DOI:
10.1039/C6RA08839B
(Paper)
RSC Adv., 2016,
6, 82862-82871
Aerobic degradation of estrogenic alkylphenols by yeasts isolated from a sewage treatment plant
Received
6th April 2016
, Accepted 24th August 2016
First published on 26th August 2016
Abstract
Long-chain alkylphenols including octylphenol (OP) are well-known toxic pollutants prevailing in the environment due to the massive demand of these chemicals in industry and have been identified as endocrine disrupting chemicals (EDCs). While a considerable amount of work has addressed the degradation capacity of EDCs carried out by bacterial and filamentous fungal strains, little is known about the EDC degradation by yeasts. In this study, we isolated three branched-chain OP-degrading yeast strains from samples collected from a wastewater treatment plant in Taipei, Taiwan, by enrichment methods using 4-t-octylphenol (4-t-OP) as the sole carbon and energy source. Analysis of ITS and LSU rDNA gene sequences revealed that these isolates belonged to two genera and were designated as Candida rugopelliculosa RRKY5, Galactomyces candidum RRK17 and G. candidum RRK22. Growth properties of the isolates were explored using different substrates such as 4-t-OP, 4-t-nonylphenol (4-t-NP), octylphenol polyethoxylates (OPEOn), nonylphenol polyethoxylates (NPEOn), phenol and isooctane. Of these three, the strain RRKY5 utilized a wide variety of substrates and grew faster than the others. Results of HPLC analysis indicated that 4-t-OP degradation took place both in the presence and absence of the co-substrate dextrose (0.05%): within 24 days, 93% or 95% of 4-t-OP was degraded by C. rugopelliculosa RRKY5 in media with or without dextrose, whereas degradation was lower (less than 75%) in the two strains of G. candidum. Therefore, in contrast to the 4-t-OP degradation by filamentous ligninolytic fungi, dextrose did not appear to exert a significant impact on this process in yeast cultures. It can be concluded that RRKY5 is a potent candidate for the biodegradation of 4-t-OP and thus can potentially be used for bio-remedial actions in the future. To our knowledge, this is the first evidence demonstrating that pure cultures of yeast strains are capable of utilizing branched-chain OP.
1. Introduction
In recent decades, a wide range of toxic organic pollutants predominantly used in domestic, industrial and agricultural activities have been found to interfere with the endocrine system by mimicking natural hormones or biosignalling pathways in humans and wildlife.1 These substances are called endocrine disrupting chemicals (EDCs). Of these chemicals, 4-t-octylphenol (4-t-OP) has been of great concern due to its widespread usage and higher estrogenic responses compared to other EDCs.2 The occurrence and accumulation of 4-t-OP in the environment and biota may originate from sewage treatment plants or result from incomplete in situ microbial degradation of octylphenol polyethoxylate (OPEOn) surfactants, which are used considerably in household and industrial purposes such as detergents, emulsifiers and plasticizers.3 It has been found that 4-t-OP may lead to hormone related cancers, decrease sperm quality and interfere with sexual development and reproductive systems.4,5 As consequence, exposure of 4-t-OP, even at low concentrations, can cause adverse effects in wildlife reproduction and human health, especially in infants. Because conventional physical and chemical methods applied in wastewater treatment processes are not effective for 4-t-OP elimination, biodegradation of 4-t-OP by potential microorganisms has been considered one of the efficient methods to remove 4-t-OP from the environment.6,7
Indeed, biodegradation of branched-chain OP by microbial consortia in soil, water, and sediments has been well documented.8 Pure cultures of many bacterial strains mainly belonging to sphingomonads have also been demonstrated to utilize branched-chain OP as sole carbon and energy source, and their metabolic pathways have been extensively studied as well.9,10 With respect to fungal species, available data to date, however, have indicated that degradation of branched-chain alkylphenols is mostly carried out by ligninolytic and mitosporic fungi that produce laccase.11–15 The observation that no yeasts has hitherto been shown to degrade branched-chain OP as a sole carbon source is surprising, given that yeasts are known to exhibit higher metabolic versatility and broad substrate specificity of their enzymes towards toxic pollutants,16 supposedly being able to grow abundantly in environmental matrices contaminated by these toxins.17 For example, the degradation capabilities of alkyl substituted aromatic hydrocarbons by yeasts have been widely acknowledged,18,19 and removal of organic chemicals by yeasts in bioreactors has also been demonstrated in many studies.20,21 Consequently, there may be a potential for yeasts in 4-t-OP degradation which can be applied in the bioremediation of 4-t-OP polluted environments as an effective and alternative approach compared to conventional treatments. Relevantly, earlier reports have shown that Candida maltosa isolated from a sewage treatment plant and later identified as Candida aquae-textoris was used for the biodegradation of linear-chain NP;22,23 however, most of the alkyl-substituted phenols occurring in the environment have branched-chains and are known to be more resistant to microbial degradation than phenols with linear-alkyl chains.24 This implies that towards clarifying whether the degradation pathway affects the aromatic ring or branched-alkyl chain or both parts of 4-t-OP, degradation experiments seem to have to be done not only with 4-t-OP, but also with phenol and isooctane separately.25,26
In the present study, yeasts capable of utilizing branched-chain OP as a sole carbon and energy source were isolated and investigated. To the best of our knowledge, this is the first study showing that yeast strains can degrade the branched-chain OP.
2. Experimental section
2.1. Chemicals
4-t-Octylphenol (4-t-OP; single isomer), 4-t-nonylphenol (4-t-NP; mixture of branched NP isomers), Triton X-100 (OPEOn; octylphenol polyethoxylates), Tergitol® NP-9 (NPEOn; nonylphenol polyethoxylates), phenol, and the corresponding standards with high purity (≥97%) were purchased from Sigma Chemical Co. (St. Louis, MO, USA). Isooctane (2,2,4-trimethylpentane) was obtained from Riedel-Dehean Chemicals Co. All other chemicals of analytical grade were purchased from Showa Chemicals Co, Japan. All solvents, including high-performance liquid chromatography (HPLC) grade of acetonitrile, ethyl acetate, and methanol were obtained from Sigma Chemical Co. (St. Louis, MO, USA).
2.2. Enrichment and isolation of 4-t-OP degrading yeast strains
The yeast strains were isolated from sewage water and sludge samples collected from Dihua Sewage Treatment Plant located at Taipei, Taiwan. For enrichment, water (5 mL) and sludge (5 g) were inoculated into 50 mL Modified Minimal Salt Broth (MMSB) composed of (per liter) 1.5 g Na2HPO4·7H2O, 1.123 g KH2PO4, 2.0 g (NH4)2SO4, 0.2 g MgSO4·7H2O, 0.01 g CaCl2·2H2O, 0.01 g FeSO4·7H2O, 0.815 mg (NH4)MO7O24·4H2O and 1 mL of trace metal solution. The trace metal solution included (per liter) 2.5 g Na2EDTA·2H2O, 11 g ZnSO4·7H2O, 5 g FeSO4·7H2O, 1.54 g MnSO4·H2O, 0.4 g CuSO4·5H2O, 0.25 g Co(NO3)2·6H2O and 0.18 g Na2B4O7·10H2O.27,28 Double distilled water was used to make the final volume to 1000 mL. The pH of the medium was adjusted to 6 with 1 N HCl and sterilized by autoclaving (121 °C, 1.5 kg cm−2 for 30 min). The medium was supplemented with 0.005% 4-t-OP as a sole carbon source and incubated at 25 °C on a rotary shaker (150 rpm) in the dark for 50 days. Every 10 days, 5 mL of the enriched cultures were transferred into freshly prepared pre-sterilized 45 mL modified minimal salt broth. After 5 transfers, 1 mL was diluted to 102 to 105 and 0.1 mL of the diluted liquid culture was streaked onto modified minimal salt agar (MMSA) plates containing 0.005% 4-t-OP as the sole carbon source. All the plates were incubated at 25 °C for 10 days in the dark. For further purification and identification of isolates, Yeast Malt Agar (YMA, composed of (g L−1) 5 g peptic digest of animal tissue, 3 g yeast extract, 3 g malt extract, 10 g dextrose, 20 g agar, pH 6.2) was used. During enrichment, tetracycline (100 mg L−1) was used to prevent bacterial growth. All the isolated strains were maintained at 4 °C for a month. For long term preservation, cells were frozen at −20 °C in broth culture supplemented with 30% (w/v) glycerol.29 Pure cultures of the yeast strains were deposited at the Bioresource Collection and Research Center (BCRC), Hsinchu, Taiwan.
2.3. DNA isolation and PCR amplification of 5.8S-ITS/LSU rDNA regions
The isolates were grown on YMA at 25 °C for 5 days and morphologically investigated with light microscopy.30 The isolates were further identified by analysis of the nuclear internal transcribed spacer (ITS = ITS1, 5.8S ribosomal RNA gene, ITS2) and partial ribosomal large subunit RNA gene (LSU rDNA, D1/D2 domain) sequence analysis. The three isolates were grown on Yeast Malt Broth (YMB) for 48 h and then total genomic DNA was extracted according to the protocol (Bioman Scientific Co., Ltd., Taiwan). The ITS region was amplified with the forward primer ITS1F (5′-CTTGGTCATTTAGAGGAAGTAA-3′)31 and reverse primer ITS4 (5′-TCCTCCGCTTATTGATATGC-3′)32 and the D1/D2 domain sequence of LSU rDNA with primers NL1 (5′-GCATATCAATAAGCGGAGGAAAAG-3′) and NL4 (5′-GGTCCGTGTTTCAAGACGG-3′).33 The PCR mixture consisted of 10× reaction buffer (5 μL), 25 mM MgCl2 (3 μL), dNTP-Mix (4 μL), 1 μL of primer pairs (NL1 & NL4) or (ITS1F & ITS 4), Taq-DNA polymerase (0.25 μL), ddH2O (up to 50 μL) and genomic DNA (5 μL). The amplification conditions were of initial temperature 94 °C for 30 s; 60 °C for 30 s; 72 °C for 1 min, 10 cycles followed by 28 cycles of 94 °C for 30 s; 50 °C for 30 s; 72 °C for 1 min; and finally 72 °C for 1 min. The amplified product was checked with 2% agarose gel electrophoresis subsequently by staining with GelRed™ (Biotium, Hayward, California, USA) visualized under UV light (312 nm). The PCR products were purified with GFX PCR DNA and Gel Band Purification Kit (GE Healthcare, UK). Sequencing of DNA was done by Mission Biotech (Nangang, Taipei) with the same primers as for the PCR. DNA sequences were edited in Codon Code Aligner version 4.0.1 (Codon Code Corporation, USA) and submitted to GenBank. The GenBank accession numbers and BCRC numbers of the isolated yeast strains were given in Table 1.
Table 1 Identification of 4-t-octylphenol degrading yeast strains from Dihua sewage waste water treatment plant in Taipei using BLAST with ITS & LSU rDNA sequences
| Isolate |
BCRC number |
Sample location |
GenBank accession number |
Proposed identification |
Compared length (ITS) |
Closely related species (GenBank) |
Compared length (LSU rDNA) |
Closely related species (GenBank) |
| ITS |
LSU rDNA |
Base pair identitya (%) |
Accession numbera |
Base pair identitya (%) |
Accession numbera |
| Base pair identity and accession number were obtained by BLAST search. |
| RRKY5 |
BCRC 23418 |
Water (aeration tank) |
KR815825 |
KR527144 |
Candida rugopelliculosa |
375/423 (89%) |
HE660072 |
Issatchenkia scutulata var. exigua |
557/563 (99%) |
EF550238 |
Candida rugopelliculosa |
| 370/419 (88%) |
AB704714 |
Candida phayaonensis |
543/544 (99%) |
U71069 |
Candida rugopelliculosa |
| 409/472 (87%) |
KP132499 |
Pichia cactophila |
553/573 (97%) |
KC832482 |
Pichia sp. |
| 405/473 (86%) |
KP131718 |
Candida inconspicua |
537/563 (95%) |
HE799658 |
Pichia exigua |
| RRK17 |
BCRC FU30156 |
Water (aeration tank) |
KR815826 |
KR527145 |
Galactomyces candidum |
409/411 (99%) |
JQ668740 |
Galactomyces geotrichum |
523/523 (100%) |
JN974268 |
Galactomyces candidum |
| 409/411 (98%) |
JN974288 |
Galactomyces candidum |
523/523 (100%) |
JQ317161 |
Galactomyces candidum |
| 408/411 (99%) |
JQ668739 |
Galactomyces geotrichum |
523/523 (100%) |
JF757237 |
Galactomyces geotrichum |
| 402/408 (99%) |
AJ279445 |
Galactomyces geotrichum |
522/523 (99%) |
JQ317163 |
Galactomyces candidum |
| 402/413 (97%) |
JN974286 |
Galactomyces candidum |
522/523 (99%) |
KP017413 |
Galactomyces candidum |
| 402/414 (97%) |
JN974287 |
Galactomyces candidum |
522/523 (99%) |
JN974267 |
Galactomyces candidum |
| 400/412 (97%) |
JN974289 |
Galactomyces candidum |
522/523 (99%) |
GQ373001 |
Saccharomycetales sp. |
| 398/412 (97%) |
JF262188 |
Galactomyces geotrichum |
|
|
|
| 398/412 (97%) |
KP132254 |
Galactomyces candidum |
|
|
|
| RRK22 |
BCRC FU30158 |
Sludge (biogas storage tank) |
KR815827 |
KR527146 |
Galactomyces candidum |
381/385 (99%) |
JQ668740 |
Galactomyces geotrichum |
504/504 (100%) |
JN974268 |
Galactomyces candidum |
| 380/385 (99%) |
JQ668739 |
Galactomyces geotrichum |
504/504 (100%) |
JQ317161 |
Galactomyces candidum |
| 380/386 (99%) |
AJ279445 |
Galactomyces geotrichum |
504/504 (100%) |
JF757237 |
Galactomyces geotrichum |
| 377/385 (98%) |
JN974288 |
Galactomyces candidum |
|
|
|
| 371/377 (98%) |
KF112070 |
Geotrichum candidum |
504/504 (100%) |
AB294540 |
Geotrichum sp. |
| 361/364 (99%) |
KF225039 |
Galactomyces geotrichum |
503/504 (99%) |
JQ317163 |
Galactomyces candidum |
| 361/364 (99%) |
GQ458034 |
Galactomyces geotrichum |
503/504 (99%) |
KP017413 |
Galactomyces candidum |
| 361/365 (99%) |
KF225027 |
Galactomyces geotrichum |
503/504 (99%) |
JN974267 |
Galactomyces candidum |
| 374/386 (97%) |
JN974289 |
Galactomyces candidum |
503/504 (99%) |
GQ458034 |
Galactomyces geotrichum |
| 374/387 (97%) |
JN974286 |
Galactomyces candidum |
503/505 (99%) |
JN974265 |
Galactomyces candidum |
2.4. Phylogenetic analysis
For phylogenetic analysis, our LSU rDNA gene sequences were aligned with those of closely related species retrieved from GenBank using BLAST search (http://www.ncbi.nih.gov/BLAST/) and from relevant publication about Candida and Galactomyces34,35 with MUSCLE implemented in MEGA6, without manual editing, except for trimming off the uneven ends of the alignment block. A phylogenetic tree was constructed by maximum likelihood with Kimura 2-parameter model with gamma distributed rate of nucleotide substitution and 1000 bootstrap replications.36 GenBank accession numbers are given in the unrooted trees (Fig. 2 and 3).
2.5. Substrate utilization tests
To investigate the substrate utilization, the isolated yeast strains were inoculated into 30 mL modified minimal salt broth in 250 mL conical flasks with the individual substrate as the sole carbon source. Because alkylphenol polyethoxylates (APEOn) consist of hydrophobic and hydrophilic moieties that would eventually be converted to hydrophobic intermediate alkylphenols due to microbial attack,37,38 and because further utilization and degradation of intermediate products have not been clarified with a single strain except bacteria,10,39 we selected OPEOn and NPEOn as the model APEOn and their potential intermediate products including 4-t-OP, 4-t-NP, phenol, and iso-octane as the substrates for the isolated yeast strains to address the complete degradation of APEOn. Each substrate was used at the concentration of 50 mg L−1. The initial yeast cell concentration was adjusted to OD600 = 0.1 ± 0.03. The molecular structures of 4-t-OP and related compounds are depicted in Fig. 1. All the flasks were incubated at 25 °C on a rotary shaker at 150 rpm in the dark. Minimal medium without substrate served as a negative control. All the experiments were conducted in triplicate analysis. Cell density was determined by UV-Vis spectrometric analysis at OD600. Cell growth was measured by removing 1 mL of sample from culture flasks and mixed with 50% methanol (v/v), final concentration to solubilize insoluble metabolites formed from biodegradation. The above step was done instantly prior to optical density assessment.38
 |
| | Fig. 1 Molecular structures of 4-t-OP and related compounds used in this study. | |
2.6. Biodegradation experiment
For growth experiments, 30 mL modified minimal salt broth (in 250 mL conical flask) with 4-t-OP (10 mg L−1) was prepared. Co-metabolism experiments were conducted with the same medium supplemented with 0.05% dextrose (w/v). Inoculum was prepared individually by growing the three yeast strains in 30 mL modified minimal broth with 0.002% 4-t-OP (w/v) and 1% dextrose (w/v). After 48 hours, cells were centrifuged at 4000 rpm for 10 minutes at 4 °C.40 The pellets were washed with pre-sterilized 30 mL modified minimal broth. After washing for 5 times, the final pellet was used as an inoculum for further experiments. Initial optical density was adjusted to OD600 = 0.1 ± 0.03. All the flasks were incubated at 25 °C on a rotary shaker at 150 rpm in the dark. Cell growth and 4-t-OP concentration were analyzed every 4 days over 24 days. In this experiment, two types of control were applied. MMSB supplemented with 0.001% 4-t-OP but devoid of yeast cells was used to evaluate abiotic degradation (Fig. 4a). In another set of experiment, MMSB with 0.001% 4-t-OP and dead yeast cells was used to assess carbon source adsorption to the biomass (Fig. 4a–c). Dead cells were prepared by autoclaving prior to inoculation. All experiments were performed in triplicate. The residual 4-t-OP was quantified by high performance liquid chromatography (HPLC). Percentage of remaining 4-t-OP was calculated according to the eqn (1)| | |
% of remaining 4-t-OP = 1 − (substrate peak area in the test sample)/(substrate peak area in the control sample) × 100.
| (1) |
2.7. Analytical methods
2.7.1. Extraction of 4-t-OP. 10 mL of yeast cultures containing 4-t-OP was mixed with 10 mL of 72.4% of MgSO4·7H2O (w/v) and 200 μL of 5 N H2SO4. After vigorous shaking, 20 mL of ethyl acetate was added to the solution. The organic layer (upper layer) was separated and dried by rotary evaporator under reduced pressure. 2 mL of acetonitrile was added and mixed thoroughly to dissolve the residual substrates for HPLC analysis.38
2.7.2. Spike recovery experiment. Extraction efficiency of the method was evaluated. The experiment was performed with 30 mL modified minimal salt medium. The working standards of 4-t-OP concentrations were prepared in methanol. Defined concentrations of 4-t-OP (1, 10 and 20 μg mL−1) were applied into individual flasks containing 30 mL modified minimal medium. Extraction and analysis were performed in triplicate (n = 3) with the procedure described above. The percentage of recovery was evaluated by comparing the known concentration and the concentration obtained from the recovered samples. The percentage of recovery of 4-t-OP in minimal medium was carried out using ethyl acetate as partitioning solvent. The efficiency of the method was satisfactory as the percentage acquired ranged from 82 to 93% with RSD (0.5–2%). From the results, we can conclude that if 4-t-OP is present in the culture medium, it can be extracted by using this method.
2.7.3. Quantitative analysis of residual 4-t-OP by HPLC. HPLC analysis was performed using a reverse phase HPLC (Agilent 1100) and an acclaim surfactant C18 column (4.6 × 150 mm, particle size 5 μm, Thermo) was used for quantitative analysis of 4-t-OP after degradation. The elution conditions were 60% acetonitrile (solvent A) and 40% water (solvent B). The flow rate was adjusted to 0.8 mL min−1 and UV-Vis detection wavelength was set to 277 nm.10 The injection volume was 20 μL and the temperature was fixed at 30 °C. The total run time was 17 min. Retention time (RT) of 4-t-OP was determined at RT 13.8 min. The calibration curve was plotted against the peak area versus 4-t-OP concentration. The correlation coefficient was acquired as 0.999. Limit of detection (LOD) and limit of quantification (LOQ) were 0.4 and 1.1 μg mL−1 with the mean precision of ±9% (n = 5). The LOD and LOQ were calculated by using signal to noise ratio of 3.3 and 10.41
3. Results & discussion
3.1. Isolation of 4-t-OP degrading yeast strains
Because wastewater in general contains an assortment of alkylphenols and success in the isolation of EDC-degrading microbes from activated sludge has been documented,42 in this study aqueous and sludge samples taken from a wastewater treatment plant were used for the isolation of 4-t-OP degrading yeast strains. Three yeast strains were successfully isolated using enrichment culture techniques. The method was used to make sure that the isolated strains were capable of utilizing 4-t-OP instead of only tolerating it. After each enrichment cycle, cells were observed under the light microscope to check the cell viability. The three isolated yeast strains were able to utilize 4-t-OP as the sole carbon and energy source under aerobic conditions. The three strains were assigned as RRKY5, RRK17 and RRK22. The characteristics of colony morphology and microscopic observations of the isolated strains are shown in Table 2. Based on the characteristics, RRK17 is rather similar to RRK22 while RRKY5 can be distinguished from the other strains. The two similar strains differed by their origin: RRK17 was isolated from an aeration tank and RRK22 was from a sludge tank (Table 1).
Table 2 Morphological and growth properties of yeast isolatesa
| Parameters |
Characteristics of isolates |
| RRKY5 |
RRK17 |
RRK22 |
| +++ – strong growth (OD600 ≥ 0.25); ++ – medium growth (OD600 ≥ 0.2); + – low growth (OD600 ≥ 0.15); — – no growth (OD600 ≤ 0.15). YMA – yeast malt agar. MMSB with individual substrates were used at a concentration of 50 mg L−1. MMSB – modified minimal salt broth. |
| Colony morphology on YMAb |
White, dry, butyrous, and wrinkled surface, erose margin |
White, flat, dry, powdery to finely hairy |
White, flat, dry, powdery to finely hairy |
| Microscopic observation |
Spherical, ovoid to elongate, singly in pairs, unipolar, budding cells, wrinkled and folded pellicle present |
Septate hyphae, cells doliform; arthroconidia hyaline and slightly inflated |
Septate hyphae, cells doliform; arthroconidia hyaline and slightly inflated |
![[thin space (1/6-em)]](https://www.rsc.org/images/entities/char_2009.gif) |
| Growth on OP-related compounds |
| 4-t-Octylphenolc (4-t-OP) |
+++ |
++ |
++ |
| 4-t-Nonylphenolc (4-t-NP) |
++ |
++ |
++ |
| Octylphenol polyethoxylatesc (OPEOn) |
+++ |
+++ |
+++ |
| Nonylphenol polyethoxylatesc (NPEOn) |
+++ |
+++ |
+++ |
| Phenolc |
+++ |
++ |
++ |
| Isooctanec |
+++ |
++ |
++ |
| MMSBd |
— |
— |
— |
3.2. Taxonomic relationship and phylogenetic analysis of yeast isolates
The identification of Candida species as C. maltosa reported in 1995 to degrade 4-n-NP was later corrected as C. aquae-textoris22,23 which shows that correct species identification is not trivial. Morphological and molecular comparison of our three strains indicated that two strains were closely related to each other and distantly related to the third strain. To identify the exact relationship of the three yeast strains, ITS and LSU rDNA sequences were compared with those retrieved from GenBank by BLAST search (Table 1). The BLAST search results showed that the ITS sequences of the strain RRKY5 had 89% similarity with the ITS sequence of Issatchenkia scutulatavar var. exigua HE660072, whereas 88%, 87% and 86% homology was observed for Candida phyayaonensis AB704714, Pichia cactophila KP132499 and Candida inconspicua KP131718.43–45 Based on LSU rDNA sequences, however, the strain had 99% similarity with Candida rugopelliculosa EF550238, FJ623263, and U71069,34,46 whereas 97% and 95% homology was observed for Pichia sp. (KC832482) and Pichia exigua (HE799658). The sequences of the other two strains RRK17 and RRK22 showed 97–99% similarity with those of Galactomyces geotrichum and G. candidum. Using LSU rDNA sequences of the two strains with BLAST, the highest similarities were 99% with sequences of G. candidum and G. geotrichum35,47,48 and below 97% with the sequences of the other species (Table 1). Based on the BLAST search with the ITS and LSU rDNA sequences, the highest percentage of similarities was observed in different species. The results revealed that the species identifications of the isolated strains were still inconclusive. The phylogenetic analysis based on D1/D2 domain sequences confirmed that the strain RRKY5 belonged to Candida rugopelliculosa (Fig. 2). Compared to inconsistent BLAST search results with ITS and LSU rDNA sequences of the strains RRK17 and RRK22, phylogenetic relationship of the sequences of 28S rDNA showed that the two yeast strains belonged to Galactomyces candidum, not G. geotrichum (Fig. 3). Although ITS sequences are proposed as barcode for fungi, deviating intraspecific variabilities in different fungal groups and a high proportion of database entries with wrong species names limit the usefulness for identification purposes.49 Particular further analysis and mycological expertise is required to interpret the data correctly. The species G. bryndzae and G. silvicola nesting in the well supported clade of G. candidum in Fig. 3 are considered synonyms of G. candidum by experts of yeast systematics. Candida rugopelliculosa and G. candidum were used for the decolorization of dye treatment under aerobic conditions50,51 which indicates their ability to decompose complex aromatic compounds and potential for application in wastewater treatment.
 |
| | Fig. 2 Phylogenetic analysis of Candida species constructed by maximum likelihood analysis of D1/D2 LSU rDNA sequences. Branch lengths are comparative to nucleotides differences, and the numbers given at the nodes are the percentage of frequencies. Bootstrap is based on 1000 replicates. Frequencies below 50% are not given. Candida bentonensis was used as out group (T = ex-type strain). | |
 |
| | Fig. 3 Phylogenetic analysis of Galactomyces species constructed by maximum likelihood analysis of D1/D2 LSU rDNA sequences. Branch lengths are comparative to nucleotides differences, and the numbers given at the nodes are the percentage of frequencies. Bootstrap is based on 1000 replicates. Frequencies below 50% are not given. Dipodascus albidus was used as out group (T = ex-type strain). | |
3.3. Substrate utilization tests
For substrate utilization, minimal medium supplemented with each substrate at a concentration of 50 mg L−1 was used. The growth of the yeast strains differed for each substrate (Table 2). All the three yeast strains were able to utilize different substrates as sole carbon and energy sources. Candida rugopelliculosa grew rapidly on a wide range of substrates such as 4-t-OP, 4-t-NP, OPEOn, NPEOn, phenol and iso-octane; the two G. candidum strains were also able to grow effectively on these substrates except phenol and iso-octane. In the negative control, no growth was observed in all the three yeast strains. The utilization of phenol and iso-octane suggested that the degradation of 4-t-OP can be initiated via both aromatic ring hydroxylation and alkyl chain oxidation. The degradation of 4-t-OP starts through either cleavage of the aromatic ring or shortening of the alkyl moiety with the subsequent detachment of the methyl group at the α-carbon.10,52 So far, none of the prokaryotic and eukaryotic microorganisms have been shown to involve both mechanisms in the degradation of 4-t-OP.
3.4. Analysis of residual 4-t-OP concentration by HPLC
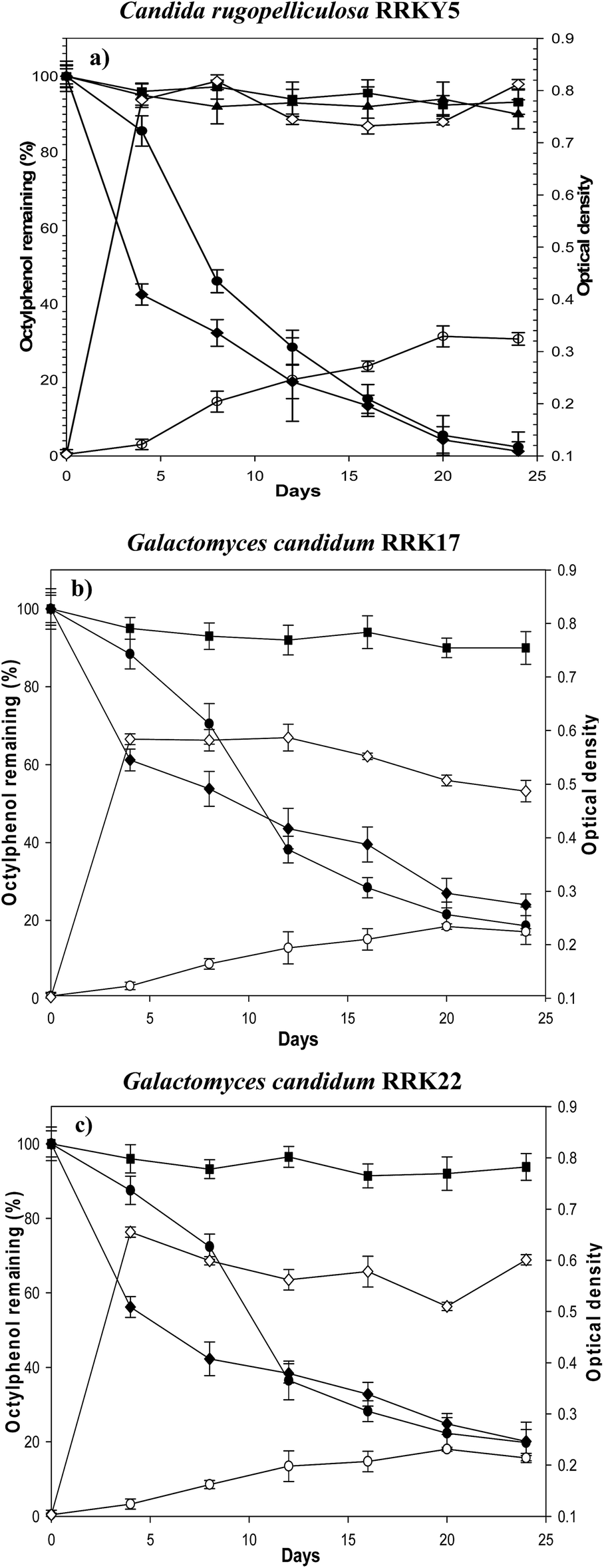
All the yeast strains were capable of degrading 4-t-OP as the sole carbon and energy source in the medium with and without dextrose (Fig. 4a–c). In the controls including modified minimal salt broth (MMSB) without yeast cells and MMSB with dead yeast cells, the results indicated that there was no significant abiotic degradation and cell adsorption of compounds during incubation periods of C. rugopelliculosa (Fig. 4a). Only dead cells were, therefore, applied as controls in the experiments with G. candidum. In both conditions, the 4-t-OP concentration was recovered between 92–95% after 24 days. Under aerobic conditions, the three yeast strains effectively degraded 4-t-OP (10 mg L−1) in media supplemented with and without dextrose. In the medium without dextrose, the isolated yeast strains RRKY5, RRK17 and RRK22 efficiently degraded 4-t-OP by removing 71.4%, 63.5% and 64.2%, respectively, within 12 days, and cell densities (OD600) increased in parallel with the 4-t-OP degradation. After 20 days, the percentage of 4-t-OP degradation rose to 97% in RRKY5, whereas only 78% and 74% 4-t-OP degradation was observed in RRK17 and RRK22. Cell biomass increased more in RRKY5 compared with RRK17 and RRK22. Interestingly, the addition of trace amount of dextrose (0.5 g L−1) only influenced the initial 4-t-OP degradation detected in this study. After 4 days, the highest degradation percentage of 58% was observed in RRKY5 compared to RRK17 and RRK22 (39% and 44%, respectively). During the subsequent period, i.e., after 20 days, the degradation rate increased to 94% in RRKY5, and 76% and 80% degradation were detected in the other yeast strains. In a study of a white-rot basidiomycete Phanerochaete sordida, the percentage of 4-t-OP degradation at an initial concentration of 20 mg L−1 after 4 days in the presence of elevated dextrose concentrations (3% w/v) was higher (80%) than that in the present study.13 Similarly, the complete 4-t-OP degradation by another filamentous basidiomycete, Gymnopus quercophilus (as “Marasmius quercophilus”), was observed also in a nutrient rich medium.14
 |
| | Fig. 4 Biodegradation of 4-t-OP by Candida rugopelliculosa RRKY5 (a), Galactomyces candidum RRK17 (b), Galactomyces candidum RRK22 (c) in 30 mL MMSB. Percentage of 4-t-OP (10 mg L−1) degradation in medium without (●) and with 0.05% dextrose (♦) respectively. Cell growth was given by increasing cell density on medium without (○) and with dextrose 0.05% (◇). Two types of control were used (in (a)). Control I contains MMSB with 4-t-OP without yeast cells (▲). Control II (in (a–c)) contains MMSB with 4-t-OP (10 mg L−1) and dead cells (■). Error bar represents standard deviation (SD) of triplicate analysis. | |
Most of previous studies reported the microbial degradation of 4-t-OP by bacteria, hyphal fungi, or microbial consortia.9–11 To our knowledge no reports have been published about the degradation of 4-t-OP by identified yeast strains isolated from wastewater treatment plants. Vallini et al., (2001)23 reported that Candida aquae-textoris was able to utilize 4-n-NP as a sole carbon source which was degraded completely after 14 days. The biodegradation of different NP isomers by the bacterium Sphingomonas sp. TTNP3 showed that the degradation was more efficient in the linear-chain than branched-alkyl chain structure.9 In addition, earlier studies demonstrated that the alkylphenols cannot be completely removed in activated sludge with batch microcosm treatment, because the alkylphenols become bound to the sewage sludge.53 The alternative approach is the application of a single strain, the so called bio-augmentation to eliminate various xenobiotics.54 So far, the removal of NP during wastewater treatment was attained by the application of bio-augmented Sphingomonas sp. in membrane bioreactor containing activated sludge.55 To the best of our knowledge, none of the studies have reported the removal of 4-t-OP in activated sludge by a single microbial strain. Using a bio-augmented pure yeast strain in a membrane bioreactor can be considered for the effective removal of 4-t-OP from activated sludge during the wastewater treatment. According to a previous report, the degradation of linear-chain nonylphenol by C. aquae-textoris was initiated via ω-oxidation of the alkyl chain followed by β-oxidation to form carboxylic acid as a major product.22 The mechanism and enzymes involved in the degradation of phenol substituted branched alkyl chains by yeast strains is completely unknown. The general mechanisms underlying degradation of branched-chain alkylphenols have been demonstrated in both bacteria and filamentous fungi.10,11,39,52 There are four major mechanisms: (1) aromatic ring hydroxylation; (2) type II ipso-substitution, resulting in the formation of alkylcatechol and hydroquinone used by bacteria mediated by dioxygenases;10,39 (3) oxidative radical polymerization and terminal oxidation of the alkyl chain to form oligomers and higher transformation products in ligninolytic fungi. Several enzymes are responsible, including extracellular lignin modifying enzyme (LMEs) and cytochrome P450 monooxygenases;11 (4) alkyl chain oxidation leading to the formation of carboxylic acid as a major product in non-ligninolytic fungi. The possible enzymes have not been characterized.52 In order to address degradation mechanisms, in a first step we are analyzing the metabolites in an ongoing study. According to these records, filamentous ligninolytic basidiomycetes require additional substrates or nutrient rich condition in order to remove toxic pollutants, such as 4-t-OP effectively from the environment. Under natural conditions, these fungi grow in solid substrates, which should be considered also for biotechnological approaches. In contrast to these fungi, the yeast strain Candida rugopelliculosa RRKY5 has the potential for biotechnological removal of 4-t-OP from contaminated water under nutrient rich as well as nutrient poor conditions.
For future development of industrial wastewater treatment under aerobic conditions, it is aimed to obtain the complete removal of alkylphenols with application of yeast strains or their enzymes. The process parameters are to be optimized for the novel strain of C. rugopelliculosa.
4. Conclusions
Branched-chain OP degradation by yeast strains isolated from a wastewater treatment plant has been demonstrated for the first time. The isolates were identified as Candida rugopelliculosa RRKY5, Galactomyces candidum RRK17 and G. candidum RRK22. Most effective degradation of 4-t-OP was observed in C. rugopelliculosa RRKY5, whereas the presence and absence of dextrose had no significant effect in any of the yeast strains. Compared to ligninolytic filamentous fungi which require nutrient rich media and in nature only grow on solid substrates, the yeast C. rugopelliculosa provides additional potential of application in bioremediation of 4-t-OP contaminated environment and enhancement of removal of 4-t-OP from industrial wastewater. Further studies are required to identify the exact mechanism of 4-t-OP degradation by yeast strains.
Acknowledgements
The authors would like to thank Shiu-Mei Liu and Ching-Fu Lee for their valuable suggestions. Ranjith kumar is also grateful for a grant from the office of international affairs of National Central University. The study was also supported by the Ministry of Science and Technology, Taiwan (NSC102-2621-B-008-001-MY3). Taipei city government is acknowledged for giving permission to take samples and initiating the work. We also thank Hsin Chen, Jr-Kuen Hsu, Li-Chia Liu and Yu-Hung Yeh for their technical support in the laboratory.
References
- T. T. Schug, A. Janesick, B. Blumberg and J. J. Heindel, J. Steroid Biochem. Mol. Biol., 2011, 127, 204–215 CrossRef CAS PubMed.
- J. P. Sumpter and S. Jobling, Environ. Health Perspect., 1995, 103, 173–178 CrossRef CAS PubMed.
- G. G. Ying, B. Williams and R. Kookana, Environ. Int., 2002, 28, 215–226 CrossRef CAS PubMed.
- F. R. Boockfor and C. A. Blake, Biol. Reprod., 1997, 57, 267–277 CrossRef CAS PubMed.
- T. Raecker, B. Thiele, R. M. Boehme and K. Guenther, Chemosphere, 2011, 82, 1533–1540 CrossRef CAS PubMed.
- N. Nakada, H. Shinohara, A. Murata, K. Kiri, S. Managaki, N. Sato and H. Takada, Water Res., 2007, 41, 4373–4382 CrossRef CAS PubMed.
- G. Bertanza, R. Pedrazzani, V. Zambarda, M. Dal Grande, F. Icarelli and L. Baldassarre, Water Sci. Technol., 2010, 61, 1663–1671 CrossRef CAS PubMed.
- G. G. Ying and R. S. Kookana, Environ. Sci. Technol., 2003, 37, 1256–1260 CrossRef CAS.
- B. Kolvenbach and P. X. Corvini, New Biotechnol., 2012, 30, 88–95 CrossRef CAS PubMed.
- T. Toyama, M. Murashita, K. Kobayashi, S. Kikuchi, K. Sei, Y. Tanaka, M. Ike and K. Mori, Environ. Sci. Technol., 2011, 45, 6524–6530 CrossRef CAS PubMed.
- T. Cajthaml, Environ. Microbiol., 2015, 17, 4822–4834 CrossRef CAS PubMed.
- C. Junghanns, M. Moeder, G. Krauss, C. Martin and D. Schlosser, Microbiology, 2005, 151, 45–57 CrossRef CAS PubMed.
- Y. Tamagawa, H. Hirai, S. Kawai and T. Nishida, Environ. Toxicol., 2007, 22, 281–286 CrossRef CAS PubMed.
- A. Farnet, A. Chevremont, G. Gil, S. Gastaldi and E. Ferre, Chemosphere, 2011, 82, 284–289 CrossRef CAS PubMed.
- D. S. Moon and H. G. Song, Appl. Biochem. Biotechnol., 2012, 168, 542–549 CrossRef CAS PubMed.
- G. Bleve, C. Lezzi, M. Chiriatti, I. D'Ostuni, M. Tristezza, D. Di Venere, L. Sergio, G. Mita and F. Grieco, Bioresour. Technol., 2011, 102, 982–989 CrossRef CAS PubMed.
- J. B. Sutherland, Fungal Biotechnology in Agricultural, Food and Environmental Applications, 2004, pp. 443–455 Search PubMed.
- C. E. Cerniglia and S. A. Crow, Arch. Microbiol., 1981, 129, 9–13 CrossRef CAS.
- W. J. Middelhoven, Antonie van Leeuwenhoek, 1993, 63, 125–144 CrossRef CAS PubMed.
- C. Juárez-Ramírez, N. Ruiz-Ordaz, E. Cristiani-Urbina and J. Galíndez-Mayer, World J. Microbiol. Biotechnol., 2001, 17, 697–705 CrossRef.
- J. Woertz, K. Kinney, N. McIntosh and P. Szaniszlo, Biotechnol. Bioeng., 2001, 75, 550–558 CrossRef CAS PubMed.
- A. Corti, S. Frassinetti, G. Vallini, S. D'Antone, C. Fichi and R. Solaro, Environ. Pollut., 1995, 90, 83–87 CrossRef CAS PubMed.
- G. Vallini, S. Frassinetti, F. D'Andrea, G. Catelani and M. Agnolucci, Int. Biodeterior. Biodegrad., 2001, 47, 133–140 CrossRef CAS.
- J. Shan, B. Jiang, B. Yu, C. Li, Y. Sun, H. Guo, J. Wu, E. Klumpp, A. Schäffer and R. Ji, Environ. Sci. Technol., 2011, 45, 8283–8289 CrossRef CAS PubMed.
- H. Y. Neujahr and J. M. Varga, Eur. J. Biochem., 1970, 13, 37–44 CrossRef CAS PubMed.
- F. Solano-Serena, R. Marchal, S. Heiss and J. P. Vandecasteele, J. Appl. Microbiol., 2004, 97, 629–639 CrossRef CAS PubMed.
- H. J. Chen, D. H. Tseng and S. L. Huang, Bioresour. Technol., 2005, 96, 1483–1491 CrossRef CAS PubMed.
- S. Chen, Q. Hu, M. Hu, J. Luo, Q. Weng and K. Lai, Bioresour. Technol., 2011, 102, 8110–8116 CrossRef CAS PubMed.
- C. F. Chang, C. C. Chen, C. F. Lee and S. M. Liu, Antonie van Leeuwenhoek, 2013, 104, 1117–1123 CrossRef CAS PubMed.
- C. Kurtzman, J. W. Fell and T. Boekhout, The yeasts: a taxonomic study, Elsevier, 2011 Search PubMed.
- M. Gardes and T. D. Bruns, Mol. Ecol., 1993, 2, 113–118 CrossRef CAS PubMed.
- T. J. White, T. Bruns, S. Lee and J. Taylor, PCR protocols: a guide to methods and applications, 1990, vol. 18, pp. 315–322 Search PubMed.
- C. Kurtzman and C. Robnett, J. Clin. Microbiol., 1997, 35, 1216–1223 CAS.
- C. P. Kurtzman, C. J. Robnett and E. Basehoar-Powers, FEMS Yeast Res., 2008, 8, 939–954 CrossRef CAS PubMed.
- M. Groenewald, T. Coutinho, M. T. Smith and J. P. van der Walt, Int. J. Syst. Evol. Microbiol., 2012, 62, 3072–3080 CrossRef PubMed.
- K. Tamura, G. Stecher, D. Peterson, A. Filipski and S. Kumar, Mol. Biol. Evol., 2013, 30, 2725–2729 CrossRef CAS PubMed.
- X. Gu, Y. Zhang, J. Zhang, M. Yang, H. Tamaki, Y. Kamagata and D. Li, Chemosphere, 2010, 80, 216–222 CrossRef CAS PubMed.
- Y. W. Lin, G. L. Guo, H. C. Hsieh and S. L. Huang, Bioresour. Technol., 2010, 101, 2853–2859 CrossRef CAS PubMed.
- N. N. Tuan, H. C. Hsieh, Y. W. Lin and S. L. Huang, Bioresour. Technol., 2011, 102, 4232–4240 CrossRef PubMed.
- A. J. Karley, S. I. Powell and J. M. Davies, Appl. Environ. Microbiol., 1997, 63, 1312–1317 CAS.
- V. Andreu, E. Ferrer, J. L. Rubio, G. Font and Y. Picó, Sci. Total Environ., 2007, 378, 124–129 CrossRef CAS PubMed.
- C. P. Yu, H. Roh and K. H. Chu, Environ. Sci. Technol., 2007, 41, 486–492 CrossRef CAS PubMed.
- N. Golić, N. Čadež, A. Terzić-Vidojević, H. Šuranská, J. Beganović, J. Lozo, B. Kos, J. Šušković, P. Raspor and L. Topisirović, Int. J. Food Microbiol., 2013, 166, 294–300 CrossRef PubMed.
- S. Limtong, S. Nitiyon, R. Kaewwichian, S. Jindamorakot, S. Am-In and W. Yongmanitchai, Int. J. Syst. Evol. Microbiol., 2012, 62, 2786–2792 CrossRef CAS PubMed.
- L. Irinyi, C. Serena, D. Garcia-Hermoso, M. Arabatzis, M. Desnos-Ollivier, D. Vu, G. Cardinali, I. Arthur, A. C. Normand and A. Giraldo, Med. Mycol., 2015, 53, 313–337 CrossRef PubMed.
- C. P. Kurtzman and C. J. Robnett, Antonie van Leeuwenhoek, 1998, 73, 331–371 CrossRef CAS PubMed.
- N. M. Koch, I. M. Soto, M. Galvagno, E. Hasson and L. Iannone, Fungal Ecol., 2015, 13, 167–180 CrossRef.
- J. Zhang, J. Shi, H. Lv and Y. Liu, Appl. Microbiol. Biotechnol., 2013, 97, 1279–1287 CrossRef CAS PubMed.
- R. H. Nilsson, E. Kristiansson, M. Ryberg, N. Hallenberg and K. H. Larsson, Evol. Bioinf. Online, 2008, 4, 193–201 Search PubMed.
- S. J. Kim and M. Shoda, Appl. Environ. Microbiol., 1999, 65, 1029–1035 CAS.
- X. Liu, J. Zhang, J. Jiang, R. Li, Z. Xie and S. Li, Int. Biodeterior. Biodegrad., 2011, 65, 135–141 CrossRef CAS.
- T. Janicki, M. Krupiński and J. Długoński, Bioresour. Technol., 2016, 200, 223–229 CrossRef CAS PubMed.
- A. C. Johnson and J. P. Sumpter, Environ. Sci. Technol., 2001, 35, 4697–4703 CrossRef CAS PubMed.
- A. O. Olaniran, D. Pillay and B. Pillay, Chemosphere, 2006, 63, 600–608 CrossRef CAS PubMed.
- M. Cirja, G. Hommes, P. Ivashechkin, J. Prell, A. Schäffer, P. F. Corvini and M. Lenz, Appl. Microbiol. Biotechnol., 2009, 84, 183–189 CrossRef CAS PubMed.
|
| This journal is © The Royal Society of Chemistry 2016 |
Click here to see how this site uses Cookies. View our privacy policy here.