
Evaluating the biological impact of polyhydroxyalkanoates (PHAs) on developmental and exploratory profile of zebrafish larvae†
Xiang Li‡
a,
Xu Li‡b,
Di Chen‡c,
Jia-Liang Guo‡d,
Dao-Fu Fenge,
Ming-Zhu Sunc,
Yun Lu*d,
Dong-Yan Chen*b,
Xin Zhao*c and
Xi-Zeng Feng*a
aState Key Laboratory of Medicinal Chemical Biology, The Key Laboratory of Bioactive Materials, Ministry of Education, College of Life Science, Nankai University, Tianjin 300071, China. E-mail: xzfeng@nankai.edu.cn
bTianjin Key Laboratory of Tumor Microenvironment and Neurovascular Regulation, Department of Histology and Embryology, School of Medicine, Nankai University, Tianjin 300071, China. E-mail: chendy@nankai.edu.cn
cThe Institute of Robotics and Automatic Information Systems, Nankai University, Tianjin 300071, China. E-mail: zhaoxin@nankai.edu.cn
dTEDA Hospital, No. 65 Third Avenue, Economic-Technological Development Area, Tianjin 300457, China. E-mail: tedaluyun@163.com
eDepartment of General Surgery, Tianjin Medical University General Hospital, No. 154 Anshan Road, Tianjin, 300052, China
First published on 7th April 2016
Abstract
In this study, we employed zebrafish as an animal model to evaluate the biological effect of polyhydroxyalkanoates (PHAs) on early development via morphological, physiological, and behavioural analyses. Developmental results indicated no obvious adverse effects of PHAs on the development of zebrafish embryos and larvae, except the occurrence of krox20 expression fluctuation. Furthermore, standard and colour-enriched open field tests were conducted to assess the natural colour preference/avoidance behaviour of zebrafish larvae as well as the effect of PHA concentration on patterns of exploratory behaviour and natural colour preference/avoidance. The behavioural results were as follows: (1) compared with un-injected larvae, PHA-injected larvae displayed enhanced exploratory behaviour with decreased thigmotaxis (central avoidance) in the open field test and increased thigmotaxis (central avoidance) in the colour-enriched open field test. (2) PHA injection attenuated anxiety-like behaviour by decreasing latency prior to exploration. (3) PHAs increased the preference for blue, red and black colours. In conclusion, PHAs have a concentration-dependent effect on strengthening exploratory behaviour, lessening anxious behaviour and altering the colour preference/avoidance patterns of larval zebrafish. These results suggest that PHAs have a potential effect on the behaviour of zebrafish.
Introduction
Polyhydroxyalkanoates (PHAs) are a series of linear polyesters consisting of hydroxy derivatives of fatty acids that are produced by a wide range of microorganisms.1,2 PHAs are typically highly biodegradable and biocompatible. In addition, PHAs are degraded to carbon dioxide and water in biological environments. Due to these superior properties of PHAs, they are widely used for biomedical materials.3 For example, PHAs have been applied extensively for medical and pharmacological purposes, such as wound dressing, tissue-engineered cardiovascular devices, bone-marrow scaffolds, tendon repair devices, nerve guides, articular cartilage repair devices, guided tissue repair devices, stents, adhesion barriers, orthopaedic pins, slings, repair patches, nerve repair devices, sutures, drug delivery systems, etc. Because of the prevalence of PHAs in medical applications, PHAs are increasingly used in patients who need tissue or organ repair or as a drug delivery system. PHAs have demonstrated promising biocompatibility for medical and pharmacological applications and have drawn the attention of many researchers over the past 20 years.4–6 E. I. Shishatskaya and his research group have spent years evaluating the biological effects of various forms of PHAs via in vitro and in vivo experiments. Overall, the authors found PHAs to be universally biocompatible and to cause no adverse changes in the physiological or biochemical parameters of laboratory animals (mice and rats) over an extended period of observation, although biocompatibility depends on the size, shape and purity of the material used in the implant.7–12 Although the biological evaluation of PHAs in animal models such as mice and rats has been exhaustive, there is a paucity of research examining the biological effect of PHAs in zebrafish. Furthermore, the effect of PHAs on the early development of zebrafish embryos and on neuro-behaviours (exploratory behaviour and colour preference/avoidance) remains undocumented.Traditionally, due to low-cost husbandry and housing, high fecundity,13 a short developmental period combined with well-characterized developmental stages, embryo transparency, and the convenience of microinjections,13–15 zebrafish (Danio rerio) has been a promising animal model for toxicity and biocompatibility assessments.16–23 An increasing number of behavioural neuroscience laboratories have begun employing zebrafish as their primary research organism.24 Important properties, such as genetic homology with most human genes,25 neurotransmitter systems similar to those of mammals,26 and a large and variable behavioural repertoire,27 render zebrafish an efficient system for behaviour-based screening used to validate the pharmacological effects of drugs, neuro-active compounds and biomaterials in medical applications.27–31 In addition, the popularity of zebrafish as a novel neuro-behavioural model for high-throughput screening has promoted the use of automated video systems to record behavioural changes following drug or biomaterial treatment.32–34
Among the various behaviour-based screening methodologies, one prevalent assay known as the “exploratory-driven anxiety model” (open field test) is regarded as a well-accepted standard in preclinical research on animals (rodents) treated with neuro-active compounds to assess behavioural reactions to environmental complexity.28,29,35,36,38–44 We developed and adapted the open field test and expanded the behavioural parameters to investigate the effects of different concentrations of PHAs on the behaviours of zebrafish larvae.
In this study, we created a wide range of concentrations (0.25 μg mL−1 to 1500 μg mL−1) by diluting PHAs in standard system water (egg water). We then microinjected the PHA solutions into embryos at the 1–4 cell stage to evaluate the effect of PHAs on the early development of embryonic and larval zebrafish. Once injected, the embryos developed to 6 day post-fertilization (dpf), and then the larval zebrafish were subjected to open field tests40 to assess the effects of different concentrations of PHAs on the exploratory behaviour of environmental complexity (colour enrichment) and on colour preference. The result of the developmental toxicology analysis demonstrated that the injection of PHA microspheres into embryonic yolk caused no adverse changes in development, morphology, mortality or the patterns of early gene expression in embryonic and larval zebrafish during the sensitive early embryonic developmental stages (from 4 hpf to 96 hpf). To detect the behavioural effect of PHAs, open field tests were conducted with larvae at 6 dpf following PHA injection. Interestingly, despite excellent biocompatibility in vivo, PHAs exerted a behavioural effect on exploratory behaviours and colour preference behaviours of zebrafish larvae. Specifically, PHAs decreased the novel anxiety/stress response of animals when they encountered a novel and complex environment (colour-enriched), according to the well-established ‘exploratory-driven anxiety model’ theory.35 Furthermore, PHA injection elevated the level of exploratory behaviour. Compared with the control (un-injected) zebrafish larvae, PHA-injected larval zebrafish were more likely to explore the colour-enhanced peripheral zones in the open field test. The present study is, to our knowledge, the first research concerning the developmental and neuro-behavioural effects of PHAs on zebrafish embryos and larvae. Moreover, we demonstrated that PHAs may exert concentration-dependent behavioural effects on animal exploratory behaviour and colour preference behaviour. These findings illustrate the potential behavioural effects of PHAs and their homologous series.
Results
Characterization of PHAs particles in the standard system water
To simulate the environmental conditions under which PHAs particulates are distributed in an aquatic system, PHAs of the different masses used in this experiment were directly added to standard system water (egg water) and stirred to a relatively homogenous state with an immersion blender. Dynamic light scattering (DLS) was then performed to determine the size, charge and stability of PHA particles in the standard tank water (egg water). Subsequently, PHA suspensions of different concentrations were microinjected into embryonic yolk at the 1–4 cell stages. The results of the DLS (Fig. 2A) demonstrated that PHA particles in the standard system water were agglomerated, and the size of the agglomeration increased as the concentration increased. The mean particle sizes and zeta potentials of the PHAs in the standard tank water are presented in Fig. 2B. Although the mean diameters increased as the concentration increased, the charge remained relatively constant. | ||
| Fig. 1 Schematic diagram of the experimental procedure. (A) A wide range of concentrations of PHA particles (from 0.25 μg mL−1 to 1500 μg mL−1) were microinjected into the yolk of zebrafish embryos at the two cell stage (n > 50 for each concentration). (B) PHA-injected embryos were then prepared for morphological and gene expression analyses. (C) When PHA-injected embryos developed into larvae, they were observed morphologically and analysed behaviourally for exploratory and colour preference behaviours. Larvae at 6 days post-fertilization (dpf) were transferred to the standard and colour-enriched open field apparatus for exploratory testing, which has been previously described in detail.40 As depicted in Fig. S4 and S5,† the experimental setup was composed of a light source (alternate white and infrared light arrays in a platform), an infrared camera (30 frames per s) mounted above the platform and a paired standard and colour-enriched open field apparatus centred beneath the camera. The colour-enriched open field test apparatus was a glass Petri dish (9 cm in diameter) divided into an inner transparent area (a concentric circle of the dish 2 cm in diameter), used as the starting location, and an outer zone which was equally divided into six sectors. The sectors were coloured yellow, red, green, orange, blue, and black by the addition of colour photographic filters to ensure the permeability of visible and infrared light from the light source. The standard open field apparatus (left) was identical to the colour-enriched open field apparatus except that the bottom and sides were transparent. The duration of the test was 15 min. Prior to the experiment, one standard and one colour-enriched open field dish were placed on top of the white/infrared light platform and the larvae were allowed 15 min to adapt to the test environment. The behaviour of PHA-treated zebrafish in both the standard and colour-enriched open field apparatus were video-recorded and analysed. | ||
 | ||
| Fig. 2 DLS results for PHAs in standard system water (egg water). (A) Structural formula of PHAs in this experiment and the hydrodynamic diameters of PHAs particles in different concentration. (B) Results for diameters and surface charges based on the DLS results for the PHAs particles in standard system water. | ||
Effect of PHAs on embryonic and larval development
Given that the minimal toxic effect of PHAs has been validated in other animal models such as Wistar rats,7,9,10,12 only three of the PHA-injected groups with concentrations of 25 μg mL−1, 300 μg mL−1 and 1000 μg mL−1 were chosen for developmental evaluation. For the early embryonic developmental validation, the percentage of cells showing epiboly (Fig. 3A) and mortality (Fig. S8†) was recorded at 4 hpf, 5.66 hpf, 9 hpf, 10 hpf, and 24 hpf. Obvious promotion in the epiboly process by PHAs was detected, but no significant embryonic abnormality or mortality rates (Fig. S8†) were detected (Fig. 3B). | ||
| Fig. 3 Effects of PHAs microinjection on embryonic development. (A) Morphological characteristics of un-injected embryos and embryos injected with PHAs (0 μg mL−1, 25 μg mL−1, 300 μg mL−1 and 1000 μg mL−1) from 4 to 24 hpf. The red lines represent the start and end of epiboly. (B) The epiboly percentages of the PHA-injected (0 μg mL−1, 25 μg mL−1, 300 μg mL−1 and 1000 μg mL−1) embryos at 9 hpf. (C) The epiboly percentages of the PHA-injected (0 μg mL−1, 25 μg mL−1, 300 μg mL−1 and 1000 μg mL−1) embryos at 10 hpf. | ||
Due to the normal morphology, morphogenesis of embryos and larvae, the three key functional genes discussed above were chosen for detection. ntl, a member of T-box transcription factor family, is a marker for the induction of the mesoderm which gives rise to posterior body structures, and was expressed in the margin, axial chordal mesoderm, and notochord during the development of uninjected embryos. Here we used ntl as a marker for cell movement during gastrulation which was an important process in embryo development. Cardiac myosin light chain 2 (cmlc2), an essential component of thick myofilament assembly and contractility of heart, has been frequently been used as a heart-specific marker. Krox20 (also known as Egr2) is a transcription factor that is involved in vertebrate hindbrain segmentation, particularly in the formation and specification of rhombomeres (r) 3 and 5, as important marker of zebrafish brain.
Whole-mount in situ hybridization of several key genes including ntl, krox20 and cmlc2 was conducted to assess the effect of PHAs on the spatial and temporal patterns of embryonic gene expression (Fig. 4B). The level of ntl and cmlc2 expression were spatially and temporally normal (Fig. 4A). Only expression of krox20 fluctuated as the PHAs concentration increased. Embryonic heart rate was also measured and analysed; no significant change was found (Fig. 4C).
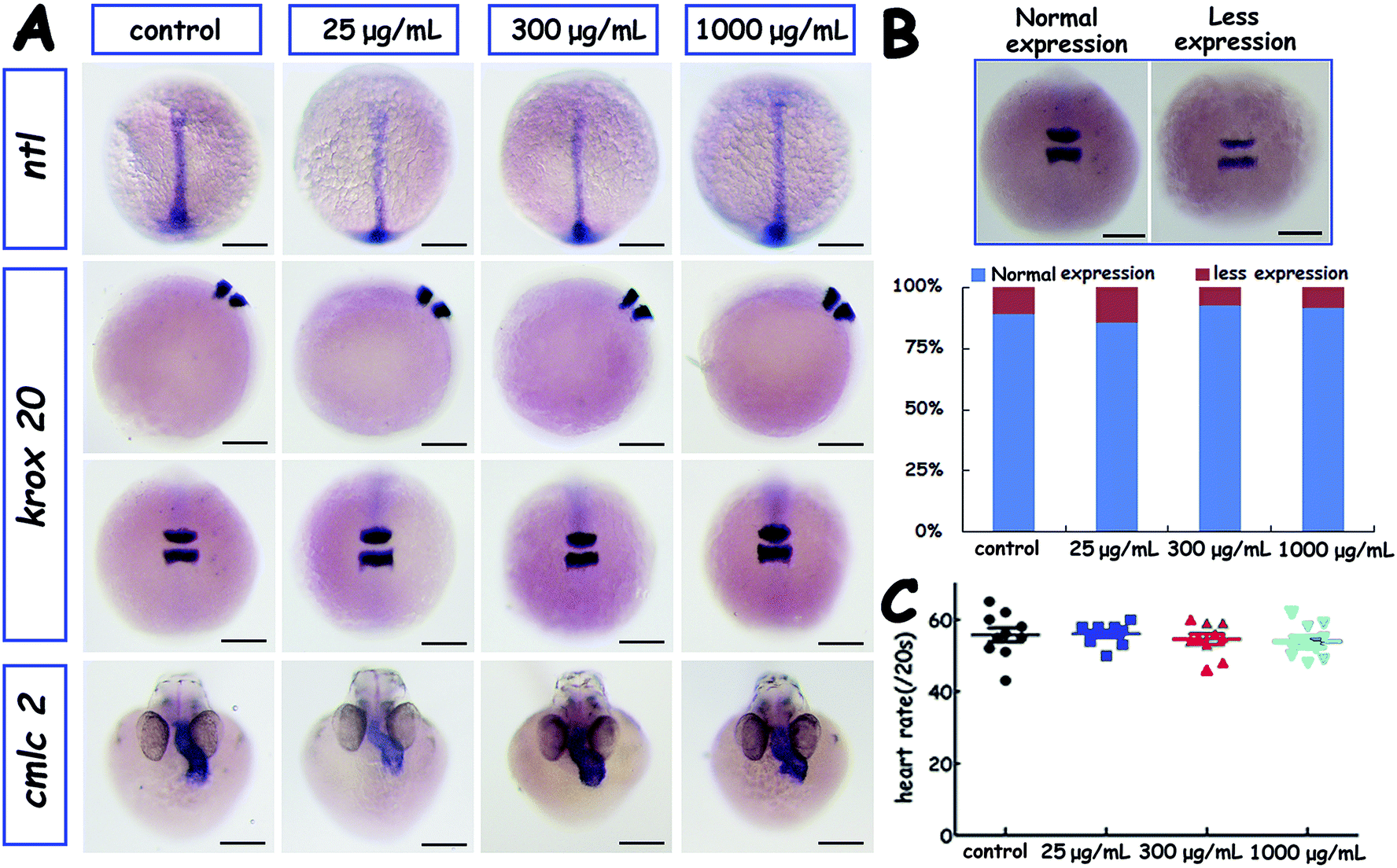 | ||
| Fig. 4 Gene expression in embryos injected with different concentrations of PHAs. (A) Expression of ntl at 10 hpf, krox20 at 12 hpf, and cmlc2 at 28 hpf in embryos injected with 0 μg mL−1, 25 μg mL−1, 300 μg mL−1, 1000 μg mL−1 PHA. (B) Expression of krox20 at 12 hpf in control and PHA-injected embryos. Scale bars = 200 μm. (C) Heart rate at 28 hpf in un-injected embryos and PHA-injected embryos. | ||
Morphological assessment and analysis were conducted for early larval development at 28 hpf, 48 hpf, 72 hpf, and 96 hpf (Fig. 5A). Developmental parameters such as body length, head area and eye area were compared among the different concentrations, and there was no significant difference for any of the parameters (Fig. 5B).
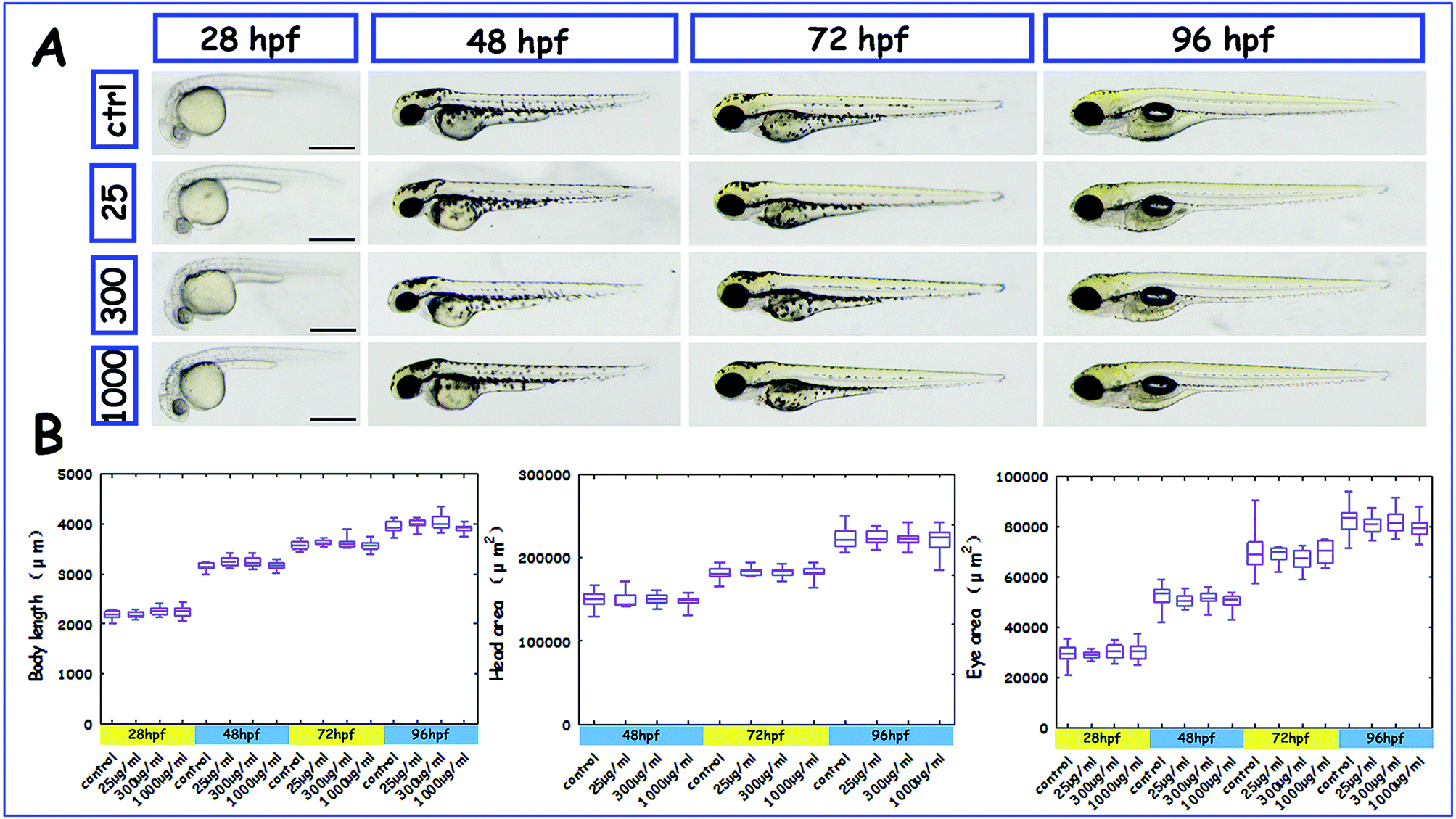 | ||
| Fig. 5 Phenotypes of zebrafish larvae injected with different concentrations of PHAs. (A) Morphological characteristics of un-injected larvae and larvae injected with PHAs (0 μg mL−1, 25 μg mL−1, 300 μg mL−1 and 1000 μg mL−1) from 28 hpf to 96 hpf. (B) Body length, head area and eye area of un-injected and injected zebrafish larvae from 28 hpf to 96 hpf. The data are presented as the mean ± SEM. Scale bars = 200 μm. | ||
Effect of PHAs on total activity level and temporal locomotor activity patterns of zebrafish larvae
In the standard open field test, a one-way ANOVA revealed no significant effect of PHA concentration on the total activity level of zebrafish larvae [F(7,184) = 1.7576, P = 0.46] (Fig. 6A). In addition, in the colour-enriched open field test, a one-way ANOVA showed a significant effect of PHA concentration on the total activity of zebrafish larvae [F(7,184) = 2.9135, P = 0.0065]. The results indicated that in the standard open field test, there was no significant concentration-dependent effect of PHAs on the total locomotor activity level. In the colour-enriched open field test, total locomotor activity level declined significantly in a dose dependent manner compared with the un-injected group (0 μg mL−1). These results show that the level of locomotor activity was not concentration-dependent in the standard open field tests but decreased in a regular manner in the colour-enriched open field tests, suggesting that colour was a key factor in altering the concentration-dependent effect of PHAs on decreasing the total locomotor activity patterns of PHA-injected zebrafish larvae. The 15 min exploratory time was divided into three blocks (first 5 min, middle 5 min and last 5 min) to assess the temporal effect of PHAs on locomotor activity patterns of un-injected and PHA-injected zebrafish larvae in the standard vs. colour-enriched open field tests. An analysis of the temporal pattern of locomotor activity (irrespective of zones) showed that levels of locomotor activity of un-injected larvae and larvae injected with PHAs did not vary over time in the standard open field test (Fig. 6A). However, in the colour-enriched open field test (Fig. 6B), although total activity level decreased as mass of PHAs increased, activity level significantly increased across the three time (5 min) blocks. | ||
| Fig. 6 Effect of PHAs on the pattern of locomotor activity in PHAs-injected and un-injected zebrafish larvae. (A) Total activity level and temporal pattern (15 min divided into three 5 min blocks) of locomotor activity of un-injected larvae and larvae injected with PHAs (0.25–1500 μg mL−1) in the standard open field test. (B) Total activity level and temporal pattern (15 min divided into three 5 min blocks) of locomotor activity of un-injected larvae and larvae injected with PHAs (0.25–1500 μg mL−1) in the colour-enriched open field test. The error bars represent the standard error of the means (SEM). The statistical significances were set at P < 0.05 (*) and P < 0.01 (**). | ||
An analysis of the temporal pattern (three 5 min blocks) of locomotor activity (regardless of zones) showed that although levels of locomotor activity of both the control group and PHA injected groups did not vary over time in the standard open field test (Fig. 6A). While the locomotor activity levels of the control group increased gradually in the colour-enriched open field test (Fig. 6B). For the locomotor activity in the inner area, a two-way ANOVA for repeated measures (time blocks, PHA concentration as main factors) revealed no significant interaction between time blocks and PHA concentration [F(14,552) = 0.8480, P = 0.6165], but there was an effect of time [F(2,552) = 6.3929, P = 0.0018]. An analysis with Dunnett's post hoc test indicated that locomotor activity levels in the third time block (minutes 10–15) were significantly elevated compared with the first time block (minutes 0–5, P < 0.01) in the colour-enriched open field test (Fig. 6B). For the locomotor activity in the outer area, the two-way ANOVA for repeated measures (time blocks, PHAs concentration as main factors) demonstrated that there was an effect of PHA concentration [F(7,552) = 2.498, P = 0.0156] on the temporal locomotor activity levels in the colour-enriched open field test, but there was no effect of time [F(2,552) = 0.9893, P = 0.3725] and no interaction between concentration and time [F(14,552) = 1.3060, P = 0.1986]. Dunnett's post hoc analysis indicated that locomotor activity levels in the third time block (minutes 10–15) were significantly elevated compared with the first time block (minutes 0–5, P < 0.05) in the colour-enriched open field test (Fig. 6B).
PHAs enhanced the exploratory behaviour of zebrafish larvae by increasing thigmotaxis (central avoidance)
As shown in Fig. 7A and B, both the standard open field test and the colour-enriched open field test apparatus could be divided into an inner and outer zone. When the effect of PHAs on the exploratory behaviour of zebrafish larvae was evaluated, all six colours in the outer zone were considered as a single variable (outer environmental complexity). | ||
| Fig. 7 Pattern of outer zone preference by PHAs-injected and un-injected zebrafish larvae. (A and B) In addition to the six radial zones, we also included two other zones, designated as the inner and outer zones. In both the standard and colour-enriched open field apparatus, the inner zone consisted of the centre area (white area, 2 cm in diameter) and the outer zone consisted of the remaining area, including all the radial zones surrounding the centre zone (shaded area). (C and G) Pattern of latency prior to exploration in PHAs-injected and un-injected zebrafish larvae in standard (C) and (G) colour-enriched open field tests. (D and I) Freezing behaviour of PHA-injected and un-injected zebrafish larvae in standard (D) and (I) colour-enriched open field tests. (E and H) Pattern of distance moved in outer zone by PHA-injected and un-injected zebrafish larvae in standard (E) and (H) colour-enriched open field tests. (F and J) Pattern of time spent in outer zone by PHA-injected and un-injected zebrafish larvae in standard (F) and (J) colour-enriched open field tests. The error bar represents the standard error of the means (SEM). The statistical significances were set at P < 0.05 (*), P < 0.01 (**) and P < 0.001 (***). | ||
Freezing behaviour
The proportion of time spent immobile was used as an indicator of freezing behaviour. Zebrafish showed different patterns of freezing behaviour as PHA concentration increased in the open field tests (Fig. 7D). A one-way ANOVA revealed that there was no significant effect [F(7,184) = 1.5367, P = 0.1573] of PHA concentration on freezing behaviour in the standard open field test. However, there was a significant effect [F(7,184) = 2.2630, P = 0.0312] of PHAs on freezing behaviour in the colour-enriched open field tests. PHAs attenuated the freezing time duration (Fig. 7I).Latency prior to exploration
The effect of PHA concentration on the time required to leave the central zone was analysed using a one-way ANOVA, and the test revealed that larvae tested in the standard open field showed a constant latency [F(7,184) = 1.8429, P = 0.0815] (Fig. 7C), whereas in the colour-enriched open field test the latency period was significantly shorter [F(7,184) = 3.8641, P = 0.0006] (Fig. 7G) and increased as PHA concentration increased. The results suggest that PHAs decreased the anxiety of zebrafish larvae caused by environmental complexity (colour) in the colour-enriched open field test.PHAs decreased thigmotaxis (central avoidance) in the standard open field test but increased thigmotaxis (central avoidance) in the colour-enriched open field test
A one-way ANOVA demonstrated a concentration-dependent effect of PHAs on distance moved within the outer zone in the standard [F(7,184) = 2.4185, P = 0.0216] (Fig. 7E) and colour-enriched tests [F(7,184) = 3.6246, P = 0.0011] (Fig. 7K) and on time spent in the outer zone in the standard [F(7,184) = 2.2365, P = 0.0332] (Fig. 7F) and colour-enriched tests [F(7,184) = 2.6527, P = 0.0123] (Fig. 7J). An analysis of the outer percent TDM and the percentage of time spent in the outer zone revealed that PHAs decreased the thigmotaxis of zebrafish larvae in the standard open field test but increased thigmotaxis in the colour-enriched open field test.PHAs injection increased the preference for blue, red and black colours
 | ||
| Fig. 8 Effect of PHAs on colour zone preference in the colour-enriched open field test. (A) (a) The colour zone layout was randomized for each PHA concentration group to eliminate the effect of spatial preference by zebrafish. (b) Effect of PHA concentration on patterns of trajectories corresponding to the colour zone layout. (c) Effect of PHA concentration on the pattern of time spent in the colour-enriched zones. Numbers mark the corresponding colour in (a–c). (B) (a) Colour zone preference/avoidance as measured by the % TDM per zone. (b) Colour zone preference/avoidance as measured by the % of time spent per zone. (c) Colour zone preference/avoidance as measured by the number of entries per zone. The error bar represents the standard error of the means (SEM). The statistical significances were set at P < 0.05 (*), P < 0.01 (**) and P < 0.001 (***). | ||
Discussion
Effect of PHAs on zebrafish embryos and larval development
To assess the toxicity of different concentrations of PHA particles during zebrafish development, PHAs were microinjected into zebrafish embryos at the 1–4 cell stage, and the effects on the embryos were observed over the course of their development. For decades, a large number of in vivo studies have demonstrated that PHAs are biodegradable and biocompatible and represent promising new materials in the ever-evolving area of biomedical materials. In this study, biocompatibility and toxicity were assessed by observing the morphology of embryos that were injected with PHAs at different stages from 4 hpf to 96 hpf. In addition, the effect of PHAs on spatial and temporal patterns of embryonic gene expression were also evaluated using whole-mount in situ hybridization of several key genes including ntl, krox20 and cmlc2. As expected, no abnormalities in embryonic and larval morphology or in gene expression patterns were found. Only expression of krox20 fluctuated as the PHAs concentration changed. Thus, PHAs have little harmful effect on the early development of zebrafish embryos and larvae but may exert a potential effect on behaviours of larvae.Effect of PHAs on total and temporal locomotor activity of zebrafish larvae
PHAs had no effect on total locomotor activity in standard open field test but reduced the total activity in colour-enriched open field test. Detailed analysis of the temporal patterns of locomotor activity over the entire open field apparatus revealed that in some PHAs concentration groups, a significant increase in the level of locomotor activity was observed over time only in colour-enriched open field tests. The results revealed that at some concentrations, PHA injection can alter the total and temporal locomotor activity patterns of larval zebrafish at 6 dpf.Effect of PHAs on thigmotaxis in standard and colour-enriched open field tests
PHA-injected larval zebrafish showed a decreased preference for the outer zones in the standard open field test but an increased preference for the outer zones in the colour-enriched test. These results suggest that PHAs appear to modify the natural pattern of exploration between the inner and outer zones, in which control zebrafish larvae moved primarily in the centre of the colour-enriched open field as a result of the complex/stressful environment created by the presence of different colours.40 Colour presence and absence in the open field test caused injected zebrafish larvae to demonstrate distinct thigmotaxis (central avoidance) behaviour, indicating that PHAs enhanced the tendency to explore the outer areas even in the presence of environmental complexity.Effect of PHAs on the pattern of exploratory behaviour by zebrafish larvae in the coloured-enriched open field
Colour was a key factor influencing the effect of PHAs on exploratory behaviour. There was a distinct difference between the effect of PHAs on the exploratory behaviour in the standard and colour-enriched open field tests. In the standard open field test, exploratory parameters such as the latency to begin exploration, percent TDM in the outer zone and time spent in the outer zone were not correlated with an increase in PHA concentration. However, a different pattern of exploratory behaviour was observed in PHAs-injected larvae tested in the colour-enriched open field. First, we observed that compared with un-injected zebrafish larvae, as PHAs concentration increased, entrance into the outer zone was initiated within a shorter period. Furthermore, as PHA concentration increased, total activity levels in the colour-enriched test gradually increased across the three time blocks (0–5 min, 5–10 min, 10–15 min), and a significant burst of locomotor activity occurred in the third time block (minutes 10–15) compared with the other time blocks. These results suggest that PHAs may enhance the ability of larval zebrafish to process the complexity of the coloured environment.Effect of PHAs on colour discrimination and natural colour preference/avoidance
At the age of 6 days, zebrafish larvae have been shown to have the physiological and behavioural capacity for colour discrimination.40,46–48 Zebrafish larvae at this life stage are already capable of responding to the complex visual stimuli in an aroused state.49 Previous reports have demonstrated that the natural pattern of colour preference/avoidance among a choice of 6 colours consists of avoidance of yellow, red, black, and blue and a preference for green and orange.40 However, PHAs injection, particularly at 300 μg mL−1, reversed this preference/avoidance order with blue > red ≈ black > yellow > white (central) > green > orange. This interesting finding suggests that PHAs injection in the early embryonic stages of zebrafish had exploratory, visual and neural effects on zebrafish larvae, which reveals the potential for neuro-behavioural effects of PHAs.Conclusion
In sum, this study provides a detailed description of the potential effects of PHAs on the embryonic development and exploratory behaviour of zebrafish larvae. We found that PHAs injection are correlated to fluctuate expression of krox20, a transcription factor that is involved in vertebrate hindbrain segmentation, particularly in the formation and specification of rhombomeres (r) 3 and 5, as important marker of zebrafish brain. Furthermore, PHAs caused changes in locomotive activity, exploratory behaviour and colour preference of larvae tested in open field assay. Further investigation should be conducted to decipher the genetic and molecular mechanisms of the behavioural changes induced by PHAs.Materials and methods
Experimental animals
All of the experimental protocols and procedures involving zebrafish were approved by the Committee for Animal Experimentation of the College of Life Science at Nankai University (no. 2008) and were performed in accordance with the NIH Guide for the Care and Use of Laboratory Animals (no. 8023, revised in 1996). The animals were maintained in aquaria at 28.5 °C with a 10/14 h dark/light cycle. All larvae were rapidly euthanized with an overdose (0.168 g L−1) of tricaine (MS-222) and disposed of according to local regulations following completion of the behavioural testing.Preparation, characterization, and stability of PHAs in standard system water
The PHAs used in this experiment were purchased from Tianjin Green-Bio Materials Co., Ltd, Tianjin, China. To simulate the actual conditions of waste PHAs in an aquatic environment, PHAs of different concentrations (0.25 μg mL−1, 2.5 μg mL−1, 25 μg mL−1, 75 μg mL−1, 300 μg mL−1, 1000 μg mL−1, 1500 μg mL−1) were prepared by weighing and adding powdered PHAs directly to the water system. The diameter and distribution of the PHAs particles in the water system were then determined using dynamic light scattering (DLS) (ZetaPALS BI-200SM, Brookhaven Instruments, USA).Embryos and microinjection
Adult wild-type zebrafish were maintained at 28.5 °C on a 14 h light/10 h dark cycle in a standard water system (KCl 0.05 g L−1, NaHCO3 0.025 g L−1, NaCl 3.5 g L−1, and CaCl2 0.1 g L−1, pH 7.0–7.2). At the appropriate time for zebrafish mating, a ratio of one male to two females was established in one tank for maximum embryo production. The embryos were cultured at 28.5 °C in system water and staged as previously described.50 Embryos at the one- to four-cell stage were visually assessed and collected after natural spawning. Embryos at the one- to four-cell stage were then transferred to the injection channel of an agarose plate with system water. One of eight concentrations of PHAs (0 μg mL−1, 0.25 μg mL−1, 2.5 μg mL−1, 25 μg mL−1, 75 μg mL−1, 300 μg mL−1, 1000 μg mL−1, 1500 μg mL−1 in the standard system water) was injected into the yolk of each embryo (n > 50). Four nanoliters of PHA solution was injected into each embryo. Control group in the experiment stand for the “injected control” group.Morphology and mortality monitoring and analysis
Embryos were observed under a stereomicroscope (Olympus SZX 10, Japan) for morphological analysis and mortality estimation at different developmental stages ranging from 4 hours post-fertilization (hpf) to 4 days post-fertilization (dpf) and were photographed according to the method of Kimmel.51 Immediately following PHA microinjection into the yolk, embryos were observed and photographed; the proportion of embryos with epiboly was quantified at 9 hpf and 10 hpf. Head area, eye area and body length of embryos in the control (0 μg mL−1) and PHA-injected groups were measured using Olympus BP2-BSW image processing software (supporting the stereomicroscope Olympus SZX 10, Japan) at 8 hpf, 48 hpf, 72 hpf and 96 hpf. Embryos in the control group (0 μg mL−1) and the 25 μg mL−1, 300 μg mL−1 and 1000 μg mL−1 injected groups were collected at 10 hpf (for ntl), at 12 hpf (for krox20), and at 28 hpf (for cmlc2) for in situ hybridization.Whole-mount in situ hybridization
Embryos were collected at the appropriate stages and fixed overnight at 4 °C in 4% paraformaldehyde, pH 7.0, in phosphate-buffered saline. Plasmids containing ntl, krox20 and cmlc2 were generous gifts from Professor Hongwei Zhang (Shandong University). Digoxigenin (DIG)-labelled antisense RNA probes were transcribed with either SP6 or T7 RNA polymerases according to the manufacturer's instructions (Roche) using linearized plasmids as DNA templates. The embryos were dechorionated with forceps after being washed three times with PBS and stored in methanol at −20 °C. Whole-mount in situ hybridization was performed as previously described.52Experimental apparatus for open field tests
Open field tests consisted of a standard open field test and a colour-enriched open field test using an apparatus and experimental procedure previously described in detail.40 The colour-enriched open field test apparatus was composed of a standard glass dish (9 cm in diameter). As Fig. 1 shows, the outer zone of the dish was equally divided into six radial sectors that were enriched with red, blue, yellow, orange and black colours (the light transmittance peaks of these colours are provided in Fig. S6†). The central zone was 2 cm in diameter and lacked colour. To exclude possible effects of position or any visual clues on colour preference/avoidance, the order of the colour layout in the six sectors was randomly reset for each concentration group. The standard open field apparatus was identical to the colour-enriched open field test apparatus except that the standard apparatus lacked colour.Experimental procedure for open field tests
We performed the open field test in strict accordance with the previous work that created the experiment standard.40 Larvae (6 dpf) that were PHA-microinjected at the two-cell embryonic stage were immediately transferred to the central zone of the standard or colour-enriched open field apparatus with a plastic Pasteur pipette. The light source consisted of white and infrared luminous diodes arranged alternately in a circuit board as depicted in Fig. S4.† The video recording system and software were custom-made (for detailed information please see the supplementary methods in the ESI†). One complete experimental procedure comprised a paired standard open field test and a colour-enriched open field test conducted simultaneously with one larva in each test apparatus. Once the larvae were transferred into the test apparatus, the 15 min video recording of the behaviour began. Experiments were performed in a quiet and secluded room with the researcher out of view of the zebrafish larvae. For each of the 8 groups (concentrations), a total of 48 (n = 24 for standard open field test and n = 24 for the colour-enriched open field test) zebrafish larvae (6 dpf) were tested. Once the behavioural tests were completed, all of the larvae were rapidly euthanized with an overdose (0.168 g L−1) of tricaine (MS-222) and disposed of according to the guidelines of the Committee for Animal Experimentation of the College of Life Science at Nankai University (no. 2008).Behavioural parameters for open field tests
Locomotor activity in the different zones was measured as the total distance moved (TDM) in the central and outer zones (for both standard and colour-enriched tests) and in colour and non-colour zones (for colour-enriched tests). These parameters were indicators of (1) the exploratory behaviour of larvae tested with or without colour (high values indicated greater exploration) and (2) colour preference (high values in particular coloured areas indicated a preference for that colour).The temporal locomotor activity level was defined as the time at which the behavioural response of a tested subject (animal) became attenuated to a continuous stimulus/environment (colour, in this experiment).45,53 The temporal locomotor activity level was determined statistically by comparing the mean TDM during the last 5 min with the mean TDM during the first 5 min; when a significant difference was detected, the subject was determined to be adapted. This method was used to determine whether the two main factors in this experiment—the complexity of the environment (i.e., presence or absence of colour) and the concentration of injected PHAs—had differentially affected the pattern of the temporal locomotor activity level.
Anxiety induced by environmental complexity was measured as centre avoidance, which was calculated as the ratio between TDM in the inner zone and TDM over the entire test arena (both inner and outer zones). Thigmotaxis (central avoidance) can also be represented as the percentage of time spent in different zones.35 Freezing behaviour was also used as an indicator of anxiety and was measured as the amount of time spent (percentage) immobile (defined as the absence of movement for ≥1 s) for the total duration of the test.37,54,55
Colour zone preference/avoidance was measured by the percent TDM within each of the 6 colour-enriched zones and the percent TDM within the central zone. To remove any spatial influence on the colour choice of the larvae, we used the standard open field test and calculated the percent TDM for each of six (non-coloured) sectors; the results indicated no preference for any radial zone in the absence of colour, with an average percent TDM of 14.28% per sector. To obtain additional information on exploratory patterns and colour preference, the number of visits to every zone was also recorded. To eliminate the effect of a spatial preference on the colour choice of zebrafish larvae in the colour-enriched open field test, the position of the colours were randomly reset for every experiment.
Data analysis
The movement data were processed using Visual Studio 2005 (Microsoft) and Matlab R2011b (Math Works). The statistical analysis was conducted in Matlab R2011b (Math Works). All data were arcsine square root transformed prior to analysis to stabilize the variance prior to running ANOVAs. A one-way ANOVA was performed to analyse the effect of PHA concentration on total activity level in the open field environments, thigmotaxis (central avoidance), latency to begin exploration, freezing behaviour and colour preference/avoidance (percent TDM per zone, percent time spent per zone and number of entries per zone). A two-way ANOVA for repeated measures (time blocks and PHA concentration as main factors) was conducted to assess concentration and time effects on total locomotor activity levels in standard and colour-enriched open field tests. Dunnett's post hoc test was used to analyse multiple comparisons. Data are presented as the mean ± SEM, and a probability level of 5% was used as the minimum criterion for significance.Author contributions
X. Z. F., Y. L. and X. L. conceived and designed the experiments. X. L. conducted the zebrafish behavioural assays and wrote the manuscript. X. L. wrote the manuscript. The video tracking software was designed by D. C., M. Z. S. and X. Z. Authors X. L. and J. L. G. collected and analysed the behavioural data. X. L., D. F. F. and D. Y. C. observed the embryonic development and assessed gene expression. All authors discussed the results and implications and reviewed the manuscript at all stages.Competing financial interests
The authors declare no competing financial interests.Acknowledgements
We thank the Special Fund for Basic Research on Scientific Instruments from the Chinese National Natural Science Foundation (grant no. 61327802) and the National Basic Research Program of China (2015CB856500) for support of this work. D. F. F. thank the support from the National Natural Science Foundation of China (grant no. 81501589). D. Y. C. also acknowledges support from the Tianjin Science Technology Research Funds of China (grant no. 14JCQNJC09600). Y. L. acknowledges support from the Tianjin Science Technology Research Funds of China (grant no. F1012921 and F1014931). X. Z. F. acknowledges support from the State Key Laboratory of Medicinal Chemical Biology, Nankai University, Tianjin, China (grant no. 201501019).References
- A. J. Anderson and E. A. Dawes, Occurrence, metabolism, metabolic role, and industrial uses of bacterial polyhydroxyalkanoates, Microbiol. Rev., 1990, 54, 450–472 CAS.
- A. Steinbüchel, Perspectives for biotechnological production and utilization of biopolymers: metabolic engineering of polyhydroxyalkanoate biosynthesis pathways as a successful example, Macromol. Biosci., 2001, 1, 1–24 CrossRef.
- K. Sudesh, H. Abe and Y. Doi, Synthesis, structure and properties of polyhydroxyalkanoates: biological polyesters, Prog. Polym. Sci., 2000, 25, 1503–1555 CrossRef CAS.
- W. Amass, A. Amass and B. Tighe, A review of biodegradable polymers: uses, current developments in the synthesis and characterization of biodegradable polyesters, blends of biodegradable polymers and recent advances in biodegradation studies, Polym. Int., 1998, 47, 89–144 CrossRef CAS.
- S. D. P. Williams, D. M. Horowitz and O. P. Peoples, PHA applications: addressing the price performance issue I. Tissue Engineering, Int. J. Biol. Macromol., 1999, 25, 111–121 CrossRef CAS PubMed.
- U. A. Stock and J. E. Mayer Jr, et al., Tissue engineering of cardiac valves on the basis of PGA/PLA Co-polymers, J. Long-Term Eff. Med. Implants, 2001, 11, 249–260 CrossRef CAS PubMed.
- E. Shishatskaya and T. Volova, A comparative investigation of biodegradable polyhydroxyalkanoate films as matrices for in vitro cell cultures, J. Mater. Sci.: Mater. Med., 2004, 15, 915–923 CrossRef CAS PubMed.
- E. I. Shishatskaya, O. N. Voinova, A. V. Goreva, O. A. Mogilnaya and T. G. Volova, Biocompatibility of polyhydroxybutyrate microspheres: in vitro and in vivo evaluation, J. Mater. Sci.: Mater. Med., 2008, 19, 2493–2502 CrossRef CAS PubMed.
- E. I. Shishatskaya. in Macromolecular symposia, Wiley Online Library, 2008, pp. 65–81 Search PubMed.
- E. Shishatskaya, T. Volova and I. Gitelson, in Doklady Biological Sciences, Springer, 2002, pp. 109–111 Search PubMed.
- X.-Y. Xu, et al., The behaviour of neural stem cells on polyhydroxyalkanoate nanofiber scaffolds, Biomaterials, 2010, 31, 3967–3975 CrossRef CAS PubMed.
- E. Shishatskaya, T. Volova, A. Puzyr, O. Mogilnaya and S. Efremov, Tissue response to the implantation of biodegradable polyhydroxyalkanoate sutures, J. Mater. Sci.: Mater. Med., 2004, 15, 719–728 CrossRef CAS PubMed.
- S. E. Lepage and A. Bruce, Zebrafish epiboly: mechanics and mechanisms, Int. J. Dev. Biol., 2010, 54, 1213–1228 CrossRef CAS PubMed.
- M. Tada and M. Kai, Noncanonical Wnt/PCP signaling during vertebrate gastrulation, Zebrafish, 2009, 6, 29–40 CrossRef CAS PubMed.
- C. J. Weijer, Collective cell migration in development, J. Cell Sci., 2009, 122, 3215–3223 CrossRef CAS PubMed.
- V. E. Fako and D. Y. Furgeson, Zebrafish as a correlative and predictive model for assessing biomaterial nanotoxicity, Adv. Drug Delivery Rev., 2009, 61, 478–486 CrossRef CAS PubMed.
- A. J. Hill, H. Teraoka, W. Heideman and R. E. Peterson, Zebrafish as a model vertebrate for investigating chemical toxicity, Toxicol. Sci., 2005, 86, 6–19 CrossRef CAS PubMed.
- Y. Wang, J. L. Seebald, D. P. Szeto and J. Irudayaraj, Biocompatibility and biodistribution of surface-enhanced Raman scattering nanoprobes in zebrafish embryos: in vivo and multiplex imaging, ACS Nano, 2010, 4, 4039–4053 CrossRef CAS PubMed.
- Y. Liu, et al., A progressive approach on zebrafish toward sensitive evaluation of nanoparticles' toxicity, Integr. Biol., 2012, 4, 285–291 RSC.
- X. Li, et al., SiO2 nanoparticles change colour preference and cause Parkinson's-like behaviour in zebrafish, Sci. Rep., 2014, 4, 3810, DOI:10.1038/srep03810.
- N. He, et al., Exploring the toxicity of a bismuth–asparagine coordination polymer on the early development of zebrafish embryos, Chem. Res. Toxicol., 2013, 26, 89–95 CrossRef CAS PubMed.
- J. Y. Xue, et al., An assessment of the impact of SiO2 nanoparticles of different sizes on the rest/wake behavior and the developmental profile of zebrafish larvae, Small, 2013, 9, 3161–3168 CrossRef CAS PubMed.
- Y.-P. Wang, et al., Developmental and cartilaginous effects of protein-coated SiO2 nanoparticle corona complexes on zebrafish larvae, RSC Adv., 2014, 4, 18541–18548 RSC.
- M. Sison, J. Cawker, C. Buske and R. Gerlai, Fishing for genes influencing vertebrate behavior: zebrafish making headway, Lab. Anim., 2006, 35, 33–39 CrossRef PubMed.
- R. Sood, et al., Methods for reverse genetic screening in zebrafish by resequencing and TILLING, Methods, 2006, 39, 220–227 CrossRef CAS PubMed.
- D. Chatterjee and R. Gerlai, High precision liquid chromatography analysis of dopaminergic and serotoninergic responses to acute alcohol exposure in zebrafish, Behav. Brain Res., 2009, 200, 208–213 CrossRef CAS PubMed.
- R. Gerlai, High-throughput behavioral screens: the first step towards finding genes involved in vertebrate brain function using zebrafish, Molecules, 2010, 15, 2609–2622 CrossRef CAS PubMed.
- H. Anisman and K. Matheson, Stress, depression, and anhedonia: caveats concerning animal models, Neurosci. Biobehav. Rev., 2005, 29, 525–546 CrossRef PubMed.
- C. Belzung and G. Griebel, Measuring normal and pathological anxiety-like behaviour in mice: a review, Behav. Brain Res., 2001, 125, 141–149 CrossRef CAS PubMed.
- T. Darland and J. E. Dowling, Behavioral screening for cocaine sensitivity in mutagenized zebrafish, Proc. Natl. Acad. Sci. U. S. A., 2001, 98, 11691–11696 CrossRef CAS PubMed.
- S. Guo, Linking genes to brain, behavior and neurological diseases: what can we learn from zebrafish?, Genes, Brain Behav., 2004, 3, 63–74 CrossRef CAS.
- S. Baraban, M. Taylor, P. Castro and H. Baier, Pentylenetetrazole induced changes in zebrafish behavior, neural activity and c-fos expression, Neuroscience, 2005, 131, 759–768 CrossRef CAS PubMed.
- J. Ninkovic and L. Bally-Cuif, The zebrafish as a model system for assessing the reinforcing properties of drugs of abuse, Methods, 2006, 39, 262–274 CrossRef CAS PubMed.
- A. Stewart et al., in Zebrafish Neurobehavioral Protocols, Springer, 2011, pp. 169–179 Search PubMed.
- J. A. Bouwknecht and R. Paylor, Pitfalls in the interpretation of genetic and pharmacological effects on anxiety-like behaviour in rodents, Behav. Pharmacol., 2008, 19, 385–402 CrossRef PubMed.
- S. D. Suarez and G. G. Gallup, An ethological analysis of open-field behavior in rats and mice, Learn. Motiv., 1981, 12, 342–363 CrossRef.
- L. Prut and C. Belzung, The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review, Eur. J. Pharmacol., 2003, 463, 3–33 CrossRef CAS PubMed.
- D. C. Rogers, et al., Behavioral and functional analysis of mouse phenotype: SHIRPA, a proposed protocol for comprehensive phenotype assessment, Mamm. Genome, 1997, 8, 711–713 CrossRef CAS PubMed.
- D. J. Echevarria, C. M. Hammack, D. W. Pratt and J. D. Hosemann, A novel behavioral test battery to assess global drug effects using the zebrafish, Int. J. Comp. Psychol., 2008, 21, 19–34 Search PubMed.
- F. Ahmad and M. K. Richardson, Exploratory behaviour in the open field test adapted for larval zebrafish: impact of environmental complexity, Behav. Processes, 2013, 92, 88–98 CrossRef PubMed.
- R. F. Thompson and W. A. Spencer, Habituation: a model phenomenon for the study of neuronal substrates of behavior, Psychol Rev., 1966, 73, 16 CrossRef CAS PubMed.
- J. Best and W. K. Alderton, Zebrafish: an in vivo model for the study of neurological diseases, Neuropsychiatr. Dis. Treat., 2008, 4, 567 CrossRef CAS PubMed.
- A. Cerbone and A. G. Sadile, Behavioral habituation to spatial novelty: interference and noninterference studies, Neurosci. Biobehav. Rev., 1995, 18, 497–518 CrossRef.
- C. Tomaz, M. S. Aguiar and P. J. Nogueira, Facilitation of memory by peripheral administration of substance P and naloxone using avoidance and habituation learning tasks, Neurosci. Biobehav. Rev., 1991, 14, 447–453 CrossRef.
- J. D. Best, et al., Non-associative learning in larval zebrafish, Neuropsychopharmacology, 2008, 33, 1206–1215 CrossRef CAS PubMed.
- S. E. Brockerhoff, J. E. Dowling and J. B. Hurley, Zebrafish retinal mutants, Vision Res., 1998, 38, 1335–1339 CrossRef CAS PubMed.
- S. S. Easter Jr and G. N. Nicola, The development of vision in the zebrafish (Danio rerio), Dev. Biol., 1996, 180, 646–663 CrossRef CAS.
- V. C. Fleisch and S. C. Neuhauss, Visual behavior in zebrafish, Zebrafish, 2006, 3, 191–201 CrossRef PubMed.
- T. Yokogawa, M. C. Hannan and H. A. Burgess, The dorsal raphe modulates sensory responsiveness during arousal in zebrafish, J. Neurosci., 2012, 32, 15205–15215 CrossRef CAS PubMed.
- M. Westerfield, The zebrafish book: a guide for the laboratory use of zebrafish (Danio rerio), University of Oregon Press, 2000 Search PubMed.
- C. B. Kimmel, W. W. Ballard, S. R. Kimmel, B. Ullmann and T. F. Schilling, Stages of embryonic development of the zebrafish, Dev. Dyn., 1995, 203, 253–310 CrossRef CAS PubMed.
- J. Shao, et al., Tissue regeneration after injury in adult zebrafish: the regenerative potential of the caudal fin, Dev. Dyn., 2011, 240, 1271–1277 CrossRef CAS PubMed.
- L. M. Gómez-Laplaza and R. Gerlai, Latent learning in zebrafish (Danio rerio), Behav. Brain Res., 2010, 208, 509–515 CrossRef PubMed.
- C. Maximino, et al., Measuring anxiety in zebrafish: a critical review, Behav. Brain Res., 2010, 214, 157–171 CrossRef PubMed.
- N. Sousa, O. Almeida and C. Wotjak, A hitchhiker's guide to behavioral analysis in laboratory rodents, Genes, Brain Behav., 2006, 5, 5–24 CrossRef PubMed.
Footnotes |
| † Electronic supplementary information (ESI) available. See DOI: 10.1039/c6ra04329a |
| ‡ These authors contributed equally to this work. |
| This journal is © The Royal Society of Chemistry 2016 |