid decarboxylase (GAD), and is widely distributed in the nervous system to reduce the activity of neurons by binding to GABA receptors. GABA plays a multifunctional role in regulating the intestinal movement, carcinogenesis, pain processing and inflammation, dependent or independent on the GABAergic system in the enteral nervous system (ENS).13–16 GABAergic system constitutes of GABA, GADs, GABA transporters (GATs) and GABA receptors (GABARs). GADs (including GAD1 and GAD2) and GATs (including solute carrier family 6 member 1 (SLC6A1) and SLC6A11–13) synergistically maintain the concentration of endocellular and extracellular GABA.17,18 GABA receptors consist of ionotropic receptors, including GABAA receptors (a complex constituted of several subunits, including alpha 1–6, beta 1–3, gamma 1–3, delta, epsilon, theta, pi, and rho 1–3), GABAC receptors, and metabotropic GABAB receptor (a G protein-coupled receptor consisting of two subunits – GABAB1 and GABAB2).19–21 Recently, it is reported that GABA also plays important roles in the growth and hormonal generation and secretion. For example, dietary GABA enhanced the feed intakes and gene expressions of GABA receptors, while inhibited the expression of cholecystokinin-related gene in growing lambs.22 Dietary GABA supplementation also improved the feed intakes and milk protein concentrations in transitioning cows.23 Our previous study demonstrated that the gut microbiota-generated GABA promoted the intestinal expression of IL-17 during ETEC infection in mice.24 Those studies indicate the potential beneficial roles of GABA in piglets during the weaning stress. We hypothesize that GABA could alleviate the piglet weaning stress through modulating the gut immunity, the intestinal GABAergic system, the amino acid metabolism, and the intestinal microbiota. Thus, this study aimed to analyze the effects of dietary GABA supplementation on the growth performance, intestinal immunity, intestinal GABAergic system, amino acid profiles and gut microflora of the weaning piglets.
Materials and methods
Animals and experiment design
A total of sixteen healthy piglets (Duroc × Landrace × Landrace, 21 days old) were purchased from Hunan new wellful Co., Ltd (China, Changsha). After an adaptation period of 3 d, piglets with an average body weight of 6.43 kg were assigned into two groups randomly: (1) GABA group: Piglets were fed on a corn- and soybean meal-based diet (Table S1†) with 20 mg kg−1 of GABA supplementation; (2) control group: Piglets were fed on a corn- and soybean meal-based diet without GABA supplementation. The basal diet was formulated according to the National Research Council (2012) requirements for piglets. GABA (Ajinomoto, Tokyo, Japan) was added to the basal diet at the expense of corn starch, and the dosage for GABA supplementation and the experimental period were selected according to previous studies.25,26 The dietary contents of dry matter, crude protein, crude fiber, net energy and digestive energy in the basal diet were determined according to the methods of the Association of Official Analytical Chemists (1996). The contents of amino acids in the basal diet were determined by high-performance liquid chromatography (HPLC) according to our previous study.27The piglets were housed individually in an environmentally-controlled facility with hard plastic slatted flooring, and maintained at an ambient temperature of 25 ± 2 °C with free access to diets and drinking water. Body weight and feed intake were monitored weekly through the whole experiment. After 3 weeks of GABA supplementation, piglets were sacrificed after anesthesia. Weights of the organs, including the heart, liver, spleen, lungs and kidneys, were monitored and recorded, while the blood, jejunum, ileum, ileal mucosa and luminal content of ilea were collected immediately, snap-frozen in liquid nitrogen and stored at −80 °C until further processing. All animal experiment procedures were approved by the Animal Welfare Committee of the Institute of Subtropical Agriculture, Chinese Academy of Sciences (2016-4A).
Growth performance and organ index
Body weights and feed intakes were measured weekly. The growth rate is the ratio of weekly body weight gain to the initial body weight, while the feed conversion ratio is the ratio of the weekly feed intake to the weekly body weight gain. Organ index is the relative weight of an organ, and is calculated by dividing the initial body weight by the organ weight.Free amino acids analysis
Ileal mucosal and serum free amino acids were analyzed by HPLC. The serum or the homogenate of ileal mucosa was centrifuged with 0.01 M hydrochloric acid solution, and then, the supernatant was thoroughly homogenized with 0.8% sulfosalicylic acid solution. After incubating overnight at 4 °C, the mixture was centrifuged and filtered before being analyzed by HPLC according to the instructions of the manufacturer. Authentic standards (Sigma Chemicals) were used to quantify the amino acids in the samples.Gene expression analysis
The expressions of GABAergic system and immune-associated genes were analyzed by RT-PCR according to our previous study.28 Briefly, liquid nitrogen-frozen jejunal and ileal samples were ground to extract the total RNA using the TRIZOL reagent (Invitrogen, USA), according to the manufacturer's recommendation. Total RNA was used for the synthesis of complementary DNA using the PrimeScript RT reagent kit with gDNA Eraser (TAKARA BIO INC., Qingdao, China) according to the product manual, following processing with DNase I (Invitrogen, USA). RT-PCR was performed on an Applied Biosystems 7900HT Fast Real-Time PCR System (Thermo, USA) according to our previous study.29 Primers (Table S2†) used in this study were designed under the principles of primer design with the Primer 6.0 software (PRIMER-E, New Zealand) and the Oligo 5.0 software (Molecular Biology Insights, Inc., USA) based on the gene sequences of Sus scrofa on GenBank. The house-keeping genes, beta-actin (ACTB) and glyceraldehyde phosphate dehydrogenase (GAPDH) were chosen for the normalization of all target gene levels, and the cycling conditions of RT-PCR were 94 °C for 40 s (40 cycles), 60 °C for 30 s (1 cycle) and 72 °C for 35 s (1 cycle). The relative gene expression was represented by the ratio of the target gene to the control gene using the formula 2−(ΔΔCt), where ΔΔCt = (CtTarget − Cthouse-keeping genes)treatment− (CtTarget − Cthouse-keeping genes)control. Normalization of the relative gene expression was expressed as a ratio of the expression of the GABA group to the controls.5,30Bacteria 16S rDNA sequencing and bioinformatics
The DNAs of the luminal contents of the ilea were extracted with the Qiagen QIAamp DNA Stool Mini Kit according to the manufacturer's protocol. Agarose gel electrophoresis was used to quantify the DNAs from the luminal contents and the amplification products. Illumina MiSeq Sequencer was used for the V3 to V4 regions of bacterial 16S rDNA sequencing. Sequencing libraries were generated using Ion Plus Fragment Library Kit 48 rxns (Thermo Scientific) following the manufacturer's recommendations. The library quality was assessed on the Qubit@2.0 Fluorometer (Thermo Scientific). The library was sequenced on an Ion S5TM XL platform and 400 bp/600 bp single-end reads were generated. Single-end reads were assigned to the samples based on their unique barcode and truncated by cutting off the barcode and the primer sequence. Quality filtering on the raw reads was performed under specific filtering conditions to obtain the high-quality clean reads according to the Cutadapt quality controlled process. Sequence analysis and operational taxonomic unit (OTU) clustering were performed by Uparse software (Uparse v7.0.1001), and the sequences with more than 97% similarity were clustered to the same OTU. RDP Classifier (V2.2, Michigan State University Board of Trustees, East Lansing MI) was used for the species annotations, based on the GreenGene database. The MUSCLE software (Version 3.8.31) was used to study the phylogenetic relationships of different OTUs, and the differences in the dominant species in different samples (groups), and for multiple sequence alignments. Abundant information in the OTUs was normalized using a standard sequence number corresponding to the sample with the least sequences. Subsequent analysis of the alpha diversity, and beta diversity and the environmental factor correlation analysis were all performed based on this normalized output data by R software and Qiime software (V 1.7). Two databases, including FAPROTAX and Tax4Fun, were used for the function prediction. Illumina MiSeq sequencing, processing of sequencing data and bioinformatics analysis were performed by Beijing Novogene Bioinformatics Technology Co., Ltd (Beijing, China).Statistical analyses
All data were pre-processed with Excel 2016 (Microsoft, Redmond, USA) and statistically analyzed by SAS version 9.4 (SAS Institute Inc., Cary, NC, USA). Word 2016 software (Microsoft, Redmond, USA) was used to prepare tables and GraphPad Prism 7.0 (GraphPad Software, Inc., La Jolla, CA) was used to generate figures. Unpaired t-test was used for the statistical analysis between the two groups, if the data followed a normal distribution; otherwise, Wilcoxon signed-rank test was used for the analysis. The results were presented as mean ± standard error of the mean (SEM) and a P value <0.05 was considered as statistically significant.31–33Results
GABA supplementation improves the growth performance of weaning piglets
Firstly, we analyzed the effects of GABA on growth performance. The results showed that the body weight and the average daily feed intake were similar between the control group and the GABA group (Fig. 1A and C). However, dietary GABA supplementation significantly promoted the growth rate in the third week (P < 0.05) and reduced the feed conversion ratio in the second week (P < 0.05) (Fig. 1B and D ). Then, we analyzed the indexes of the organs, including the heart, liver, spleen, lung and kidney, and the results illustrated that GABA administration significantly promoted the organ index of kidney (P < 0.05), while it had no effect on the indexes of the other organs (Table 1). | ||
| Fig. 1 Effects of GABA on the growth performance of the piglets. A, body weight; B, growth rate; C, average daily feed intake; D, feed conversion ratio. Unpaired t-test was used for analyzing the data (N = 8; mean ± SEM). **P < 0.05. | ||
| Con | GABA | t value | P value | |
|---|---|---|---|---|
| Organ index: The ratio of organ weight to the body weight. The organ indexes of the heart, liver, spleen, lungs and kidney were analyzed. Piglets from the control group (Con, n = 8) and the GABA group (GABA, n = 8) were given access to the basal diet without and with supplementation of GABA, separately. Unpaired t-test was used for analyzing the data (mean ± SEM). *P < 0.05. | ||||
| Heart | 0.477 ± 0.011 | 0.466 ± 0.011 | 0.652 | 0.523 |
| Liver | 2.664 ± 0.048 | 2.549 ± 0.07 | 1.385 | 0.177 |
| Spleen | 0.185 ± 0.005 | 0.196 ± 0.009 | −1.092 | 0.287 |
| Lungs | 1.104 ± 0.033 | 1.174 ± 0.049 | −1.191 | 0.243 |
| Kidney | 0.453 ± 0.012 | 0.507 ± 0.012* | −3.213 | 0.003 |
Dietary GABA plays a modulatory role in the intestinal immunity
To test the effects of dietary GABA on gut immunity, we performed RT-PCR analysis to test the expressions of immunity-related factors, including toll-like receptors (TLR)-2, 4, 5, and 6, myeloid differential protein-88 (MyD88), polymeric immunoglobulin receptor (pIgR), mucin (Muc) 1, 2, and 4, tumor necrosis factor alpha (TNF-α), interferon gamma (IFN-γ), and interleukin (IL)-1, 2, 4, 6, 10, 17, 18, and 22. The mRNA expressions of IL-1, 18 and 22, and Muc1 were inhibited (P < 0.05) and the mRNA levels of TNF-α, IFN-γ, IL-10, TLR-6 and MyD88 were increased (P < 0.05) in the ilea of piglets (Fig. 2). The expressions of other factors were similar between the control and GABA groups (Fig. 2).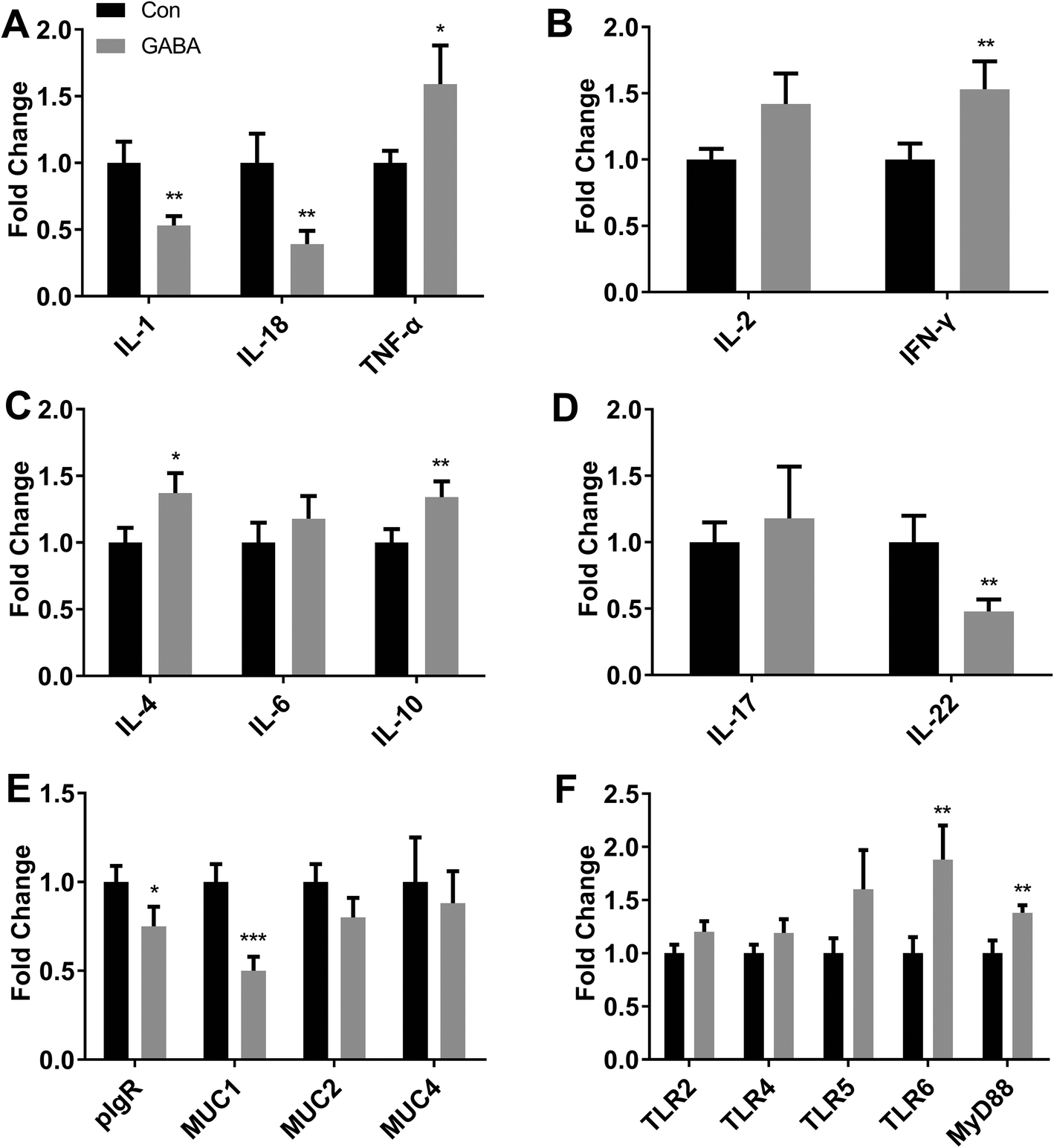 | ||
| Fig. 2 Ileal mRNA expressions of immune related factors in the piglets. Relative gene expressions of IL-1, 18, and TNF-α (A), IL-2 and IFN-γ (B), IL-4, 6, 8 (C), IL-17, 22 (D), pIgR, Muc1, 2, 4 (E), Tlr2, 4, 5, 6, and MyD88 (F) were analyzed by RT-PCR. N = 8, unpaired t-test was used for analyzing the data (mean ± SEM). *0.1 < P < 0.05; **P < 0.05; ***P < 0.01. | ||
Dietary GABA regulates the intestinal GABAergic system
Gene expression of gut GABAergic system was further detected by RT-PCR. GATs (including SLC6A1, SLC6A11, SLC6A12 and SLC6A13) were expressed in the ileum of piglets, and GABA remarkably inhibited (P < 0.05) ileal expression of SLC6A13 (Table 2). The gene expressions of GABAB1–2, GABAAα1–5, GABAAβ2, GABAAγ1–2, GABAAδ, GABAAε, GABAAπ, GABAAθ and GABAAρ1 were detected, and GABA suppressed the mRNA expressions of GABAB2 and GABAAβ2 (P < 0.05), while it showed a slight trend of inhibition on the GABAAθ expression (P = 0.0952) (Table 2). The results show that GATs and GABA receptors but not GADs were expressed in the small intestines of the piglets and GABA could modify the small intestinal GABAergic system.| Con | GABA | t value | P value | |
|---|---|---|---|---|
| Ileal gene expressions of GAT, GABA receptors and GAD (undetected) were analyzed by RT-PCR. Piglets from the control group (Con, n = 8) and the GABA group (GABA, n = 8) were given access to the basal diet supplemented without and with GABA. Unpaired t-test was used for analyzing the date (mean ± SEM). *P < 0.05. | ||||
| SLC6A1 | 1.00 ± 0.17 | 1.24 ± 0.18 | −0.96 | 0.351 |
| SLC6A11 | 1.00 ± 0.10 | 1.14 ± 0.12 | −0.91 | 0.377 |
| SLC6A12 | 1.00 ± 0.06 | 1.03 ± 0.20 | −0.14 | 0.895 |
| SLC6A13 | 1.00 ± 0.12 | 0.69 ± 0.08* | 2.27 | 0.041 |
| GABAB1 | 1.00 ± 0.11 | 0.77 ± 0.10 | 1.59 | 0.137 |
| GABAB2 | 1.00 ± 0.21 | 0.45 ± 0.05* | 2.56 | 0.039 |
| GABAAβ2 | 1.00 ± 0.16 | 0.58 ± 0.05* | 2.56 | 0.038 |
| GABAAδ | 1.00 ± 0.12 | 1.08 ± 0.13 | −0.46 | 0.656 |
| GABAAε | 1.00 ± 0.23 | 1.02 ± 0.06 | −0.10 | 0.923 |
| GABAAρ1 | 1.00 ± 0.22 | 0.7 ± 0.08 | 1.31 | 0.224 |
| GABAAπ | 1.00 ± 0.14 | 0.84 ± 0.22 | 0.64 | 0.536 |
| GABAAθ | 1.00 ± 0.17 | 0.66 ± 0.09 | 1.81 | 0.095 |
| GABAAγ1 | 1.00 ± 0.18 | 0.74 ± 0.14 | 1.17 | 0.264 |
| GABAAγ2 | 1.00 ± 0.13 | 0.89 ± 0.17 | 0.50 | 0.627 |
| GABAAα1 | 1.00 ± 0.03 | 1.03 ± 0.15 | −0.21 | 0.842 |
| GABAAα2 | 1.00 ± 0.30 | 0.49 ± 0.04 | 1.69 | 0.134 |
| GABAAα3 | 1.00 ± 0.13 | 0.91 ± 0.07 | 0.58 | 0.574 |
| GABAAα4 | 1.00 ± 0.17 | 1.02 ± 0.16 | −0.06 | 0.949 |
| GABAAα5 | 1.00 ± 0.09 | 1.02 ± 0.19 | −0.11 | 0.916 |
Dietary GABA promotes ileal mucosal amino acids, but reduces serum amino acids
The results of intestinal mucosal amino acid profiles were shown in Table 3. GABA significantly improved (P < 0.05) the concentrations of 11 amino acids, including L-alanine, L-phenylalanine, L-tryptophan, L-proline, L-serine, L-threonine, L-cystine, L-tyrosine, L-histidine, L-ornithine, L-carnosine, and showed a trend to increase the levels of GABA, L-leucine, L-isoleucine and L-aspartic acid (P < 0.05) in the ileal mucosa. GABA administration significantly (P < 0.05) lowered the levels of DL-α-amino-n-butyric acid, o-phosphoethanolamine and hydroxy-L-proline in the ileal mucosa. Then, we detected the serum profiles of 18 amino acids by HPLC. The results revealed that the concentrations of L-histidine, L-serine, L-arginine, L-alanine, L-proline, L-cystine, L-lysine, L-methionine, L-isoleucine, L-leucine, L-phenylalanine were notably lower (P < 0.05) in the sera of piglets in the GABA group than those of the controls (Table 3). These results show that the dietary GABA supplementation increases the levels of most amino acids in the ileal mucosa but reduces those in the serum, indicating that GABA might affect amino acid absorption and metabolism.| Con | GABA | t value | P value | |
|---|---|---|---|---|
| Amino acid profiles of the ileal mucosa and sera were detected by HPLC. Piglets from the control group (Con, n = 8) and the GABA group (GABA, n = 8) were given access to the basal diet supplemented without and with GABA. Unpaired t-test was used for analyzing the data (mean ± SEM). *P < 0.05. | ||||
| Ileal mucosa | ||||
| L-Alanine | 283.97 ± 6.73 | 328.63 ± 25.78 | −1.68 | 0.133 |
| L-Valine | 92.79 ± 9.38 | 118.86 ± 5.39* | −2.20 | 0.049 |
| L-Leucine | 224.38 ± 15.99 | 265.48 ± 8.97 | −2.04 | 0.064 |
| L-Isoleucine | 108.69 ± 8.43 | 132.32 ± 5.75 | −2.15 | 0.053 |
| L-Phenylalanine | 132.41 ± 8.88 | 163.02 ± 4.32* | −2.78 | 0.017 |
| L-Tryptophan | 21.47 ± 1.68 | 29.46 ± 1.43* | −3.46 | 0.005 |
| L-Methionine | 88.82 ± 8.02 | 103.22 ± 2.18 | −1.73 | 0.121 |
| L-Proline | 136.36 ± 11.68 | 188.33 ± 12.16* | −3.08 | 0.010 |
| Glycine | 509.37 ± 47.76 | 585.33 ± 47.72 | −1.13 | 0.279 |
| L-Serine | 179.35 ± 12.04 | 236.95 ± 10.46* | −3.46 | 0.005 |
| L-Threonine | 97.76 ± 10.76 | 131.16 ± 5.61* | −2.49 | 0.029 |
| L-Cystine | 24.24 ± 2.23 | 50.53 ± 3.35* | −6.52 | 0.000 |
| L-Tyrosine | 125.18 ± 9.77 | 156.85 ± 2.54* | −3.14 | 0.014 |
| L-Aspartic acid | 242.8 ± 15.61 | 301.01 ± 24.2 | −2.02 | 0.063 |
| L-Glutamic acid | 1183.58 ± 56.45 | 1208.12 ± 55.96 | −0.31 | 0.762 |
| L-Lysine | 148.06 ± 12.42 | 157.73 ± 17.03 | −0.47 | 0.648 |
| L-Arginine | 131.12 ± 12.22 | 144.42 ± 16.4 | −0.66 | 0.521 |
| L-Histidine | 45.74 ± 2.23 | 67.32 ± 8.09* | −2.57 | 0.033 |
| L-Ornithine | 25.15 ± 1.25 | 30.51 ± 0.91* | −3.47 | 0.005 |
| L-Citrulline | 73.17 ± 5.46 | 95.28 ± 9.08 | −2.09 | 0.056 |
| β-Alanine | 23.68 ± 1.67 | 24.9 ± 0.48 | −0.71 | 0.499 |
| o-Phospho-L-serine | 19.42 ± 1.15 | 20.34 ± 1.31 | −0.53 | 0.606 |
| L-1-Methylhistidine | 0.65 ± 0.15 | 0.68 ± 0.26 | −0.09 | 0.933 |
| L-3-Methylhistidine | 7.81 ± 1.52 | 10.69 ± 0.71 | −1.63 | 0.127 |
| L-Alpha-aminoadipic acid | 85.42 ± 4.04 | 94.54 ± 4.57 | −1.50 | 0.157 |
| DL-α-Amino-n-butyric acid | 10.31 ± 0.99 | 0.00 ± 0.00* | 10.41 | 0.000 |
| DL-β-Aminoisobutyric acid | 49.19 ± 3.86 | 41.45 ± 1.63 | 1.85 | 0.096 |
| GABA | 12.2 ± 0.52 | 14.15 ± 0.79 | −1.98 | 0.069 |
| Taurine | 267.44 ± 27.86 | 227.21 ± 6.81 | 1.40 | 0.199 |
| o-Phosphoethanolamine | 247.32 ± 40.8 | 143.93 ± 6.43* | 2.50 | 0.039 |
| Sarcosine | 32.33 ± 1.73 | 31.93 ± 1.30 | 0.18 | 0.856 |
| L-Anserine | 140.89 ± 24.74 | 162.5 ± 19.7 | −0.68 | 0.506 |
| L-Cystathionine | 26.84 ± 1.79 | 28.88 ± 1.41 | −0.91 | 0.382 |
| Ethanolamine | 59.75 ± 9.01 | 72.5 ± 4.29 | −1.28 | 0.222 |
| Hydroxy-L-proline | 24.9 ± 1.28 | 20.49 ± 1.44* | 2.28 | 0.039 |
| DL-Plus allo-llhydroxylysine | 7.24 ± 0.36 | 7.05 ± 0.24 | 0.44 | 0.665 |
| Urea | 38.19 ± 1.5 | 46.48 ± 1.51* | −3.87 | 0.002 |
| L-Carnosine | 454.16 ± 44.66 | 506.36 ± 17.05 | −1.28 | 0.226 |
| Serum | ||||
| L-Histidine | 7.01 ± 0.62 | 4.29 ± 0.05* | 4.37 | 0.005 |
| L-Serine | 16.87 ± 0.30 | 14.28 ± 0.51* | 4.16 | 0.002 |
| L-Arginine | 20.92 ± 0.76 | 17.98 ± 0.78* | 2.69 | 0.020 |
| Glycine | 42.92 ± 4.39 | 37.66 ± 2.32 | 1.01 | 0.329 |
| L-Aspartic acid | 4.3 ± 0.52 | 4.24 ± 0.50 | 0.08 | 0.939 |
| L-Glutamate | 42.33 ± 1.74 | 36.61 ± 3.89 | 1.34 | 0.201 |
| L-Threonine | 5.4 ± 0.15 | 6.68 ± 0.84 | −1.49 | 0.183 |
| L-Alanine | 42.84 ± 2.12 | 24.29 ± 0.76* | 8.25 | 0.000 |
| L-Proline | 30.17 ± 1.26 | 23.07 ± 1.04* | 4.34 | 0.001 |
| L-Cystine | 2.02 ± 0.41 | 0.95 ± 0.18* | 2.38 | 0.035 |
| L-Lysine | 42.93 ± 3.34 | 21.99 ± 1.02* | 5.99 | 0.000 |
| L-Tyrosine | 19.55 ± 1.54 | 15.19 ± 1.46 | 2.06 | 0.059 |
| L-Methionine | 9.49 ± 0.85 | 6.65 ± 0.91* | 2.28 | 0.039 |
| L-Valine | 19.41 ± 1.44 | 17.40 ± 1.42 | 0.99 | 0.340 |
| L-Isoleucine | 12.11 ± 0.67 | 9.41 ± 0.52* | 3.19 | 0.008 |
| L-Leucine | 17.83 ± 0.96 | 12.06 ± 0.20* | 5.91 | 0.002 |
| L-Phenylalanine | 12.71 ± 1.01 | 8.08 ± 0.46* | 3.98 | 0.002 |
| L-Tryptophan | 4.75 ± 0.21 | 5.48 ± 0.43 | −1.45 | 0.171 |
| GABA | 1.26 ± 0.10 | 1.26 ± 0.08 | 0.01 | 0.992 |
Dietary GABA supplementation regulates the intestinal microbiota
We then analyzed the effects of GABA on intestinal microbiota. The results generated from 16sRNA sequencing of thirteen samples, including Raw PE, Raw Tags, Clean Tags, Effective Tags, Base, Average Length, Q20, Q30, GC% and Effective%, were presented in Table S3.† The rarefaction curves (Fig. S1A and B†) and Good's-coverage (Table S4†) revealed that the sequencing data, sequencing depth and coverage met the demands of the further analysis. Venn diagram indicated that the ileal microbiota of both groups shared 410 OTUs, with 609 unique OUTs in GABA group and 103 unique OTUs in control group (Fig. S1C†). Relative abundances of the 10 or 15 of the most abundant members at different microbial taxonomy levels were presented in Fig. S2A and D (on phylum level), B and E (on class level), C and F (on genus level).† Firmicutes were the first dominant population in both groups (Table 4), and GABA reduced (P < 0.05) the relative abundances of Firmicutes, while increased (P < 0.05) the relative abundances of other members, such as Proteobacteria, Bacteroidetes, Cyanobacteria, Fusobacteria, Actinobacteria, Chloroflexi and Saccharibacteria, on the phylum level. GABA notably suppressed (P < 0.05) the growth of Clostridia and Clostridium_sensu_stricto_1, which were the dominant constituents at the class level and genus level, respectively, and promoted (P < 0.05) other microbial populations (Table 4). Thus, the dietary GABA supplementation modulated the relative abundances of the dominant microflora from the phylum level to the genus level.| Con | GABA | Z score | P value | |
|---|---|---|---|---|
| Top 10 microbial populations of the control group (Con, n = 6) and the GABA group (GABA, n = 7) at the phylum level and class level, and the top 15 microbial population of the Con (n = 6) and the GABA (n = 7) at the genus level. Unpaired t-test was used for analyzing the data (mean ± SEM). *P < 0.05. | ||||
| Phylum | ||||
| Firmicutes | 0.930 ± 0.027 | 0.609 ± 0.063* | 2.786 | 0.005 |
| Cyanobacteria | 0.007 ± 0.002 | 0.166 ± 0.040* | −2.929 | 0.003 |
| Tenericutes | 0.044 ± 0.023 | 0.093 ± 0.043 | −0.643 | 0.520 |
| Proteobacteria | 0.014 ± 0.004 | 0.101 ± 0.022* | −2.929 | 0.003 |
| Fusobacteria | 0.000 ± 0.000 | 0.006 ± 0.004* | −2.643 | 0.008 |
| Bacteroidetes | 0.002 ± 0.001 | 0.010 ± 0.002* | −2.786 | 0.005 |
| Actinobacteria | 0.002 ± 0.001 | 0.010 ± 0.002* | −2.929 | 0.003 |
| Chloroflexi | 0.000 ± 0.000 | 0.001 ± 0.001* | −2.362 | 0.018 |
| Saccharibacteria | 0.001 ± 0.001 | 0.002 ± 0.000 | −1.357 | 0.175 |
| Acidobacteria | 0.000 ± 0.000 | 0.001 ± 0.000 | −1.642 | 0.101 |
| Others | 0.000 ± 0.000 | 0.002 ± 0.001 | — | — |
| Class | ||||
| Clostridia | 0.810 ± 0.045 | 0.310 ± 0.087* | 2.786 | 0.005 |
| Bacilli | 0.081 ± 0.042 | 0.259 ± 0.076* | −2.214 | 0.027 |
| Chloroplast | 0.007 ± 0.002 | 0.165 ± 0.040* | −2.929 | 0.003 |
| Mollicutes | 0.044 ± 0.023 | 0.093 ± 0.043 | −0.643 | 0.520 |
| Gammaproteobacteria | 0.010 ± 0.003 | 0.079 ± 0.022* | −2.929 | 0.003 |
| Erysipelotrichia | 0.035 ± 0.026 | 0.023 ± 0.005 | −1.214 | 0.225 |
| Negativicutes | 0.003 ± 0.001 | 0.017 ± 0.005* | −2.357 | 0.018 |
| Fusobacteriia | 0.000 ± 0.000 | 0.006 ± 0.004* | −2.786 | 0.005 |
| Betaproteobacteria | 0.001 ± 0.000 | 0.010 ± 0.003* | −2.929 | 0.003 |
| Alphaproteobacteria | 0.001 ± 0.001 | 0.011 ± 0.002* | −2.929 | 0.003 |
| Others | 0.007 ± 0.003 | 0.026 ± 0.003 | — | — |
| Genus | ||||
| Clostridium_sensu_stricto_1 | 0.667 ± 0.063 | 0.138 ± 0.037* | 2.929 | 0.003 |
| Lactobacillus | 0.027 ± 0.008 | 0.157 ± 0.061 | −1.929 | 0.054 |
| Terrisporobacter | 0.033 ± 0.007 | 0.106 ± 0.049 | −0.214 | 0.830 |
| Unidentified_Chloroplast | 0.007 ± 0.002 | 0.165 ± 0.040* | −2.929 | 0.003 |
| Romboutsia | 0.104 ± 0.039 | 0.049 ± 0.009 | 1.071 | 0.284 |
| Mycoplasma | 0.044 ± 0.023 | 0.093 ± 0.043 | −0.643 | 0.520 |
| Enterococcus | 0.044 ± 0.043 | 0.005 ± 0.002 | −1.931 | 0.054 |
| Turicibacter | 0.034 ± 0.026 | 0.016 ± 0.005 | −1.071 | 0.284 |
| Halomonas | 0.001 ± 0.000 | 0.040 ± 0.019* | −2.643 | 0.008 |
| Streptococcus | 0.009 ± 0.004 | 0.047 ± 0.010* | −2.786 | 0.005 |
| Gemella | 0.001 ± 0.000 | 0.017 ± 0.011* | −2.643 | 0.008 |
| Weissella | 0.001 ± 0.000 | 0.015 ± 0.008* | −2.503 | 0.012 |
| Veillonella | 0.003 ± 0.001 | 0.016 ± 0.005* | −2.357 | 0.018 |
| Acinetobacter | 0.001 ± 0.000 | 0.014 ± 0.003* | −2.929 | 0.003 |
| Pediococcus | 0.000 ± 0.000 | 0.007 ± 0.003* | −2.074 | 0.038 |
| Others | 0.027 ± 0.007 | 0.115 ± 0.013 | — | — |
Estimators of community richness in the GABA group, such as observed species, Chao1 and Abundance-based Coverage Estimator (ACE), were significantly higher than those in the Con group (P < 0.05), and the Shannon index, the Simpson's index and phylogenetic diversity (PD) of the whole tree were also significantly improved (P < 0.05) upon GABA administration (Table S4†). These findings from the alpha diversity analysis indicated a regulatory role of GABA on enhancing the community richness and diversity of the intestinal microbiota, as rank abundance curves shown in Fig. S1D and E.† Then, the weighted UniFrac values were calculated, and GABA significantly improved (P < 0.05) this value (Fig. 3A). Results from the unweighted pair-group method with arithmetic means (UPGMA) (Fig. 3B), principal component analysis (PCA) (Fig. 3C), and nonmetric multidimensional scaling (NDMS) (Fig. 3D) also showed the difference in the UniFrac values. The function prediction indicated that dietary GABA altered the microbial function (Fig. 3E), such as increasing chemoheterotrophy, fermentation, chloroplasts and nitrate/nitrite/nitrogen respiration (Fig. 3F). Environmental factor correlation analysis revealed that the Firmicutes, the dominative population at the phylum level, were positively associated with the feed conversion ratio (Fig. 4A), the intestinal expression of pIgR and Muc1 (Fig. 4B), and the profiles of serum amino acid (including histidine, serine, arginine, alanine, proline, lysine, leucine and phenylalanine) (Fig. 4C and D), and negatively associated with the growth rate (Fig. 4A), the intestinal gene expression of IFNγ, Tlr6 and MyD88 (Fig. 4B), and the profiles of ileal mucosal amino acid (including cystine, histidine, proline, serine, threonine, tryptophan and tyrosine) (Fig. 4C and D). Other populations, such as Proteobacteria and Bacteroidetes, in the phylum showed an adverse relationship with the above-mentioned parameters (Fig. 4). These results demonstrated that dietary GABA supplementation could affect both alpha and beta diversity of the intestinal microbiota and change the microbial function, which were statistically correlated with the growth performance, intestinal immunity and amino acid profiles of the weaned piglets.
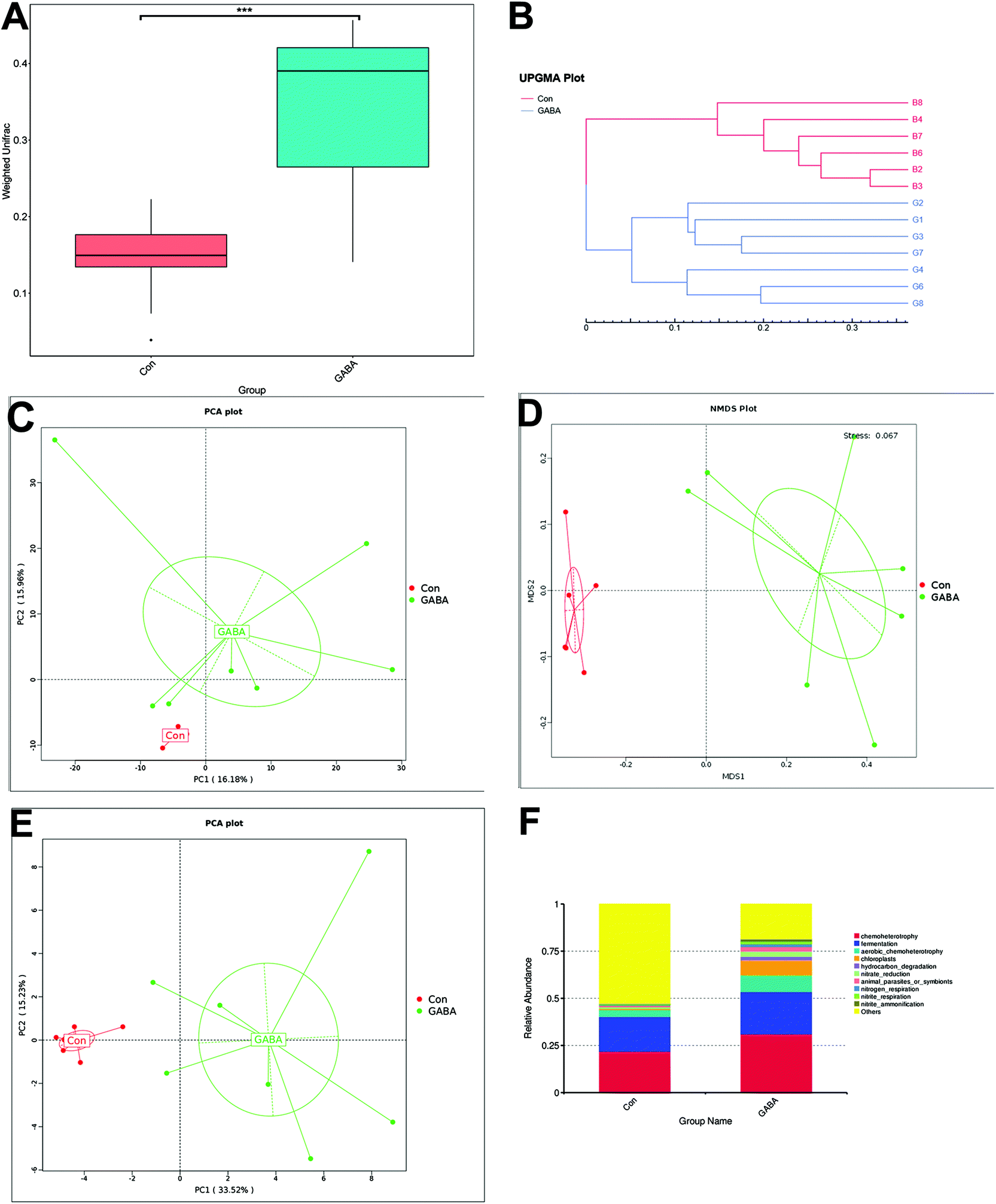 | ||
| Fig. 3 Dietary GABA highly regulated the beta diversity and function of the ilea. Rank abundance curves in each sample are shown in (A). Beta diversity analysis, including UPMGA (B), PCA (C) and NMDS (D), contrasted the differences between GABA and Con groups. Function prediction indicated that dietary GABA altered the microbial function (E), such as increasing chemoheterotrophy, fermentation, chloroplasts and nitrate/nitrite/nitrogen respiration (F). Con, control group (N = 6); GABA, GABA group (N = 7). ***P < 0.01. | ||
 | ||
| Fig. 4 GABA-induced change in the intestinal microbiota correlated with the growth performance, intestinal immunity and profiles of amino acids. Environmental factor correlation analysis revealed that the Firmicutes, the dominative population at the phylum level, were positively associated with the feed conversion ratio (A), intestinal expression of pIgR and Muc1 (B), and profiles of serum amino acid (including histidine, serine, arginine, alanine, proline, lysine, leucine and phenylalanine) (C and D), and were negatively associated with the growth rate (A), intestinal gene expression of IFNγ, Tlr6 and MyD88 (B), and profiles of ileal mucosal amino acid (including cystine, histidine, proline, serine, threonine, tryptophan and tyrosine) (C and D). Other populations, such as Proteobacteria and Bacteroidetes, in the phylum show an adverse relationship with the mentioned parameters. Con, control group (N = 6); GABA, GABA group (N = 7). *P < 0.05;**P < 0.01. | ||
Discussion
Piglets suffer from big challenges in the growth, gut health and immunity during weaning, due to inadequate feed intakes and lack of necessary hormones and immuno-enhancing substances in milk. Amino acids are used to prevent weaning stress and this practice has been strongly supported by the recent studies.34 We report that the dietary GABA supplementation improves the growth performance, and highly regulates the intestinal immunity, the amino acid profiles in the intestinal mucosa, and the intestinal microbiota in weaned piglets.Inadequate feed intake and growth restriction following weaning impaired the feed intake and growth of the weaned piglets.35,36 GABA is reported to promote growth performance in poultry and livestock, such as chickens, pigeons, sheep and cows.26,37,38 Wang et al. reported that dietary GABA enhances the feed intake and promotes the gene expressions of GABAB receptors, while inhibiting the expression of CCK-related gene in growing lambs.22 Similarly, we found that the dietary GABA supplementation promotes the growth rate, the feed efficiency and the kidney organ index of piglets. The mechanism may lie in the role of GABA in promoting secretions of growth hormones, insulin, neuropeptide Y (NPY), and cholecystokinin,39 which are closely related to growth performance. Our findings are different from the report by Li, who reported that the 30 mg kg−1 dietary supplementation of GABA has little effect on the average daily feed intakes, average daily gains and feed conversion ratios in the weaned pigs.25 The difference may come from the different weaning age of piglets or a different dosage of GABA supplementation.
Besides the growth performance, the intestinal immune system also suffers from the challenge of weaning. Intestinal mucosa is considered as the first defensive line to prevent entry of pathogens in the intestinal tract40,41 and it can express multiple immune factors, including inflammatory cytokines produced by the intestinal immune system, alpha defensins produced by the Paneth cells, secretory immunoglobin A from the plasma cells, and the goblet cell-producing mucin protein.42,43 A series of novel studies indicated that GABA has a neuro-immune interaction that is potentially involved in the regulation of immune cell activity.44,45 We found that dietary GABA plays a modulatory role in the intestinal immunity by regulating the intestinal immune system, such as inhibiting the gene expressions of IL-22, proinflammatory cytokines (IL-1 and IL-18) and Muc1, and promoting the gene expressions of anti-inflammatory cytokines (IFN-γ, IL-4 and IL-10), TLR6 and MyD88. Some previous studies also claimed that GABA inhibits the gene expression of pro-inflammatory cytokines, such as IL-1.46–48 The results indicate that GABA could regulate the gut immune responses in weaned piglets.
Increasing number of reports illustrated the roles of GABAergic system on the intestinal health and diseases. Thus, we analyzed the effect of GABA on the intestinal GABAergic system as well, which may further regulate the motility, secretion and immunity of the intestinal tract in ENS dependent or independent ways.16,49 GADs are expressed not only in the nervous system, but also exist in the thymus, ovaries, pancreas, stomach,50–52 and the intestines of humans, cats, rats and fetal pigs.53,54 However, they are undetectable in the small intestine of piglets, possibly due to the low expression of GAD that could not be detected in our model by RT-PCR. GATs show high affinity to the plasma membrane and are found in the intestines, including the duodenum, ileum, and colon.14 In this study, all four GATs are detected in the ileum of piglets, and GABA supplementation affects the SLC6A13 expression in the ileum. Li et al. reported that the mRNAs of 19 subunits of GABRA and GAD are detected in the rat intestinal epithelial cell line-18, and some subunits of GABAA and GAD also exist in the small intestines of mice, rats and humans.54 This study also demonstrated that both GABAA and GABAB are expressed in small intestines of piglets, and dietary GABA modulates some components of the intestinal GABA receptors. However, it is limited in clarifying the relation of GABA receptors with the intestinal immune responses.
Amino acid metabolism is a complex network and often changes coordinately.55–57 Amino acids are absorbed through the intestinal luminal plasma membrane into the intestinal mucosa and then transported to the blood.58 Interestingly, we found that the GABA administration increases the absorption of the intestinal mucosal natural amino acids, while lowers the levels of most natural amino acids in the sera of piglets. Similarly, Xie et al. reported that the GABA administration ameliorates the high-fat diet-induced plasma free amino acid disorder by arresting the arginine reduction and promoting the metabolism of other natural amino acids.59 The contrasting effects of GABA on the profiles of amino acids in the intestinal mucosa and the sera reveal that GABA may play a role in promoting both the absorption and the consumption of amino acids, and this might be associated with the protein synthesis or energy metabolism of amino acids. It is widely known that amino acids such as Gln and Glu are regulators of the growth performance, gut microbiota and intestinal immunity, indicating that the promotion of growth performance and the regulation of intestinal immunity by GABA might depend on its effects on the amino acid profiles. However, the detailed mechanism of alterations in the amino acid profiles driven by GABA needs to be further studied.
Gut microbiota play important roles in the growth, immune regulation, anti-infection, metabolism and intestinal health of the host.60–63 We found that GABA markedly modulates the compositions and diversity, and changes the functions of gut microbiota, suggesting that GABA plays an important role in maintaining the intestinal microbial homeostasis. Previous studies showed that GABA-producing bacteria (i.e., Lactobacillus, Lactococcus, Enterococcus, Leuconostoc, Pediococcus, Rhizopus and Streptococcus) could affect the host in multiple ways by producing GABA.64 For example, gut microbial-producing GABA enhanced intestinal IL-17 secretion in ETEC-challenged mice.42 Also, GABA notably suppressed the percentage of Clostridium sensustricto (cluster I), a group of bacteria that could use glucose to produce short-chain fatty acids.65 Meanwhile, some other probiotic and lactic acid producing microbes, such as Lactobacillus,66 were increased in this study after GABA supplementation. We also found that the shifted microbiota correlates with the growth performance, intestinal immunity and amino acid profiles of the weaned piglets. These novel results indicate that GABA serves as an intestinal microbiota regulator, which may further mediate the host growth, metabolism and immune system.
In conclusion, dietary GABA plays a beneficial role in relieving the weaning stress, such as improving the growth performance, kidney organ index and regulating the intestinal immunity of weaned piglets, which may be through the regulation of amino acid profiles and intestinal microbiota. The results would be helpful for understanding the function of GABA in the intestine and benefit for the application of GABA in animal production.
Abbreviations
| GABA | Gamma-aminobutyric acid |
| Glu | Glutamate |
| Gln | Glutamine |
| CNS | Central nervous system |
| ENS | Enteric nervous system |
| GAD | Glutamic acid decarboxylase |
| GAT | GABA transports |
| HE | Hematoxylin and eosin |
| V/C | Villus length to crypt depth |
| HPLC | High-performance liquid chromatography |
| RT-PCR | Real-time quantitative reverse transcriptase-polymerase chain reaction |
| TLR | Toll-like receptors |
| MyD88 | Myeloid differential protein-88 |
| pIgR | Polymeric immunoglobulin receptor |
| Muc | Mucin |
| TNF-α | Tumor necrosis factor alpha |
| IFN-γ | Interferon gamma |
| IL | Interleukin SEM, standard error of the mean |
| BW | Body weight |
| ADFI | Average daily feed intake. |
Author contributions
S. C., G. L., R. W. and B. T. designed the experiment. S. C., S. L., M. W., Y. X. and M. Q. conducted the experiment. J. Y., P. B. and G. L. analyzed the data and prepared tables and figures. S. C. and B. T. prepared the manuscript. R. W. and Y. Y. revised the manuscript.Conflicts of interest
There are no conflicts to declare.Acknowledgements
This research was jointly supported by the National Key R&D Program (2017YFD0500503), Key Programs of frontier scientific research of the Chinese Academy of Sciences (QYZDYSSW-SMC008) and National Natural Science Foundation of China (31872365, 31790411 and 31672433).References
- B. Jayaraman and C. M. Nyachoti, Husbandry practices and gut health outcomes in weaned piglets: A review, Anim. Nutr., 2017, 3, 205–211 CrossRef PubMed.
- M. Castillo, S. M. Martin-Orue, M. Nofrarias, E. G. Manzanilla and J. Gasa, Changes in caecal microbiota and mucosal morphology of weaned pigs, Vet. Microbiol., 2007, 124, 239–247 CrossRef CAS PubMed.
- G. Wu, Functional amino acids in nutrition and health, Amino Acids, 2013, 45, 407–411 CrossRef CAS PubMed.
- J. Yin, W. Ren, X. Huang, T. Li and Y. Yin, Protein restriction and cancer, Biochim. Biophys. Acta, 2018, 1869, 256–262 CAS.
- S. Chen, S. Liu, F. Zhang, W. Ren, N. Li, J. Yin, J. Duan, Y. Peng, G. Liu, Y. Yin and G. Wu, Effects of dietary L-glutamine supplementation on specific and general defense responses in mice immunized with inactivated Pasteurella multocida vaccine, Amino Acids, 2014, 46, 2365–2375 CrossRef CAS PubMed.
- R. Rezaei, D. A. Knabe, C. D. Tekwe, S. Dahanayaka, M. D. Ficken, S. E. Fielder, S. J. Eide, S. L. Lovering and G. Wu, Dietary supplementation with monosodium glutamate is safe and improves growth performance in postweaning pigs, Amino Acids, 2013, 44, 911–923 CrossRef CAS PubMed.
- X. Wu, Y. Zhang, Z. Liu, T. J. Li and Y. L. Yin, Effects of oral supplementation with glutamate or combination of glutamate and N-carbamylglutamate on intestinal mucosa morphology and epithelium cell proliferation in weanling piglets, J. Anim. Sci., 2012, 90(Suppl 4), 337–339 CrossRef PubMed.
- C. J. Hu, Q. Y. Jiang, T. Zhang, Y. L. Yin, F. N. Li, J. P. Deng, G. Y. Wu and X. F. Kong, Dietary supplementation with arginine and glutamic acid modifies growth performance, carcass traits, and meat quality in growing-finishing pigs, J. Anim. Sci., 2017, 95, 2680–2689 CAS.
- G. Liu, W. Ren, J. Fang, C. A. Hu, G. Guan, N. A. Al-Dhabi, J. Yin, V. Duraipandiyan, S. Chen, Y. Peng and Y. Yin, L-Glutamine and L-arginine protect against enterotoxigenic Escherichia coli infection via intestinal innate immunity in mice, Amino Acids, 2017, 49, 1945–1954 CrossRef CAS PubMed.
- J. Duan, J. Yin, M. Wu, P. Liao, D. Deng, G. Liu, Q. Wen, Y. Wang, W. Qiu, Y. Liu, X. Wu, W. Ren, B. Tan, M. Chen, H. Xiao, L. Wu, T. Li, C. M. Nyachoti, O. Adeola and Y. Yin, Dietary glutamate supplementation ameliorates mycotoxin-induced abnormalities in the intestinal structure and expression of amino acid transporters in young pigs, PLoS One, 2014, 9, e112357 CrossRef PubMed.
- J. Duan, J. Yin, W. Ren, T. Liu, Z. Cui, X. Huang, L. Wu, S. W. Kim, G. Liu, X. Wu, G. Wu, T. Li and Y. Yin, Dietary supplementation with L-glutamate and L-aspartate alleviates oxidative stress in weaned piglets challenged with hydrogen peroxide, Amino Acids, 2016, 48, 53–64 CrossRef CAS PubMed.
- G. Wu, Amino acids: metabolism, functions, and nutrition, Amino Acids, 2009, 37, 1–17 CrossRef CAS PubMed.
- J. B. Furness, Integrated Neural and Endocrine Control of Gastrointestinal Function, Adv. Exp. Med. Biol., 2016, 891, 159–173 CrossRef CAS PubMed.
- E. L. Fletcher, M. J. Clark and J. B. Furness, Neuronal and glial localization of GABA transporter immunoreactivity in the myenteric plexus, Cell Tissue Res., 2002, 308, 339–346 CrossRef CAS PubMed.
- M. Lyte, Probiotics function mechanistically as delivery vehicles for neuroactive compounds: Microbial endocrinology in the design and use of probiotics, BioEssays, 2011, 33, 574–581 CrossRef CAS PubMed.
- M. Auteri, M. G. Zizzo and R. Serio, GABA and GABA receptors in the gastrointestinal tract: from motility to inflammation, Pharmacol. Res., 2015, 93, 11–21 CrossRef CAS PubMed.
- M. F. Tsai, P. McCarthy and C. Miller, Substrate selectivity in glutamate-dependent acid resistance in enteric bacteria, Proc. Natl. Acad. Sci. U. S. A., 2013, 110, 5898–5902 CrossRef CAS PubMed.
- R. Mazzoli and E. Pessione, The Neuro-endocrinological Role of Microbial Glutamate and GABA Signaling, Front. Microbiol., 2016, 7, 1934 Search PubMed.
- D. Zhang, Z. H. Pan, M. Awobuluyi and S. A. Lipton, Structure and function of GABA(C) receptors: a comparison of native versus recombinant receptors, Trends Pharmacol. Sci., 2001, 22, 121–132 CrossRef CAS PubMed.
- M. Watanabe, K. Maemura, K. Kanbara, T. Tamayama and H. Hayasaki, GABA and GABA receptors in the central nervous system and other organs, Int. Rev. Cytol., 2002, 213, 1–47 CrossRef CAS PubMed.
- A. Billinton, A. O. Ige, J. P. Bolam, J. H. White, F. H. Marshall and P. C. Emson, Advances in the molecular understanding of GABAB receptors, Trends Neurosci., 2001, 24, 277–282 CrossRef CAS PubMed.
- D. M. Wang, B. Chacher, H. Y. Liu, J. K. Wang, J. Lin and J. X. Liu, Effects of gamma-aminobutyric acid on feed intake, growth performance and expression of related genes in growing lambs, Animal, 2015, 9, 445–448 CrossRef CAS PubMed.
- D. M. Wang, H. Y. Liu, C. Wang, J. X. Liu and J. D. Ferguson, Effects of rumen-protected gamma-aminobutyric acid on feed intake, performance and antioxidative status in transition cows, Livest. Sci., 2013, 153, 66–72 CrossRef.
- W. Ren, J. Yin, H. Xiao, S. Chen, G. Liu, B. Tan, N. Li, Y. Peng, T. Li, B. Zeng, W. Li, H. Wei, Z. Yin, G. Wu, P. R. Hardwidge and Y. Yin, Intestinal Microbiota-Derived GABA Mediates Interleukin-17 Expression during Enterotoxigenic Escherichia coli Infection, Front. Immunol., 2016, 7, 685 Search PubMed.
- Y. H. Li, F. Li, M. Liu, J. J. Yin, B. J. Cheng, B. M. Shi and A. S. Shan, Effect of gamma-aminobutyric acid on growth performance, behavior and plasma hormones in weaned pigs, Can. J. Anim. Sci., 2015, 95, 165–171 CrossRef CAS.
- D. M. Wang, C. Wang, H. Y. Liu, J. X. Liu and J. D. Ferguson, Effects of rumen-protected gamma-aminobutyric acid on feed intake, lactation performance, and antioxidative status in early lactating dairy cows, J. Dairy Sci., 2013, 96, 3222–3227 CrossRef CAS PubMed.
- W. Ren, J. Yin, M. Wu, G. Liu, G. Yang, Y. Xion, D. Su, L. Wu, T. Li, S. Chen, J. Duan, Y. Yin and G. Wu, Serum amino acids profile and the beneficial effects of L-arginine or L-glutamine supplementation in dextran sulfate sodium colitis, PLoS One, 2014, 9, e88335 CrossRef PubMed.
- S. Chen, P. Bin, W. Ren, W. Gao, G. Liu, J. Yin, J. Duan, Y. Li, K. Yao, R. Huang, B. Tan and Y. Yin, Alpha-ketoglutarate (AKG) lowers body weight and affects intestinal innate immunity through influencing intestinal microbiota, Oncotarget, 2017, 8, 38184–38192 Search PubMed.
- M. M. Wu, H. Xiao, G. Liu, S. Chen, B. Tan, W. K. Ren, F. W. Bazer, G. Y. Wu and Y. L. Yin, Glutamine promotes intestinal SIgA secretion through intestinal microbiota and IL-13, Mol. Nutr. Food Res., 2016, 60, 1637–1648 CrossRef CAS PubMed.
- J. Yin, Y. Li, H. Han, J. Zheng, L. Wang, W. Ren, S. Chen, F. Wu, R. Fang, X. Huang, C. Li, B. Tan, X. Xiong, Y. Zhang, G. Liu, J. Yao, T. Li and Y. Yin, Effects of Lysine deficiency and Lys-Lys dipeptide on cellular apoptosis and amino acids metabolism, Mol. Nutr. Food Res., 2017, 61 DOI:10.1002/mnfr.201600754.
- J. Yin, W. Ren, G. Liu, J. Duan, G. Yang, L. Wu, T. Li and Y. Yin, Birth oxidative stress and the development of an antioxidant system in newborn piglets, Free Radical Res., 2013, 47, 1027–1035 CrossRef CAS PubMed.
- J. Yin, W. Ren, J. Duan, L. Wu, S. Chen, T. Li, Y. Yin and G. Wu, Dietary arginine supplementation enhances intestinal expression of SLC7A7 and SLC7A1 and ameliorates growth depression in mycotoxin-challenged pigs, Amino Acids, 2014, 46, 883–892 CrossRef CAS PubMed.
- J. Yin, M. M. Wu, H. Xiao, W. K. Ren, J. L. Duan, G. Yang, T. J. Li and Y. L. Yin, Development of an antioxidant system after early weaning in piglets, J. Anim. Sci., 2014, 92, 612–619 CrossRef CAS PubMed.
- W. W. Wang, S. Y. Qiao and D. F. Li, Amino acids and gut function, Amino Acids, 2009, 37, 105–110 CrossRef CAS PubMed.
- H. S. Yang, X. Xiong, X. C. Wang, T. J. Li and Y. L. Yin, Effects of weaning on intestinal crypt epithelial cells in piglets, Sci. Rep., 2016, 6 DOI:10.1038/srep36939.
- F. Smith, J. E. Clark, B. L. Overman, C. C. Tozel, J. H. Huang, J. E. F. Rivier, A. T. Blisklager and A. J. Moeser, Early weaning stress impairs development of mucosal barrier function in the porcine intestine, Am. J. Physiol.: Gastrointest. Liver Physiol., 2010, 298, G352–G363 CrossRef CAS PubMed.
- W. Y. Xie, X. Y. Hou, F. B. Yan, G. R. Sun, R. L. Han and X. T. Kang, Effect of -aminobutyric acid on growth performance and immune function in chicks under beak trimming stress, Anim. Sci. J., 2013, 84, 121–129 CrossRef CAS PubMed.
- N. C. Jo, J. Jung, J. N. Kim, J. Lee, S. Y. Jeong, W. Kim, H. G. Sung and S. Seo, Effect of vaccination against foot-and-mouth disease on growth performance of Korean native goat (Capra hircus coreanae), J. Anim. Sci., 2014, 92, 2578–2586 CrossRef CAS PubMed.
- E. R. Kim, Z. F. Wu, H. Sun, Y. Z. Xu, L. R. Mangieri, Y. Xu and Q. C. Tong, Hypothalamic Non-AgRP, Non-POMC GABAergic Neurons Are Required for Postweaning Feeding and NPY Hyperphagia, J. Neurosci., 2015, 35, 10440–10450 CrossRef CAS PubMed.
- T. H. Yen and N. A. Wright, The gastrointestinal tract stem cell niche, Stem Cell Rev., 2006, 2, 203–212 CrossRef CAS PubMed.
- A. J. Battersby and D. L. Gibbons, The gut mucosal immune system in the neonatal period, Pediatr. Allergy Immunol., 2013, 24, 414–421 CrossRef PubMed.
- W. Ren, K. Wang, J. Yin, S. Chen, G. Liu, B. Tan, G. Wu, F. W. Bazer, Y. Peng and Y. Yin, Glutamine-Induced Secretion of Intestinal Secretory Immunoglobulin A: A Mechanistic Perspective, Front. Immunol., 2016, 7, 503 Search PubMed.
- W. K. Ren, S. Chen, L. W. Zhang, G. Liu, T. Hussain, X. Hao, J. Yin, J. L. Duan, B. Tan, G. Y. Wu, F. W. Bazer and Y. L. Yin, Interferon Tau Affects Mouse Intestinal Microbiota and Expression of IL-17, Mediators Inflammation, 2016, 2839232, DOI:10.1155/2016/2839232.
- B. D. Gulbransen and K. A. Sharkey, Novel functional roles for enteric glia in the gastrointestinal tract, Nat. Rev. Gastroenterol. Hepatol., 2012, 9, 625–632 CrossRef CAS PubMed.
- J. R. Grider, Neurotransmitters mediating the intestinal peristaltic reflex in the mouse, J. Pharmacol. Exp. Ther., 2003, 307, 460–467 CrossRef CAS PubMed.
- M. G. Reyes-Garcia, F. Hernandez-Hernandez, B. Hernandez-Tellez and F. Garcia-Tamayo, GABA (A) receptor subunits RNA expression in mice peritoneal macrophages modulate their IL-6/IL-12 production, J. Neuroimmunol., 2007, 188, 64–68 CrossRef CAS PubMed.
- R. Bhat, R. Axtell, A. Mitra, M. Miranda, C. Lock, R. W. Tsien and L. Steinman, Inhibitory role for GABA in autoimmune inflammation, Proc. Natl. Acad. Sci. U. S. A., 2010, 107, 2580–2585 CrossRef CAS PubMed.
- N. Soltani, H. Qiu, M. Aleksic, Y. Glinka, F. Zhao, R. Liu, Y. Li, N. Zhang, R. Chakrabarti, T. Ng, T. Jin, H. Zhang, W. Y. Lu, Z. P. Feng, G. J. Prud'homme and Q. Wang, GABA exerts protective and regenerative effects on islet beta cells and reverses diabetes, Proc. Natl. Acad. Sci. U. S. A., 2011, 108, 11692–11697 CrossRef CAS PubMed.
- J. Vlainic, J. Suran, T. Vlainic and A. L. Vukorep, Probiotics as an Adjuvant Therapy in Major Depressive Disorder, Curr. Neuropharmacol., 2016, 14, 952–958 CrossRef CAS PubMed.
- S. L. Erdo and J. R. Wolff, gamma-Aminobutyric acid outside the mammalian brain, J. Neurochem., 1990, 54, 363–372 CrossRef CAS.
- B. E. Faulkner-Jones, D. S. Cram, J. Kun and L. C. Harrison, Localization and quantitation of expression of two glutamate decarboxylase genes in pancreatic beta-cells and other peripheral tissues of mouse and rat, Endocrinology, 1993, 133, 2962–2972 CrossRef CAS PubMed.
- J. S. Petersen, S. Russel, M. O. Marshall, H. Kofod, K. Buschard, N. Cambon, A. E. Karlsen, E. Boel, W. A. Hagopian and K. R. Hejnaes, et al., Differential expression of glutamic acid decarboxylase in rat and human islets, Diabetes, 1993, 42, 484–495 CrossRef CAS PubMed.
- Y. Miki, K. Taniyama, C. Tanaka and T. Tobe, GABA, glutamic acid decarboxylase, and GABA transaminase levels in the myenteric plexus in the intestine of humans and other mammals, J. Neurochem., 1983, 40, 861–865 CrossRef CAS.
- Y. Li, Y. Y. Xiang, W. Y. Lu, C. Liu and J. Li, A novel role of intestine epithelial GABAergic signaling in regulating intestinal fluid secretion, Am. J. Physiol.: Gastrointest. Liver Physiol., 2012, 303, G453–G460 CrossRef CAS PubMed.
- Y. Noguchi, Q. W. Zhang, T. Sugimoto, Y. Furuhata, Y. Sakai, M. Mori, M. Takahashi and T. Kimura, Network analysis of plasma and tissue amino acids and the generation of an amino index for potential diagnostic use, Am. J. Clin. Nutr., 2006, 83, 513s–519s CrossRef CAS.
- N. Shikata, Y. Maki, Y. Noguchi, M. Mori, T. Hanai, M. Takahashi and M. Okamoto, Multi-layered network structure of amino acid (AA) metabolism characterized by each essential AA-deficient condition, Amino Acids, 2007, 33, 113–121 CrossRef CAS PubMed.
- G. Hernandez-Montes, J. J. Diaz-Mejia, E. Perez-Rueda and L. Segovia, The hidden universal distribution of amino acid biosynthetic networks: a genomic perspective on their origins and evolution, Genome Biol., 2008, 9, R95 CrossRef PubMed.
- S. Broer, Amino acid transport across mammalian intestinal and renal epithelia, Physiol. Rev., 2008, 88, 249–286 CrossRef CAS PubMed.
- Z. X. Xie, S. F. Xia, Y. Qiao, Y. H. Shi and G. W. Le, Effect of GABA on oxidative stress in the skeletal muscles and plasma free amino acids in mice fed high-fat diet, J. Anim. Physiol. Anim. Nutr., 2015, 99, 492–500 CrossRef CAS PubMed.
- M. G. Rooks and W. S. Garrett, Gut microbiota, metabolites and host immunity, Nat. Rev. Immunol., 2016, 16, 341–352 CrossRef CAS PubMed.
- M. Nieuwdorp, P. W. Gilijamse, N. Pai and L. M. Kaplan, Role of the microbiome in energy regulation and metabolism, Gastroenterology, 2014, 146, 1525–1533 CrossRef CAS PubMed.
- M. Arumugam, J. Raes, E. Pelletier, D. Le Paslier, T. Yamada, D. R. Mende, G. R. Fernandes, J. Tap, T. Bruls, J. M. Batto, M. Bertalan, N. Borruel, F. Casellas, L. Fernandez, L. Gautier, T. Hansen, M. Hattori, T. Hayashi, M. Kleerebezem, K. Kurokawa, M. Leclerc, F. Levenez, C. Manichanh, H. B. Nielsen, T. Nielsen, N. Pons, J. Poulain, J. Qin, T. Sicheritz-Ponten, S. Tims, D. Torrents, E. Ugarte, E. G. Zoetendal, J. Wang, F. Guarner, O. Pedersen, W. M. de Vos, S. Brunak, J. Dore, H. I. T. C. Meta, M. Antolin, F. Artiguenave, H. M. Blottiere, M. Almeida, C. Brechot, C. Cara, C. Chervaux, A. Cultrone, C. Delorme, G. Denariaz, R. Dervyn, K. U. Foerstner, C. Friss, M. van de Guchte, E. Guedon, F. Haimet, W. Huber, J. van Hylckama-Vlieg, A. Jamet, C. Juste, G. Kaci, J. Knol, O. Lakhdari, S. Layec, K. Le Roux, E. Maguin, A. Merieux, R. Melo Minardi, C. M'Rini, J. Muller, R. Oozeer, J. Parkhill, P. Renault, M. Rescigno, N. Sanchez, S. Sunagawa, A. Torrejon, K. Turner, G. Vandemeulebrouck, E. Varela, Y. Winogradsky, G. Zeller, J. Weissenbach, S. D. Ehrlich and P. Bork, Enterotypes of the human gut microbiome, Nature, 2011, 473, 174–180 CrossRef CAS PubMed.
- M. Schwarzer, M. Strigini and F. Leulier, Gut Microbiota and Host Juvenile Growth, Calcif. Tissue Int., 2018, 102, 387–405 CrossRef CAS PubMed.
- R. Dhakal, V. K. Bajpai and K. H. Baek, Production of gaba (gamma - Aminobutyric acid) by microorganisms: a review, Braz. J. Microbiol., 2012, 43, 1230–1241 CrossRef CAS PubMed.
- A. Willems and M. D. Collins, Phylogenetic relationships of the genera Acetobacterium and Eubacterium sensu stricto and reclassification of Eubacterium alactolyticum as Pseudoramibacter alactolyticus gen. nov., comb. nov, Int. J. Syst. Bacteriol., 1996, 46, 1083–1087 CrossRef CAS PubMed.
- P. Hemarajata and J. Versalovic, Effects of probiotics on gut microbiota: mechanisms of intestinal immunomodulation and neuromodulation, Ther. Adv. Gastroenterol., 2013, 6, 39–51 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c8fo02161a |
| This journal is © The Royal Society of Chemistry 2019 |