DOI:
10.1039/C6FO01207H
(Paper)
Food Funct., 2017,
8, 291-298
Medium-chain fatty acids reduce serum cholesterol by regulating the metabolism of bile acid in C57BL/6J mice
Received
13th August 2016
, Accepted 5th December 2016
First published on 7th December 2016
Abstract
Hypercholesterolemia is one of the important risk factors of atherosclerosis (AS). The aim of this study is to explore the effect of medium-chain fatty acids (MCFAs) on serum cholesterol levels and their mechanism of action. Hyperlipemia, as a model of abnormal lipid hypermetabolism, was established by using a high fat diet in C57BL/6J mice. Forty eight mice with dyslipidemia were randomly divided into 4 groups, 12 mice per group, including the control group, the 2% caprylic acid (C8:0)-treated group, 2% capric acid (C10:0)-treated group, and 2% oleic acid (C18:1)-treated group. All mice were fed with a high fat diet. After 16 weeks, the mice were anesthetized with chloral hydrate. The mouse portal vein blood, the liver and the start site of the ileum (1 cm) were collected. The body weight of the mice and blood lipid profiles were measured. Gene transcription and the expression level associated with bile acid metabolism in the liver and small intestine were determined by real-time PCR and the western blotting method. The concentrations of bile acid metabolites in bile and feces were analysed. After 16 weeks of treatment, the concentrations of TC and LDL-C in the caprylic acid group were significantly lower than those in the control group (P < 0.05); the transcription and expression level of LXR, CYP7A1, CYP27A1 and ABCG8 in the caprylic acid and capric acid groups were significantly higher than those in the control group in the liver (P < 0.05), however the transcription and expression level of the small heterodimer partner (SHP) were significantly lower than those in the control group (P < 0.05); the transcription and expression level of LXR, ABCG5 and ABCG8 in the caprylic acid, capric acid and oleic acid groups were significantly higher than those in the control group in the small intestine (P < 0.05). The concentrations of total bile acid, mainly cholic acid and cholesterol in bile and feces were significantly higher in the caprylic and capric acid groups than those of in the control group (P < 0.05). Thus, MCFA increased the expression of LXR and ABCG8, enhanced CYP7A1 and CYP27A1 expression, decreased and SHP expression in the liver, thereby promoted liver bile acid synthesis and excretion. In addition MCFA increased the expression of ABCG5, ABCG8 and LXR in the small intestine, thereby inhibiting small intestinal bile acid absorption, increasing the concentrations of cholesterol and bile acid in bile and feces and reducing the level of serum cholesterol.
Introduction
Coronary artery disease (CAD) is a leading cause of morbidity and mortality in both developing and developed countries.1 High cholesterol is one of the risk factors for CAD, and many randomized controlled trials have demonstrated reduced CAD event rates when this risk factor is treated with medications.2 In addition, lifestyle modifications such as smoking cessation, achievement and maintenance of an ideal body weight, regular exercise, reduction in the intake of saturated fat and sugars, and decreasing the level of stress are the cornerstone of CAD prevention strategies and are the first steps in risk factor management.3,4 Medium-chain fatty acids (MCFAs) composed primarily of glycerides of caprylic (C8:0) and capric (C10:0) acids which differs from long-chain fatty acids (LCFAs, 14 or more carbon atoms) and has no effect on cholesterol biosynthesis,5,6 can lower total cholesterol and maintain low density lipoprotein cholesterol levels.7–9 It was reported that MCFAs are metabolized somewhat differently from LCFA.10 Esterification of MCFA is limited in hepatocytes and MCFA undergoes preferential oxidation and behaves more like glucose rather than fat. Eventually, a larger proportion of MCFA might be used for oxidation rather than being stored in tissues.11,12 The difference between the metabolism of MCFA and LCFA may contribute to the hypocholesterolemic effect of MCFA. In our previous study, we investigated that MCFA might reduce blood cholesterol by promoting the excretion of fecal cholesterol.7 However, the definite mechanism in charge of the reduction of cholesterol is unclear. In this study we will observe the effect of MCFA on bile acid metabolism in C57BL/6J mice fed with a high fat diet, compare with oleic acid (C18:1) and investigate the mechanism of action.
Materials & methods
Chemicals and reagents
Antibodies targeting the BSEP (bile salt export pump), ABCG8 (ATP-binding cassette sub-family G member 8), ABCG5 (ATP-binding cassette sub-family G member 5), FXR (Farnesoid X Receptor), SHP (small heterodimer partner), LXR (Liver X Receptor), CYP7A1 (cytochrome P450 7A1), CYP27A1 (cytochrome P450 27A1) and GAPDH (glyceraldehyde 3-phosphate dehydrogenase) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Secondary antibodies against rabbit, mouse or goat IgG were obtained from Cell Signaling Technology (Danvers, MA, USA). A prestained protein marker p7708V was purchased from New England Biolabs, Ltd (Beijing, China). A western blotting luminol reagent and PVDF membranes were purchased from Millipore (Billerica, MA, USA). Cholesterol (no. C8667), cholestenol (no. 47129), coprostanol (C7578), cholic acid (CA, no. C1129), deoxycholic acid (DCA, no. D2510), chenodeoxycholic acid (CDCA, no. C9377), ursodeoxycholic acid (UDCA, no. U5127), lithocholic acid (LCA, no. L6250), taurocholic acid (TCA, no. T4009), taurochenodeoxycholic acid (TCDCA, no. T6260), glycocholic acid (GCA, no. G2878), and glycochenodeoxycholic acid (GCDCA, no. 739715), purchased from Sigma-Aldrich, were used as standards.
Animals
Male C57BL/6J mice (n = 100) purchased from the Institute of Laboratory Animal Science, the Chinese Academy of Medical Science (license no. SCXK: Jing2009-007) were housed in a temperature-controlled environment (22 ± 2 °C, humidity 40% to 60%) with a 12 h light/dark cycle. The animals (18 ± 4 g) had free access to food and water. The experiments with the animals were conducted in accordance with the guidelines of the Animal Care and Use Committee of the Chinese PLA General Hospital (Beijing, China), and approved by the animal ethical committee of Chinese PLA General Hospital (Beijing, China).
Diets
During the first week, all animals were fed a commercial standard diet (purchased from the Academy of Military Medicine Science and based on the AIN-93G diet) and afterwards, fifteen mice were randomly selected and fed with the standard diet as a normal control and the rest of the mice were fed with a high-fat diet. After 4 weeks, mice whose serum total cholesterol (collected from the mandibular venous plexus) were over 40% more than those of the normal control mice were assigned to one of the four weight-matched groups for each experiment. Thereafter, three groups were fed one of the three high-fat diets containing caprylic acid, capric acid or oleic acid for 16 weeks (Table 1). The mice in the control group were fed a high-fat diet.
Table 1 Compositions of diets with different fatty acids (C8:0, C10:0 and C18:1) for C57BL/6J mice
| Ingredients |
HF |
C8:0 |
C10:0 |
C18:1 |
| HF, high fat diet; C8:0, caprylic acid; C10:0, capric acids; C18:1, oleic acid. |
| High fat diet (%) |
92 |
90 |
90 |
90 |
| Casein (%) |
8 |
8 |
8 |
8 |
| C8:0 (%) |
|
2 |
|
|
| C10:0 (%) |
|
|
2 |
|
| C18:1 (%) |
|
|
|
2 |
| Energy (kJ g−1) |
18.81 |
18.81 |
18.81 |
18.81 |
| |
|
Percentage of nutrients
|
| Protein (%) |
17.65 |
17.65 |
17.65 |
17.65 |
| Fat (%) |
21.03 |
21.03 |
21.03 |
21.03 |
| Carbohydrate (%) |
47.5 |
47.5 |
47.5 |
47.5 |
| Mineral mixture (%) |
0.79 |
0.79 |
0.79 |
0.79 |
| Vitamin mixture (%) |
0.51 |
0.51 |
0.51 |
0.51 |
| Fiber (%) |
1.5 |
1.5 |
1.5 |
1.5 |
| Water (%) |
9.5 |
9.4 |
9.4 |
9.4 |
| Others (%) |
5.5 |
5.6 |
5.6 |
5.6 |
Feces, blood, tissue and bile sampling
After 16 weeks of feeding, six mice were chosen randomly from each group to record the diet intake and feces sample output through limited metabolic cages. Daily diet intake and the feces output of each mouse were recorded for 3 days at the end of the study. The feces were lyophilized, weighed, pulverised, and stored at −80 °C until further analysis. At the end of the experiment, the mice were deprived of the diet, but not water, for fasting overnight (at least 12 h). In the morning after fasting, the mice were anesthetized and their blood samples were collected from the aorta ventralis. Several technicians worked together to complete the blood and tissue sampling as rapidly as possible. The mesenteric, epididymal and perirenal fat pads, and the tissues of the liver and small intestine (mainly ileum) were excised, rinsed with ice-cold saline and weighed. Parts of the liver and small intestine were immediately frozen in liquid nitrogen and stored at −80 °C. The microcapsule was used to puncture the gallbladder wall to extract the bile, and each bile sample was centrifuged at 16![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) 000g for 30 min, and the supernatant was collected and stored −80 °C for later analysis.
000g for 30 min, and the supernatant was collected and stored −80 °C for later analysis.
Measurement of serum chemistry
At the end of the experiment, serum was collected through centrifuging the blood samples. TG (triglyceride), TC (total cholesterol), LDL-C (low density lipoprotein-cholesterol), HDL-C (high density lipoprotein-cholesterol) and bile acids (BAs) in the serum were measured using standard protocols in the clinical laboratory.
Real-time quantitative PCR analysis
Primers were designed by using a type of Primer Express 3.0 software, based on the mRNA sequences from a database (Table 2) and were synthesized by Invitrogen (Beijing, China). Total RNA was isolated by using a Trizol reagent. RT-PCR was performed using a One Step SYBR® PrimeScript® RT-PCR Kit (Takara Biotechnology Co., Ltd, Dalian, China). Amplification was carried out with a BIO-RAD iCycler Thermal Cycler (BIO-RAD, Hercules, CA, USA). The PCR conditions were as follows: reverse transcription 42 °C, 5 min, denaturation at 95 °C for 10 s, followed by 40 cycles of 95 °C for 5 s, and 60 °C for 20 s. Relative mRNA expression levels were determined by the comparative critical threshold (Ct) method (in a separate tube). The house keeping gene, β-actin was used as a control for normalization.
Table 2 The PCR primers used in this study
| Target genes |
Forward primer (5′–3′) |
Reverse primer (5′–3′) |
| BSEP |
ACCACTACGGAGTTCACGC |
GACAGGCACTTGTGAAAACG |
| NTCP |
ACCGTTTCTTCGTGGATGG |
ACACAGTTCAGTGCTCGCTC |
| ABCG8 |
CACCCTTGTCCTCGCTAT |
TCCTTTGCCTCAGCTTTC |
| ABCG5 |
ATTGTCACCATCCACCAG |
GACAGGGGTAACCACAGT |
| LXR |
GTCAGCAGAGCCTACAGC |
TGTAATGGAAGCCAGAGG |
| SHP |
AGGATGCTGTGACCTTCG |
CAAGGCTCCAGAAAGACT |
| CYP7A1 |
ATGGAGAAGGCTAAGACG |
CACTTCTTCAGAGGCTGC |
| CYP27A1 |
CCAATGTGGACAACCTCCT |
CTTGTGGTCTCGGTGGTC |
| ASBT |
TTATTCCTGTTTCCTTTGG |
TTTGGTACAGTATTCCTCC |
| FXR |
TCCCATTTACAGGCTACG |
TCTGTGGCTGAACTTGAG |
| GAPDH |
CGTGTTCCTACCCCCAATG |
ATGTCATCATACTTGGCAGGTT |
Western blotting analysis
A 20 mg amount of frozen liver or small intestine tissue was added to 200 μL of a protein lysate solution, and homogenized with a glass grinder on ice. The homogenate was centrifuged at 14000g for 15 min at 4 °C to create a supernatant for subsequent measurement. The protein of the supernatant was measured by using the BCA kit, before the electrophoretic sample of 40 μg of the total protein was mixed with a 5-fold loading buffer; the mixture was boiled for 5 min and then immediately cooled in an ice box. The sample was size-fractionated on 10% SDS–PAGE gel and then electrophoretically transferred onto a polyvinylidene fluoride membrane. Next, the membrane was incubated overnight at 4 °C with the appropriate primary antibodies and then incubated with the appropriate secondary antibodies at room temperature for 2 h. Each membrane was developed using an enhanced ChemiImager5500 chemiluminescence system (Alpha Innotech Corporation, Miami, FL, USA).
Bile acid metabolites analysis
The gas chromatographic method and liquid chromatography–mass spectrometry were used. The instruments were Shimadzu GCMS-QP 2010 plus, Shimadzu UFLC-20AD and an Applied Biosystem 3200Q TRAP mass spectrometer with an electrospray ionization (ESI) source. Neutral steroid (including cholesterol, cholestanol and coprostanol), free bile acid (including CA, DCA, CDCA, UDCA, LCA) and binding bile acid (including TCA, TCDCA, GCA, GCDCA) in bile and feces were determined. The method of sample preparation and determination were carried out according to the report of Huanling Y,13 Kikuchi-Hayakawa H14 and our previous study.7
Statistical analysis
All data are expressed in mean ± standard deviation (SD). Data were analyzed by using one-way analysis of variance followed by the independent t-test to determine the significance of difference between groups using SPSS software version 17.0. Values at p < 0.05 were set to be statistically significant.
Results
Effects of MCFA on weight of body, liver and white adipose tissue
There were no significant differences in body weight between the control group, caprylic acid group, capric acid group and oleic acid group in the first 8 weeks of treatment. However the body weights of the caprylic acid group, capric acid group and oleic acid group were significantly lower than that of the control group after 12 or 16 weeks of treatment (Table 3). At the end of the experiment, liver, mesenteric white adipose tissue, perirenal fat and epididymal fat were collected and weighed. It was indicated that the liver weight, the weight of the mesenteric white adipose tissue, epididymal fat and body fat percentage in the caprylic acid group, capric acid group and oleic acid group were lower than those of the control group. However the weight of perirenal fat and the liver index were not different among every group (Table 4).
Table 3 The effect of different fatty acids (C8:0, C10:0 and C18:1) on body weight (mean ± SD, n = 12)
| Groups |
HF |
C8:0 |
C10:0 |
C18:1 |
| HF, high fat diet; C8:0, caprylic acid; C10:0, capric acids; C18:1, oleic acid; ap < 0.05, versus HF group. |
| 0 (weeks) |
26.23 ± 0.13 |
26.28 ± 0.12 |
26.27 ± 0.19 |
26.24 ± 0.18 |
| 4 (weeks) |
29.06 ± 1.23 |
28.73 ± 1.07 |
28.04 ± 1.29 |
28.33 ± 0.80 |
| 8 (weeks) |
31.24 ± 1.67 |
30.52 ± 1.55 |
30.01 ± 1.24 |
30.04 ± 1.28 |
| 12 (weeks) |
33.89 ± 2.19 |
31.78 ± 2.06a |
31.33 ± 1.62a |
31.48 ± 0.89a |
| 16 (weeks) |
36.21 ± 2.55 |
32.95 ± 1.97a |
32.66 ± 1.46a |
32.73 ± 0.95a |
Table 4 The effect of different fatty acids (C8:0, C10:0 and C18:1) on body fat mass (mean ± SD, n = 12)
| Groups |
HF |
C8:0 |
C10:0 |
C18:1 |
| HF, high fat diet; C8:0, caprylic acid; C10:0, capric acids; C18:1, oleic acid; ap < 0.05, versus HF group. |
| Liver weight (g) |
1.73 ± 0.08 |
1.53 ± 0.16a |
1.56 ± 0.13a |
1.55 ± 0.11a |
| Liver index (%) |
4.80 ± 0.36 |
4.64 ± 0.47 |
4.78 ± 0.40 |
4.73 ± 0.35 |
| Mesenteric fat (g) |
1.25 ± 0.57 |
0.79 ± 0.30a |
0.76 ± 0.32a |
0.77 ± 0.15a |
| Perirenal fat (g) |
0.46 ± 0.04 |
0.44 ± 0.04 |
0.43 ± 0.05 |
0.44 ± 0.03 |
| Epididymal fat (g) |
0.32 ± 0.08 |
0.22 ± 0.02a |
0.22 ± 0.02a |
0.23 ± 0.03a |
| Body fat percentage (%) |
5.53 ± 1.31 |
4.38 ± 0.82a |
4.27 ± 0.85a |
4.39 ± 0.50a |
Effects of MCFA on lipid profiles in blood
The levels of serum TC and LDL-C in the caprylic acid group, TC in the oleic acid group and TG in the capric group were significantly lower than that of the control group. The levels of blood HDL-C/LDL-C in the caprylic acid group, capric acid group and oleic acid group were significantly higher than that of the control group. In addition, total bile acids were not different among every group (Table 5).
Table 5 The effect of different fatty acids (C8:0, C10:0 and C18:1) on the blood lipid profile (mean ± SD, n = 12)
| Blood lipid profile |
HF |
C8:0 |
C10:0 |
C18:1 |
| HF, high fat diets; C8:0, caprylic acid; C10:0, capric acids; C18:1, oleic acid; ap < 0.05, versus HF group. |
| TC (mmol L−1) |
0.78 ± 0.18 |
0.58 ± 0.16a |
0.65 ± 0.19 |
0.54 ± 0.13a |
| LDL-c (mmol L−1) |
2.99 ± 0.28 |
2.06 ± 0.46a |
2.20 ± 0.37 |
2.12 ± 0.23 |
| HDL-c (mmol L−1) |
2.69 ± 0.84 |
3.29 ± 1.14 |
3.70 ± 1.38 |
3.21 ± 1.42 |
| HDL-c/LDL-c |
0.9 ± 0.54 |
1.60 ± 0.28a |
1.68 ± 0.46a |
1.51 ± 0.52a |
| TG (mmol L−1) |
3.38 ± 0.47 |
2.74 ± 0.47 |
2.54 ± 0.33a |
3.09 ± 0.26 |
| BA (μmol L−1) |
2.29 ± 0.50 |
2.58 ± 0.52 |
2.09 ± 0.74 |
2.37 ± 0.74 |
Effects of MCFA on mRNA of relevant factors of bile acid metabolism in liver and in small intestine
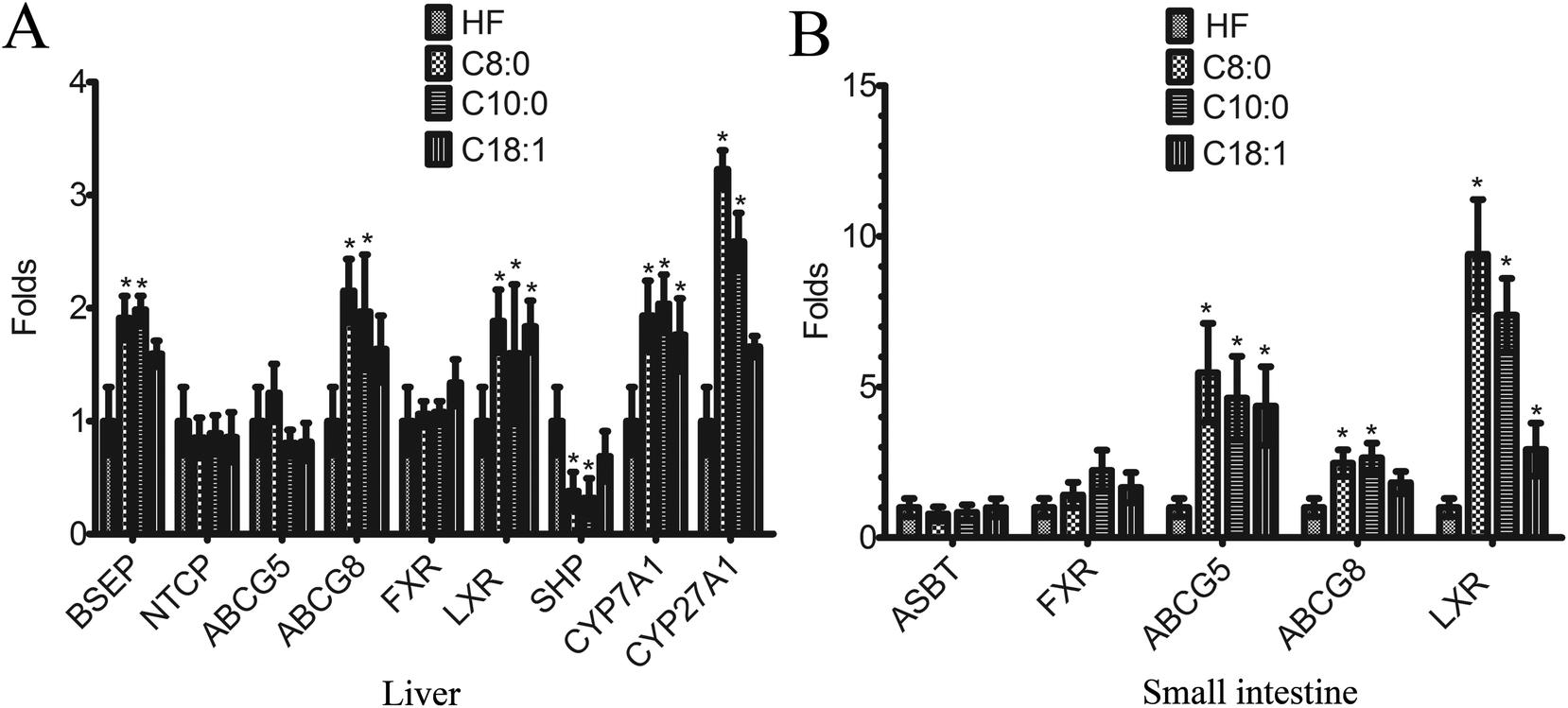
In liver, the transcription levels of CYP7A1 and LXR in caprylic acid, capric acid and oleic acid groups were higher than that in the control group, and mRNA expression of CYP27A1, BSEP and ABCG8 in caprylic acid and capric acid groups were higher than that in the control group, whereas SHP was lower in the control group. In the small intestine, the transcription levels of LXR and ABCG5 in the caprylic acid, capric acid and oleic acid groups were higher than the control group, and mRNA expression of ABCG8 in caprylic acid and capric acid groups were higher than that in the control group. However, the transcription levels of sodium taurocholate co-transport peptide (NTCP), FXR and ABCG5 in the liver and the apical sodium-dependent bile acid transporter (ASBT) and FXR in the small intestine had no difference among every group (Fig. 1).
 |
| | Fig. 1 Effect of different fatty acids (C8:0, C10:0 and C18:1) on the transcription of cholesterol metabolism related genes in the liver and small intestine of C57BL/6J mice. Total RNA was extracted from liver and small intestine tissues and mRNA transcription level was measured by real-time PCR analysis. The housekeeping gene, GAPDH, was used to normalize the expression level, and critical threshold (Ct) values were compared. *p < 0.05, versus high fat diet group. A. in liver; B. in small intestine. | |
Effects of MCFA on protein expression of relevant factors of bile acid metabolism in liver and in small intestine
In liver, the expression levels of CYP7A1, CYP27A1, LXR and ABCG8 in the caprylic acid and capric acid group were higher than that in the control group, however, the expression level of SHP was lower in the control group. In the intestine, the expression levels of LXR and ABCG5 in the caprylic acid, capric acid and oleic acid groups were higher than that in the control group. The expression levels of BSEP, FXR and ABCG5 in the liver and FXR and ABCG8 in the small intestine had no difference among every group (Fig. 2).
 |
| | Fig. 2 Effect of different fatty acids (C8:0, C10:0 and C18:1) on the expression of cholesterol metabolism related genes in liver and small intestine of C57BL/6J mice. Total protein was extracted from liver and small intestine tissues and the protein expression level was measured by western blot analysis. The housekeeping gene, GAPDH, was used to normalize the expression level, and relative light density values were compared. *p < 0.05, versus high fat diet group. A. section of blots in liver; B. gray-scale analysis in liver; C. section of blots in small intestine; D. gray-scale analysis in small intestine. | |
Effects of MCFA on bile acid metabolites in bile and feces
In bile, the concentrations of total BAs, including CA, CDCA, TCDCA, and GCDCA, and the level of cholesterol in the caprylic acid, capric acid and oleic acid groups were higher than that in the control group significantly (Table 6). In feces, the concentrations of total BAs, including CA, CDCA, LCA, TCDCA, and GCDCA, and the level of cholesterol in the caprylic acid and capric acid groups were higher than those in the control group significantly (Table 7). Furthermore, excretion of total BAs, CA, LCA in the caprylic acid group were significantly higher than that in the oleic acid group (Table 7). There were no significant differences in the excretion of DCA, UDCA, TCA, cholestanol, and coprostanol among the groups in either bile or feces (Tables 6 & 7). For the concentration of GCA in bile, no significant difference was shown, but in feces, there was a significant increase in the concentration in the caprylic acid group compared to that in the control group (Tables 6 & 7).
Table 6 The effect of different fatty acids (C8:0, C10:0 and C18:1) on bile acid metabolites in bile (mmol L−1, mean ± SD, n = 12)
| |
HF |
C8:0 |
C10:0 |
C18:1 |
| HF, high fat diets; C8:0, caprylic acid; C10:0, capric acids; C18:1, oleic acid; ap < 0.05, versus HF group. |
| Total BAs |
68.7 ± 22.2 |
93.7 ± 20.8a |
85.2 ± 10.7a |
90.0 ± 16.8a |
| CA |
58.2 ± 21.2 |
80.7 ± 21.3a |
71.6 ± 11.1a |
76.4 ± 16.2a |
| DCA |
0.62 ± 0.15 |
0.68 ± 0.16 |
0.66 ± 0.10 |
0.58 ± 0.13 |
| CDCA |
2.18 ± 0.73 |
2.80 ± 0.50a |
2.97 ± 0.47a |
2.89 ± 0.34a |
| UDCA |
0.27 ± 0.07 |
0.27 ± 0.05 |
0.28 ± 0.08 |
0.27 ± 0.04 |
| LCA |
2.11 ± 0.45 |
2.20 ± 0.68 |
2.13 ± 0.67 |
2.33 ± 0.26 |
| TCA |
2.23 ± 0.85 |
2.29 ± 1.02 |
2.26 ± 1.28 |
2.21 ± 0.70 |
| TCDCA |
1.42 ± 0.70 |
2.46 ± 0.67a |
2.45 ± 0.38a |
2.56 ± 0.26a |
| GCA |
1.67 ± 0.67 |
1.74 ± 1.42 |
1.64 ± 1.61 |
1.63 ± 0.72 |
| GCDCA |
0.29 ± 0.19 |
0.54 ± 0.23a |
0.57 ± 0.20a |
0.56 ± 0.13a |
| Total |
6.09 ± 1.11 |
8.36 ± 1.56a |
7.25 ± 1.96a |
8.35 ± 1.12a |
| Cholesterol |
2.99 ± 1.03 |
4.82 ± 1.49a |
4.04 ± 1.21a |
4.84 ± 0.60a |
| Cholestanol |
1.12 ± 0.44 |
1.41 ± 0.22 |
1.32 ± 0.15 |
1.41 ± 0.18 |
| Coprostanol |
1.67 ± 0.67 |
2.04 ± 1.42 |
1.94 ± 1.61 |
2.13 ± 0.72 |
Table 7 The effect of different fatty acids (C8:0, C10:0 and C18:1) on bile acid metabolites in feces (mg per 3 d, mean ± SD, n = 6)
| |
HF |
C8:0 |
C10:0 |
C18:1 |
| HF, high fat diets; C8:0, caprylic acid; C10:0, capric acids; C18:1, oleic acid; ap < 0.05, versus HF group. bp < 0.05, versus C18:1 group. |
| Total BAs |
96.25 ± 32.11 |
176.37 ± 29.22ab |
151.98 ± 25.1a |
122.56 ± 18.92 |
| CA |
59.55 ± 14.03 |
125.96 ± 8.65ab |
93.90 ± 17.06a |
70.83 ± 5.91 |
| DCA |
8.67 ± 1.75 |
9.29 ± 0.65 |
9.80 ± 1.36 |
9.48 ± 0.77 |
| CDCA |
6.50 ± 0.69 |
14.40 ± 0.75a |
12.66 ± 0.89a |
11.54 ± 0.76a |
| UDCA |
3.83 ± 0.14 |
3.85 ± 1.11 |
3.80 ± 0.21 |
3.85 ± 1.27 |
| LCA |
4.51 ± 0.34 |
13.33 ± 0.51ab |
12.47 ± 0.40a |
9.34 ± 0.45a |
| TCA |
2.37 ± 1.3 |
2.44 ± 1.27 |
2.79 ± 1.8 |
2.48 ± 1.53 |
| TCDCA |
3.11 ± 0.32 |
5.77 ± 0.38a |
5.63 ± 0.34a |
5.91 ± 0.46a |
| GCA |
1.34 ± 0.56 |
1.99 ± 0.45a |
1.33 ± 0.38 |
1.42 ± 0.18 |
| GCDCA |
4.14 ± 2.5 |
8.99 ± 1.7a |
8.64 ± 1.8a |
7.44 ± 1.2 |
| Total |
215.6 ± 37.2 |
328.6 ± 20.17a |
305.4 ± 34.11a |
238.7 ± 26.11 |
| Cholesterol |
204.8 ± 26.23 |
316.2 ± 19.78a |
292.4 ± 14.98a |
225.8 ± 5.77 |
| Cholestanol |
8.55 ± 1.11 |
9.34 ± 0.5 |
9.90 ± 1.05 |
9.77 ± 1.2 |
| Coprostanol |
1.71 ± 0.14 |
2.11 ± 0.02 |
2.15 ± 0.13 |
2.19 ± 0.14 |
Discussion
It is generally accepted that a high-fat diet (HF) can induce obesity in mice.15 In the present study, we observed that compared with the control group, MCFA present in the diet can slow down the increase of body weight, and decrease the liver weight, the weight of mesenteric white adipose tissue, epididymal fat and body fat percentage. Thus it can be seen that MCFA can decrease the production of fat in HF mice. What is the effect of MCFA on the serum prolife? We investigated the plasmatic levels of TG, TC, LDL-C, HDL-C and bile acids to test the effects of MCFA on lipid metabolism. As predicted, when MCFA was present in the diet, the increases in serum TC, LDL-C and TG which were the main risk factors leading to atherosclerosis16,17 were significantly smaller than those observed in the control group. However there is only TC decrease in serum in the oleic acid treated group compared with the control group. As we known, HDL-C is regarded as “good” cholesterol, while LDL-C as “bad” cholesterol.18,19 In our study, MCFA and oleic acid could preserve HDL-C to a higher level and the ratio of HDL-C/LDL-C was significantly higher. MCFA not only decreases serum TC and increases serum HDL-C/LDL-C, which is similar to oleic acid, but also decreases serum LDL-C and TG. It can be concluded that caprylic acid or capric acid is the best in decreasing TC, LDL-C and TG and increasing HDL-C/LDL-C. Therefore, considering the benefits of MCFA in lipid metabolism, MCFA should be further investigated as a protective agent for the cardiovascular system. In addition, serum TC can be regulated by the liver, small intestine and various genes, at the same time, the level of serum lipids had a dynamic change, and it might not fully reflect the respective gene expression level. Therefore the concentrations of serum TC were similar in the C8:0, C10:0 and C18:1 groups, although some genes were related with bile acid synthesis, excretion and uptake had differences (Fig. 1 & 2).
Cholesterol homeostasis in mammals is maintained through integrated regulation of three major metabolic pathways in the liver which include an endogenous biosynthetic pathway, exogenous intake pathway and excretion of bile.20 Removal of the excess of cholesterol from the body is essential for the maintenance of homeostasis. ATP-binding cassette sub-family G member 5 (ABCG5) and member 8 (ABCG8) are the members of the superfamily of ATP-binding cassette (ABC) transporters. They form a sterol transporter that acts in the liver and small intestine to prevent accumulation of dietary sterols, and intestinal ABCG5/G8 contributes to the extrahepatic cholesterol efflux in mice.21,22 In this study we not only observed that compared with the control group, MCFA increased the expressions of ABCG5 or ABCG8 transporters in the liver or small intestine of obese mice fed with a high fat diet, but also reported that the concentrations of total BAs and cholesterol in bile and feces in the MCFA group increased significantly compared with that of the control group. This result indicated that MCFA stimulates cholesterol efflux in mice, and up-regulation of ABCG5 or ABCG8 transporters was the likely mechanism underlying the decreased concentration of serum cholesterol.
Cytochrome P450 7A1 (CYP7A1) and P450 27A1 (CYP27A1) are rate-limiting enzymes involved in the classic pathway and the alternative pathway of bile acid biosynthesis. Chenodeoxycholic acid (CDCA), a major component of bile acids, represses CYP7A1 by activating a transcriptional repressor named the small heterodimer partner (SHP).23 In this study we observed that compared with the control group, MCFA decreased the expressions of SHP in the liver of obese mice fed with a high fat diet, however increased the expressions of CYP7A1 and CYP27A1. This result indicated that the MCFA enhance the synthesis of bile acid from cholesterol in the liver. From Table 6, we can see that the level of total BAs, including free bile acid and binging bile acid, in bile in MCFAs were significantly higher than that in the control group. It can be concluded that MCFA can lower serum cholesterol by decreasing the expressions of SHP and increasing the expressions of CYP7A1 and CYP27A1, as a result, promoting the synthesis of bile acid in the liver of mice.
The liver X receptor (LXR) is one of the key regulators of genes involved in cholesterol homeostasis. LXR agonists can increase cholesterol excretion into the bile by upregulation of ABCG5 and ABCG8 and reduce small intestinal cholesterol absorption.24–26 In our study, we investigated significant reductions of serum cholesterol and increase of BA synthesis and excretion in the MCFA group, but we didn't find serum BA elevation in the MCFA group as compared with the control group. This suggested that absorption of BAs was not increased. Moreover, the concentrations of BAs in bile and feces were both increasing; it also indicated that the absorption of BAs was reduced. It might be thought that MCFAs increase the expression of ABCG8 or ABCG5 relevant to the expression of LXR in the liver or in the small intestine, and so decrease the absorption of BAs in the intestine. Similar results have been reported in other studies of fish oil.27,28 However, the reason that the expression of ABCG5 in the liver was the same among every group, it needed further investigation. Thereby, it might be another mechanism to reduce blood cholesterol through MCFAs reducing the intestinal cholesterol or BA absorption.
In summary, in present study, our findings reveal that MCFA increases the expression of LXR and ABCG8 and enhances CYP7A1 and CYP27A1 activity by decreasing SHP expression in the liver, thereby promoting bile acid synthesis and excretion. The level of BAs and cholesterol in bile and feces increased, but serum BA was normal in the MCFA group, so we considered that MCFA increases the expression of ABCG5 and LXR in the small intestine, thereby inhibiting small intestinal bile acid absorption. As a result, we considered that MCFA reduced serum cholesterol through promoting the synthesis and excretion of liver bile acid, inhibiting the absorption of small intestine bile acid, and increasing the concentration of cholesterol and bile acid in bile and feces. These processes are illustrated in Fig. 3.
 |
| | Fig. 3 Outline of the effects of MCFA on the main metabolic pathway of bile acid. The arrows (↑) or (↓) represent an up or down-regulation of protein or mRNA expression. MCFA increased the expression of LXR and ABCG8 and enhanced CYP7A1 and CYP27A1 expression, decreased SHP expression in liver, and MCFA increased the expression of ABCG5, ABCG8 and LXR in small intestine, thereby MCFA promoted liver bile acid synthesis and excretion and inhibited small intestinal bile acid absorption. It can be concluded that liver bile acid excretion may be the primary pathway. | |
Acknowledgements
This study was partially supported by the Natural Science Fund of China (no. 81202203, 81541067) and the Youth Innovation Fund of the Chinese PLA General Hospital (no. 15KMM45).
References
- A. Aggarwal, S. Aggarwal, A. Goel, V. Sharma and S. Dwivedi, JRSM Cardiovasc. Dis., 2012, 1, 8 CrossRef PubMed . [PubMed: 24175065].
- M. J. Pletcher, E. Vittinghoff, A. Thanataveerat, K. Bibbins-Domingo and A. E. Moran, PLoS One, 2016, 11, e154288 Search PubMed , [PubMed: 27138014].
- J. Frohlich and A. Al-Sarraf, Cardiovasc. Pathol., 2013, 22, 16–18 CrossRef PubMed . [PubMed: 22502868].
- N. Tada, C. Maruyama, S. Koba, H. Tanaka, S. Birou, T. Teramoto and J. Sasaki, Atheroscler. Thromb., 2011, 18, 723–734 CrossRef . [PubMed: 21685707].
- D. Kritchevsky, H. K. Kim and S. A. Tepper, Atherosclerosis, 1972, 15, 101–105 CrossRef CAS PubMed . [PubMed: 5013274].
- E. M. Ooi, G. F. Watts, T. W. Ng and P. H. Barrett, Nutrients, 2015, 7, 4416–4425 CrossRef CAS PubMed . [PubMed: 26043038].
- Q. Xu, C. Xue, Y. Zhang, Y. Liu, J. Wang, X. Yu, X. Zhang, R. Zhang, X. Yang and C. Guo, Biosci., Biotechnol., Biochem., 2013, 77, 1390–1396 CrossRef CAS PubMed . [PubMed: 23832358].
- Y. Liu, C. Xue, Y. Zhang, Q. Xu, X. Yu, X. Zhang, J. Wang, R. Zhang, X. Gong and C. Guo, Biosci., Biotechnol., Biochem., 2011, 75, 1939–1944 CrossRef CAS PubMed . [PubMed: 21979067].
- Y. H. Liu, Y. Zhang, Q. Xu, X. M. Yu, X. S. Zhang, J. Wang, C. Xue, X. Y. Yang, R. X. Zhang and C. Y. Xue, Biosci., Biotechnol., Biochem., 2012, 76, 1213–1218 CrossRef CAS PubMed . [PubMed: 22790949].
- Y. H. Tsai, S. Park, J. Kovacic and J. T. Snook, Lipids, 1999, 34, 895–905 CrossRef CAS PubMed . [PubMed: 10574653].
- B. Wang, J. Fu, L. Li, D. Gong, X. Wen, P. Yu and Z. Zeng, Int. J. Food. Sci. Nutr., 2016, 67, 288–297 CrossRef CAS PubMed . [PubMed: 26932533].
- D. B. van Schalkwijk, W. J. Pasman, H. F. Hendriks, E. R. Verheij, C. M. Rubingh, K. van Bochove, W. H. Vaes, M. Adiels, A. P. Freidig and A. A. de Graaf, PLoS One, 2014, 9, e100376 Search PubMed . [PubMed: 25049048].
- Y. Huanling, L. Yong, W. Junbo, Z. Liping and Y. Weixing, Br. J. Nutr., 2005, 94, 315–320 CrossRef PubMed . [PubMed: 16176600].
- H. Kikuchi-Hayakawa, N. Onodera, S. Matsubara, E. Yasuda, Y. Shimakawa and F. Ishikawa, Br. J. Nutr., 1998, 79, 97–105 CrossRef CAS PubMed . [PubMed: 9505807].
- U. J. Jung, Y. Y. Cho and M. S. Choi, Nutrients, 2016, 8, 305 CrossRef PubMed . [PubMed: 27213439].
- P. Libby, P. M. Ridker and G. K. Hansson, Nature, 2011, 473, 317–325 CrossRef CAS PubMed . [PubMed: 21593864].
- X. Ni and H. Wang, Am. J. Transl. Res., 2016, 8, 1073–1081 Search PubMed . [PubMed: 27158393].
- H. Bjorkbacka, Curr. Opin. Lipidol., 2014, 25, 319–320 CrossRef PubMed . [PubMed: 25003740].
- H. Lu and A. Daugherty, Curr. Opin. Lipidol., 2014, 25, 157–158 CrossRef CAS PubMed . [PubMed: 24622057].
-
K. R. Feingold and C. Grunfeld, Introduction to Lipids and Lipoproteins, 2015, [PubMed: 26247089] Search PubMed.
- J. Wang, M. A. Mitsche, D. Lutjohann, J. C. Cohen, X. S. Xie and H. H. Hobbs, J. Lipid Res., 2015, 56, 319–330 CrossRef CAS PubMed . [PubMed: 25378657].
- X. H. Yu, K. Qian, N. Jiang, X. L. Zheng, F. S. Cayabyab and C. K. Tang, Clin. Chim. Acta, 2014, 428, 82–88 CrossRef CAS PubMed . [PubMed: 24252657].
- Y. Moon, B. Park and H. Park, BMB Rep., 2016, 49, 173–178 CrossRef CAS PubMed . [PubMed: 26521940].
- A. Mohammadi and E. A. Oshaghi, J. Diabetes Metab. Disord., 2014, 13, 20 CrossRef PubMed . [PubMed: 24476027].
- T. Pfeifer, M. Buchebner, P. G. Chandak, J. Patankar, A. Kratzer, S. Obrowsky, G. N. Rechberger, R. S. Kadam, U. B. Kompella, G. M. Kostner, D. Kratky and S. Levak-Frank, Curr. Pharm. Biotechnol., 2011, 12, 285–292 CAS . [PubMed: 21190543].
- E. Calayir, T. M. Becker, A. Kratzer, B. Ebner, U. Panzenbock, J. Stefujl and G. M. Kostner, Curr. Pharm. Biotechnol., 2008, 9, 516–521 CAS . [PubMed: 19075690].
- T. Kamisako, Y. Tanaka, T. Ikeda, K. Yamamoto and H. Ogawa, Hepatol. Res., 2012, 42, 321–326 CrossRef CAS PubMed . [PubMed:22364600].
- A. M. Berard, M. F. Dumon and M. Darmon, FEBS Lett., 2004, 559, 125–128 CrossRef CAS PubMed . [PubMed: 14960319].
|
| This journal is © The Royal Society of Chemistry 2017 |
Click here to see how this site uses Cookies. View our privacy policy here.