DOI:
10.1039/C6FO01329E
(Paper)
Food Funct., 2017,
8, 429-436
Protective effects of dietary glycine and glutamic acid toward the toxic effects of oxidized mustard oil in rabbits
Received
8th September 2016
, Accepted 5th January 2017
First published on 6th January 2017
Abstract
The protective role of glycine and glutamic acid against the toxic effects of oxidized oil was studied for the first time. Mustard seed oil was thermally oxidized and characterized for quality characteristics and polyphenolic composition using reversed phase HPLC-DAD. Significant changes in the quality characteristics occurred with thermal oxidation. Fourteen polyphenolic compounds were identified and quantified in oils. Quercetin-3-glucoside, quercetin-3-feruloylsophoroside, catechin, quercetin-3-rutinoside, quercetin-3,7-diglucoside, sinapic acid and vanillic acid hexoside were the major compounds in the fresh and oxidized oil. Oxidized, un-oxidized mustard oils, glycine and glutamic acid were given to rabbits alone or in combination. The biochemical responses were studied in terms of haematological and biochemical parameters and histopathology. It has been observed that biochemical and haematological parameters were adversely affected by the oxidized oil, while supplementation of both amino acids was beneficial in normalizing these parameters. Both amino acids alone have no significant effects, however, oxidized oil affected the liver by enhancing fat accumulation, causing hepatitis, reactive Kupffer cells and necrosis. The co-administration of oxidized oils with glycine or glutamic acid revealed significant recovery of the liver structure and function. In conclusion, glycine or glutamic acid is beneficial and protective against food toxicity and can be considered as an ameliorative food supplement.
Introduction
Glycine is the simplest and non-essential amino acid, which is important for the synthesis of glucose, purine, chlorophylls, conjugation of bile acids and heme.1 It is the key amino acid to reduce fibrosis and eliminate the liver injury caused by experimental drugs and blocks programmed cell death.2 Moreover, metabolites of glycine also play an important role in cancer.3 Glycine has a protective role against anoxic and oxidative stress produced by ethanol. The derivatives of glycine decrease the activation of lipid peroxidation in stress, reduce the duration of the alarm stage of stress reaction and limit stress damage to the heart.4 Glycine intake was effective against the decrease in the plasma free fatty acids, adipose cell size and blood pressure in sucrose fed rat models.5 The role of glycine against oxidative stress in animal models makes it beneficial, protective6 and helpful in restoring the normal glutathione levels. Glycine has protective effects against lipid peroxidation,7 however, oxidative stress produced by dietary oxidized lipids in animal models may have similar protective effects.
Glutamic acid is the most predominant amino acid naturally present in many foods either in the free or bound state of the animal or vegetable origin such as mushroom, tomato, fish and cheese.8 Glutamate maintains the balance of nitrogen in the biological system. Proteins rich in proline, glutamic acid and leucine have been found to be essential for the optimal response of the p53 gene.9 It is useful in the treatment of different cancers and its metallo-drugs have strong anticancer properties.10 It acts as a neurotransmitter and very helpful in the treatment of hypoglycaemic coma, ulcers, neurological conditions, mental retardation, Parkinson's disease, muscular dystrophy and epilepsy.11 The recent work of Yin et al.12 revealed that glutamate and aspartate are beneficial against oxidative stress in piglets. There is however no information regarding the role of dietary supplementation of glycine & glutamate against the toxic effects produced by oxidized lipids.
Lipids are oxidized to primary and secondary oxidation products13 during frying of foods. Ingestion of dietary oxidized lipids has been found to cause hyperlipidaemia,14 increase oxidative stress, toxicity, fat deposition in the liver, necrosis15,16 and ultimately cancer. The present work was carried out to determine the protective role of glycine and glutamic acid against the toxic effects produced by thermally oxidized mustard oil in rabbits.
Materials and methods
Materials
Fresh mustard (Brassica Juncea L.) seed oil was obtained using the pressing technique at 35 °C from the local extraction plant. Glycine (PubChem CID: 750), catechin, sinapic acid, quercetin-3-glucoside, quercetin-3-rutinoside, malondialdehyde (MDA), and thiobarbituric acid (TBA) were purchased from Sigma-Aldrich (USA) and glutamic acid (PubChem CID: 611) was from Scharlau (Spain). All other chemicals and reagents were of high purity and ACS analytical grade.
Analyses of oil
Mustard oil was thermally oxidized on a hot plate at 170 ± 10 °C in air, for consecutive 9 hours and was stored in a refrigerator at −20 °C to avoid further oxidation process. The oil samples were characterized for the following parameters:
Total phenolic & flavonoid contents.
Total phenolic contents (TPC) were extracted with methanol![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :water (9:1, v/v) and analysed using Folin Ciocalteu's reagent against the standard calibration curve of gallic acid. TPC was expressed as milligrams of gallic acid equivalents per 100 g of mustard oil. Total flavonoids were measured according to the method of Mohdaly et al.17 Briefly, an aliquot of 0.5 mL ethanolic AlCl3 solution was added to the extract solution (0.5 mL). After 1 h at room temperature the absorbance at 420 nm was measured. Total flavonoid contents were expressed as quercetin equivalents (QEs) per gram of oil.
:water (9:1, v/v) and analysed using Folin Ciocalteu's reagent against the standard calibration curve of gallic acid. TPC was expressed as milligrams of gallic acid equivalents per 100 g of mustard oil. Total flavonoids were measured according to the method of Mohdaly et al.17 Briefly, an aliquot of 0.5 mL ethanolic AlCl3 solution was added to the extract solution (0.5 mL). After 1 h at room temperature the absorbance at 420 nm was measured. Total flavonoid contents were expressed as quercetin equivalents (QEs) per gram of oil.
Lipid oxidation.
Mustard oil samples were evaluated for peroxide contents using the standard AOCS method18 and were expressed as meq of oxygen per kg of oil. Secondary lipid oxidation was studied using the thiobarbituric acid reactive substances (TBARS) method as reported recently.19
Pigment contents.
Pigments such as chlorophyll a, b and β-carotene were extracted with acetone:hexane (4:6) and absorbance was measured at 663, 645, 505 and 453 nm using a UV-visible spectrophotometer (Shimadzu, Japan) against the blank. The pigments were expressed as mg per 100 mL.
Polyphenolic profile.
Polyphenolic compounds were repeatedly extracted using a methanol–water (60:40) mixture from the oil solution dissolved in hexane. The concentrated fractions were filtered in HPLC vials. The polyphenolic compounds were separated and identified using HPLC-DAD (Agilent 1260 Infinity Better, Agilent Technologies, Germany). The column used was Agilent Rapid Resolution Zorbax Eclipse Plus C18 (4.6 × 100 mm, 3.5 μm) (Agilent Technologies, Germany), maintained at 25 °C. The gradient system consists of methanol–acetic acid–deionized water (10:2:88) as solvent A and methanol–acetic acid–deionized water (90:2:8) as solvent B with a flow rate of 1 min.20 The chromatograms were obtained at 320 nm and absorption spectra in the range of 200–650 nm. The identification of phenolic compounds was based on the available reference standards (catechin, sinapic acid, quercetin-3-glucoside, quercetin-3-rutinoside) or identified by comparison with reported literature and quantified against the compounds of similar response factors of the reference standards.
Animal feeding
Rabbits (local Himalayan breed) were selected for the study, because of their highly selective and developed organ system and were easily available animals for these experiments.16 All experiments were approved by the ethical committee for proper care and experimental of animals of the Department of Biotechnology (complies with Helsinki protocols). The study was then approved by the Graduate Studies Committee of the Department and consequently by the advanced studies and research board (ASRB) of the University of Malakand (No. UOM/DDA/2013/481). Rabbits were fed on fresh diet daily and had free access to water and food during the study. A total of fifty-four rabbits (further divided into two groups, each containing 27 rabbits) were selected randomly for experimental purposes and were familiarized in the Bio-Park University of Malakand for a one week period. Group N was control, groups P, Z and R were given 1, 2 and 3 g kg−1 of glycine, group O was given 3 g kg−1 of oxidized mustard oil, group U was fed with 3 g kg−1 of un-oxidized mustard oil, while groups W, X and Y were fed with 3 g kg−1 of oxidized mustard oil with 1, 2 and 3 g kg−1 of glycine respectively. Similarly, C was a control group of glutamic acid, groups A, B and D were fed with 1, 2 and 3 g kg−1 of glutamic acid respectively, group E was fed with 3 g kg−1 of oxidized mustard oil, group F was fed with 3 g kg−1 of un-oxidized mustard oil, while groups G, H and M were fed with 3 g kg−1 of oxidized mustard oil together with 1, 2 and 3 g kg−1 of glutamic acid, respectively. The oral gavage feeding was continued for two weeks. The amounts of amino acids given were in accordance with previously published work.21
Serum biochemical analyses
After completing 2 weeks feeding session blood was taken from the jugular vein and kept in the serum and EDTA tubes. The blood samples were centrifuged at 5000g for a period of five minutes and the serum was decanted in labelled Eppendorf tubes and was preserved at −20 °C until analyses. Biochemical analysis was performed through a blood chemistry analyser (model Micro-Lab 200, MERCK, Japan). The analysed parameters were TG, cholesterol, HDL, LDL, SGPT and glucose.
Haematology
For haematology, 2 mL blood was taken in EDTA tubes from rabbits. The CELL-DYN 3200 Abbott Diagnostic Division, Canada (automatic digital machine) was used for the determination of haematological parameters.
Liver histopathology
The rabbit's liver was preserved in formalin before its analysis. Microscopic examination of slides was done with the help of electronic microscope model M 7000 D (SWIFT, Japan) and using a digital camera of microscope model DCM 130 (SWIFT, Japan) a picture was captured.
Statistical analyses
Analyses were carried out in triplicate or otherwise mentioned, and the values were presented in the tabular form with mean and standard deviation. The data were measured statistically using one way analysis of variance and the t test was applied at α = 0.05 using GraphPad Prism 6 (GraphPad Software, Inc., 2012).
Results and discussion
Characteristics of mustard oils
The oxidized and un-oxidized mustard oil was characterized for quality parameters like total phenolic contents, total flavonoid contents, pigment contents, peroxide values and TBARS contents as shown in Table 1. In the current investigation the total phenolic contents in un-oxidized and oxidized mustard oil were 229 and 569 mg of GAE per 100 g respectively. The value of TPC was higher than reported by Janu et al.22 who found that there are 56 mg of GAE per 100 g of TPC in mustard oil. The difference in total phenolic contents might be due to the difference in the extraction methods or the temperature of extraction, and the variety of mustard. The total flavonoid contents were 5.57 and 4.71 mg g−1 in un-oxidized and oxidized mustard oil. These results are in agreement with the previous findings which showed that flavonoid contents were declined during thermal treatment in rapeseed oil.23 Peroxide values of the un-oxidized and oxidized mustard oils were 4.52 and 115.5 meq kg−1, respectively. Similarly, an increase in the TBARS was also observed from 1.12 μmol g−1 of un-oxidized to 14.71 μmol g−1 of the oxidized mustard oil. The increase in the PV and TBARS may be due to the oxidation of the fatty acid moieties of triacylglycerols.13 Chlorophyll a decreased to 0.69 mg per 100 mL from 0.94 mg per 100 mL of the oxidized and un-oxidized mustard oils respectively, while there were no significant changes in the chlorophyll b contents. The content of β-carotene was 1.81 mg per 100 mL in un-oxidized mustard oil, while its amounts were not detected in oxidized oil samples. The high stability of the mustard seed oil may thus be due to these natural antioxidants.
Table 1 Composition and characteristics of un-oxidized and oxidized mustard oil
| Samples |
Un-oxidized mustard oil |
Oxidized mustard oil |
| Values are mean with standard deviations of replicate measurements (n = 5). The superscripts a and b represent extractions in methanol (100%) and methanol:water (1:1) respectively. TBARS stands for thiobarbituric acid reactive substances. |
| Total phenolic contents (mg per 100 g) |
229.0 ± 3.2 |
569.0 ± 4.5 |
| Total flavonoids (mg g−1) |
a5.57 ± 0.2 |
a4.71 ± 0.06 |
|
b7.11 ± 0.3 |
b9.19 ± 0.9 |
| Peroxide value (meq kg−1) |
4.52 ± 0.6 |
115.5 ± 2.3 |
| TBARS (μmol g−1) |
1.12 ± 0.04 |
14.71 ± 0.06 |
| Chlorophyll a (mg per 100 mL) |
0.95 ± 0.01 |
0.69 ± 0.01 |
| Chlorophyll b (mg per 100 mL) |
0.94 ± 0.008 |
0.96 ± 0.01 |
| β-Carotene (mg per 100 mL) |
1.81 ± 0.01 |
N.D |
Polyphenolic profile of mustard oils
Fig. 1 shows the separation and identification of polyphenolic compounds in oxidized and un-oxidized mustard oils. Fourteen phenolic compounds were identified and quantified with the details as shown in Table 2. Peak 1 was identified as gallic acid with a λmax of 271 nm against the reference standard. The amount of gallic acid increased significantly with thermal stress. This may be due to the degradation of other phenolic compounds to gallic acid. Jun et al.24 showed that gallic acid is present in the canola seeds suggesting that it can be used for finger printing of brassica seeds and oils. Peak 2 was identified as coumaroyl hexoside with a λmax of 316 and 232 nm with significant losses during oxidation. Sinapine was peak 3 with characteristic λmax of 328, 266, and 240 nm and a concentration of 3.15 & 1.63 μg g−1 in control and oxidized oil, respectively. Similarly, sinapic acid and sinapic acid glucoside were found to be compounds 3 and 4 respectively. Sinapic acid was 4.02 μg g−1 in the control oil sample and decreased upon heating to 1.89 μg g−1. Sinapic acid and its derivatives have been identified in the seeds and oils of the brassica family.25 These results indicate that sinapic acid or its derivatives can be used for finger printing seed oils from the brassica family. Peaks 6 & 7 were tri-galloylquinic acid and vanillic acid hexoside respectively. Peaks 8 to 11 were the derivatives of quercetin such as quercetin-3-feruloylsophoroside, quercetin-3-glucoside, quercetin-3,7-diglucoside, and quercetin-3-rutinoside. These compounds were present in a higher amount than other phenolic compounds representing a total of 36.62 μg g−1 in control samples. A significant decline occurred in the individual or collective amount (23.49 μg g−1) of quercetin derivatives with 36% losses. Peak 12 was kaempferol-3-(caffeoyldiglucoside)-7-rhamnoside with characteristic λmax of 338, 270 and 248 nm. Sinapoyl dihexoside was peak 13 with a λmax of 329 and 239 nm. This compound was identified in mustard seeds of the Canadian variety by Engels et al.25 Peak 14 was identified as cyanidin-3-glucoside with characteristic λmax of 278 and 234 nm. These results indicate that significant losses occurred in the all phenolic compounds with the exception of compounds 13 and 14.
 |
| | Fig. 1 Representative HPLC-DAD chromatograms of mustard oils at 320 nm. (A) Control, (B) oxidized mustard oil. Each peak in the chromatogram represents individual phenolic compound with characteristicdetails and amounts shown in Table 2. | |
Table 2 Identification & quantification of polyphenolic compounds using HPLC-DAD in oxidized & un-oxidized mustard oil
| Peak |
Rt |
Absorption spectra (nm) |
Identification |
Amount (μg g−1) |
| Control |
Oxidized |
| Values are expressed as mean with standard deviations of triplicate independent extraction & measurements. |
| 1 |
1.1 |
271 |
Gallic acid |
7.14 ± 0.1 |
10.5 ± 0.1 |
| 2 |
4.0 |
316, 232 |
Coumaroyl hexoside |
3.53 ± 0.1 |
1.83 ± 0.1 |
| 3 |
10.8 |
328, 266, 240 |
Sinapine |
3.15 ± 0.2 |
1.63 ± 0.2 |
| 4 |
12.6 |
322, 234 |
Sinapic acid |
4.02 ± 0.2 |
1.89 ± 0.2 |
| 5 |
13.3 |
328, 232 |
Sinapic acid glucoside |
1.85 ± 0.1 |
0.89 ± 0.01 |
| 6 |
15.1 |
272 |
Tri-galloylquinic acid |
1.36 ± 0.2 |
0.29 ± 0.01 |
| 7 |
22.2 |
292, 254 |
Vanillic acid hexoside |
6.03 ± 0.4 |
3.23 ± 0.4 |
| 8 |
25.8 |
334, 256 |
Quercetin-3-feruloylsophoroside |
7.98 ± 0.6 |
4.84 ± 0.6 |
| 9 |
28.2 |
355, 252 |
Quercetin-3-glucoside |
17.6 ± 0.3 |
11.7 ± 0.3 |
| 10 |
29.4 |
332, 264, 252 |
Quercetin-3,7-diglucoside |
5.01 ± 0.2 |
4.08 ± 0.2 |
| 11 |
29.8 |
355, 284 |
Quercetin-3-rutinoside |
2.73 ± 0.1 |
2.0 ± 0.1 |
| 12 |
30.6 |
338, 270, 248 |
Kaempferol-3-(caffeoyldiglucoside)-7-rhamnoside |
3.76 ± 0.2 |
1.96 ± 0.2 |
| 13 |
33.3 |
329, 239 |
Sinapoyl dihexoside |
0.89 ± 0.07 |
0.85 ± 0.07 |
| 14 |
33.7 |
278, 234 |
Cyanidin-3-glucoside |
1.72 ± 0.1 |
1.76 ± 0.1 |
Change in the body weight of rabbits
Glycine and glutamic acid increased the body weight by increasing the dose rate. The un-oxidized and oxidized mustard oil decline the body weight. But when the glycine was added to oxidized mustard oil it normalizes the body weight of rabbits as shown in Table 3. These results are in agreement with our previous findings,26 which showed that thermally oxidized sunflower oil decreased the body weight of rabbits compared to the control group. The decline in the weight may be due to disturbance in the lipid metabolism and protein oxidation in the body. The increase in the body weight with the supplementation of the selected amino acids is in accordance with the previous results,21 which showed that glycine increases the body weight as compared to the control group. These results indicate that oxidized lipids caused a decrease in the weight of the experimental animals, while the supplementation of the selected amino acids resulted in the normalization of the body weight.
Table 3 Effect of glycine and glutamic acid on the whole body weight of the rabbits
| Glycine groups |
Glutamic acid groups |
| Group |
Code |
Net gain/loss (g) |
Group |
Code |
Net gain/loss (g) |
| Values are expressed as mean ± SD of triplicates. Different letters (a–h) represent significant difference from N and C groups (control) at P < 0.05 using one way analysis of variance in column statistics. OM represents oxidized mustard oil. |
| Control |
N |
7.3 ± 1.2a |
Control |
C |
11.0 ± 1.5a |
| Glycine (1 g kg−1) |
P |
15.0 ± 1.0b |
Glutamic acid (1 g kg−1) |
A |
56.0 ± 1.7b |
| Glycine (2 g kg−1) |
Z |
21.0 ± 1.0c |
Glutamic acid (2 g kg−1) |
B |
79.0 ± 1.0c |
| Glycine (3 g kg−1) |
R |
30.0 ± 1.5d |
Glutamic acid (3 g kg−1) |
D |
100.0 ± 1.0d |
| Oxidized oil (3 g kg−1) |
O |
−21.0 ± 1.2e |
Oxidized oil (3 g kg−1) |
E |
−79.0 ± 1.0e |
| Un-oxidized oil (3 g kg−1) |
U |
−18.0 ± 1.5f |
Un-oxidized oil (3 g kg−1) |
F |
−71.0 ± 1.0f |
| OM (3 g kg−1) + glycine (1 g kg−1) |
W |
−5.7 ± 1.5g |
OM (3 g kg−1) + glutamic acid (1 g kg−1) |
G |
8.0 ± 1.0a |
| OM (3 g kg−1) + glycine (2 g kg−1) |
X |
8.7 ± 1.2a |
OM (3 g kg−1) + glutamic acid (2 g kg−1) |
H |
14.0 ± 1.0g |
| OM (3 g kg−1) + glycine (3 g kg−1) |
Y |
15.0 ± 1.0b |
OM (3 g kg−1) + glutamic acid (3 g kg−1) |
M |
21.0 ± 1.5h |
Organ weight of rabbits
The effect of oxidized, un-oxidized oils, glycine and glutamic acid alone or in combination on the weights of different organs (liver, kidney, heart and brain) was evaluated. It was found that glutamic acid and glycine increase the liver weight, while not affecting other body organs. Oxidized mustard oil significantly increased the weights of the liver and brain, while no significant changes were observed by the supplementation of un-oxidized oils. It was also found that when the two amino acids were administered along with oxidized oil, the weight of the rabbits reached the normal limit in the case of the liver and brain. The weight of the liver declined in the present study after ingestion of oxidized and un-oxidized mustard oil. In the present study there was no effect of amino acids and the oils on the kidney weight (Table 4). It was observed that amino acids and oils have no effects on the heart weight. The present results are in contrast to the study conducted by Saha et al.27 who showed that the weight of the heart declined by ingestion of mustard oil, it may be due to the change in the thermal temperature or the duration of the heating.
Table 4 Effect of dietary supplementation of glycine, glutamic acid alone or in combination with oxidized mustard oil on the weight of the liver, kidneys, heart and brain of rabbits
| Treatments |
Code |
Liver weight (g) |
Kidney weight (g) |
Heart weight (g) |
Brain weight (g) |
| Values are mean ± SD of triplicates. Different letters (a–f) represent significant difference from N (control of glycine group) or C (control of glutamic acid) at P < 0.05 using one way analysis of variance in each group. OM represents oxidized mustard oil. |
| Control |
N |
35.0 ± 0.5a |
7.7 ± 0.5a |
4.0 ± 0.0a |
6.0 ± 0.0a |
| Glycine (1 g kg−1) |
P |
22.0 ± 0.5b |
6.0 ± 0.6a |
2.0 ± 0.0b |
6.0 ± 0.0a |
| Glycine (2 g kg−1) |
Z |
27.0 ± 0.5c |
7.3 ± 1.2a |
2.7 ± 1.2b |
6.0 ± 0.0a |
| Glycine (3 g kg−1) |
R |
34.0 ± 0.5a |
7.3 ± 1.2a |
3.3 ± 1.2a |
6.0 ± 0.0a |
| Oxidized oil (3 g kg−1) |
O |
63.0 ± 1.2d |
9.3 ± 3.1a |
4.0 ± 2.0a |
8.0 ± 0.0b |
| Un-oxidized oil (3 g kg−1) |
U |
30.0 ± 0.5e |
8.0 ± 0.0a |
4.0 ± 0.0a |
6.7 ± 0.5a |
| OM (3 g kg−1) + glycine (1 g kg−1) |
W |
34.0 ± 1.0a |
10.0 ± 0.0b |
4.0 ± 0.0a |
7.7 ± 0.5b |
| OM (3 g kg−1) + glycine (2 g kg−1) |
X |
35.0 ± 0.5a |
11.0 ± 1.2b |
4.0 ± 0.0a |
6.7 ± 1.2a |
| OM (3 g kg−1) + glycine (3 g kg−1) |
Y |
39.0 ± 0.5f |
6.7 ± 1.2a |
2.0 ± 0.0a |
6.0 ± 0.0a |
| Control |
C |
33.0 ± 1.5a |
9.0 ± 1.0a |
4.0 ± 0.0a |
6.0 ± 0.0a |
| Glutamic acid (1 g kg−1) |
A |
28.0 ± 0.5b |
6.7 ± 1.2a |
4.0 ± 0.0a |
5.3 ± 1.2a |
| Glutamic acid (2 g kg−1) |
B |
45.0 ± 1.0c |
8.0 ± 0.0a |
4.0 ± 0.0a |
6.0 ± 0.0a |
| Glutamic acid (3 g kg−1) |
D |
50.0 ± 0.0d |
8.7 ± 1.2a |
4.0 ± 0.0a |
6.7 ± 1.2a |
| Oxidized oil (3 g kg−1) |
E |
62.0 ± 0.5e |
13.0 ± 1.2b |
5.3 ± 1.2a |
8.0 ± 0.0b |
| Un-oxidized oil (3 g kg−1) |
F |
29.0 ± 1.2b |
8.1 ± 2.0a |
4.0 ± 2.0a |
6.0 ± 0.0a |
| OM (3 g kg−1) + glutamic acid (1 g kg−1) |
G |
47.0 ± 1.0c |
8.0 ± 0.0a |
4.7 ± 1.2a |
6.0 ± 0.0a |
| OM (3 g kg−1) + glutamic acid (2 g kg−1) |
H |
57.0 ± 1.2f |
9.3 ± 1.2a |
4.0 ± 0.0a |
6.0 ± 0.0a |
| OM (3 g kg−1) + glutamic acid (3 g kg−1) |
M |
62.0 ± 1.5e |
11.0 ± 1.2c |
4.0 ± 0.0a |
5.3 ± 1.2a |
Serum biochemistry
The serum cholesterol increased by the ingestion of un-oxidized and thermally oxidized mustard oil, while supplementation of glycine and glutamic acid normalizes the serum cholesterol level. When the amino acids were fed along with oxidized mustard oil the cholesterol level tends toward normal as shown in Table 5. The results are in agreement with previous work which showed that heated mustard oil at 210 °C significantly increased the level of serum cholesterol in rats compared to fresh mustard oil.28 A significant (P < 0.05) increase in the triglyceride level was also observed when the rabbits were fed with the oxidized mustard oil. The amino acids in the present study normalize triglyceride levels when fed in combination with oxidized mustard oil. The serum high density lipoproteins (HDL)-cholesterol was elevated after the use of oxidized mustard oil. Both amino acids have been found to normalize low density lipoproteins (LDL) and HDL-cholesterol. The negative effects on serum lipids were highly correlated with the reported work of Chacko and Rajamohan.28 Khan et al.29 showed that mustard seeds when supplemented with diet normalize the serum lipid profile in rats. This suggests that mustard seeds or its oil were helpful in normalizing the serum biochemical parameters. The supplementation of glycine and glutamic acid was also found to normalize the glucose level. The present findings are in accordance with ref. 16 which showed that oxidized lipids significantly decrease the serum glucose level. The supplementation of glycine and glutamic acid was beneficial in normalizing the serum glucose level and may be helpful in the management of diabetes.30 The liver cells generate an enzyme called serum glutamate pyruvate transaminase, the excess production of this enzyme was considered as an indicator of damage to the liver tissues. The inflammation of the liver occurs by the administration of drugs, infiltration of fats or disease of the bile duct.15 When an injury occurs to the liver cells the level of ALT rises. In the present study the level of the ALT has increased with the increase in the dose of amino acids, but a significant (P < 0.05) decrease was found in the ALT after administration of mustard seed oil and the selected amino acids. These results showed that the toxic effects of oxidized mustard oil can be overcome by the supplementation of glycine and glutamic acid.
Table 5 Effect of dietary supplementation of glycine, glutamic acid alone or in combination with oxidized mustard oil on the serum biochemistry of rabbit
| Treatments |
Code |
Serum total cholesterol (mg dL−1) |
Serum total TG (mg dL−1) |
Serum HDL cholesterol (mg dL−1) |
Serum LDL-cholesterol (mg dL−1) |
Serum glucose (mg dL−1) |
Serum ALT/SGPT (IU) |
| Values are mean ± SD of triplicates. Different superscript letters (a–f) represent significant difference from N (control of glycine group) or C (control of glutamic acid) at P < 0.05 using one way analysis of variance in column statistics. OM represents oxidized mustard oil. TG, triacylglycerols; HDL, high density lipoproteins; LDL, low density lipoproteins; ALT, alanine aminotransferase. |
| Control |
N |
62.0 ± 2.6a |
63.0 ± 2.6a |
36.0 ± 1.5a |
19.0 ± 2.4a |
70.0 ± 1.0a |
44.0 ± 1.0a |
| Glycine (1 g kg−1) |
P |
63.0 ± 1.5a |
62.0 ± 0.58a |
35.0 ± 1.0a |
16.0 ± 0.5a |
76.0 ± 1.0b |
45.0 ± 0.5a |
| Glycine (2 g kg−1) |
Z |
61.0 ± 1.0b |
60.0 ± 1.0a |
33.0 ± 1.0a |
16.0 ± 1.9a |
79.0 ± 0.5c |
47.0 ± 0.5a |
| Glycine (3 g kg−1) |
R |
56.0 ± 1.5c |
58.0 ± 1.5a |
32.0 ± 0.5a |
12.0 ± 2.2b |
81.0 ± 2.0c |
51.0 ± 1.0b |
| Oxidized oil (3 g kg−1) |
O |
84.0 ± 4.9d |
84.0 ± 3.8b |
46.0 ± 1.0b |
21.0 ± 2.5a |
50.0 ± 4.0d |
31.0 ± 1.5c |
| Un-oxidized oil (3 g kg−1) |
U |
75.0 ± 1.2e |
76.0 ± 1.5c |
42.0 ± 1.0c |
18.0 ± 1.4a |
53.0 ± 3.1d |
34.0 ± 0.5d |
| OM (3 g kg−1) + glycine (1 g kg−1) |
W |
81.0 ± 2.1d |
82.0 ± 3.1d |
39.0 ± 1.5c |
25.0 ± 3.0c |
55.0 ± 1.5d |
38.0 ± 1.0e |
| OM (3 g kg−1) + glycine (2 g kg−1) |
X |
76.0 ± 2.5f |
77.0 ± 1.0e |
36.0 ± 1.5a |
25.0 ± 1.1c |
59.0 ± 1.5e |
42.0 ± 2.0a |
| OM (3 g kg−1) + glycine (3 g kg−1) |
Y |
71.0 ± 3.1a |
77.0 ± 5.9c |
36.0 ± 1.0a |
19.0 ± 1.7a |
74.0 ± 1.1b |
44.0 ± 1.0a |
| Control |
C |
66.0 ± 1.0a |
64.0 ± 1.0a |
37.0 ± 1.0a |
17.0 ± 0.6a |
71.0 ± 1.5a |
42.0 ± 2.1a |
| Glutamic acid (1 g kg−1) |
A |
63.0 ± 2.9a |
63.0 ± 1.0a |
37.0 ± 2.0a |
14.0 ± 1.6b |
71.0 ± 1.9a |
43.0 ± 1.5a |
| Glutamic acid (2 g kg−1) |
B |
60.0 ± 1.5b |
62.0 ± 0.5a |
35.0 ± 1.0a |
12.0 ± 0.6c |
73.0 ± 1.0a |
43.0 ± 1.0a |
| Glutamic acid (3 g kg−1) |
D |
57.0 ± 1.0c |
57.0 ± 1.5b |
33.0 ± 1.5b |
12.0 ± 2.2c |
74.0 ± 2.1a |
45.0 ± 2.6a |
| Oxidized oil (3 g kg−1) |
E |
85.0 ± 1.5d |
76.0 ± 2.0c |
46.0 ± 1.7c |
24.0 ± 2.3d |
49.0 ± 1.0b |
30.0 ± 1.0b |
| Un-oxidized oil (3 g kg−1) |
F |
77.0 ± 1.5e |
72.0 ± 2.1c |
43.0 ± 1.0d |
19.0 ± 0.9e |
64.0 ± 1.0c |
38.0 ± 1.5a |
| OM (3 g kg−1) + glutamic acid (1 g kg−1) |
G |
76.0 ± 0.5f |
69.0 ± 2.1c |
41.0 ± 0.5d |
20.0 ± 1.5a |
58.0 ± 1.5d |
34.0 ± 2.1c |
| OM (3 g kg−1) + glutamic acid (2 g kg−1) |
H |
71.0 ± 1.5g |
66.0 ± 1.5a |
38.0 ± 1.0a |
19.0 ± 1.4a |
61.0 ± 2.0d |
41.0 ± 3.1a |
| OM (3 g kg−1) + glutamic acid (3 g kg−1) |
M |
67.0 ± 1.2a |
64.0 ± 1.2a |
37.0 ± 1.5a |
17.0 ± 2.3a |
66.0 ± 1.7c |
45.0 ± 1.5a |
Haematological indices
Table 6 shows no significant changes in the haematological parameters such as haemoglobin, platelets, total leukocyte count, lymphocytes, neutrophils, eosinophils and monocytes with the supplementation of glycine and glutamic acid alone. The ingestion of oxidized lipids produced toxic effects on all the parameters. Our previous reports15,16 also showed that thermally oxidized lipids produced toxic effects in the haematological indices of the experimental animals. The co-administration of glycine and glutamic acid was highly beneficial in normalizing the levels of haematological parameters.
Table 6 Effect of dietary supplementation of glycine, glutamic acid alone or in combination with oxidized mustard oil on the haematological parameters of rabbits
| Treatments |
Code |
Hb (g dL−1) |
Platelets (×103) cells per μL |
TLC (×103) cells per μL |
Blood lymphocytes (%) |
Neutrophils (%) |
Eosinophils (%) |
Monocytes (%) |
| Values are mean ± SD of triplicates. Different letters (a–f) represent significant difference from N (control of glycine group) or C (control of glutamic acid) at P < 0.05 using one way analysis of variance in column statistics. OM represents oxidized mustard oil. TLC, total leukocytes counts. |
| Control |
N |
13.0 ± 0.6a |
223.0a |
6.7 |
61.0 ± 2.0a |
31.0 ± 1.5a |
2.0 ± 0.0a |
2.7 ± 0.5a |
| Glycine (1 g kg−1) |
P |
13.0 ± 0.3a |
223.1a |
7.0 |
64.0 ± 1.5a |
31.0 ± 2.0a |
2.1 ± 0.05a |
3.3 ± 1.5a |
| Glycine (2 g kg−1) |
Z |
14.0 ± 0.1a |
227.2b |
7.1 |
65.0 ± 2.0a |
31.0 ± 1.5a |
2.1 ± 0.1a |
2.3 ± 0.5a |
| Glycine (3 g kg−1) |
R |
14.0 ± 0.1a |
230.3c |
7.1 |
63.0 ± 1.5a |
30.0 ± 1.0a |
2.1 ± 0.05a |
2.3 ± 0.5a |
| Oxidized oil (3 g kg−1) |
O |
11.0 ± 0.2b |
213.2d |
9.1 |
39.0 ± 2.0b |
53.0 ± 2.5b |
2.2 ± 0.2a |
3.3 ± 0.5a |
| Un-oxidized oil (3 g kg−1) |
U |
12.0 ± 0.3c |
217.0e |
8.5 |
47.0 ± 2.5c |
48.0 ± 2.1c |
2.1 ± 0.05a |
3.0 ± 0.0a |
| OM (3 g kg−1) + glycine (1 g kg−1) |
W |
11.0 ± 0.1b |
215.0f |
8.6 |
43.0 ± 2.5d |
43.0 ± 1.5d |
1.8 ± 0.1b |
3.0 ± 1.0a |
| OM (3 g kg−1) + glycine (2 g kg−1) |
X |
12.0 ± 0.2a |
218.0e |
8.1 |
46.0 ± 3.1c |
39.0 ± 1.0e |
2.1 ± 0.2a |
2.3 ± 0.5a |
| OM (3 g kg−1) + glycine (3 g kg−1) |
Y |
12.0 ± 0.2a |
220.0a |
7.8 |
48.0 ± 3.1e |
34.0 ± 1.5a |
1.9 ± 0.1a |
2.0 ± 0.0a |
| Control |
C |
13.0 ± 0.8a |
221.0a |
6.9 |
59.0 ± 1.5a |
33.0 ± 2.6a |
1.7 ± 0.2a |
2.7 ± 0.5a |
| Glutamic acid (1 g kg−1) |
A |
13.0 ± 0.3a |
223.0a |
7.1 |
62.0 ± 1.5b |
29.0 ± 1.5a |
1.8 ± 0.2a |
3.0 ± 1.0b |
| Glutamic acid (2 g kg−1) |
B |
14.0 ± 0.3a |
227.1b |
7.2 |
59.0 ± 1.5b |
29.0 ± 2.5a |
1.8 ± 0.2a |
3.0 ± 0.0b |
| Glutamic acid (3 g kg−1) |
D |
14.0 ± 0.7a |
230.4c |
7.3 |
60.0 ± 2.1b |
30.0 ± 3.1a |
2.0 ± 0.0a |
2.7 ± 0.5a |
| Oxidized oil (3 g kg−1) |
E |
11.0 ± 0.3b |
113.2d |
8.7 |
47.0 ± 2.6c |
51.0 ± 3.5b |
2.4 ± 0.1a |
3.7 ± 0.5c |
| Un-oxidized oil (3 g kg−1) |
F |
12.0 ± 0.4c |
118.1e |
7.9 |
52.0 ± 1.2d |
38.0 ± 1.5c |
2.1 ± 0.1a |
2.7 ± 1.2a |
| OM (3 g kg−1) + glutamic acid (1 g kg−1) |
G |
11.0 ± 0.1b |
114.0d |
8.3 |
49.0 ± 2.5e |
43.0 ± 1.5d |
1.9 ± 0.1a |
2.7 ± 0.5a |
| OM (3 g kg−1) + glutamic acid (2 g kg−1) |
H |
12.0 ± 0.1c |
116.0e |
8.1 |
53.0 ± 1.0d |
39.0 ± 2.5c |
2.0 ± 0.1a |
3.0 ± 1.0a |
| OM (3 g kg−1) + glutamic acid (3 g kg−1) |
M |
13.0 ± 0.2a |
118.1e |
7.7 |
59.0 ± 1.2b |
35.0 ± 3.2a |
2.0 ± 0.05a |
2.3 ± 1.2a |
Liver histopathology
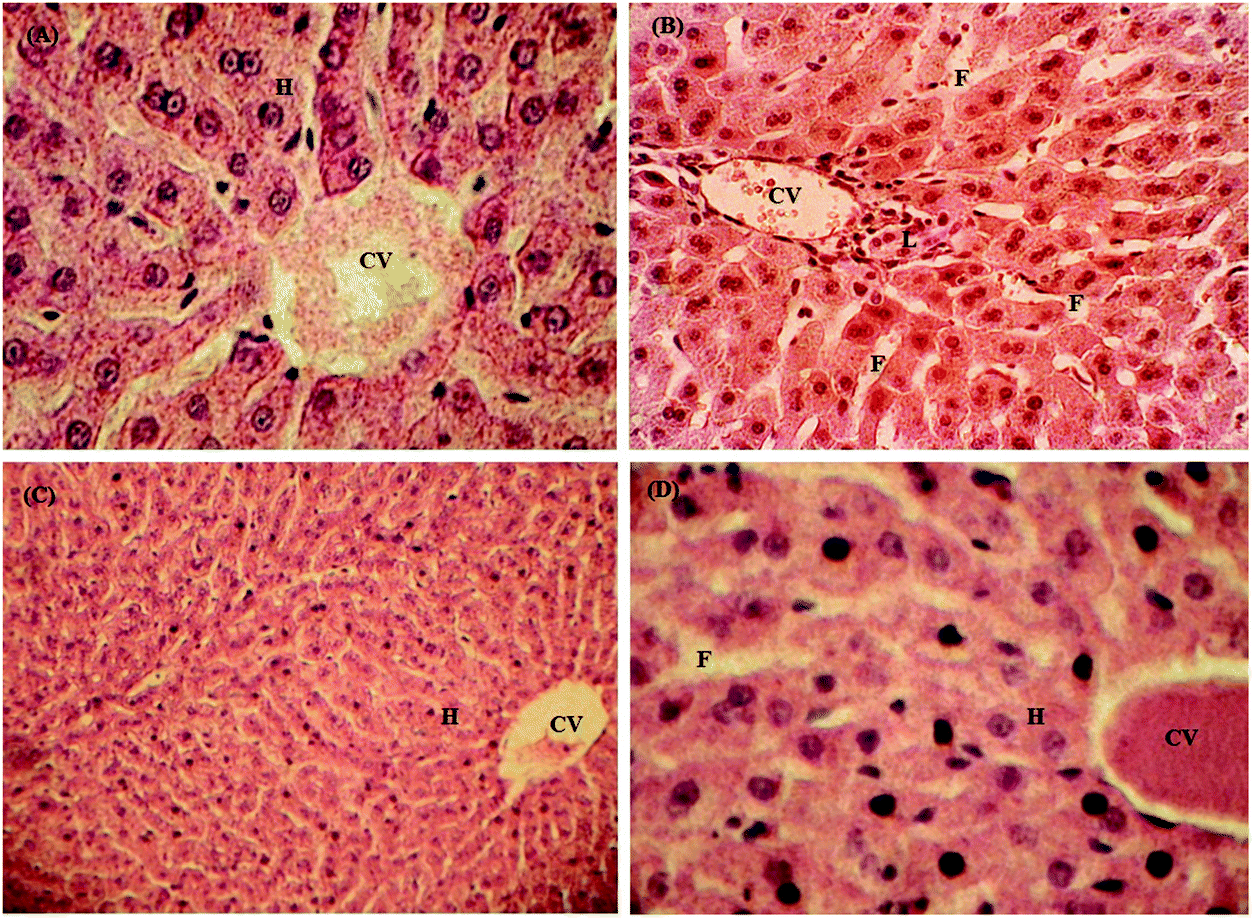
The oxidized mustard oil produces toxic effects on the liver architecture as shown in Fig. 2. The oxidized oil affects the hepatocytes by damaging their boundaries; there is also the accumulation of fats and reactive Kupffer cells (Fig. 2B) compared to control (Fig. 2A), while there was less toxicity after feeding the un-oxidized mustard oil to rabbits. The feeding of oxidized mustard oil resulted in obvious fat accumulation marked as F (Fig. 2B). The lymphocyte accumulation marked as L was higher with coagulative necrosis. In the case of supplementation of both amino acids alone, no harmful effects were observed. The feeding of both amino acids in combination with OM showed a recovery of the liver structure and functions as indicated in Fig. 2C, D and ALT levels in Table 4. Glutamic acid supplementation in combination with OM showed a mild fat accumulation marked as F as shown in Fig. 2D. Senthilkumar et al.21 also reported that glycine significantly inhibits severe oxidative stress in the liver. Thus it may be concluded that both glycine and glutamic acid act as antioxidants and are beneficial. This beneficial effect of glycine and glutamic acid may be partly explained by the fact that glycine and glutamic acid increased the influx of chloride into Kupffer cells, leading to reduction of the production of the tumour necrosis factor (TNF-α).31 These results indicate that ingestion of diet containing oxidized lipids may cause negative effects which can be overcome by co-administration of glycine or glutamic acid.
 |
| | Fig. 2 Representative liver cross section of the biochemical response of glycine and glutamic acid toward the toxic effects of oxidized mustard oil in rabbits. (A) Control group, (B) oxidized mustard oil, (C) glycine with oxidized oil and (D) glutamic acid with oxidized oil. Photomicrograph at X40. Letters CV, H, L and F represent the central vein, hepatocytes, lymphocytes and fats respectively. | |
Conclusions
Mustard seed oil was thermally oxidized and analysed for quality characteristics and phenolic profile. Fourteen polyphenolic compounds were identified and quantified with significant losses during oxidation. Glycine and glutamic acid protect against oxidative stress produced by oxidized mustard oil. The oxidized mustard oil also declined the body weight, but co-administration of amino acids increased the body weight of rabbits. The oxidized mustard oil increased the level of cholesterol, triglycerides, and HDL-cholesterol, while the ALT and glucose level were declined. There was no effect of oxidized mustard oil on the level of LDL-cholesterol. Both amino acids declined the level of cholesterol, triglycerides and HDL-cholesterol and increased the level of glucose. The oxidized oil also decreases the levels of haemoglobin, blood lymphocytes and platelet concentrations. But the co-administration of the amino acids normalizes the level of these parameters. The oxidized oil increased the TLC, neutrophils and eosinophils counts, but were normal after the co-administration of amino acids. The oxidized mustard oil significantly alters the architecture of the liver, enhances fat accumulation, and causes hepatitis and necrosis. Co-administration of glycine or glutamic acid normalized the liver structure and function. It is therefore suggested that glycine and glutamic acid are beneficial and may be considered as supplements for oxidative stress and adverse effects induced by dietary oxidized lipids.
Conflict of interest
The authors have no conflict of interest with anybody or institution.
Acknowledgements
The authors would like to thank Higher Education Commission (HEC) Pakistan for supporting this work through research grant no. 2344. Mr Fareed Ullah Research Assistant at Department of Biotechnology is highly acknowledged for his support during oil analyses.
References
- W. Wang, Z. Wu, Z. Dai, Y. Yang, J. Wang and G. Wu, Amino acids, 2013, 45, 463–477 CrossRef CAS PubMed.
- C.-Y. Chen, B.-T. Wang, Z.-C. Wu, W.-T. Yu, P.-J. Lin, W.-L. Tsai and S.-C. Shiesh, Clin. Chim. Acta, 2013, 420, 150–154 CrossRef CAS PubMed.
- I. Amelio, F. Cutruzzolá, A. Antonov, M. Agostini and G. Melino, Trends Biochem. Sci., 2014, 39, 191–198 CrossRef CAS PubMed.
- Y. Ding, G. F. T. Svingen, E. R. Pedersen, J. F. Gregory, P. M. Ueland, G. S. Tell and O. K. Nygård, J. Am. Heart Assoc., 2016, 5, e002621 CrossRef PubMed.
- M. E. Hafidi, I. Pérez, J. Zamora, V. Soto, G. Carvajal-Sandoval and G. Baños, Am. J. Physiol. Regul. Integr. Comp. Physiol., 2004, 287, R1387–R1393 CrossRef PubMed.
- A. Ruiz-Ramirez, E. Ortiz-Balderas, G. Cardozo-Saldana, E. Diaz-Diaz and M. El-Hafidi, Clin. Sci., 2014, 126, 19–29 CrossRef CAS PubMed.
- R. Senthilkumar, M. Sengottuvelan and N. Nalini, Cell Biochem. Funct., 2004, 22, 123–128 CrossRef CAS PubMed.
- P. Morales, I. C. F. R. Ferreira, A. M. Carvalho, M. C. Sánchez-Mata, M. Cámara, V. Fernández-Ruiz, M. Pardo-de-Santayana and J. Tardío, LWT–Food Sci. Technol., 2014, 55, 389–396 CrossRef CAS.
- B. C. Nair, S. R. Krishnan, G. R. Sareddy, M. Mann, B. Xu, M. Natarajan, P. Hasty, D. Brann, R. R. Tekmal and R. K. Vadlamudi, Cell Death Differ., 2014, 21, 1409–1418 CrossRef CAS PubMed.
- I. Ali, W. A. Wani, K. Saleem and M.-F. Hsieh, RSC Adv., 2014, 4, 29629–29641 RSC.
- S. Dutta, S. Ray and K. Nagarajan, Saudi Pharm. J., 2013, 21, 337–343 CrossRef PubMed.
- J. Yin, M. Liu, W. Ren, J. Duan, G. Yang, Y. Zhao, R. Fang, L. Chen, T. Li and Y. Yin, PLoS One, 2015, 10, e0122893 Search PubMed.
- A. Zeb, Free Radical Res., 2015, 49, 549–564 CrossRef CAS PubMed.
- A. Zeb and A. Mehmood, Turk. J. Biochem., 2012, 37, 417–423 CrossRef CAS.
- A. Zeb and I. Haq, J. Food Biochem., 2016, 40, 428–435 CrossRef CAS.
- A. Zeb and S. Ullah, Food Chem., 2015, 186, 6–12 CrossRef CAS PubMed.
- A. A. A. Mohdaly, I. Smetanska, M. F. Ramadan, M. A. Sarhan and A. Mahmoud, Ind. Crops Prod., 2011, 34, 952–959 CrossRef CAS.
-
AOCS, Official methods and recommended practices of the American oil chemist society, AOCS press, Champaign, 1998 Search PubMed.
- A. Zeb and F. Ullah, J. Anal. Methods Chem., 2016, 2016, 5 Search PubMed.
- A. Zeb, Anal. Methods, 2015, 7, 7753–7757 RSC.
- R. Senthilkumar, P. Viswanathan and N. Nalini, Pharmazie, 2004, 59, 55–60 CAS.
- C. Janu, D. Kumar, M. Reshma, P. Jayamurthy, A. Sundaresan and P. Nisha, J. Food Biochem., 2014, 38, 38–49 CrossRef CAS.
- A. Spielmeyer, A. Wagner and G. Jahreis, Food Chem., 2009, 112, 944–948 CrossRef CAS.
- H.-I. Jun, D. P. Wiesenborn and Y.-S. Kim, Food Sci. Biotechnol., 2014, 23, 1753–1760 CrossRef CAS.
- C. Engels, A. Schieber and M. G. Gänzle, Eur. Food Res. Technol., 2012, 234, 535–542 CrossRef CAS.
- A. Zeb and S. Hussain, J. Food Biochem., 2014, 38, 498–508 CrossRef CAS.
- S. Saha, N. Ahmad, S. Majumder, M. Hosain and M. Miah, Bangladesh J. Vet. Med., 2012, 3, 79–81 CrossRef.
- C. Chacko and T. Rajamohan, Int. J. Biol. Med. Res., 2011, 2, 991–997 Search PubMed.
- B. A. Khan, A. Abraham and S. Leelamma, Plant Foods Hum. Nutr., 1996, 49, 295–299 CrossRef CAS PubMed.
- X. Bi, J. Lim and C. J. Henry, Food Chem., 2017, 217, 281–293 CrossRef CAS PubMed.
- M. Yin, K. Ikejima, G. E. Arteel, V. Seabra, B. U. Bradford, H. Kono, I. Rusyn and R. G. Thurman, J. Pharmacol. Exp. Ther., 1998, 286, 1014–1019 CAS.
|
| This journal is © The Royal Society of Chemistry 2017 |
Click here to see how this site uses Cookies. View our privacy policy here. 
 * and
Saleem Ur
Rahman
* and
Saleem Ur
Rahman