
Efficient production of citric acid in segmented fermentation using Aspergillus niger based on recycling of a pellet-dispersion strategy†
Baoshi Wangabc,
Jian Chena,
Hua Lia,
Fuxin Sunc,
Youran Liab and
Guiyang Shi*ab
aNational Engineering Laboratory for Cereal Fermentation Technology, Wuxi 214122, PR China
bSchool of Biotechology, Jiangnan University, Wuxi 214122, PR China. E-mail: gyshi@jiangnan.edu.cn; Fax: +86 1085918229; Tel: +86 1085918229
cJiangsu Guoxin Union Energy Co., Ltd., Wuxi 214203, PR China
First published on 19th October 2016
Abstract
Citric acid (CA), as the most important platform compound, has attracted much attention because of its broad applications in many areas. Here, we constructed a segmentation fermentation process which effectively separated cell growth and CA production. Through a pellet-dispersion strategy, the CA yield decline caused by the mycelium structure of Aspergillus niger was solved. In particular, CA production was significantly increased from 70.3 g L−1 to 97.0 g L−1. Furthermore, the segmentation fermentation conditions were optimized to improve the CA production. Evidence showed that the process was stable during ten batches of recycling fermentation in 5 L five-conjoined-fermenters, from which the CA yield even increased by 1.4 g L−1 (by 1.26%) compared with the control. All of these results confirm that the strategy could enhance productivity and avoid the traditional long process (about 30 days) of seed culturing (including spore preparation). It could be also applied in other fermentation industries involving filamentous fungi.
1. Introduction
Citric acid (2-hydroxy-1,2,3-propanetricarboxylic acid, CA), an intermediate of the tricarboxylic acid cycle, is a significantly important platform compound. With its unique attributes, CA has been widely used in food and beverages, and for chemical and pharmaceutical applications.1–4 Currently, the global demand for CA is continuously increasing at a high annual rate of 5%, along with new applications in advanced fields such as drug delivery5 and tissue engineering.6 To cover the ever-increasing demand, the development of efficient technologies is becoming more and more important.7,8Conventionally, CA is mainly produced by submerged fermentation with batch cultivation, using the filamentous fungus Aspergillus niger.9 However, this traditional cultivation mode has many disadvantages, such as a long auxiliary time, poor energy efficiency and low utilization rate of the equipment. A continuous cultivation mode, in comparison, could achieve a higher yield with less labor and energy consumption. Moreover, the continuous culture liquid is also favorable for the recovery of the products. To date, implementation of this culture mode is generally difficult due to the fact that CA synthesis is only partially growth-related. The special mycelium structure of A. niger influences the mass transfer of dissolved oxygen, a primary obstacle in maintaining normal cell metabolism.10 Even so, many experts have made great attempts to develop a continuous process for CA production through immobilization of the cells of A. niger based on control of the pellet size.11–15 There are some limitations to these immobilized viable cell systems, such as unwanted side-reactions and, most importantly, impaired mass transfer. In this case, it is still difficult to maintain the culture activity due to the absence of cell culture regeneration and the gradual aging of the culture. Yeast has also been used for the continuous production of CA in recent years.9,16–18 Arzumanov et al. designed a cultivation process using Yarrowia lipolytica VKM Y-2373, where the culture liquid was withdrawn from cells and then added to a fresh medium, which ensured that the culture activity could remain stable after a long period of time.10 Likewise, Rywińska et al. employed a fermentation process using a strain of Yarrowia lipolytica, Wratislavia AWG7, where a portion of the culture liquid was withdrawn and the same volume of production medium was added. When 40% of freshly replaced medium was added, the activity of the culture remained stable long-term.16 However, a considerable amount of isocitric acid (ICA) accumulated when yeast was used as the production strain. The presence of ICA made the isolation and purification process of CA more complicated, which was a great obstacle in terms of its application.17,19 Recently, although genetic engineering has made improvements in terms of reducing ICA formation, the instability of recombinant strains and concerns over food safety must be considered when CA is applied to food additives and in healthcare fields. In fact, in comparison with yeast, A. niger still remains the best candidate for the continuous production of CA due to its ease of handling, wide substrate spectrum, high yields at low pH and extremely low by-product secretion.20–22 Therefore, development in the continuous cultivation mode of yeast has highlighted the benefits of using A. niger in the field of CA production.
Filamentous fungi typically exhibits a complex morphological life cycle that is related to production performance in the submerged culture processes.23 The fungal forms can vary from dense spherical pellets to dispersed mycelium, and have been shown to be strongly influenced by process parameters, including power input through stirring and aeration. Belmar-Beiny et al. found a cycle of hypha fragmentation and regrowth at a higher stirred speed during clavulanic acid production with S. clavuligerus.24 A similar phenomenon could be observed during CA production using A. niger, during which the hypha fragments chipped off the pellet surface due to mechanical forces and a reseeded pellet population.25 Moreover, Lu et al. confirmed that the seed morphology with a dispersed pattern of A. niger AN151, induced by the higher agitation speed, achieved higher gluconic acid formation compared to the pelleted fermentations.26 Xin et al. developed a feasible method that utilized mycelium fragments to grow fungal pellets and this presented a good application for treated wastewater.27 All together, the performance of the morphological life cycle provided information on the control of the mycelium structure of A. niger in the continuous cultivation.
In the present study, we firstly propose a segmentation fermentation process to separate cell growth and CA production, laying a foundation for the continuous production of CA. Then, a pellet-dispersion strategy was introduced to avoid CA reduction caused by the mycelium formation of A. niger. Moreover the segmentation fermentation conditions were optimized. Finally, the CA fermentation process was reconstructed by combining segmentation fermentation with pellet-dispersion strategies and significant improvement was obtained in the productivity. The present work also provides useful information for the morphological control of fermentations involving filamentous fungus.
2. Methods
2.1 Microorganism
The strain of A. niger used in this study was an industrial strain that is used for CA production, which was obtained from Jiangsu Guoxin Union Energy Co., Ltd. Fresh spores were obtained after stepwise-expansion cultivation including plate selection and formation of a slant culture, eggplant-flask culture and moldy-bran bucket culture at 35 °C for 30 d (Fig. 1). The spore suspension was prepared by adding distilled water to Tween-80 (2%, v/v) and this was then used for inoculation. | ||
| Fig. 1 Schematic diagram of citric acid production via the two synthesis modes of traditional fermentation and constructed segmentation fermentation. The traditional fermentation process is a batch cultivation process as shown in the above dotted box, containing two stages of seed culture (including spore preparation in the form of stepwise-expansion cultivation) and fermentation culture. The segmentation fermentation was a recycling-batch process as shown in the below dotted box. A portion of the fermentation broth was segmented out to a new fermenter where the fermentations were cultured for a certain stage. The segmented mycelium pellet was dispersed into the dispersed mycelium by the disperser and then fresh medium was added into the fermenter. The dispersed mycelium can also further develop into a mycelium pellet when cultured for a period of time. By repeating the process above, a novel segmentation fermentation process is constructed (F1–F10 represent the ten recycling fermentation batches). | ||
2.2 Media and cultivation conditions
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) :3 (w/w) and then the slurry pH was adjusted to 5.8–6.0 with calcium hydroxide. Thermostable α-amylase (40000 U mL−1, optimal temperature range from 95–105 °C, Novozymes) was added to the slurry at 25 U per gram of cassava power, followed by heating of the slurry at 100 °C and holding it for 2 h.28 The water loss during the liquefaction process was firstly supplemented. Then the liquefaction slurry was filtered through a Buchner funnel and the liquefaction filtrate was collected. The filtrate was diluted to the required substrate concentration for the preparation of the seed medium and fermentation medium.
:3 (w/w) and then the slurry pH was adjusted to 5.8–6.0 with calcium hydroxide. Thermostable α-amylase (40000 U mL−1, optimal temperature range from 95–105 °C, Novozymes) was added to the slurry at 25 U per gram of cassava power, followed by heating of the slurry at 100 °C and holding it for 2 h.28 The water loss during the liquefaction process was firstly supplemented. Then the liquefaction slurry was filtered through a Buchner funnel and the liquefaction filtrate was collected. The filtrate was diluted to the required substrate concentration for the preparation of the seed medium and fermentation medium.2.3 Image analysis
The fermentation broth was directly diluted with saline (0.9% NaCl, w/w) to a certain amount of the mycelium pellets’ concentration. Mycelia images were captured by a CCD camera (Leica DFC 450) that was installed on a microscope (Leica DM 1000) using a 10× objective. Morphological measurements were conducted using Leica application Suite V4 software (Leica microsystems, Wetzlar, Germany). The pellet quantity was counted under the microscope (Leica DM 1000) using a 10× objective with the concentration diluted to about 102 to 103 pellets per milliliter.292.4 Analytical methods
In order to analyze the residual total sugar and CA yield, the fermentation culture was centrifuged at 10000 × g and 4 °C for 10 min, and the clear supernatant was filtered through a 0.22 μm syringe filter for analysis. The residual total sugar was determined using a biosensor (SBA-40B, Shandong Academy of Sciences, China) after the samples were subjected to acid hydrolysis (200 g L−1 sulfuric acid, 100 °C, 20 min).30 The CA yield was determined by high-performance liquid chromatography using an SH1011 column (Agilent, USA) and was subsequently detected at 210 nm with a UV detector (UVD 170U, DIONEX Co., Ltd.) with a mobile phase of 0.01 M H2SO4 at 50 °C. The cell pellet was washed with distilled water and filtered through filter paper, then dried at 105 °C in the oven until it reached a constant weight for biomass analysis.
3. Results and discussion
3.1 Segmentation recycling fermentation with the mycelium pellet for CA production
CA production using traditional batch fermentation was performed in a 5 L fermenter. As expected, the CA fermentation process contained two stages of cell growth and product synthesis (ESI Fig. 1†). In the early stages of fermentation (0–24 h), cell growth was rapidly increased along with a little CA accumulation. After 24 h, the amount of CA increased with a higher synthesis rate, though biomass consumption evidently slowed down and approached a steady-state. The total sugar concentration decreased gradually throughout the CA fermentation process. The reducing sugar presented a trend where the sugar concentration was first increased and then decreased, since a high amount of glucoamylase was secreted in the early stages, consistent with cell growth. Collectively, the CA synthesis mode is partially growth-related.It has previously been shown that separating growth and synthesis is necessary in time or in space for the continuous production of CA by A. niger, since CA biosynthesis is indirectly growth-associated.31 To this end, we constructed a novel process, where a portion (2/10, v/v) of the fermentation culture was segmented into another fermenter containing fresh medium at fixed periods of time. Subsequently, six-recycling batches were conducted in the same manner. The fermentation results are shown in Fig. 2. Interestingly, CA accumulation was significantly decreased from 112.3 g L−1 to 70.3 g L−1 with an increase in the number of segmentation batches (Fig. 2a). In particular, the CA-production was clearly reduced by 37.4% in the sixth batch. The biomass almost appeared with an analogous regularity, dropping from an initial concentration of 23.4 g L−1 to 14.9 g L−1 by the sixth batch, and decreased by 36.3%. As expected, the recycling batches with decreasing biomass displayed lower productivity. Additionally, the morphology of filamentous organisms in submerged cultivation is critically important since it can significantly influence the productivity. Therefore, the morphological features including mean diameter, pellet quantity and mycelium pellet structure in the six-recycling batches were further investigated (Fig. 2c and d). Surprisingly, the mean diameter of the mycelium pellets was significantly increased from an initial size of 150.6 μm to 476.7 μm as the number of recycling batches increased, along with a remarkable reduction in the pellet quantity from 3.7 × 104 to 1.4 × 104 pellets per milliliter (Fig. 2c). Moreover, the hypha in the mycelium pellet were densely intertwined and the compactness of the pellets also increased with an increase in the number of recycling batches, with the pellet diameter increasing distinctly (Fig. 2d). As is well known, mycelium pellets with a larger size and denser layer limit the supply of oxygen and nutrients to the pellet cores, resulting in “dead zones” with greatly reduced productivity.32 Taken together, the sharp decrease in CA production and biomass with an increase in the number of recycling-batches should be ascribed to the dramatic changes in the morphological characteristics. These results suggest that control of the strain morphology during the segmentation fermentation should be an important target to increase the CA yield.
 | ||
| Fig. 2 (a) CA production, (b) biomass concentration, (c) mean diameter and quantity of mycelium pellets, and (d) mycelium pellet morphological changes during segmentation fermentation recycling (F1–F6 represent the six recycling fermentation batches). Data are means ± SD (n = 3). | ||
3.2 Segmentation recycling fermentation with the dispersed mycelium for CA production
Although the cell growth and CA synthesis were successfully separated via the constructed segmentation fermentation process, a sharp increase in the pellet diameter and hypha compactness with an increase in the number of the recycling batches severely hindered the cell growth and CA accumulation. The filamentous morphology has been shown to be strongly influenced by the shearing action that is caused by mechanical agitation and aeration. Interestingly, it could be found that the hypha fragmentation chipped off the surface of the pellet, due to mechanical forces, and the reseeded pellet population.24–26 In consideration of the strain morphology characteristics upon exposure to the mechanical environment, we dispersed the pellet using the disperser to precisely control the morphology during segmentation fermentation. Similarly to the above segmentation fermentation process, a portion (2/10, v/v) of fermentation broth was segmented out into a new fermenter, and the described culture process was continuously recycled for six batches of fermentations (Fig. 1). Notably, the dispersed mycelium in the recycling segmentation could also form a pellet (Fig. 3d). Surprisingly, the mean diameter almost remained approximately at 164.8 μm on average and the pellet quantity remarkably increased from initially 3.7 × 104 to 5.3 × 104 per milliliter (Fig. 3c). The pellet structure was obviously improved with hypha that were less-loosely intertwined in the pellet core (Fig. 3d), which was favorable for the oxygen and nutrition transfer, facilitating CA accumulation. As expected, CA production in this segmentation fermentation with recycled dispersed mycelium was almost completely restored and stabilized at 94.4 g L−1 in the recycling batches (Fig. 2a). Additionally, the biomass was also slightly enhanced compared to the mycelium pellet recycling fermentations. These results demonstrate that control of the pellet morphology in the segmentation fermentation, using a pellet-dispersion strategy, is a feasible and effective way to improve the CA production. | ||
| Fig. 3 (a) CA production, (b) biomass concentration, (c) mean diameter and quantity of mycelium pellets, and (d) mycelium pellet morphological changes during segmentation fermentation recycling with mycelium pellets (DF1–DF6 represent the six recycling fermentation batches). Data are means ± SD (n = 3). | ||
3.3 Influence of segmentation fermentation conditions on CA production
Although the pellet-dispersion strategy successfully improved the CA production and biomass, the results were still lower than those in the initial control. So ensuring that adequate biomass is retained in the fermenter at all times is crucial for CA accumulation in the continuous culture process.33 To strengthen the fermentation processes, the segmentation conditions such as segmentation level, soybean addition and segmentation time were further investigated in the shake-flask. To identify the influence of the segmentation proportion on fermentation, segmentation levels (1/10, 2/10, 2/10, 4/10 and 5/10; v/v) under the conditions of soybean addition of 15 g L−1 and a segmentation time of 24 h were analyzed in a shaken-flask experiment. As shown in Fig. 4a and b, CA titer and biomass were 5.3% and 11.1% lower than those of the control when the segmentation level was 1/10 (v/v), whereas CA production and biomass approached control levels with an increase in the segmentation level from 2/10 to 5/10 (v/v). This revealed that the segmentation of more fermentation broth could lower the pH of the initial fermentation medium, which was favorable for CA production. These results, which are perfectly in accordance with previous reports, show that a lower pH in the initial medium could accelerate the accumulation of CA.34 Therefore, it should be noted that the segmentation level should be maintained at a moderate level to ensure adequate biomass for fermentation. In the following experiment, we selected the segmentation level 2/10 (v/v) to be investigated.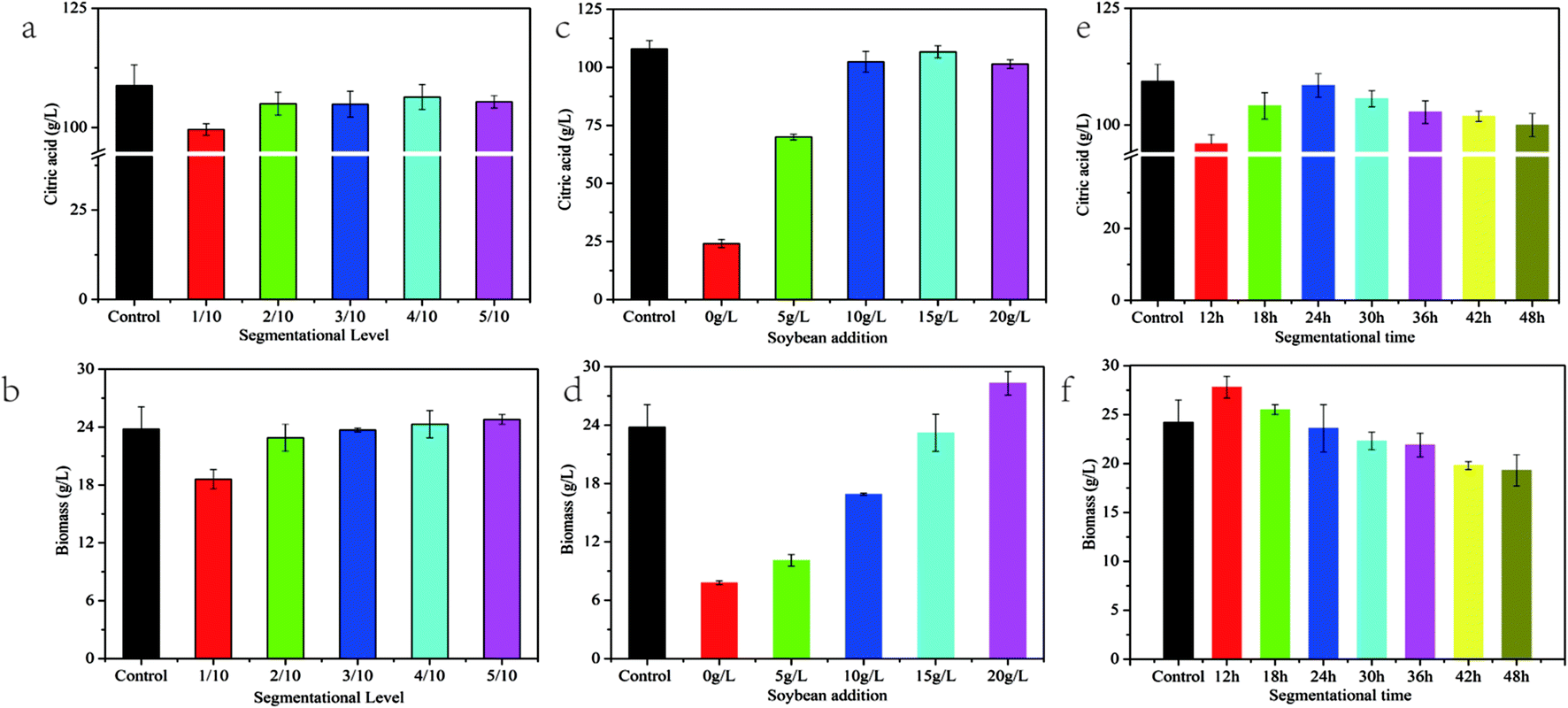 | ||
| Fig. 4 Influence of segmentation fermentation conditions including segmentation level ((a) and (b) soybean addition 15 g L−1; segmentation time 24 h), soybean addition ((c) and (d) segmentation level 2/10, v/v; segmentation time 24 h) and segmentation time ((e) and (f) segmentation level 2/10, v/v; soybean addition 15 g L−1) on the biomass and CA production. Data are means ± SD (n = 3). | ||
It has been shown that excess production of CA only occurs under nitrogen-limited conditions while biomass growth and reproduction are suppressed under these conditions.34 The accumulation of CA was strongly influenced by the composition of the fermentation medium, especially in the submerged fermentation. To determine a moderate concentration of the nitrogen source, we investigated the effects of adding different amounts of soybean, as the sole nitrogen source, on CA production under the conditions of a segmentation level of 2/10 (v/v) and segmentation time of 24 h (Fig. 4c and d). CA concentration presented an increasing trend with the addition of soybean from 5 g L−1 to 15 g L−1 and approached the control results at a level of 15 g L−1 while it became reduced at a level of 20 g L−1. However, biomass continuously increased with soybean addition (from 5 g L−1 to 20 g L−1). For CA production, when the strain grew under the conditions of a low nitrogen source supply, the nitrogen source could not meet the demand of the strain and resulted in a reduction of metabolic activity, giving a lower yield. However, an excessive nitrogen supply could cause a decrease in productivity because of the resulting losses of substrate due to overgrowth during the fermentation process.35 Therefore, moderate control of a nitrogen-limited supply can balance the competition between cell growth and CA production. In this study, soybean addition at 15 g L−1 was evidently favorable to the segmentation fermentation.
Throughout the CA production process, the cell generally presented different viabilities during different stages.10 Thus, selection of an appropriate phase to initiate the segmentation fermentation is of great importance. Different segmentation times were investigated under the conditions of a segmentation level of 2/10 (v/v) and soybean addition of 15 g L−1. The results showed that the segmentation fermentation started at an earlier stage when there was more biomass and less CA compared to the control while having less biomass and CA made the segmentation fermentation start at a later stage (Fig. 4e and f). The CA production attained the highest results, close to the control, at 24 h as the biomass was lying in an active-metabolism stage.
3.4 Optimization of CA production by segmentation fermentation coupled with the pellet-dispersion strategy
For the effective production of CA, segmentation fermentation processes coupled with the pellet-dispersion strategy were performed in a 5 L five-conjoined-fermenter under the optimized segmentation conditions (segmentation level 2/10, v/v; soybean addition 15 g L−1; segmentation time 24 h) (Fig. 1). Surprisingly, the final CA production in the tenth-batch approached up to 115.1 g L−1 after a fermentation time of 60 h, which was even higher than that of the initial control (111.3 g L−1, Fig. 5a). However, CA yield did have a slight decline in the initial batches, during which the dispersed mycelium might have a slightly adaptive process in the nitrogen-limited medium. Overall, the CA production stabilized the whole recycling batches (112.7 g L−1) on average, even increasing 1.4 g L−1 (by 1.26%) compared with the control. In comparison, the total sugar consumption in the recycling batches (2nd to 10th) significantly decreased (by 10.01%) since more nutrients were used to synthesize the product (Fig. 5b). Meanwhile, the volumetric CA productivity and specific rate of CA production of the fermentations throughout the whole recycling batches (2nd to 10th) were stable and even slightly higher than the control (Table 1), demonstrating that the cells could retain high viability even in the tenth batch. Furthermore, the yield of CA was remarkably improved from the initial batch, 0.83 g g−1 (the control), to the tenth batch, 0.96 g g−1, with a 15.7% increase (Table 1). Consequently, much more CA can be produced in the optimized fermentation process under consumption of the equivalent amount of substrate compared to the traditional fermentation process, which greatly reduces production costs and could have extensive prospects for industrial applications. Taken together, these results demonstrate that it is feasible and efficient to produce CA by segmentation fermentation coupled with the pellet-dispersion strategy.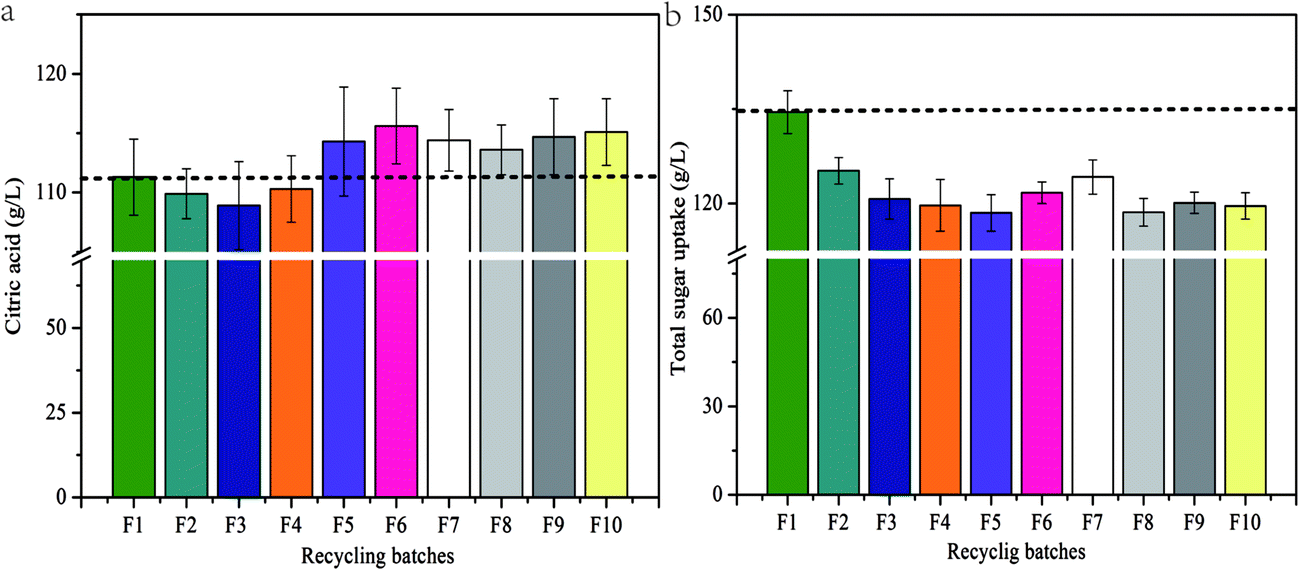 | ||
| Fig. 5 CA production (a) and total sugar uptake (b) in the ten-recycling batches (F1–F10 represent the ten recycling fermentation batches). Data are means ± SD (n = 3). | ||
| Batches | QCAb (g L−1 h−2) | qCAc (10 × g g−1 h−2) | YCAd (g g−1) |
|---|---|---|---|
| a (F1–F10 represent the ten recycling fermentation batches).b QCA = volumetric citric acid productivity.c qCA = specific rate of citric acid production.d YCA (g g−1) = yield of citric acid (g citric acid per total sugar).e Data are means ± SD (n = 3). | |||
| F1 | 1.86 ± 0.31 | 0.76 ± 0.03 | 0.83 ± 0.07 |
| F2 | 1.83 ± 0.24 | 0.83 ± 0.04 | 0.88 ± 0.12 |
| F3 | 1.82 ± 0.42 | 0.83 ± 0.02 | 0.90 ± 0.09 |
| F4 | 1.84 ± 0.34 | 0.75 ± 0.04 | 0.92 ± 0.07 |
| F5 | 1.91 ± 0.29 | 0.94 ± 0.03 | 0.96 ± 0.18 |
| F6 | 1.93 ± 0.16 | 0.90 ± 0.01 | 0.95 ± 0.07 |
| F7 | 1.91 ± 0.32 | 0.78 ± 0.02 | 0.92 ± 0.13 |
| F8 | 1.89 ± 0.26 | 0.82 ± 0.03 | 0.96 ± 0.20 |
| F9 | 1.91 ± 0.09 | 0.80 ± 0.05 | 0.96 ± 0.13 |
| F10 | 1.92 ± 0.11 | 0.79 ± 0.04 | 0.96 ± 0.08 |
A scale-up simulation experiment was conducted in a 30 L fermenter. Interestingly, CA production in the tenth-batch still approached up to 117.1 g L−1 after 60 h, 2.6 g L−1 higher than that in the first batch (114. 5 g L−1). Overall, CA production in the recycling batches (2nd to 10th) was 116.1 g L−1 on average with a 1.57% increase (ESI Fig. 2†), which was even better than those in the 5 L fermenter. Meanwhile, the total sugar consumption in the recycling batches significantly decreased since more nutrients were used for the CA synthesis, consistent with the results in the 5 L experiments (Fig. 2b). In view of the process parameters in the recycling batches, the productivity and production rate of CA and the yield were all stable and improved compared with the first batch (ESI Table 1†). All together, cells could retain high viability even in the tenth batch and this demonstrated that the segmentation fermentation coupled with the pellet-dispersion strategy was feasible.
4. Conclusions
In this study, we constructed a segmentation fermentation process to separate cell growth and CA production. Against this background, the CA yield was significantly increased from 70.3 g L−1 to 97.0 g L−1 via a pellet-dispersion strategy. Moreover, CA production in a 5 L five-conjoined-fermenter stabilized during the ten recycling batches, even increasing to 1.4 g L−1 (by 1.26%) compared with the control. Collectively, these results demonstrate that it is feasible and efficient to produce CA by segmentation fermentation coupled with the pellet-dispersion strategy. This work also serves as a promising stepping-stone for other fermentations involving filamentous fungi.Acknowledgements
This research was financially supported by the National High Technology Research and Development Program of China (863 Program, 2015AA020302) and the Cooperation Project of Jiangsu Province among Industries, Universities and Institutes (BY2015019-13) and the Jiangsu Guoxin-Union Energy Co. Ltd., China. We are thankful for their support.References
- A. R. Angumeenal and D. Venkappayya, LWT--Food Sci. Technol., 2013, 50, 367–370 CrossRef CAS.
- G. Singh Dhillon, S. Kaur Brar, M. Verma and R. D. Tyagi, Food Bioprocess Technol., 2011, 4, 505–529 CrossRef.
- L. Karaffa and C. P. Kubicek, Appl. Microbiol. Biotechnol., 2003, 61, 189–196 CrossRef CAS PubMed.
- H. Ikram-ul, S. Ali, M. A. Qadeer and J. Iqbal, Bioresour. Technol., 2004, 93, 125–130 CrossRef CAS PubMed.
- A. T. Naeini, M. Adeli and M. Vossoughi, Nanomedicine: Nanotechnology, Biology and Medicine, 2010, 6, 556–562 CrossRef CAS PubMed.
- A. Guillermo, Y. Jian and H. Ryan, US Pat. Application, 20090325859, 2010.
- J. N. W. A. de Jongh, Metab. Eng., 2008, 10, 87–96 CrossRef PubMed.
- I. Weyda, M. Lübeck, B. K. Ahring and P. S. Lübeck, J. Ind. Microbiol. Biotechnol., 2014, 41, 733–739 CrossRef CAS PubMed.
- A. Förster, A. Aurich, S. Mauersberger and G. Barth, Appl. Microbiol. Biotechnol., 2007, 75, 1409–1417 CrossRef PubMed.
- T. Arzumanov, N. Shishkanova and T. Finogenova, Appl. Microbiol. Biotechnol., 2000, 53, 525–529 CrossRef CAS PubMed.
- S.-K. Kim, P.-J. Park and H.-G. Byun, Biotechnol. Bioprocess Eng., 2002, 7, 89–94 CrossRef.
- A. P. J. N. V. Sankpal and B. D. Kulkarni, Process Biochem., 2001, 36, 1129–1139 CrossRef.
- K. Garg and C. B. Sharma, J. Gen. Appl. Microbiol., 1992, 38, 605–615 CrossRef CAS.
- H. Eikmeier and H. J. Rehm, Z. Naturforsch., C: Biosci., 1987, 42, 408–413 CAS.
- J. Vaija and P. Linko, J. Mol. Catal., 1986, 237–253 CrossRef CAS.
- A. Rywińska and W. Rymowicz, J. Ind. Microbiol. Biotechnol., 2010, 37, 431–435 CrossRef PubMed.
- A. Rywińska, P. Juszczyk, M. Wojtatowicz and W. Rymowicz, J. Biotechnol., 2011, 152, 54–57 CrossRef PubMed.
- L. Moeller, M. Grünberg, A. Zehnsdorf, A. Aurich, T. Bley and B. Strehlitz, J. Biotechnol., 2011, 153, 133–137 CrossRef CAS PubMed.
- S. Anastassiadis, I. G. Morgunov, S. V. Kamzolova and T. V. Finogenova, Recent Pat. Biotechnol., 2008, 2, 107–123 CrossRef CAS PubMed.
- H. Grewal and K. Kalra, Biotechnol. Adv., 1995, 13, 209–234 CrossRef CAS PubMed.
- E. Schuster, N. Dunn-Coleman, J. C. Frisvad and P. W. M. van Dijck, Appl. Microbiol. Biotechnol., 2002, 59, 426–435 CrossRef CAS PubMed.
- F. Vendruscolo, P. M. Albuquerque, F. Streit, E. Esposito and J. L. Ninow, Crit. Rev. Biotechnol., 2008, 28, 1–12 CrossRef CAS PubMed.
- R. Krull, T. Wucherpfennig, M. E. Esfandabadi, R. Walisko, G. Melzer, D. C. Hempel, I. Kampen, A. Kwade and C. Wittmann, J. Biotechnol., 2013, 163, 112–123 CrossRef CAS PubMed.
- M. T. Belmar-Beiny and C. R. Thomas, Biotechnol. Bioeng., 1991, 37, 456–462 CrossRef CAS PubMed.
- G. C. Paul, M. A. Priede and C. R. Thomas, Biochem. Eng. J., 1999, 3, 121–129 CrossRef CAS.
- F. Lu, K. Ping, L. Wen, W. Zhao, Z. Wang, J. Chu and Y. Zhuang, Process Biochem., 2015, 50, 1342–1348 CrossRef CAS.
- B. Xin, Y. Xia, Y. Zhang, H. Aslam, C. Liu and S. Chen, Bioresour. Technol., 2012, 105, 100–105 CrossRef CAS PubMed.
- Y. Deng, S. Li, Q. Xu, M. Gao and H. Huang, Bioresour. Technol., 2012, 107, 363–367 CrossRef CAS PubMed.
- X. D. Ren, Y.-J. Xu, X. Zeng, X.-S. Chen, L. Tang and Z.-G. Mao, RSC Adv., 2015, 82138–82143 RSC.
- J. Xu, X.-F. Su, J.-W. Bao, Y.-Q. Chen, H.-J. Zhang, L. Tang, K. Wang, J.-H. Zhang, X.-S. Chen and Z.-G. Mao, Bioresour. Technol., 2015, 189, 186–194 CrossRef CAS PubMed.
- G. J. L. Elmer, J. Biochem. Microbiol. Technol. Eng., 1959, 413–429 Search PubMed.
- H. Driouch, R. Hänsch, T. Wucherpfennig, R. Krull and C. Wittmann, Biotechnol. Bioeng., 2012, 109, 462–471 CrossRef CAS PubMed.
- K. D. Rane and K. A. Sims, Biotechnol. Bioeng., 1995, 46, 325–332 CrossRef CAS PubMed.
- M. Papagianni, Biotechnol. Adv., 2007, 25, 244–263 CrossRef CAS PubMed.
- M. Papagianni, F. Wayman and M. Mattey, Appl. Environ. Microbiol., 2005, 71, 7178–7186 CrossRef CAS PubMed.
Footnote |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c6ra13648f |
| This journal is © The Royal Society of Chemistry 2016 |