
Individual and combined antioxidant effects of ginsenoside F2 and cyanidin-3-O-glucoside in human embryonic kidney 293 cells†
Abstract
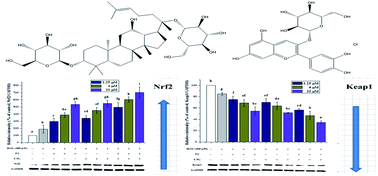
Ginsenosides and anthocyanins have already been reported to exhibit protective effects on intracellular defense against oxidative stress. Nuclear factor erythroid-derived 2 (NF-E2)-like 2 (Nrf2) plays a key role in signal pathways to counteract oxidative stress and controls expression of antioxidant enzymes. The present study is aimed at investigating the individual and combined antioxidant effects of ginsenoside F2 (F2) and cyanidin-3-O-glucoside (C3G) against H2O2-induced oxidative stress in Human Embryonic Kidney (HEK-293) cells. Cell viability, MDA level and activities of antioxidant enzymes were measured using corresponding assay kits. A DCFH-DA fluorescent probe assay was used to measure the level of intracellular reactive oxygen species (ROS). Quantitative real-time PCR and Western blotting were used to detect the expression of Nrf2 and Kelch-like ECH associated protein 1 (Keap1). The results showed that F2 and C3G significantly inhibited the generation of MDA and intracellular ROS, and increased the activities of antioxidant enzymes. Furthermore, F2 and C3G significantly enhanced the protein and mRNA expressions of Nrf2 and reduced the expressions of Keap1 in a concentration-dependent manner. On the basis of the stronger individual effects, synergistic effects were present in the combinations of F2 and C3G. In conclusion, these results demonstrated that F2 and C3G can protect HEK-293 cells against H2O2-induced oxidative stress and possibly act synergistically through reducing intracellular ROS, as well as activating the Nrf2/Keap1 signaling pathway and increasing antioxidant enzyme levels.
 Please wait while we load your content...
Please wait while we load your content...

