
Metabolic engineering of TiO2 nanoparticles in Nitzschia palea to form diatom nanotubes: an ingredient for solar cells to produce electricity and biofuel†

Abstract
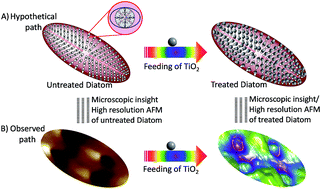
Diatoms are nature's nanobot because they can be described as cells in a glass house. Three-dimensional nanobio-engineered frustules of diatoms have vast applications in nanomaterials and bio-photonics. The amorphous silica of diatom frustules is not a good semiconductor, but when coated with titanium dioxide (TiO2), the semiconducting efficiency of the frustules becomes adequate for use in numerous device applications, including solar cells. The metabolic incorporation of titanium dioxide nanoparticles in the diatom frustules is primarily used for the construction of titanium nanotubes. In the present study, TiO2 was metabolically inserted by a two-stage cultivation process and its incorporation in the diatom frustules was studied by spectroscopic and atomic force microscopic methods. It was found that diatom frustules metabolically inserted with nanostructured TiO2 by a two-stage cultivation in f/2 medium could replace nanostructured surface doping with titanium in dye-sensitized solar cells for heat and electricity production. The DSSC made from the titanium-doped diatom frustule has a power efficiency almost double (9.45%) that of the simple DSSC (4.20%) without any diatom frustule metabolically inserted with TiO2. Alternatively, these nanoarray diatom containing tubes may also be used for the construction of novel dye-sensitized solar cells, which may help in oozing lipids from living diatom cells, such as in algal cells, for the generation of an electric current.
 Please wait while we load your content...
Please wait while we load your content...

