
Effects of cholic acid modified glucosamine on chondrogenic differentiation†
Abstract
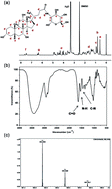
Glucosamine hydrochloride is a widely used drug for the treatment of osteoarthritis and can be easily modified by other molecules because of its alterable functional groups. Cholic acid is an amphiphilic molecule which may regulate properties of glucosamine once introduced into it. In this study, a model cell line ATDC5 was used to investigate the effect of cholic acid modified glucosamine on chondrogenic differentiation. The Alcian blue staining indicated that 1.6 mM of cholic acid modified glucosamine enhanced cartilage-relevant extracellular matrix deposition to the greatest extent. Besides, the results of real-time polymerase chain reaction, western blotting and immunofluorescence staining demonstrated that the expression of cartilage-relevant genes and proteins was up-regulated most and the fibrous cartilage- and hypertrophic cartilage-relevant genes and proteins were down-regulated. Overall, these results demonstrated that 1.6 mM cholic acid modified glucosamine exhibited the best effect on promoting chondrogenic differentiation, which not only enhanced chondrogenesis but also inhibited fibrous cartilage and hypertrophic cartilage formation.
 Please wait while we load your content...
Please wait while we load your content...

