
Enhanced long-term nitrogen removal by organotrophic anammox bacteria under different C/N ratio constraints: quantitative molecular mechanism and microbial community dynamics†
Abstract
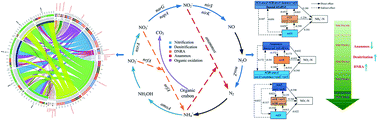
The anaerobic ammonium oxidation (anammox) process has mainly been applied to NH4+–N-rich wastewater with very low levels of organic carbon (<0.5 g COD per g N). Some anammox bacteria species have the capacity to oxidize organic carbon with nitrate as the electron acceptor. However, little is known about the organotrophic anammox nature of “Ca. Brocadia sinica”. To elucidate the metabolic versatility and microbial succession of “Ca. Brocadia sinica” under TOC/TN stress conditions, the influence of TOC/TN ratios on the nitrogen transformation pathway and the dynamics of microbial communities were investigated. It was found that an appropriate TOC/TN ratio (<0.2) could promote the anammox activity over the short-term. Meanwhile, “Ca. Brocadia sinica” had higher tolerance to higher TOC/TN (>0.4) abiotic stresses. Mass balance indicated that organotrophic anammox could outcompete denitrifiers under a TOC/TN ratio of 0.1–0.2. Quantitative response relationships and pathway analysis revealed that (AOA amoA + AOB amoA + anammox + nrfA)/bacteria, nrfA/(narG + napA), and nrfA were the key functional gene groups determining the organotrophic anammox contribution. Additionally, MiSeq sequencing showed that Planctomycetes, Proteobacteria, Chloroflexi, and Chlorobi were the most abundant phyla in the organotrophic anammox system. Furthermore, higher TOC/TN ratios (>0.40) could result in the community succession of anammox species, in which “Ca. Jettenia caeni” and “Ca. Kuenenia stuttgartiensis” were the dominant organotrophic anammox bacteria species. Overall, combined analyses revealed that the coupling of anammox, DNRA (organotrophic anammox), and denitrification comprised the primary pathway that accounted for TOC and nitrogen removal.
 Please wait while we load your content...
Please wait while we load your content...

