
Preventive effects of zingerone on cardiac mitochondrial oxidative stress, calcium ion overload and adenosine triphosphate depletion in isoproterenol induced myocardial infarcted rats
Abstract
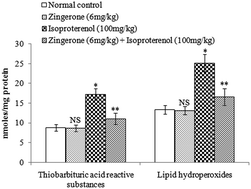
Cardiac mitochondrial oxidative stress, calcium ion (Ca2+) overload and adenosine triphosphate (ATP) depletion play an important role in the pathogenesis of myocardial infarction. The preventive effects of zingerone on oxidative stress, Ca2+ overload and ATP depletion in isoproterenol induced myocardial infarction were evaluated in rats. Rats were pretreated with zingerone (6 mg kg−1 body weight) daily for a period of 14 days. Isoproterenol (100 mg kg−1 body weight) was injected subcutaneously into rats twice at an interval of 24 h (on 15th and 16th day) to induce myocardial infarction. Isoproterenol induced myocardial infarcted rats showed a significant increase in the levels/concentrations of cardiac diagnostic marker, heart mitochondrial lipid peroxidation, Ca2+, and a significant decrease in the activities/concentrations of heart mitochondrial superoxide dismutase, glutathione peroxidase, reduced glutathione, tricarboxylic acid cycle and respiratory chain enzymes and ATP. Zingerone pretreatment showed significant preventive effects on all the biochemical parameters evaluated. Furthermore, the biochemical findings were correlated with transmission electron microscopic study on the structure of heart mitochondria. The in vitro study revealed the reactive oxygen species (ROS) scavenging effects of zingerone. Thus zingerone prevented mitochondrial damage by preventing cardiac mitochondrial oxidative stress, Ca2+ overload and ATP depletion in isoproterenol induced myocardial infarcted rats. The observed effects could be due to zingerone's anti-inotropic and ROS scavenging properties.
 Please wait while we load your content...
Please wait while we load your content...

